|
RESPUESTAS A RIESGO DE DEPREDACIÓN Y OPORTUNIDADES
ALIMENTICIAS EN PECES PELÁGICOS FORMADORES DE
CARDÚMENES
Trabajo recibido el 1º de Julio de 1993 y aceptado
para su publicación el 30 de Marzo de 1994.
Carlos J. Robinson
Instituto de Ciencias del Mar y Limnología,
Universidad Nacional Autónoma de México, Apartado Postal 70-305.
México, D.F. 04510, MÉXICO. Contribución No. 743 del
Instituto de Ciencias del Mar y Limnología.
Virgilio Arenas-F.
Instituto de Ciencias del Mar y Limnología,
Universidad Nacional Autónoma de México, Apartado Postal 70-305.
México, D.F. 04510, MÉXICO. Contribución No. 743 del
Instituto de Ciencias del Mar y Limnología.
El conocimiento
que se tiene sobre el compromiso presentado por peces gregarios que
viven en ambientes de riesgo de depredación es limitado. Experimentos
y observaciones previos han subestimado el hecho de que las especies
de peces gregarios pueden tomar ventaja de vivir en grupo cuando se
presenta un riesgo de ser depredados y la necesidad de obtener alimento.
En este estudio se analizan las respuestas dadas por el sprat (Sprattus
sprattius). Los resultados indican que cuando se busca alimento
en un ambiente de riesgo, los individuos dentro del cardumen son capaces
de explotar los beneficios anti-depredador que les confiere el vivir
en grupo. Sin embargo, cuando el hambre aumenta, estos peces gregarios
comprometen el beneficio de asociarse para obtener alimento. Por medio
de la manipulación del estado de hambre de algunos de los individuos
con respecto al resto, se concluye que los menos hambrientos de un cardumen
obtienen mayores ventajas. Sin embargo, cuando un pez se encuentra más
hambriento que el resto se enfrenta a la situación más
comprometedora de todas las analizadas. PALABRAS-CLAVE:
Cardumen, peces pelágicos, depredación. Current knowIedge
of behavioural compromises made by shoaling fish foraging in risky environments
is limited. Some previous experiments haye overlooked the fact that
shoaling fish species may take ad.vantage of living in groups to trade-off
risk of predation and food rewards. In the present study the compromises
presented by sprat (Sprattus sprattus) are analysed. Results
indicate that when searching for food in risky environments individuals
in a shoal are capable of exploiting the anti-predator benefits providing
of grouping. However, when bunger arises, shoaling fish may compromise
the benefits of shoaling to obtain food. By manipulating the bunger
state of the individuals with respect to the rest of the shoal, results
show that being less hungry in a shoal of hungry fish provides some
advantages to the fish. However, being more hungry in a less hungry
shoal put the fish in the most disadvantageous situation of those analysed.
KEY WORDS:
Schooling, shoaling, risk of predation, trade-off, pelagic fish.
The presence of a predator may influence the hehaviour of animals in diverse manners, from an increase of vigilance or escape response, to changes in habitat (Magurran and Pitcher, 1987; Metcalfe et al., 1987a; Werner et al., 1983). When antipredator behaviour is in conflict with other activities such as feeding, animals have been shown to modify their foraging tactics to reduce their risk of predation. The nature of the compromise may be modulated by motivational aspects such as the forager's hunger state or the temporal variation in predation risk (Milinski, 1985; Metcalfé et al., 1987a). Furthermore, the strategy employed may be related to the habitat and way of life of the species (Lima et al., 1985; Metcalféet al.,1987b; Huntingfórd et al., 1988). ANTI-PREDATOR ADVANTAGES OF SHOALINGLiving in grotips provides some benefits to the members of the group which are not shared by sofitary individuals (Bertram, 1978). Many of these benefits have been associated with predator avoidance. In numerous fish species for instance, shoaling is thought to have evolved as an anti-predator strategy (Seghers, 1974). In a group, the probability that an individual will be captured in a single attack decreases as group size increases (Hamilton, 1971). Dilution of individual risk on any given predator's attack has been supported by results of several authors (Neill and Cullen, 1974. Major, 1978; Morgan and Godin, 1985). The "dilution effecC theory is based on the premise that the probability of a predator catching a prey in a group is approximately 1/N where N = the number of fish in the shoal. Meanwhile, when a forager is searching for food, the chances of obtaining a particle of food increase with the number of visits to the area where food is supposed to be found, then, the probability of gathering food shotild be proportional to the frequency (f) of visits by the fish to the profitable area. Following these arguments, one may assume that for an individual in the shoal the safest and more profitable way to visit the area shotild be minimizing the ratio (1/N)/f. That is, incrementing its presence in a risky but profitable zone, but minimising its own "domain of dange? (Hamilton, 1971). However, if hunger increases it is expected that a hungry fish should compromise safety in group for food reward. To test this, we manipulated both the hunger state of an entire shoal and the hunger state of a single individual with respect to the rest of the shoal. MATERIAL AND METHODS CONDITION
CONDITION 1. ALL FISH FED AND ALL FISH HUNGRYFourteen sprat (Sprattus sprattus) (mean total length 6.5 cm, s.d. 0.22 cm) a pelagic fish abundant around the British Isles, were maintained in a circular tank (2.8m diam., 1.5 m high, 1.0 m deep water) divided in halves by a black plastic sheet across its diameter. Fish were able to swini into each side of the tank through a rectangular (100 x 60 cm) window located in the centre of the plastic 30 cm above the tank floor. A ration of 5.8 g day-¹ of mysid shrimp was delivered by a remotely-operated tube during seven days only to one side of the tank. On the seventh day, a life-size model of a sea gull was presented at 10.00 hours in the area were food was delivered. The model was released three consecutive times from a height of 1.5 m and allowed to dive into the water. At 14.00 hours of the same day, a fish swimining in the safé area i. e. the area of the tank where the model was not presented, was randonily chosen and observed. We will call this fish "target f¡sh". Its movements were followed using a video-camera equipped with a wide angle fl/4/ 12.5 mm lens and mounted beside the tank. The following events were recorded: a) Number of fish in the risky area: recorded as the number of fish in the risky area when the target fish was there. b) Elapsed time: defíned as the time that the target fish spent in the risky area. Once the target fish returned to the safe area another fish was chosen and then followed. This procedure was repeated with another fish, and so on during three hours. Following this, the shoal was removed from the tank and a new set of 14 sprats (mean body length= 6.2 cm, s.d. 0.45 cm) maintained in a spare tank was introduced. The shoal was undisturbed during two weeks with a ration of 5.8 g day-¹ of food. At the beginning of the next week diet was reduced to 0.8 g day-1. At 10.00 hours of the fourth day with this diet, the seagull model was again presented in the area where the food was delivered. Starting again at 14.00 hours and during three hours the same procedure of following a target fish was applied as with the previous shoal. CONDITION 2. A FISH WITHIN THE SHOAL DIFFERENUY HUNGRY MOTIVATED.A third set of 14 sprats (mean body length= 6.8 cm. sd. 0.31 cm) was introduced to the experimental tank Fish were mantained with a daily diet of 5.8 g of frozen shrimp for two -YN,eeks. After this period, the shoal was herded to one side of the tank using a large black plastic sheet and then a fish was captured with a dip net, anaesthefised with MS 222 (tricaine methane sulphonate) (1:15,000) and marked with a spot of standard methylene blue on both fianks of the fish's body using an inoculátor. With these marks, the fish was recognisable from the rest of the shoal. The following seven days the shoal was fed the regular 5.8 g diet. At the end of this regime the shoal was carefully herded to one side of the tanta, and the window in míddle of the curtain was blocked. However, the marked fish was left in the side where food used to be delivered. In order to avoid further stress to the marked fish, a group of Vive sprat was transferred from a spare tank to the side were the single fish esas maintained (fish in the spare tank were mantained with a similar regime of about 10% of fish body weight). The following day food was delivered to the side where the marked fish and the spare ones were swimming. In order to assure that the marked fish fed, a ration of 1.5 9 of food was detivered every 30 min destino 5 hours starting at 14.00 hours. Although the precise amount of food consumed by the fish was not known, an estimation was made by siphoning the remainder of the food at the end of cach section. From the total supplied, about a third was consumed cach day. The following day the same procedure of food administration was performed. However, just after the last ration, the spare fish were carefully removed from the tank and the window was again reopened allowing the marked fed fish to re-join the rest of the experimental shoal. The following day fish were swimming freely from one side of the tanta to the other and at 10.00 hours the sea gull model was again presented. At 14.00 hours and during the next three hours, the pattem of swimming of the less-hungry fish was followed. Register of counting and elapsed time was recorded as before. A new set of 14 sprats Amaran body length= 7.1 cm, sd. 0.44 cm) was introduced to the experimental tank and maintained for two weeks with the diet of 5.8 g of food. Afterwards, the shoal was herded to one side of the tank, a fish was captured with a dip net, anaesthetised and marked as before. During the following week, fish were lea undisturbed with the regime of 5.8 g of food and at the end of the 7th day, before tuming the light off the marked fish was isolated in the non- food arca of the tank, whereas the rest of the group was carefully herded to the other side. The window was clases and five fish from the spare tank were transferred joining the isolated fish. As previously, the next two days we started delivering portions of food every 30 minutes to one side of the tank during 5 hours starting at 14.00 hours. This time each portion was of 3.0 a. Fish did not show signs of lack of appetite. At the end of each day the remains were siphoned from the bottom of the tank. Of 30g delivered fish had consumed approximately 8.7 g during the first day and 10.3 during the second. No food was provided to the si where the second marked fish and the spare fish were swimming. Just before the ti went off, spare fish were removed from the tank and the window opened. The next day fish were swimming in a single shoal and at 10.00 h the seagull model was presented as More Finally, at 14.00 hours the more-hungry marked fish was chosen as a target fish and during three hours u behaviour was registered as with the previous fishes. During these observations the mean temperature of the water in the tank was 15.2ºc (14.6-15.9, 95% c.i.). RESULTS
CONDITION 1 The number of fish swimming in the risky arca during the threc-hours recordíng session varied between diets. When all-hungry, fish visited the risky arca more often and spent more time than when allfed (Table 1). A frecuencia analysis of number of fish in the arca is presented in Fig. 1. Results show that well-fed fish were present in groups of larger size than when fish were all hungry. A Kolmogorov Smirnov two-sample test (Siegel and Castellan, 1988) was performed to test the significance of the differences of shoal sizes between diets for each event (results of this analysis are presented in Fig. 1). 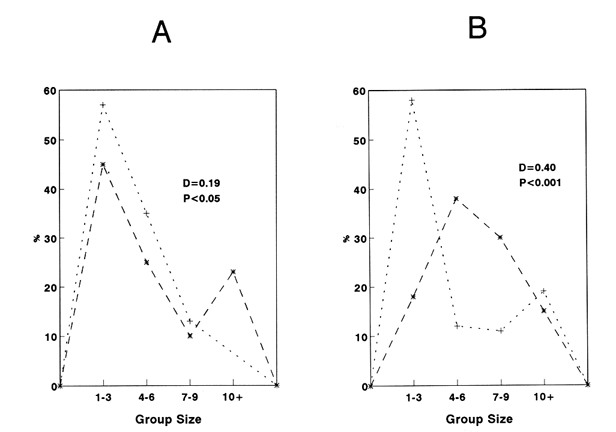 Figure 1. Distribution of relative frequencies of shoal size from a group of 14 sprat in the risky-area. Results are presented when all fish were fed and all hungry (A), and when a fish was fed, the rest hungry and when a fish was hungry and the rest was fed (B). Results of a Kolmogorov-Smirnov test, two-tailed, are presented to compare between conditions (* fed, + hungry). 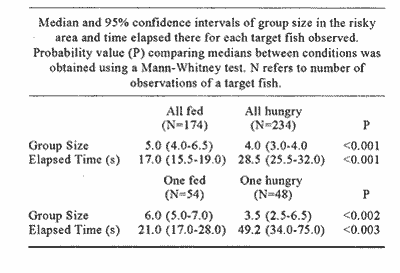 Table 1 Median and 95% confortense intervals of group size in the risky area and time elapsed there for each target fish observad. Probability valor (P) comparing medians between conditions was obtained using a Mann-Whitney test. N refers to number of observations of a target fish. Results of relation (I/N)/f for each diet are presented in Table 2. A KoImogorov-Smirnov two-sample test ¡radicales that when fed, fish tend to have smaller values of Index of Risk in their visits to the risky area than when they were hungry (D = 0.24, P<0.001, two-tailed) (Fig. 2). CONDITION 2In this case the nombre of occasions the target fish, either hungry or no hungry, were in the risky area was very similar (48 and 54 respectively). However, hungry fish tended to spend a significantly longer time per visit (Table 1). As More a further frequency analysis of nombre of fish in the risky area was performed (Fig. 1). Results show that when the target fish was less hungry than the others the fish was found in group of larger size. The results of a KoImogorov-Smirnov two sample test for each event are given in the same figure. The relation (1/N)/f was obtained for eachobservation, the results are presented in Table 3. When the target fish was the fed, it tended to present smaller values of index of rísk significantly lower than when the target fish was the hungry one (KoImogorov-Smirnov, two sample test, D = 0.62, P<0.00 1, two- callad) (Fig. 2). 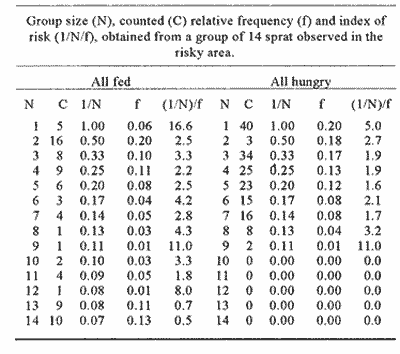 Table 2 Group size (N), counted (C) relative frecuencia (f) and index of risk (I/N/f), obtained from a group of 14 sprat observed in the risky area. 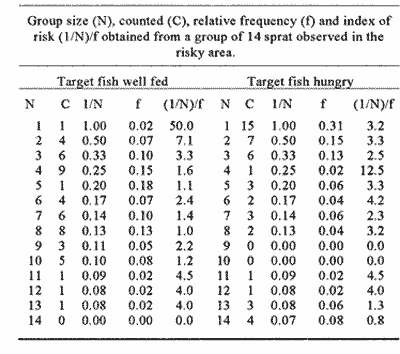 Figure 2. Cumulative distribiation of relative frequeney of visits for various values of index of risk obtained from a shoal of 14 sprat under four experimental conditions 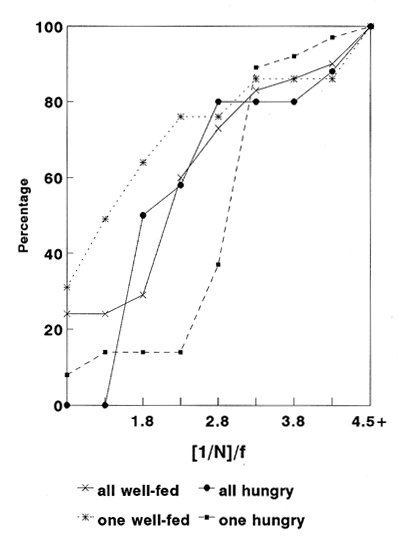 Table 3 Group size (N), counted (C), relative frequency (f) and index of risk (l/N)/f obtained from a group of 14 sprat observed in the risky area. DISCUSSIONPrevious works (e.g. Fraser and Huntingfórd, 1986; Pitcher et al., 1988) have analysed the strategies presented by shoaling fish when the demands for food and the avoidance of predators are in conflict. However, ¡n such works the main advantage associated to fish shoaling, security against predators (Seghers, 1974), due in part to the confusion and avoidance effect (Morgan and Godin, 1985; Landeau and Terborgh, 1986) was not taken in account. If it is accepted that shoaling provides antipredator advantages to an individual in a group, then risk incurred by a shoaling fish shotild not depend only on how y times or for how long a fish is present in a dangerous area but, essentially with how many companions the fish swims with. Results in the present work indicate that fish in a shoal may take advantage of the number of fish in a group visiting a risky area and control its own frequency of appearance. The ability to achieve this be nefit seems closely related to the hunger state of the individuals (specifically, to the relation of a partícular fish with the hunger condition of other members of the group). For instance, being less hungry in a shoal of hungry fish provides more advantages to the odd individual. It can reduce its visits to the unsafe area and be present in a way that allows it to minimise its own risk. However, being more hungry in a less hungry shoal puts a fish in the most disadvantageous situation of those analysed. In this case, the odd fish has to compromise food rewards and risk of predation more often, each. time longer and with a high risk. Results from other studies have demostrated that fish in a shoal take account of what others are doing (Pitcher et al., 1982). Similarly, during the development of the present experiment when the target fish was the less hungry fish, it was not difficult for the observer to predict when the red fish would cross to the risky area. As the fish were swiniming around in the sale area, the well-fed fish seemed to recognise how many fish were in a position to cross and how many fish were already in the other area. On several occasions, if there was a group of fish ready to cross e.g. swimming very close to the window, the target fish fóllowed and crossed with them. lf the group was too small, it remained in the safe area until another opportunity to cross arose. Therefore, it seems that fed fish took advantage of the behaviour of the rest of the hungry fish, allowing it to decide when and how to visit the unsafe area in a way which could minimise its own risk. The question is why, when hungry, fish did not visit the risky arca in a large group to reduce risk of predation? Results of other works have indicated that hungry fish within a shoal tend to be more independent and individualistíc (Morgan, 1988; Robinson and Pitcher, 1989). Individuals in a shoal may gain advantage of the benefits of grouping only when coordinases manoeuvres are displayed (Neill and Cullen, 1974- Pitcher and Wyche, 1983). It seems that in this experiment it did not pay for a hungry sprat to wait for other members of the shoal to cross to the risky area in search for food. Instead they crossed in small granos or individually, though by so doing they were incrementing their probability of being captured (Morgan and Godin, 1985). AgradecimientosACKNOWLEDGEMENTSThis work forms part of the requirements for CRM's doctoral degree from the University of Wales, Bangor, U.K. and was supported by UNAM and CONACyT, Mexico. We thank Dr. T.J. Pitcher for carly cristicisms to the manuscript. LITERATURABERTRAM, B.C.R. In:. Behavioral Ecology: An evolutionary approach. John Wiley. pp Living in groups:predator and prey J.R. Krebs and N.B. Davies New York 1978 213-234 FRASER, D.F. and F.A. HUNTINGFORD Ethology Feedíng and avoiding predation hazard: the behavioral response of the prey 1986 56-68 73 HAMILTON, W.D Sobornal of Theoretical Biology, Geometry for selfish haré. 1971 295-311 31 HUNTINGFORD. F.A., N.B., METCALFE and J.E., THORPE Journal of Fish Ecology Choice of feeding sintiera in Atlantic salmon, Salino salar, pare: effects of predatíon risk-, season and life history strategy. 1988 917-924 33 LANDEAU, L. and J., TERBORGH Animal Behaviour, Oddity and the "confusion Effect'' in predation 1986 1372-1380 34 LIMA, Se., TI., VALONE and T. CAPACES Animal Behaviour Foraging-efficiency-predation-risk tradicional in the grey squirrel. 1985 155-165 33 MAGURRAN, AY and T.J., PITCHER Proceeding of the Royal Society Provenance, shoal size and the sociobiology of predator-evasion behaviour in minnow shoals. London, B 1987 439-465 229 MAJOR, P. F., Animal Behaviour Predator-prey interactions in two schooling fishes Caranx ignobilis and Stolephorus purpureus. 1978 760-777 26 METCALFE, N.B., F.A. HUNTINGFORD and J.E. THORPE Animal Behaviour The influence of predation risk on the feeding motivation and foraging strategy ofjuvenile Atíantic sacaron. 1987a 901-911 35 METCALFE. N.B, EA HUNTINGFORD and U THORPE, Animal Behaviorir Predation risk inipairs diet selection in juvenile salmon. 1987b 931-933 35 MILINSKI, M. Behaviour Risk of predation of parasitized stickiebacks Gasteroster4s acideattis L. under composition for food. 1985 203-216 93 MORGAN, M.J. Journal of Fish Biology The effect of hunger, shoal size and the presence of a predator on shoal cohesiveness in bluntnose minnows, Pitnephales nosotras Rafinesque. 1988 963-971 32 MORGAN, M.J. and GODIN Zeitschrift fur Tierpsychologie Antipredator benefits of schooling behaviour in a Cyprinodontid fish, the banded killifish (Fundulus diaphanus) 1985 236-246 70 NEILL, SR. and J.M. CULLEN Journal of Zoology Experiments on whether schooling by their prey affects the huntíng behaviour of cep- and fish predators. London 1974 549-569 172 PITCHER, T.J, A.E. MAGURRAN and I.J WINFIELD, Behavioral Ecology and Sociobiology Fish in larger shoals find food faster. 1982 149-151 10 PITCHER, T.J., S.W. LANG and J.A., TURNER Behavioral Ecology and Sociobiology A risk-balancing trade off between Corajina rewards and predation hazard in a 1 shoaling fish. 1988 225-228 22 PITCHER, T.J., and CJ.. WYCHE In: Predator andprey in fishes The Hage: Dr. Junk. Predator-avo ¡dance behaviours of sand-eel schools: why schools seldom split. D.L.G., Noakes, B.G. Lindquist, G.S., Helfman and .JA., Ward 1983 193-204 ROBINSON, C.J and T.J PITCHER Journal of Fish Biology Hunger motivation as a prometer of different behaviours; within a shoal of herring: selection for hornogeneity in fish shoal? 1989 459-460 35 SEGHERS, B.H., Evolution Schooling behaviour in the guppy (Poecilia reticulata) an evolutionary response to predation. 1974 486-489 28 SIEGEL, S. and N.J., CASTELLANA Nonparametric statis- the behavioral sciences McGraw-Hill. New York 1988 WERNER, E.E., J.F., GILLIAM, W. HALL and G. G. MITTELBACH, Ecology An experimental test of the effects of predation risk on habitat use in fish. 1983 1540-1548 64
|