|
CORRELACIONES DE PARÁMETROS HIDROBIOLÓGICOS
DE LA LAGUNA DE CELESTÚN, YUCATÁN
Trabajo recibido el 29 de
enero de 1993 y aceptado para su publicación el 15 de mayo de
1994.
Jorge A. Herrera-Silveira
Centro de Investigación y Estudios
Avanzadosdel Instituto Polítécnico Nacional, Mérida,
Apartado 73, CORDEMEX y Biología, UADY, Apartado postal 4-116,
Mérida, Yucatán, MÉXICO.
Se evaluaron los patrones temporales de nutrimentos,
clorofila-a, ciorofilas totales y productividad primaria
bruta fitoplanctónica (PPB) durante un ciclo anual en la Laguna de
Celestún, Yucatán. La información se analizó
construyendo las matrices de correlación. El régimen
climático en la zona de estudio tiene importante influencia en la
magnitud y sentido de las relaciones entre los parámetros para cada
temporada. La laguna presenta un constante gradiente de salinidad de la zona
interna (14-19%,) a la región marina (36-41%). El nitrato y el silicato
reactivo soluble se correlacionaron inversamente con la salinidad, lo cual
sugiere que los manantiales son la principal fuente de estos dos nutrientes.
Sin embargo, las concentraciones del fosfato reactivo soluble y del amonio
podrían estar controladas por la actividad biológica. Estos
procesos podrían favorecer el cambio de nutriente limitante para la
productividad del fitoplancton. En la temporada de secas hay alto número
de correlaciones significativas y las condiciones reflejan estabilidad, con
bajas tasas de producción (100 mgC/m3/día) por parte del
fitoplancton. La descarga de materiales al inicio de la temporada de lluvias
por los manantiales favorece el desarrollo del fitoplancton (847
mgC/m³/día), algunas relaciones cambian el sentido y por lo tanto
la estructura del sistema. En la temporada de nortes las condiciones de
temperatura y viento propician estados de tensión, que relajan las
interacciones. El análisis anual enmáscara las correlaciones que
existen entre los parámetros en cada temporada y resalta la importancia
de tener presente el contexto espacio -temporal en el cual las
características hidrobiológicas están sometidas.
PALABRAS-CLAVE: Nutrientes,
productividad, variación estacional, correlaciones, lagunas
costeras.
The seasonal patterns of nutrients, chlorophyll-a,
total clilorophyl1 and phytoplanktonic gross primary productivity (Pl?B) were
evaluated during an annual cycle in Celestun Lagoon, Yucatan. The observed data
was analized through a correlation matrix. The weather regimen in the study
zone has high influence in the magnitude and sense of relationships between
parameters for each season. The lagoon shows a permanent salinity gradient from
the inner zone (14-19%) to the seaward region (36-41%). Nitrate and soluble
silicate reactive were inversely correlatd with salinity, suggesting to the
springs as the principal source of these two nutrients. However, soluble
reactive fosfate and animoniuni concentrations could be controled by biological
activity. These processes likely favored a shift in nutrient limitant to
phytoplankton productivity. During the dry season, there is a high number of
signíficative correlations and the conditions show stability, with low
production rates of phytoplankton (100 mgC/m³/day). The discharge of
nutrients during the rainy season throgliout springs, promotes the development
of phytoplankton (847 mgC/m³ Iday) and some relationships change of sense
and then, they changed of system structure. Finally, during the Nortes season
the characteristics of temperature and wind, promote stress conditions and the
interactions relax. The annual analysis másks the correlations between
parameters in each season, wherefore the importance to keep in mind the
space-lemporal context in wich the hidrobiological parameters are
submited.
KEY WORDS: Nutrients, productivity,
seasonal variation, correlations, coastal lagoons.
Las interacciones entre diferentes parámetros físicos, químicos y biológicos de un sistema varían de acuerdo con los cambios en las condiciones ambientales globales, lo cual favorece tambien los cambios estacionales en la presencia, abundancia y distribución de los organismos. Con frecuencia las relaciones que hay entre los parámetros hidrobiológicos de sistemás costeros no son tan evidentes como para observarse a través de figuras de dispersión de puntos y, sin embargo, son importantes en la dinámica del sistema, por lo que una alternativa al tratamiento tradicional ha sido emplear el análisis de correlación. Esta estrategia puede ser útil para encontrar el grado de asociación o covariación dentro de una serie de características hidrobiológicas, como un acercamiento a explicar los cambios en el componente biótico del ecosistema. Las correlaciones estadísticas entre los valores de distintos parámetros relativos a propiedades de las aguas se han discutido para su uso en la predicción (Margalef, 1967). Este esquema ha sido propuesto por diversos autores (Cassie, 1963; Moore, 1966; Nixon, 1982). Pero el enfoque de utilizar la información generada durante un cíclo de observaciones, que por lo general es anual, y no considerar que las interacciones están sujetas a cambios de magnitud y de dirección de acuerdo con la estacionalidad ambiental local puede llegar a enmáscarar relaciones importantes; más aún en sistemás con gradientes espaciales, como lagunas costeras y estuarios, que se ven influidos por factores externos como precipitación, insolacion, etcétera. Es evidente que las concentraciones de elementos y organismos en un momento dado no dependen sólo de las condiciones prevalecientes de ese momento en particular, sino también de las que han estado antes ahí o en zonas circundantes, las cuales si no son consideradas en el análisis pueden confundirse estadísticamente. Por lo tanto, se recomienda examinar el contexto espacio-temporal al cual los parámetros hidrobiológicos están sometidos. Este análisis de datos, en combinación con los gradientes espaciales que ocurren en el ecosistema, permite asociar los cambios en la hidrología como una probable respuesta a los cambios del clima local. Si se repiten estaciónalmente las condiciones, se podrán reconocer ciclos intemos del ecosistema, lo que favorece la observación de características estructurales y funcionales de las poblaciones biológicas. Durante el presente estudio se evaluaron características físicas (salinidad, temperatura), concentraciones de oxígeno disuelto, la composición y distribución de los nutrientes esenciales, así como los pigmentos y productividad fítoplanctónica en la Laguna de Celestún, Yucatán. Esta información fue usada para estimar las variaciones estacionales de las correlaciones entre estos parámetros y su asociación con el patrón climático de la zona. ÁREA DE ESTUDIOLa Laguna de Celestún está localizada en la costa oeste de la Península de Yucatán (20°45'N; 90°25'O) y se extiende 22.4 km; es estrecha (entre 0.5 y 2.4 km), somera (de 0.5 a 3.5 m) y con una profundidad media de 1.2 m (Herrera-Silveira, 1988) (Fig. 1). La precipitación en la zona de Celestún promedia 760 mm/año y la temperatura media es de 28.5 °C. Los vientos dominantes son del sureste (0-15 km/h) la mayor parte del año, pero de noviembre a febrero se presentan temporales caracterizados por fuertes vientos del noreste (50-90 km/h) y precipitaciones, asociados a frentes polares y conocidos como "nortes". Estas peculiaridades definen el patrón temporal de la zona de estudio en: temporada de secas (marzo-mayo), de lluvias (junio-octubre) y nortes (noviembre -febrero) (SARH, 1989; Fig. 2). La velocidad de los vientos y la baja profundidad media de la laguna favorecen la mezcla vertical de la columna de agua (Capurro, 1985). El suelo de la región es calizo-kárstico (Back, 1985) y no se presentan corrientes de agua superficiales. Sin embargo, los aportes de agua dulce son a través de afloramientos del manto freático en forma de manantiales dispersos en la laguna. La flora acuática está compuesta por macroalgas (Chara fibrosa, Batophora oesterdi y Chaetomorpha linum) y los pastos acuáticos Halodule wrightú (Herrera-Silveira, 1987) y Ruppia sp. La vegetación que rodea a la laguna consiste en un bosque de manglar (Trejo, 1986). Por su alta diversidad biológica e importancia como zona de alimentación del flamenco rosado Phoenicopterus ruber rijber (Barrios-Espino, 1988) y el pato real Chairina moschata, y por ser hábitat de una gran variedad de especies de aves y reptiles en peligro de extinción, la Laguna de Celestún fue declarada por el gobierno mexicano Reserva Faunística desde 1979. 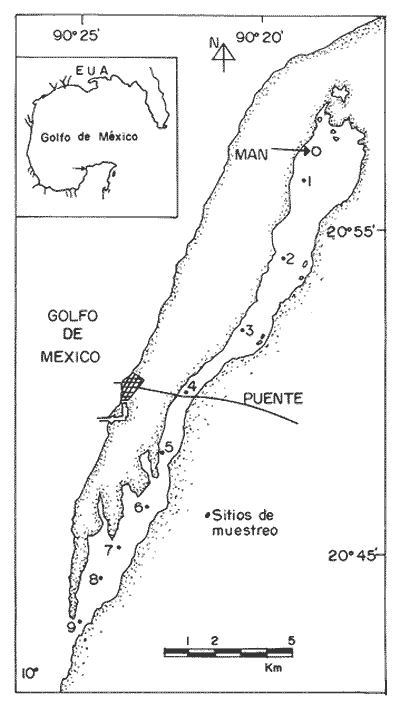 Figura 1. Localización del área de estudio, Laguna de Celestún y sitios de muestreo. 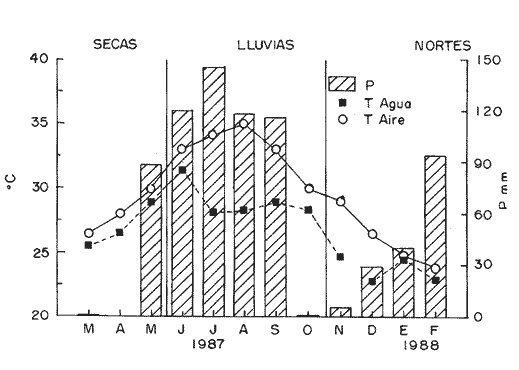 Figura 2. Variación mensual de temperatura del aire, agua y precipitación durante el período de estudio (marzo 1987-febrero 1988). MATERIAL Y MÉTODOSDoce campañas de muestreo mensuales se llevaron a cabo durante un ciclo anual a lo largo de la laguna en diez estaciones y en un manantial (Fig. 1). En cada estación se tomaron muestras a nivel medio de la columna de agua, con la ayuda de una botella van Dorn de 2.5 1 de capacidad. La temperatura (T°) se midió con un termómetro analógico Cole-Palmer (0°- 50°C, ±0.1); la salinidad se determinó usando un salinómetro de inducción Khalsico RS-9, con pre- cisión de 0.001%o (partes por mil). El oxígeno disuelto se evaluó empleando el método de Winkler (Strickland y Parsons, 1972). Las muestras para nutrientes fueron obtenidas a través de filtros de membrana de 0.45µ; éstos fueron utilizados para determinar las concentraciones de clorofila-a (Cla) y clorofilas Totales (CIT; como la suma de las clorofilas a, b y c), por extracción con acetona al 90%, siguiendo la técnica y ecuaciones descritas por Jeffrey y Humphrey (1975). El filtrado se conservó al afiadirle 0.5 ml de cloroformo por cada 250 m1 de muestra (Ho et al., 1970) y se mantuvieron a 4°C hasta su análisis (no más de 48 hr). El fosfato reactivo soluble (SRP). corregido por las interferencias de arsenato/silicato, fue determinado con el método de azul de molibdeno de Murphy y Riley (1962), y modificado por Strickland y Parsons (1972). Las concentraciones de amonio (NH4+) fueron precisadas por el método de fenol-hipoclorito (Solórzano, 1969). El silicato reactivo disuelto (SRSi) fue determinado por el método de azul de molibdeno y se usó ácido ascórbico como reductante. El nitrógeno como nitritos (NO2-) fue analizado por medio del método de sulfánilarnida en solución ácida y los NO3- fueron medidos como nitritos después de su reducción en una columna de Cd-Cu (Parsons et al., 1984). La estimación de la productividad primaria planctónica se realizó mediante el cambio de oxígeno (Vollenweider, 1969), usando botellas DBO de 300 m1 e incubaciones de 3 a 4 hr in situ. El análisis de la información consistió en construir las matrices de correlación entre las variables empleando el coeficiente de Pearson (P<0.01) para los datos de cada temporada (secas 30, lluvias 50 y nortes 40) del ciclo anual (120). Para evitar sesgos debidos a las magnitudes propias de cada variable se utilizaron las transformaciones propuestas por Margalef (1967). La información climatológica proviene de la estación meteorólogica de la Secretaría de Recursos Hidraulicos del Puerto de Celestún. RESULTADOSEl rango anual de variación de la temperatura observada fue de 9°C, con el mayor valor en julio (31.4 °C) y el menor en febrero (22.4°C). Las variaciones estacionales de temperatura del agua y del aire en la temporada siguen el mismo patrón (Fig. 2); las diferencias durante la temporada de lluvias se deben a la intrusión de agua del manto freático, la cual tiene menor temperatura (<20°C) que el agua de la laguna (Tabla 1). El gradiente espacial no refleja cambios importantes a pesar de la baja profundidad de la zona interna (estaciones 1, 2 y 3) con respecto al resto de la laguna (Fig. 3A). La evaporación es favorecida por las altas temperaturas y la baja profundidad media, sin embargo, los aportes del manto freático compensan estas pérdidas. La salinidad presenta un gradiente horizontal durante todo el año. En la zona interna de la laguna la salinidad es siempre menor a 20%o, mientras que en la zona cercana a la boca es mayor de 30% (Fig. 3B). El oxígeno disuelto fluctuó entre 1.5 y 7.5 mg/l. Los porcentajes de saturación de oxígeno fueron de 50 a 170%. Los valores altos corresponden a estaciones cercanas al mar y durante la temporada de nortes (Fig. 3C). 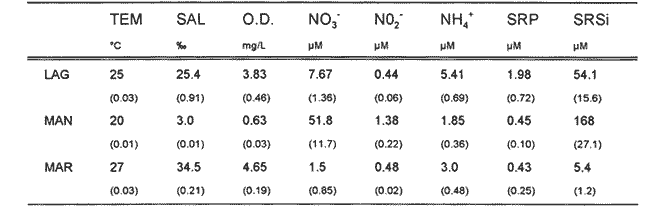 Tabla 1 Media anual, ± un error estándar (en paréntesis), de temperatura (TEM), oxígeno disuelto (O.D.), salinidad (SAL), nitratos (NO3-). nitritos(NO2-),amonio (NH4+), fósfato reactivo soluble (SRP) y silicato reactivo soluble (SRSi), en la laguná (LAG), el manantial (MAN) y el mar, durante el período de estudio. 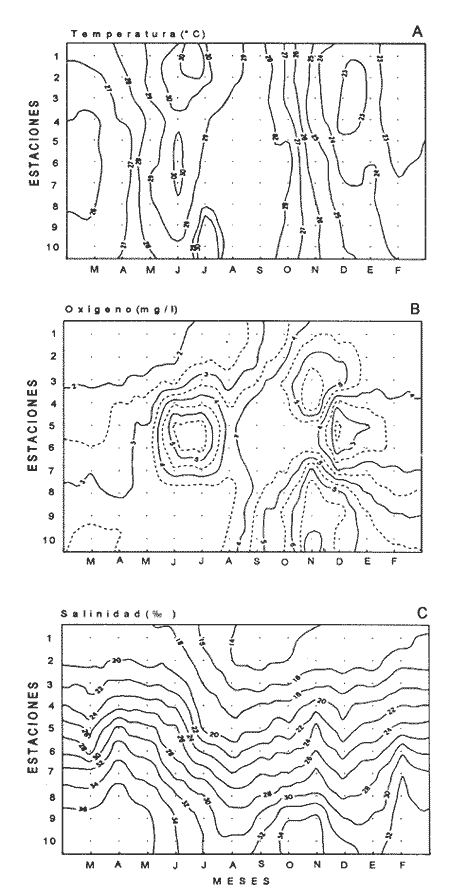 Figura 3. Diagrama espacio-temporal de la temperatura (°C) A,de salinidad %o By de oxígeno disuelto (mg/l) C durante el período de estudio en la laguna de Celestún. Con respecto a los nutrientes, las concentraciones de NO3- Y SRSi presentaron las mayores amplitudes de variación. Los NO3- presentan concentraciones mayores de 40 µM en la estación 1 durante la época de lluvias y <1 µM en las estaciones 9 y 10 durante las épocas de secas y lluvias; este patrón espacial se conserva durante casi todo el año (Figs. 4 y 5). Las concentraciones de SRSi variaron de 1 a >200 IM, siguiendo el mismo comportamiento espacio-temporal de los NO3- durante las temporadas de secas y lluvias (Figs. 4B y 5B). Sin embargo, durante la época de nortes las variaciones de SRSi son mayores que las de N03- en la zona interna. Los NO2- presentan bajas concentraciones (<2 µM) durante todo el ciclo de observaciones. En la época de secas la zona interna presenta las mayores concentraciones del ciclo (1.5 µM). 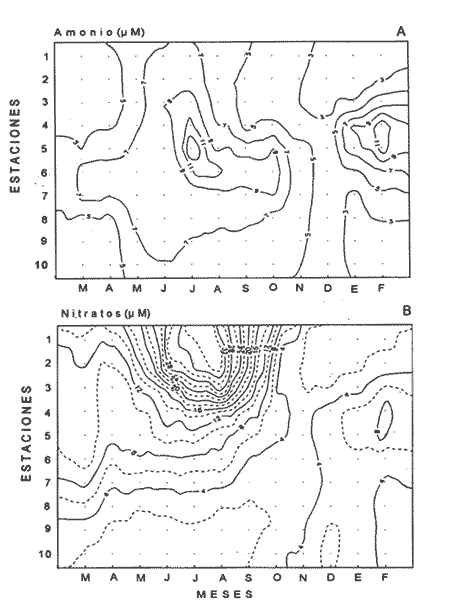 Figura 4. Diagrama espacio-temporal de amonio (µM) A y Nitratos (µM) B, durante el período de estudio de la Laguna de Celestún. Las concentraciones de amonio fueron mayores en julio (15 µM) y febrero (11 µM) en la zona media de la laguna. El gradiente espacial durante gran parte del estudio se observó de la zona interna (estaciones 4, 5, 6 y 7) hacia ambos extremos; sólo durante la temporada de secas este patrón desaparece (Fig. 4A). 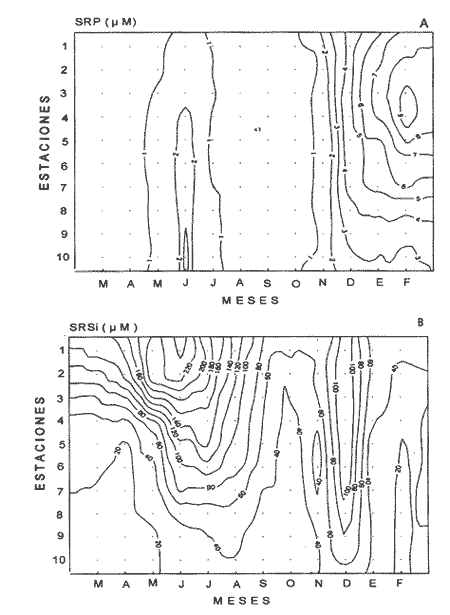 Figura 5 Diagrama espacio-temporal de fosfato reactivo soluble (µM) A, y de silicato reactivo soluble (M) B, durante el período de estudio en la laguna de Celestún. El fosfato reactivo soluble (SRP) está presente en bajas concentraciones (1µM) durante casi todo el año y sin mostrar un gradiente espacial; sólo durante el período de nortes se observaron altas concentraciones en la zona interna (>8µM) (Fig. 5A). Las concentraciones medias de clorofila-a variaron 3 de <1 a 12 mg/m³, fueron mayores en la parte media y menores en la interna. E stacional mente, las concentraciones más altas se dan a principios de la época de lluvias (Fig. 6a). El mismo patrón espacio-temporal lo siguen las concentraciones de clorofilas totales con valores de 5 a 22 mg/m³ . La productividad bruta del fitoplancton tiene el mismo patrón que la clorofila-a; las tasas de producción bruta van de <100 a 1250 mgC/m³/día (Fig. 6b). Del análisis de la correlación anual sólo son significativas la salinidad con el oxígeno (r=0.75), así como entre los pigmentos (CI-a y CIT) y el amonio (r=0.82) (Fig. 8A). 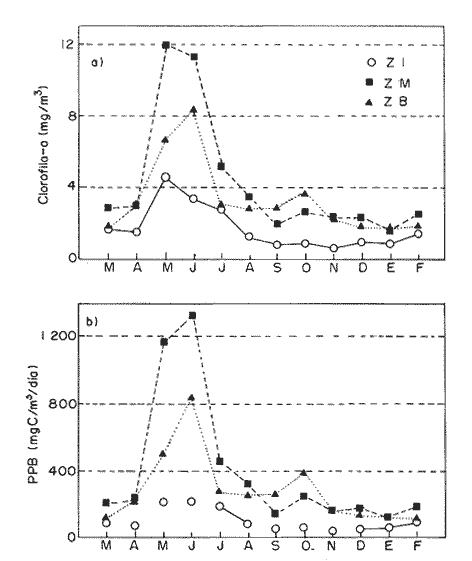 Figura 6. Cambios mensuales de elorotila-a (a) y productividad primaria bruta (b) en el área de estudio. (ZI) zona interna, estaciones 1 a 3; (ZM) zona media, estaciones 4 a 7; (ZB) zona de la boca, estaciones 8 a 10. Al estudiar la información hidrobiológica de acuerdo con las temporadas climáticas de la zona, los patrones difieren de forma importante del análisis anual (Figs. 8B y 9). En la temporada de secas los pigmentos y la PPB estan correlacionados negativamente con los nutrientes (Fig. 8B). Las altas concentraciones de NO3- Y SRSi se presentan en zonas de baja PPB (Figs. 4 y 5). por lo que el componente espacial es un aspecto que se debe tener presente en la interpretación. En la temporada de lluvias el grado y tipo de correlaciones cambian nuevamente (Fig. 8A). Los pigmentos y la PPB están correlacionados positivamente con el NH4+ (r=0.93) y el SRP (r=0.73). Las mayores PPB se evaluaron durante esta temporada (1250 mgC/m3/día). Durante la época de nortes las correlaciones nuevamente tienen cambios importantes (Fig. 8B). Los pigmentos y PPB se correlacionan positivamente con los NO3- (Cla r=0.73; CIT r=0.71; PPB r=0.75) y negativamente con los SRP (r-0.91). En esta temporada se observa un incremento importante en la concentración de SRP, no así en la de N03- (Figs. 4 y 5). 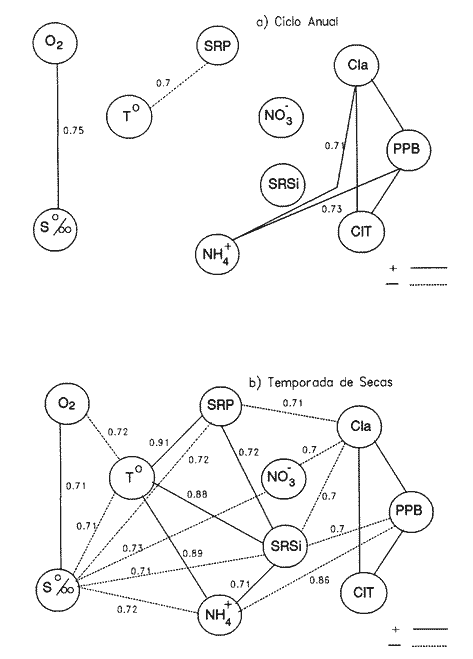 Figura 7. Correlaciones entre parámetros hidrobiológicos durante el cielo anual, a) (marzo 1987-febrero 1988) y la temporada de secas, b) (marzo-mayo). 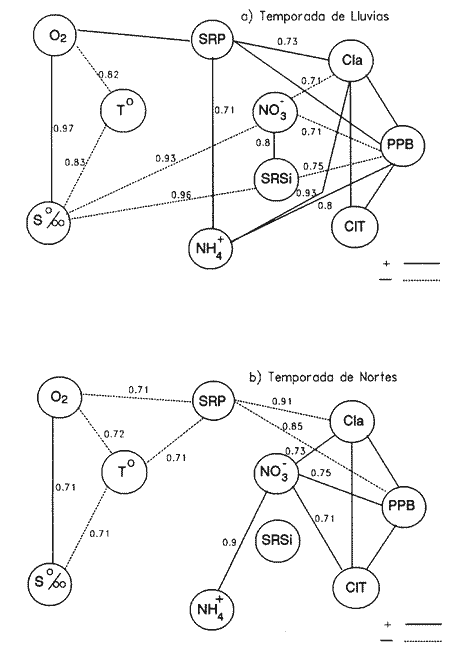 Figura 8. Correlaciones entre parámetros hidrobiológicos durante la temporada de lluvias, a)(junio-octubre) y temporada de nortes,b) (noviembre-febrero). DISCUSIÓN
EL AMBIENTE HIDROLÓGICOLas variaciones en la salinidad, NO3- Y SRSi en la Laguna de Celestún, sugieren el importante papel que los manantiales tienen sobre el comportamiento hídrológico del sistema. El análisis de muestras del manantial y del mar muestra la baja salinidad (3%) y altas concentraciones de NO3- (>10O µM) y SRSi (>300 µM) en el manantial; mientras que las altas safinidades (36-40%o) y bajas concentraciones de nutrientes (NO3- 0.1-2µM; SRSi 2-10 µM) se observaron en las muestras del agua marina (Tabla 1). En otras regiones, como Australia, Israel. Jamaica y California, hay evidencias de descargas de agua dulce a través del manto freático, con bajas salinidades y altas concentraciones de NO3- Y SRSi (D'Elia, 1981; Ronen y Kanfi, 1981, Johanes y Hearn, 1985; Oberdorfer et al., 1990). El intervalo de concentraciones de NO3en Celestún está entre los niveles reportados en otros sistemás del Golfo de México (Tuxpan 40 µM, Chautengo, 100 µM; Contreras, 1983). Las concentraciones de SRSi son mayores que las reportadas para otras lagunas del Golfo de México y el Pacífico (62µM Tuxpan-Tampamachoco Contreras, 1983; 38 µM en Bahía San Quintín, Álvarez-Borrego y Che-Barragán, 1976). pero con similar comportamiento con respecto a los sistemás con afloramientos del manto freático (Australia, 239µM, Johanes y Hearn, 1985-, California, 433µM, Oberdorfer et al., 1990). Los silicatos pueden presentar cambios importantes durante la mezcla de agua dulce y salada en ambientes costeros; estos cambios quizá se deban a la absorción en sedimentos suspendidos (Liss y Spencer, 1970; Bien et al., 1985). En Celestún el gradiente espacial durante las temporadas de secas y lluvias podría estar explicado por los mecanismos de simple dilución. Durante la temporada de nortes los cambios en sus concentraciones probablemente fueron resultado de la regeneración en los sedimentos y al incremento en el transporte vertical (Kamatani y Takano, 1984), a consecuencia de los intensos vientos de esta época. Las concentraciones de SRP en la Laguna de Celestún están en el intervalo citado por Nixon (1982) para lagunas costeras. En zonas donde los aportes de agua dulce se dan a través de ríos las concentraciones de SRP son altas; a estos cuerpos de agua se les considera como exportadores de P en forma particulada (Valiela et al., 1978, Nixon,, 1981). Sin embargo, los manantiales en Celestún no aportan altas concentraciones de SRP (Tabla 1); esto puede deberse a que su alta alcalinidad (Trejo, 1988) favorece la precipitación de los sedimentos (Golterman, 1984). Esta característica sugiere que los procesos de mineralización e importación biológica pueden desempeñar un papel importante en la dinámica de este elemento en la columna de agua. Durante la temporada de nortes la concentración de SRP aumenta signifícativamente y es mayor en la zona interna del sistema. Este incremento puede estar asociado a la mineralización de materia orgánica y al efecto de fertilización de las poblaciones de aves que usan la laguna en esta época. Por ejemplo, la abundancia de flamencos cambió de mil a >19 mil durante la temporada de nortes (Barrios-Espino, 1988). En ambientes templados y subtropicales se ha destacado el papel de las aves en la importación y dinámica de nutrientes mediante la excreción y bioperturbación (Powell et al., 1991; Bildstein et al., 1992). CORRELACIONES ENTRE PARÁMETROSDel análisis anual, la correlación entre pigmentos, PPB y NH4+ insinúa la afinidad del fitoplancton por esta fuente de nitrógeno; característica demostrada experimentalmente (McCarthy et al., 1977; Paasche y Kristiansen, 1982; Bishop et al., 1984). La correlación entre salinidad y oxígeno, así como el gradiente observado de ambos factores sugieren que la mayor actividad heterotrófica fue en la zona interna; los procesos de mezcla y autotróficos dominaron en la zona media y en la boca (Figs. 4, 5 y 6). Durante la época de secas las relaciones positivas se dan entre la temperatura y los nutrientes, lo que podría indicar que la mineralización, principalmente del NH4+ del SRP estaría favorecida por las temperaturas altas. Las poblaciones de macrófitos de la laguna liberan durante esta temporada estructuras epígeas y material particulado (Herrera-Silveira, 1988), el cual podría ser mineralizado y contribuir al aumento de nutrientes en este período. Las oscilaciones del clima son de escasa importancia durante esta época: bajas precipitaciones, vientos débiles y fluctuaciones de la marea poco perceptibles (Capurro, 1985). Estas condiciones de estabilidad de clima pueden favorecer el alto número de correlaciones entre los parámetros químicos, ya que la columna de agua está sujeta a pequeñas variaciones. Durante la temporada de lluvias la correlación entre el fósforo y la PPB sugiere que, a pesar del desfasamiento en la velocidad de respuesta al incremento de P, este elemento podría ser limitante al desaffollo del fitoplancton. La correlación negativa entre pigmentos y PPB con los NO3- (r=0.71) nuevamente se explica por los gradientes en la laguna. La correlación negativa entre la salinidad, NO3y los SRSi (r=0.93 y r=0.96) apoya la observación de que la entrada de agua dulce del manto freático es rica en estos dos nutrientes. En contraste, la falta de relación entre el SRP y la salinidad respalda el planteamiento de la existencia de bajas concentraciones de este nutriente en el agua que aportan los manantiales a la laguna, por lo que los procesos biológicos pueden ser más importantes que las entradas por el manto freático y/o el mar (Tabla 1). Durante la temporada de nortes la mayor oscilación climatológica (descenso de temperatura, precipitación), el aumento en biomása del alga Chara fibrosa en la zona interna (Selem, 1991) y la presencia de abundante fauna ornitológica (Barrios-Espino, 1988) son características que pueden tener gran efecto sobre las correlaciones de esta temporada. La correlación entre las formas nitrogenadas (r=0.9) indica mineralización en la columna de agua. La correlación positiva entre pigmentos y PPB con los NO3- e inversa con el SRP (Fig. 13) indica un cambio en el nutriente limitante para el fitoplancton, que puede ser en esta época el nitrógeno (Fig. 9). 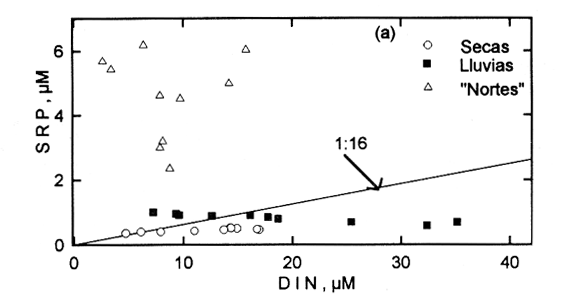 Figura 9. Relación entre DIN:SRP en las tres temporadas climáticas de 1987-1988 de la Laguna de Celestún. Los cambios en la razón nitrógeno inorgánico disuelto (N03-+NO2-+NH4+): fosfato reactivo soluble (DIN:SRP) apoyan la propuesta del cambio en el elemento que sería el limitante para el fitoplancton en cada temporada (Fig. 9). En el grupo de gráficos que componen las correlaciones se observa que las relaciones entre las variables hidrobiológicas cambian siguiendo un patrón asociado a los cambios en las condiciones de clima. Al analizar las correlaciones del ciclo anual sólo algunas son importantes, mientras que al estudiar cada temporada se observan correlaciones que eran enmáscaradas al agrupar la información disponible. Los resultados reflejan que las correlaciones entre parámetros son afectadas no sólo por los cambios temporales, sino que los gradientes espaciales tienen importancia relativa en el significado de las correlaciones. Por ello, el análisis no intenta definir la conveniencia de un procedimiento específico, sino una alternativa que permita obtener de manera sintética patrones sobre el grado de asociación de variables hidrológicas y su covariación con las condiciones ambientales. La Laguna de Celestún presenta un patrón hidrológico algo diferente con respecto a otras lagunas del Golfo de México. Botello y Mandelli (1975), Contreras (1983) y Villalobos et al. (1984) reportan cambios en las variables hidrológicas de lagunas costeras con aportes de agua dulce por ríos, sin resaltar las diferencias que durante la temporada de nortes se observan. Sin embargo, la Laguna de Celestún comparte características hidrológicas con otros sistemás de la Península de Yucatán, en el sentido de presentar manantiales y estar fuertemente influidos por la variabilidad climática (Merino et al., 1990; González et al., 1992). Caffrey y Day (1986) resaltan la importancia de la variabilidad estacional de los eventos meteorológicos sobre la dinámica de nutrientes y sedimentos en suspensión en Atchafalaya River. Por lo tanto, la propuesta de ajustar el análisis de la información en un contexto espacio-temporal puede acercar al entendimiento de la variablidad de estos cuerpos de agua. ConclusionesLos cambios entre las correlaciones en cada temporada sugieren diferentes fases de comportamiento hidrobiológico de la Laguna de Celestún. En la época de secas el agua de mar tiene mayor influencia y la estabilidad en la columna de agua favorece los procesos de mineralización. La temporada de lluvias es más dinámica, los aportes del manto freático son la característica más importante, ya que aumentan las concentraciones de algunos nutrientes y se alcanzan las mayores tasas de producción fitoplanctónica. Durante la temporada de nortes las oscilaciones clima tológicas favorecen la pérdida de materia orgánica en la zona costera. En consecuencia, el comportamiento hidrológico de la Laguna de Celestún refleja, la importancia de los períodos climatológicos sobre las características internas del sistema en el contexto espacio-temporal. La caracterización de esta variabilidad a diferentes escalas de tiempo, acerca al entendimiento de la estructura y dinámica de estos ecosistemás y los recursos que pueden estar asociados a ella. AgradecimientosSe agradece a SEPESCA y DUMAC A.C., Yucatán, por las facilidades prestadas en sus instalaciones; A. Trejo, J. Ramírez y C. Selem por su colaboración en las campañas de muestreo. Al Departamento de Recursos del Mar del CINVESTAV-Mérida por el apoyo otorgado. LITERATURAÁLVAREZ-BORREGO, S. y A. CHEE-BARRAGÁN Ciencias.Marinas Distribución superficial de fosfases y silicatos en Bahía de San Quintín, B. C. 1976 51-61 3 (1) BACK, W. Hydrogeology of the Yucatan. In: Geology and Hydreogeology of the Yucatan and Quaternary Geology of Nirtheastern Yucatan Peninsula, New Orleans Geological Society, New Orleans. 1985 BARRIOS-ESPINO, G.R. MsC.Thesis. Aspects of Ecology of the Carib- Flamingo(Phoenicopterus ruber ruber) in Yucatan, Mexico. Auburn University 1988 65 p. BISHOP. J.J., K.A. EMMANUELE y J.A. YODER Estuaries Nutrient limitation of phytoplankton growth in Georgia nearshore warters. 1984 506-512 7 (4) BILDSTEIN, K.L., E. BLOOD y F. FREDERICK Estuaries The rela- importance of biotic and abiotic vector in nutrienten transport. 1992 147-157 15 (2) BIEN, G.S., D.E DOIS y W.H. THOMAS Geochimica et Cosmochimica Acta The removal of soluble silicas from freshwater entering the sea. 1985 35-54 14 BOTELLO, A,V. y E. F. MANDELLI Final Report Project A study of variables related to the water quality of Terminos Lagoon and adjacent coastal areas, Campeche, Mexico GU CCML, UNAM. 1975 92 p. 853 CAPURRO, L. F Informe técnico. Estudio integral del ecosisteina de Celestún y sus implicaciones en lo socioeconómico.CONACYT clave: PCECCNA-021479 CAPURRO, L. F(Ed) 1985 365 p CAFFREY, J.M. y J.W. DAY Jr. Estuaries Control of the variability of nutrients and suspended sediments in a Gulf Coast Estuary by climatic forcing and spring discharge of the Atchafalaya River. 1986 295-300 9 (4a) CASSIE, M. New Zeland Journal Science Multivariate analysis in the interpretation of numerical plankton data. 1963 36-59 6 CONTRERAS, F. Biótica Variaciones en la hidrología y concentraciones de nutrientes del área estuarino-lagunar de Tuxpan, Tampamachoco, Veracruz, México. 1983 201-213 8 (2) D´ELIA, C.F., K.L WEBB y J.W PORTER Bulletin of Marine Science Nitrate-rich groundwater inputs to Discovery BAy Jamaica: a significant source of N to coral reefs. 1981 903-910 31 GOLTERMAN, H.L. Verhandlungen internationale Vereingung Limnologie Sediments, modifying and equilibrating factors in the chemistry of freswater. 1984 23-59 22 GONZÁLEZ, A., M. MERINO y S. CZITROM An. del Centro de Cienc. del Mar y Limnol. Univ. Nal. Autón. Méx. Laguna Bojórquez, Cancún: un sistema de características marinas controlado por la atmósfera. 1992 59-71 19 (1) HERRERA-SILVEIRA, J. Tesis licenciatura Productividad fitoplarictónica del Estero de Celestún, Yucatán Univ. Autón. de Guadalajara 1987 56p durante el período octubre 1983-abril 1984. ---, Tesis maestría. Productividad Primaria Fitopianctónica de la Laguna de Celestún, Yucatán CINVESTAV-IPN. Unidad Mérida. 1988 126 p. HO, C. L., E. H. SCHEWINGBERG y L. REEVES Coastal Studies Bulletin Chemistry of water and sediments in Barataira Bay. Louisiana State Univ. 1970 41-56 5 JEFFREY, S.W. y G.F. HUMPHREY Bioch. Physiol. Ptlazen New spectrophoto- equations for determining chIorophylls a, b, el and c2 in higher plants, algae and natural phytoplankton. 1975 191-194 167 JOHANES, R.E. y C.J HEARN Estuarine, Coastal and Shelf Science The effect of submarine groundwater discharge on nutrient and salinity regimes in a coastal Lagoon off Pearth Westem, Australia. 1985 789-800 21 KAMATANI, A. y M. TAKANO, Estuarine, Coastal and Shelf Science, The behaviour of dissolved silica during the mixing of river and sea water in Tokyo Bay. 1984.19 505-512 19 LISS. P.S. y C.P. SPENCER Geochimíca et Cosmochimica Acta Abiological processes in the removal of silicate from sea water 1970. 1073-1088 34 McCARTHY, J.J., W.R. TAYLOR y J.L. TAFT Limnology and Oceanography Nitrogenous nutrition of the phytoplankton in the Chesapeake Bay. 1. Nutrient availability and phytoplankton preferences. 1977 996- 1011 22 MARGALEF, R. Inv. Pesq. Correlaciones entre parámetros oceanográficos del Caribe. 1967 1-15 31 (1) MERINO, M., S. CZITROM, E. JORDAN, E. MARTIN, P. THOME y O. MORENO Estuarine, Coastal and Shelf Science Hydrology and Rain Flushing of the Nichupte Lagoon System, Constan, Mexico 1990 223-237 30 MOORE, H.B. Final report of the Institute of Marine Science. University of Miami, submitted to the U.S. Dept. of Navy Mathematical model of environmental in- on phytoplankton movement. 1966 11 p MURPHY, J. y J.P. RILEY Anal. Chem. Acta Modified single solution method for the determination of phosphate in natural waters. 1962 31-36 27 NIXON, S.W. U.S. Fish and Wieldlife Service, Official Biology Service, Publication. FWS/OBS-81/04 In: R. Cross y D. William (Eds.) Freshwater inputs and estuarine productivity. Proceedins of the National Sympisium. on Freshwater Flow to Estuaries. 1981 31-57 NIXON, S.W. Oceanologica Acta, Nutrient dynamies, primary production and fisheries yields of lagoons. 1982 357-371 Special Vol. OBERDORFER, J.A., M.A. VALENTINO y S.V. SMITH Biogeocheinistry Groundwater contribution to the nutrient budget of Tomales Bay, California. 1990 199-216 10 (3) PAASCHE, E. y S. KRISTIANSEN Estuarine, Coastal and Shelf Science Nitrogen nutrition of the phytoplankton in the Oslofjord. 1982 237-249 14 PARSONS, T.R., Y. MAITA y C.M LALLI Pergamon Press A manual of chemical and biological methods for seawater analysis. 1984 173 p POWEL, G.V.N., J.W. FOURQREAN, W.J KENWORTHY y C. ZIEMAN Estuarine Coastal and Shelf Science Bird Colonies cause seagrass enrichment in a subtropical estuary: Observational and Experimental Evidence 1991 567-579 32 SARH. Resumen anual de temperatura, precipitación, evaporación y vientos de la costa norte de la Península de Yucatán Secretaría de Agricultura y Recursos Hidráulicos, Delegación Yucatán. 1989 125 p SELEM, C. Tesis de licenciatura, Univ. Autón. de Yucatán Contribución al conocimiento de los macrófitos en la Laguna de Celestún. 1991 85 p SOLÓRZANO, L. Limnology, and Oceanography Determination of ammonia in natural water by the phenolhypochlorite method. 1969 799-801 14 STRICKLAND, J.D.H. y T.R. PARSONS Fish. Res. Board. Can. Bull. A Practical Handbook of Sea Water Analysis. Ottawa, Canada 1972 311 p 122 TREJO, A. Reporte de Servicio Social. Univ. Auton. Metropolitana-Iztapalapa Estudio de la vegetación de la zona costera inundable perteneciente a los bordes de la Laguna de Celes- Yuc., México. Los Manglares. 1986 30 p TREJO, J. Tesis de maestría (Biología Marina), CINVESTAV-IPN, Mérida Estudio del sistema CO2-carbonato en la Laguna de Celestún, Yucatán. 1988 65 p VALIELA,I., J.M. TEAL, S. VOLKMAN, D. SHAFER y E.J. CARPENTER Limnology and Oceanography, Nutrient and particulate fluxes in a salt marsh ecosystem: Tidal exchanges and inputs by precipitation and groundwater. 1978 798-812 23 (4) VILLALOBOS, F.A., V.R. DE LA PARRA, P.B.E. GALVÁN, R.J.O. CACHO, M.A.P. IZAGUIRRE Cuadernos de Divulgación Estudio hidrobiológico en la Laguna de la Mancha, municipio de Actopan, Veracruz. INIREB-Xalapa. 1984 51 p.. 15 1979-1980 VOLLENWEIDER, R.A. Blackwell Scientific VOLLENWEIDER, R.A. (Ed.), A manual on methods for measuring primary production in aquatie environments. .I.B.P Handbook 12. Oxford; 1969 213 p
|