|
EDAD Y CRECIMIENTO DE LARVAS DE Achirus
lineatus, Etropus crossotus, Syacium gunteri y Symphurus civitatus EN EL SUR
DEL GOLFO DE MÉXICO
Trabajo recibido el 19 de junio de 1992 y
aceptado para su publicación el 14 de octubre de 1992.
AGE AND GROWTH OF Achirus lineatus, Etropus crossotus, Syacium gunteri AND Symphurus civitatus LARVAE IN SOUTHERN GULF OF MEXICO
César Flores-Coto, Arturo Sánchez-lturbe, Faustino Zavala-García y David López-García
Instituto de Ciencias del Mar
y Limnología, Universidad Nacional Autónoma de México.
Apartado Postal 70-305, México, D.F. 04510, MÉXICO.
Contribución número 724 del Instituto de Ciencias del Mar y
Limnología
Se determinaron la edad y el
crecimiento de larvas de cuatro especies de pleuronectiformes, a partir de
colecciones ictioplanctónicas tomadas en el sur del Golfo de
México entre 1984 y 1988. La edad se determinó como la cantidad
de anillos de crecimiento diario, más dos, leídos en los otolitos
sagitta de las larvas. Las edades para las tallas extremas
registradas fueron: Achirus lineatus, 6-23 d (1.4-2.3 mm);
Etropus crossotus, 4-30 d (2.0-6.8 mm); Syacium
gunteri, 7-40 d (2.0-10 mm), y Symphurus
civitatus, 13-32 d (2.1-6.8 mm). El crecimiento se ajustó a un
modelo lineal en los casos de A. lineatus y E.
crossotus, con tasas de 0.10 y 0.20 respectivamente, y a uno
exponencial para S. civitatus y S.
gunteri, que tuvieron tasas diarias de crecimiento longitudinal de
alrededor de 5%. Achirus lineatus fue la especie que tuvo la
menor tasa de crecimiento, atribuible a que se registraron sólo tallas
muy pequeñas (< 3.3 mm), etapa en la que el crecimiento es más
lento. Tallas tan pequeñas en el plancton derivan del hecho que las
especies costeras como ésta cambian muy pronto a hábitos
bénticos, en contraste con S. gunteri, cuya tasa de
crecimiento superior puede estar ligada a abundancias y dispersiones mayores en
aguas oceánicas, permaneciendo en el plancton por más
tiempo.
Age and growth of early life history stages of four
Pleuronectiformes species were determined using ichthyoplank-tonic samples from
the Southern Gulf of México, collected out between 1984 and 1988. The
ages were determined from the daily growth increments on
sagitta otolith of larvae. The ages of the extreme sizes
recorded were Achirus lineatus, 6-23 d (1 .4-2.3 mm);
Etropus crossotus, 4-30 d (2.0-6.8 mm); Syacium
gunteri, 7-40 d (2.0- 10 mm), and Symphurus
civitatus, 13-32 d (2.1-6.8 mm). The growth of A.
lineatus and E. crossotus fits up to a lineal
model, with rates of 0. 10 and 0.20, respectively, meanwhile S.
gunteri and S. civitatus, growth fits up to an
exponential model, with an instantaneous growth rate - 5% daily.
Achirus lineatus showed the lowest growth rate, probably as
a result of the very small sizes of the captured larvae (< 3.3 mm), when the
growth rate is slow. The small sizes in the plankton collection are the
consequence that habits of coastal species as A. lineatus,
soon change to benthonic habits. On the other hand, S.
gunteri had a higher growth rate, which could be related to its
higher abundance and dispersion, remaining longer in the plankton.
El análisis de la edad de los peces se considera básico en las investigaciones acerca de la dinámica de poblaciones, pues permite calcular de manera más precisa el crecimiento y la mortalidad. Estos parámetros resultan muy importantes en el conocimiento integral de las primeras etapas de vida de los peces, cuya sobrevivencia determinará en gran medida la abundancia de adultos de las generaciones siguientes. En los decenios más recientes se ha tenido un avance significativo en la implementación de métodos que permiten la determinación de la edad de las larvas de peces; destacan los que se basan en las marcas diarias (anillos) en los otolitos, sobre todo por la facilidad de su lectura, aun en organismos jóvenes (Panella, 1971, 1974, 1980; Barkman, 1978; Brothers, 1978; Radtke y Dean, 1982; Warlen, 1982; Tzeng y Yu, 1988). Existe aceptación amplia del método, en función de un número cada vez mayor de trabajos que confirman la periodicidad diaria de las marcas (Peters et al., 1978; Fives y Warlen, 1986; Messieh, et al., 1987; Savoy y Crecco, 1987; Simoneaux y Warlen, 1987; Moksness y Wespestad, 1989; Parson y Peters, 1989; Siegfried y Weinstein, 1989; Hales y Hurley, 1991). Los peces planos (Pleuronectiformes) constituyen un porcentaje elevado de la fauna acompañante en las capturas comerciales de camarón en el sur del Golfo de México. Aunque no hay datos específicos de su captura, comúnmente muchas especies se consumen y constituyen una pesquería potencial (Flores-Coto et al., 1991). La información relativa a sus etapas larvarias es muy escasa; en particular, nada existe referente a las tasas de crecimiento de 15 de las 16 especies registradas en el sur del Golfo de México (Arias-Hernández, 1989; Flores-Coto et al., 1991). La excepción es el caso de Achirus lineatus, especie sobre la cual Houde et al., (1970), con trabajo experimental de laboratorio, determinaron las tasas de crecimiento y mortalidad de las larvas. Por lo anterior, como objetivo del presente trabajo se plantea determinar la relación entre la edad y la longitud de las larvas para estimar las tasas de crecimiento de las especies Achirus lineatus (Soleidae), Syacium gunteri, Etropus crossotus (Bothidae) y Symphurus civitatus (Cynoglossidae), seleccionadas en virtud de su abundancia en el sur del Golfo de México. MATERIAL Y MÉTODOSLas larvas de S. gunteri, E. crossotus y S. civitatus empleadas en este trabajo provinieron de muestras de zooplancton recolectadas en el sur del Golfo de México, con red Bongo (505 µm de malla), a bordo del buque oceanográfico "Justo Sierra", durante seis campañas oceanográficas realizadas entre abril de 1984 y agosto de 1988. Las de A. lineatus se recolectaron en la zona litoral adyacente a la Laguna de Términos, Campeche, durante seis campañas bimestrales, de julio de 1986 a mayo de 1987. Las muestras de zooplancton provenientes de las diversas campañas se fijaron con formol al 4%, neutralizado con borato de sodio; posteriormente se extrajeron todas las larvas de peces y se trasvasaron para su almacenamiento en alcohol al 70%. Las larvas se identificaron a nivel de especie y de cada una se midió su longitud patrón, tomada de la punta del hocico al extremo final de la notocorda o del complejo hiputal (Houde et al., 1970). Se seleccionó una serie de larvas que representara adecuadamente el intervalo de tallas presentes de cada especie. Con ayuda de un microscopio de disección, a cada ejemplar se le extrajeron los otolitos sagitta, de la cápsula ótica, y se montaron en un portaobjetos con bálsamo de Canadá, para la posterior lectura de los anillos (marcas diarias de crecimiento). Utilizando un microscopio óptico, se contaron los anillos o marcas diarias de cada otolito, en series de cinco lecturas en sesiones diferentes y por lectores diversos. Para cada una de las especies se analizaron los datos de la longitud patrón y el promedio de las cinco lecturas de anillos en los otolitos, mediante varios modelos (lineal, exponencial, de Gompertz, y el de von Bertalanffy) hasta encontrar el mejor, según dos criterios: 1) los ajustes por minimos cuadrados, de los nuevos valores de la longitud patrón, a la dispersión de los datos originales; 2) el valor resultante de la ordenada (b), que representa la talla de eclosión. 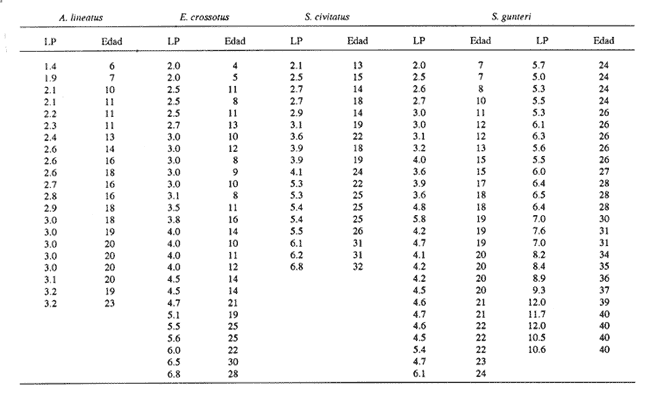 TABLA 1. Longitud patrón (LP) observada y número de anillos de crecimiento + 2 (edad, en días) de las larvas de Achirus lineatus, Etropus crossotus, Symphurus civitatus y Syacium gunteri, empleadas en los modelos de crecimiento. RESULTADOSSe analizaron los otolitos sagitta de 118 organismos. Las tallas de las larvas incluyeron las más pequeñas y las más grandes obtenidas en las recolectas: A. lineatus, 1.4 a 3.2 mm; E. crossotus, 2.0 a 6.8 mm; S. civitatus, 2.1 a 6.8 mm, y S. gunteri, de 2.0 a 10.6 mm (Tabla 1). En ningún caso se registraron larvas recién eclosionadas ni juveniles. Los especímenes más grandes fueron de S. gunteri, que se encontraban en el período de transformación. Las larvas menores de 3 mm correspondieron al período de preflexión. Tomando en cuenta que las larvas más pequeñas analizadas tenían ya varios anillos en sus otolitos, la talla de eclosión necesariamente debe ser menor; por lo tanto, en el ajuste de los distintos modelos para el crecimiento de las larvas, se consideró la dispersión de los datos originales, principalmente el valor resultante de la ordenada, que equivale a la talla de eclosión. En todos los casos se agregaron dos días al número de anillos de crecimiento registrados; de este modo la talla de eclosión derivada del modelo fue menor que la de la larva más pequeña empleada y permitió que la estimación de la edad de las larvas fuera más cercana a la real. Las edades (en días) determinadas para las tallas extremas mencionadas arriba, de las larvas ernpleadas en los modelos (Tabla 1) fluctuaron de 6 a 23 d, 4 a 30 d, 13 a 32 d y de 7 a 40 d para A. lineatus, E. crossotus, S. civitatus y S. gunteri, respectivamente. El crecimiento de las especies analizadas tuvo su mejor ajuste a un modelo lineal en los casos de A. lineatus y E. crossotus (Fig. 1 A y B) y a uno exponencial en las especies S. civitatus y S. gunteri (Fig. 2A y B). La correlación entre longitud y edad fue alta, siempre mayor de 0.90 (Tabla 2). En todos los casos los valores obtenidos de t fueron mayores que los tabulados para p=0.001; por lo tanto las regresiones fueron altamente significativas. 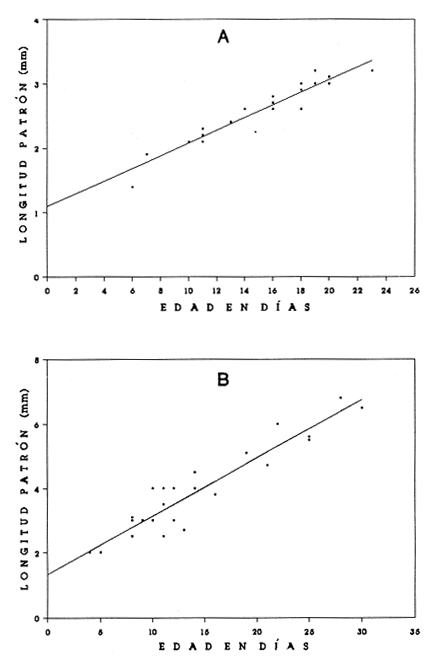 Figura 1. Crecimiento de larvas de Achirus lineatus (A) y Etropus crossotus (B), recolectadas en el sur del Golfo de México. Para describirlos datos se usó un modelo líneal LP = a + bx. 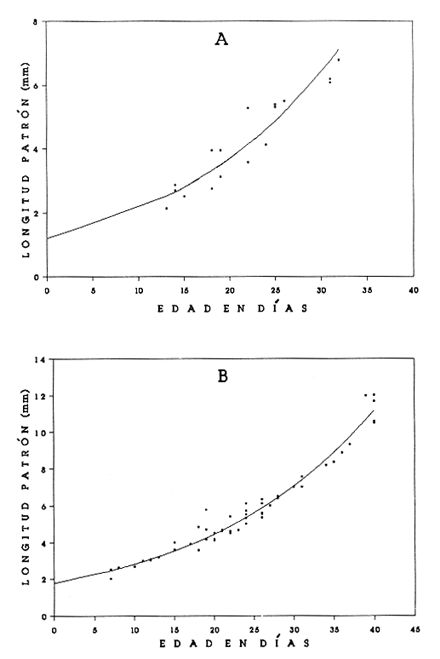 Figura 2. Crecimiento de larvas de Symphurus civitatus (A) y Syacium gunteri (B), recolectadas en el sur del Golfo de México. Para describirlos datos se usó un modelo lineal: LP = a (ebx). La tasa de crecimiento, constante en el intervalo de tallas disponibles, según el ajuste lineal fue de 0.098 mm/d para A. lineatus y 0.20 mm/d para E. crossotus. Las especies S. civitatus y S. gunteri, cuyo crecimiento fue exponencial, tuvieron coeficientes instantáneos de crecimiento muy similares, 0.055 y 0.046, respectivamente, que representan incrementos diarios de su longitud patrón de alrededor del 5.0%. De acuerdo con el modelo ajustado, el crecimiento de S. civitatus se incrementó de 0.08 mm/d, en edades menores de 5 d, hasta 0.33 mm/d a los 30 d. Para S. gunteri el incremento fue de 0. 10 mm/d en edades menores de 5 d hasta 0.48 mm/d a la edad de 40 d. DISCUSIÓNLa edad de las larvas se determinó a partir de marcas de crecimiento (anillos) en los otolitos, aunque no se ha validado la periodicidad diaria de éstas, para ninguna de las especies en estudio; sin embargo, la amplia literatura convalidatoria de tal condición de los anillos de crecimiento, en las larvas de prácticamente todas las especies estudiadas (Peters et al., 1978; Fives y Warlen, 1986; Savoy y Crecco, 1987; Messieh et al., 1987; Simoneaux y Warlen 1987; Moksness y Wespestad, 1989; Parsons y Peters, 1989; Siegfried y Weinstein, 1989; Hales y Hurley, 1991), permite asumir que tal condición prevalece en estos taxa. Por otro lado, tampoco se ha determinado para las especies de este estudio la cantidad de días que transcurren entre el momento de la eclosión y la aparición de la primera marca; ésta generalmente ocurre al término de la formación de la mandíbula, suceso concurrente con el agotamiento del vitelo y el inicio de la alimentación exógena de las larvas. Este período es muy variable; hasta donde se registra en la literatura, puede ser desde uno hasta 17 días, dependiendo sobre todo de la temperatura (Peters et al., 1978; Fives y Warlen, 1986; Messieh et al., 1987; Moksness, y Wespestad, 1989; Powles y Warlen, 1988; Siegfried y Weinstein, 1989; Kramer, 1991).
Una manera de saber el número de días que transcurren
entre la eclosión y la primera marca es incrementar días a la
cantidad de anillos leídos en los otolitos, para que con el modelo que
ajuste mejor se obtenga una ordenada que corresponda con la talla de
eclosión. Sin embargo, no existen registros de ésta referentes a
las especies estudiadas, excepto para A. lineatus: 1.59 mm
(Houde et al., 1970), que empero no pudo emplearse por
resultar mayor que la larva más pequeña en el material de este
estudio, que ya tenía cuatro anillos. Por lo anterior, estrictamente no
se tuvo un punto de partida para calcular el número de días
previos a la primera marca y definir con precisión la edad de las
larvas. Sin embargo, la decisión de aumentar dos días y obtener
la edad verdadera de las larvas (Tabla 1) de las especies estudiadas
derivó del caso similar de Chloroscombrus chrysurus
en la misma área de estudio, para la cual, conociendo su talla de
eclosión (0.8- 1.0 mm), hubo de agregarse dos días a la cantidad
de anillos registrados para obtener este valor en el modelo (Sánchez
Ramírez y Flores-Coto, datos inéditos).
Edad y crecimiento
de Chloroscombrus chrysurus.
Inédito.
Por otro lado, el número de días previos a la formación del primer anillo parece ser menor en las poblaciones ícticas del Golfo de México que en las de latitudes mayores, como se deduce de las tallas de eclosión mucho más pequeñas obtenidas de modelos con larvas del Golfo de México (Cowan, 1988; Weihzong et al., 1992), que las tallas observadas en cultivos experimentales en latitudes mayores en costas atlánticas de los Estados Unidos de América, áreas para las que se registran de 4 a 5 días previos a la aparición del primer anillo (Peters et al., 1978; Warlen, 1982). Las tasas de crecimiento obtenidas para las especies estudiadas (Tabla 2) resultan similares a las registradas para otras especies; así por ejemplo, Fives y Warlen (1986) refieren que, en las costas orientales de los Estados Unidos de América, Anchoa mitchilli crece alrededor de 4.0%/d; Warlen y Chester (1985) encuentran que en la misma área Leiostomus xanthurus crece desde 1.5 %/d en sus tallas menores hasta 7 %/d, alrededor de 17-19 mm LP (longitud patrón). En el norte del Golfo de México, Cowan (1988) encuentra para Micropogonias undulatus tasas de 0. 16 a 0.27 mm/d; y Houde et al., (1970) registran, de sus experimentos de laboratorio, que A. lineatus crece 6 % diario entre la eclosión y los 27 días (8.73 mm). Esta especie tuvo la tasa menor de crecimiento, lo que puede atribuirse a que sólo se econtraron tallas muy pequeñas (< 3.3 mm), en cuya etapa el crecimiento es más lento, antes de entrar a una fase exponencial como lo refieren Houde et al., (1970). Estas tallas pequeñas corresponden con lo señalado por Lasker et al., (1981) en el sentido de que poblaciones de larvas de peces de aguas someras o estuarinas se transforman y tienden al fondo en tallas más pequeñas que las de aguas profundas, como un mecanismo adaptativo que evita su dispersión hacia zonas oceánicas no aptas para su desarrollo. Por otro lado, la menor talla de eclosión de A. lineatus estimada en este trabajo, respecto al valor obtenido por Houde et al., (1970), podría deberse a la temperatura, que para el caso de los organismos cultivados fue de alrededor de 27°C, mientras que para el de las colectas en este trabajo fue de 29°C, y según Tzeng y Yu (1988) podría esperarse una eclosión más rápida. 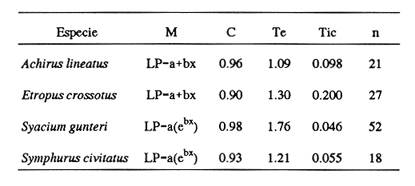 TABLA 2. Modelos (M), parametros de crecimiento (C= correlación de longitud-edad, Te = talla de eclosión, en mm; Tic = tasa instantánea de crecimiento) y número de observaciones (n) de larvas de cuatro especies de pleuronectiformes del sur del Golfo de México. Las larvas más grandes y las mayores tasas de crecimiento, registradas para S. civitatus y S. gunteri podrían relacionarse con su mayor abundancia y distribución en el área de estudio (Arias-Hernández, 1989; Flores-Coto et al., 1991), al considerar el crecimiento como factor de dominancia; ya que, además de permitirles sobrepasar las primeras tallas con mayor rapidez y con esto incrementar su sobrevivencia en las primeras etapas, favorece también la dispersión a otras áreas, al mantenerse en el plancton en estado larvario. La dispersión de los datos obtenidos para cada especie puede atribuirse parcialmente a que provienen de años, períodos y estaciones de colecta diversos donde las larvas estuvieron sujetas a condiciones externas distintas, como disponibilidad de alimento, temperatura, etc.; pero por otro lado se debe a factores (poblacionales) intrínsecos de las especies que desempeñan una función importante en la variabilidad del crecimiento. Discernir entre el efecto de unos y otros factores requeriría de trabajos experimentales. El uso de organismos de períodos diversos en este trabajo permite el establecimiento de una tasa general que se aproxima a un crecimiento promedio de la población de larvas silvestres de cada especie, en la zona estudiada. AgradecimientosLos autores desean agradecer al personal del buque ocenográfico (B.O.) Justo Sierra por su colaboración en el trabajo de campo, y CONACyT por el financiamiento del Proyecto PCCNCNA-031676, del cual es parte el presente trabajo. LITERATURAARIAS-HERNÁNDEZ, L., Tesis Profesional. Abundancia y distribución larvaria de las familias Bothidae, Pleuronectidae y Cynoglossidae (Pisces) en la Sonda de Campeche (1980-1982). Facultad de Ciencias. Universidad Nacional Autónoma de México. 1989 77 BARKMAN, R. C., The use of otolith growth rings to age young Atlantic silverside, Menidia menidia. Trans. Am. Fish. Soc., 1978 790-792 107 6 BROTHERS, E. B., Exogenous factors and the formation of daily and subdaily growth increments in fish otoliths. Am. Zool, 1978 613 18 COWAN, J. H., Bull. of Mar. Sci., Age and growth of Atlantic croaker Micropogonias undulatus, larvae collected in the coastal waters of the northem Gulf of Mexico as determined by the increments in saccular otoliths. 1988 349-357 42 3 FIVES, J. M., and S. M. WARLEN, Estuaries Aging and growth of larval Bay anchovy Anchoa mitchilli, from New Port River estuary. North Carolina. 1986 362-367 9 4B FLORES-COTO, C., F. ABUNDIO-LÓPEZ, and F. ZAVALA-GARCÍA Estuarine, Coastal and Shelf Science, Larval distribution and abundance of Pleuronectiformes from the southern Gulf of México. 1991 439-450 32 HALES, L. S., and D. H. HURLEY, Estuaries, Validation of daily increment formation in the otoliths of juvenile silver perch Bairdiella chrysoura. 1991 199-206 14 2 HOUDE, E., C. R. FUTCH, and R. DETWILER, Development of the líned sole, Achirus lineatus, described from laboratory reared and Tampa Bay specimens. Dep. of Nat. Resources, Tech. Ser. 1970 42 62 KRAMER, S. H., Fish. Bull., Growth mortality and movements of juvenile California halibut Paralicthys californicus in hollow coastal and bay habits of San Diego County, California. 1991 195-207 89 2 LASKER, R., P. E. SMITH, J. R. HUNTER, and H. G. MOSER, Marine Fish Larvae. R. Lasker (Ed.). University of Washington. 1981 131 MESSIEH, S. N., D. S. MOORE, and P. RUBEC The age and growth of fish. Estimation of age and growth of larval Atlantic herring as inferred from examination of daily growth increments of otoliths. In: R. C. Summerfelt, and G. E. Hall (Eds.). The Iowa State University Press, 1987 433-442 MOKSNESS, E., and V. WESPESTAD Fish. Bull. Ageing and back-calculating growth rates of Pacific herring Clupea pallasii larvae by reading daily otolith increments. 1989 509-513 87 3 PANELLA, G., Science, Fish otoliths: Daily Growth Layers and Periodical Patterns. 1971 1124-1127 173 ____, The Ageing of fishes. Mem. of International Symposium. Otolith growth patterns: an aid age detemination in temperate and tropical fishes. In: T. Bagenal (Ed.). Univ. Read. England 1974 28-39 19-20 Jul. 1973 _____, Skeletal growth of aqualic organisms; biological records of environmental change. Growth paterns in fish sagittae. In: D.C Rhoads, and R.A. Lutz (Eds.). Plenum Press, New York, 1980 519-560 PARSON, G. R., and K. M. PETERS, Fish. Bull., Age determination in larval and juvenile sheephead Archosargus probatocephalus. 1989 985-988 87 PETERS, D. S., J. C. DeVANE, M. T. BOYD, L. C. CLEMEMS, and A. B. POWELL, In: Annual Report of the NMFS, Beaufort Laboratory, Beaufort, N.C., to US Department of Energy, Preliminary observations on feeding, growth and energy budget of larval spot (Leiostomus xanthurus). 1978 377-397 POWLES, M. P., and S. M. WARLEN, American Fisheries Society Symposium, Estimation of hatch periods for yellow perch, based on otolith readings from juveniles (age-0). 1988 60-67 5 RADTKE, R. L., and J. M. DEAN, Fish. Bull, Increment formation in the otoliths of embryos, larvae and juveniles of the mummichog, Fundulus heteroclitus. 1982 102-216 80 2 SAVOY, T. F., and V. A. CRECCO, The age and growth of fish. Daily increments on the otoliths of larval American shad and their potential use in population dynamics studies. In: R. C. Summerfelt, and G. E. Hall (Eds.). The Iowa State University Press, 1987 413-431 SIEGFRIED, P., and M. D. WEINSTEIN, Estuaries, Validation of daily increments deposition in the otoliths of spot (Leiostomus xanthurus). 1989 180-185 3 2 SIMONEAUX, L. F., and S. M. WARLEN, The age and groswh of fish. Occurrence of daily growth increments in otoliths of juvenile Atlantic menhaden. In: R. C. Summerfelt, and G. E. Hall (Eds.). The Iowa State University Press, 1987 443-451 TZENG, W. N., and S. Y. YU, J. Fish. Biol., Daily growth increments in otoliths of milkfish, Chanos chanos (Forsskal), larvae 1988 495-504 32 WARLEN, S.M., Proceedings of the Annual Conference of Southeastern Association of Fish and Wildlife Agencies Age and growth of larvae and spawning time of Atlantic croaker in North Carolina. 1982 204-214 34 WARLEN, S. M., and A. J. CHESTER, Fish. Bull., Age, growth and distribution of larval spot, Leiostomus xanthurus, off North Carolina. 1985 587-599 4 83 WEIHZONG, C., J. J. GOVONI, and S. M. WARLEN, Fish. Bull., Comparison of feeding and growth of larval round herring Etrumeus teres and Gulf menhaden Brevoortia patronus. 1992 183-189 90 1
|