|
CARACTERIZACIÓN
CITOGENÉTICA DEL MARLÍN BLANCO Tetrapturus albidus Poey, 1860
(PISCES: ISTIOPHORIDAE) DEL CARIBE MEXICANO
Trabajo recibido el 13 de
marzo de 1992 y aceptado para su publicación el 14 de octubre de 1992.
Trabajo presentado en el XI Congreso Nacional de Zoología, efectuado del
28 al 31 de octubre de 1991 en Mérida, Yucatán,
México.
CYTOGENETIC CHARACTERIZATION OF THE SAILFISH Tetrapturus albidus POEY, 1860 (PISCES: ISTIOPHORIDAE) FROM THE MEXICAN CARIBBEAN SEA
Alicia Luz Durán-González y Alfredo Laguarda-Figueras
Instituto de Ciencias del Mar y Limnología, Universidad
Nacional Autónoma de México. Apartado Postal 70-305,
México, D.F. 04510, MÉXICO. Contribución 718 del Instituto
de Ciencias del Mar y Limnología.
Los peces picudos se encuentran
agrupados en la Familia Istiophoridae, pero su posición
taxonómica no ha sido bien establecida. Al respecto, en México se
han realizado pocas investigaciones, a pesar de su considerable importancia
pesquera tanto deportiva como comercial. En el presente trabajo se emplea el
criterio citotaxonómico para contribuir a la solución del
problema, mediante el estudio citogenético de ejemplares de la especie
Tetrapturus albidus, marlín blanco del Mar Caribe
mexicano, capturados durante el Crucero,Oceanográfico TUPICC II, en
1989. El análisis del complemento cromosómico de los organismos
estudiados demuestra la existencia de un número diploide de 48
cromosomas. De éstos, 46 (23 pares) son autosomas: 22 pares
monorrámeos (acrocéntricos) y un solo par birrámeo
(metacéntrico). Los cromosomas sexuales muestran heteromorfismo: la
hembra presenta un par de cromosomas submetacéntricos, y el macho,
heterogamético, posee un cromosoma submetacéntrico y el otro
acrocéntrico. El mayor número total de brazos cromosómicos
(número fundamental) es n. f=52, porque consta de 22 elementos
monorrámeos y 2 birrámeos. La longitud del complemento haploide
es de 26.81 µm. La presencia de un par de autosomas metacéntricos
y del heteromorfismo sexual cromosómico indican un considerable grado
evolutivo de la especie, la cual es especializada en el nivel
citogenético.
The billfishes are gathered into the
Istiophoridae Family, but their taxonomic location is not well established.
With respect to this problem, few studies have been done in México, in
spite of its commercial and sportive fishing importance. In the present work
the cytotaxonomic criterion is employed in order to solve the problem, by means
of the cytogenetic analysis of the specimens of Tetrapturus
albidus, white marlin of the Mexican Caribbean Sea, gathered in the
oceanographic campaign TUPICC II, in 1989. The chromosomic complement of
Tetrapturus albidus shows a diploid number of 2n=48
chromosomes, from which 46 (23 pairs) are autosomes: 22 pairs are monoarmed
(acrocentric) and one pair is biarmed (metacentric). The sexual chromosomes
show heteromorphism: the female shows a pair of submetacentric and the male is
heterogametic, showing one submetacentric and one acrocentric chromosome. The
major chromosome arm number (fundamental number) is f. n. =52, because it
presents 22 monoarmed and 2 biarmed elements. The presence of a metacentric
pair of autosomes and a sexual chromosomic heteromorphism. suggest certain
degree of evolution at cytogenetic level. T. albidus is
specialized, with haploid complement length of 26.81 µm.
Los estudios citogenéticos proporcionan información valiosa que ayuda a ubicar adecuadamente a los organismos en las diversas entidades taxonómicas, mediante el análisis de los cromosomas que constituyen los cariotipos característicos de las poblaciones, razas o subespecies y especies emparentadas, que se adaptan y evolucionan en el espacio y el tiempo. Así mismo, lo anterior ayuda a ordenar sistemáticamente a los seres vivos que integran los distintos taxa (Sota, 1982 y Durán-González et al., 1990). 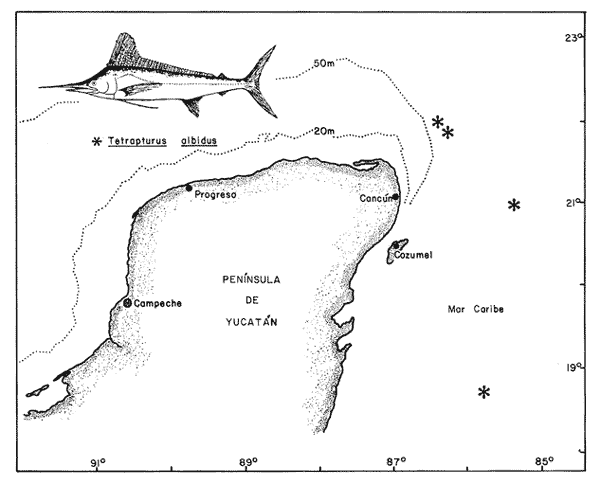 Figura 1. Estaciones de colecta de Tetrapturus albidus (marlín blanco) del TUPPICC-II. Estos estudios se han aplicado con muy buenos resultados para el análisis citotaxonómico de algunas familias de peces, como la Goodeidae y la Cichlidae (Denton, 1973; Thompson, 1979; Uyeno et al., 1983). No obstante, se han hecho pocos trabajos en los peces teleósteos marinos, a pesar de su gran diversidad (Denton, 1973; Durán y Laguarda, 1990). Los teleósteos marinos agrupados en las familias Xiphiidae e Istiophoridae, comúnmente conocidos como picudos, representan un recurso pesquero de importancia considerable, tanto comercial como deportiva, en parte debido al gran tamaño alcanzado por sus integrantes, como el pez espada, el pez vela y el marlín. Sin embargo, aunque su importancia pesquera es grande, se han realizado pocas investigaciones en torno a estas especies, y se desconocen diversas características de su biología (Dickson y Moore, 1977; Polanco et al., 1988), tomando en cuenta que la velocidad de sus desplazamientos, su distribución amplia y su dinámica compleja dificultan precisar su ubicación geográfica y el grado de aislamiento de sus poblaciones. No obstante, diversos autores han realizado contribuciones importantes acerca de la delimitación taxonómica, la nomenclatura y la filogenia de los peces picudos; entre ellas, las de Nakamura (1974 y 1985), Schultz (1987) y Fierstine (1989). A pesar de lo anterior, muchas de las estructuras, consideradas características, que definen algunas de las especies, son completamente similares entre sí y otras presentan cambios drásticos durante el crecimiento. Todo ello, aunado a la falta de conocimiento referente a los ciclos de vida de las especies, genera dificultad para determinar la taxonomía de los integrantes del grupo (Nakarnura, 1985). El género Tetrapturus, de la Familia Istiophoridae, está integrado por seis especies, de las cuales T. georgei requiere de más pruebas para consolidar su posición taxonómica. Además, es posible que exista otra especie, conocida con el nombre de marlín hacha. Todo esto hace variar los criterios relativos a la composición de este grupo (Nakamura, op. cit.). De hecho, se ha propuesto que al género Tetrapturus se le considere como una nueva familia (Tetrapturidae), de acuerdo con la interpretación de la estructura rostral realizada por Schultz (1987). En el Caribe mexicano se encuentran poblaciones de marlín blanco, Tetrapturus albidus, poco estudiadas. El presente trabajo contribuye con la caracterización citogenética de esta especie, a fin de coadyuvar al ordenamiento sistemático de la Familia Istiophoridae. MATERIAL Y MÉTODOSLos organismos estudiados se colectaron durante la campaña oceanográfica TUPICC II —correspondiente al proyecto "Prospección y evaluación de los peces pelágicos mayores del Caribe", de la Estación Puerto Morelos del ICMyL de la UNAM—, efectuada en el buque oceanográfico (B.O.) Justo Sierra en junio de 1989. Para la colecta de los organismos se utilizó un palangre que consta de un cable de 9 km de longitud y 180 anzuelos, cubriendo las estaciones de colecta (Fig. 1). Los especímenes capturados fueron determinados con base en su morfología y datos merísticos; por medio de disección y análisis directo de la gónada se identificaron tres hembras y dos machos. Posteriormente se procesaron los arcos bronquiales correspondientes, mediante las técnicas citogenéticas propuestas por McPhail y Jones (1966), Denton (1973) y Kligerman y Bloom (1977), con las ligeras modificaciones requeridas, de acuerdo con las características del material biológico y las condiciones de trabajo. Las laminillas obtenidas se tiñeron con Giemsa al 10% en solución amortiguadora (buffer) de fosfatos y se revisaron minuciosamente al microscopio óptico. Los campos cromosómicos se seleccionaron y fotografiaron; las fotografías se amplificaron e imprimieron en papel, para armar los cariotipos. Para ello se tomaron los datos merísticos de los cromosomas y se analizaron estadísticamente con el promedio de los resultados se determinaron la proporción de los brazos de los cromosomas, los índices centroméricos y las diferencias respectivas. Lo anterior permitió caracterizar citogenéticamente al material biológico estudiado y obtener el cariotipo y el idiograma correspondientes, de acuerdo con los principios metodológicos propuestos por Levan et al., (1964) y Al Aish (1969). RESULTADOSSe obtuvieron 67 campos cromosómicos, de los cuales se seleccionaron 23 para fotografiarse, amplificarse e imprimirse Se armaron los cariotipos de machos y de hembras; en todos los casos poseían 48 cromosomas en diferentes grados de espiralización (Fig. 2). El heteromorfismo cromosómico caracterizó la presencia de cromosomas sexuales. Los doce complementos cromosómicos, analizados estadísticamente, muestran la presencia de 23 pares de autosomas, de los cuales 22 son monorrámeos (acrocéntricos de tamaños decrecientes entre los dos valores extremos de 1.5 y 0.7 µm, y un par birrámeo (metacéntrico) de 1.2 µm. El par sexual de la hembra es submetacéntrico (1.5 µm) y el del macho, heterogamético, contiene un cromosoma acrocéntríco (1.06 µm) y uno submetacéntrico (1.5 µm). 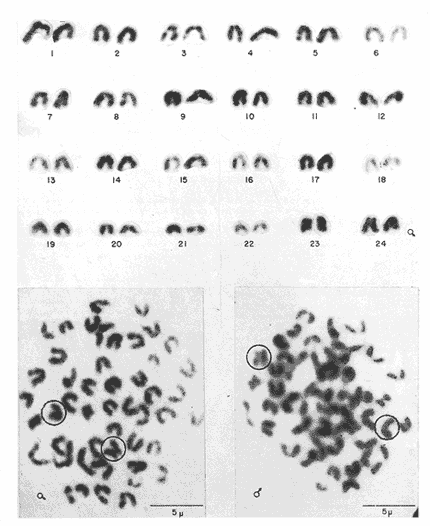 Figura 2 . Cariotipo de Tetrapturus albidus Poey, 1860. La longitud del complemento haploide de Tetrapturus albidus es de 26.81 µm (A+X); el número fundamental de brazos cromosómicos que lo caracterizan es 52 (Tabla 1, Fig. 3). DISCUSIÓNDurante los estudios realizados por Lozano-Álvarez y Negrete (1989) con respecto a la prospección y la evaluación de peces pélagicos mayores del Golfo de México y del Mar Caribe, los picudos aparecieron con mayor frecuencia en el Caribe, aunque suelen estar presentes en ambos mares. En particular durante el crucero oceanográfico TUPICC II, del ICMyL, UNAM, Tetrapturus albidus (marlín blanco) fue la especie más frecuente en la zona del Caribe; se capturaron cinco organismos [tres hembras y dos machos (Fig. 1)]. Las características morfoanatómicas de los especímenes capturados correspondieron claramente a las descritas por Nakamura (1974) y Dickson y Moore (1977) para dicha especie, que desde el punto de vista citogenético manifiesta la presencia de un número diploide de 48 cromosomas. De ellos, 23 pares son autosomas (22 pares acrocéntricos y un par metacéntrico); poseen además un par de cromosomas sexuales: el par sexual de la hembra es submetacéntrico y el del macho contiene un cromosoma acrocéntrico y uno submetacéntrico. 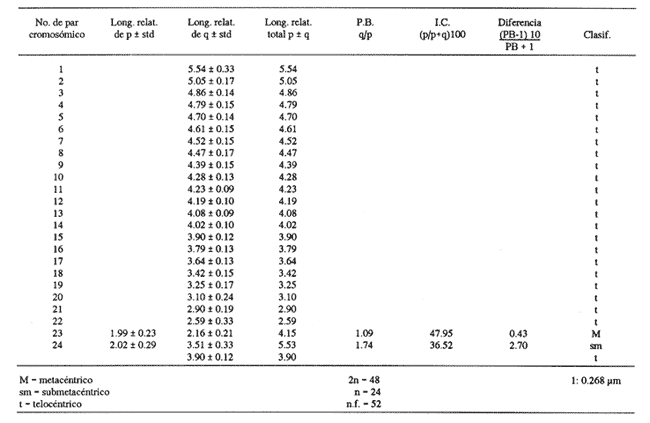 TABLA 1. Resultados del análisis estadístico de los pares cromosómicos de doce cariotipos de Tetrapturus aibidus Autores como Ohno et al., (1968), Thompson (1969) y Uyeno et al., (1983) consideran que el cariotipo ancestral de los peces óseos está constituido por 48 cromosomas acrocéntricos y de éste derivan las otras formas cromosómicas birrámeas metacéntricas y submetacéntricas. Aunque en el cariotipo obtenido existe la presencia de 91.6% de cromosomas acrocéntricos y sólo el 8.4% restante corresponde a las formas birrámeas, esto podría suponer un escaso desarrollo evolutivo de esta especie; sin embargo, datos comparativos demuestran lo contrario. La suma de las medidas promedio obtenidas para los 24 pares cromosómicos de T. albidus determina una longitud de 26.81 µm del complemento haploide (Tabla 1 y Fig. 3), mientras que Haemulon aurolineatum - otro perciforme considerado moderno por Mago (1961)- posee 24 pares de cromosomas acrocéntricos y el promedio de longitud del complemento haploide es 45.72 mm. (Durán-González et al., 1990). Esto indica que el marlín blanco presenta disminución considerable del tamaño del genoma, lo cual está de acuerdo con lo expuesto por Gold (1979), que considera a los perciformes como un orden altamente especializado, con números cromosómicos y tamaño del genoma relativamente homogéneos. Aunque la evaluación cariotípica en los peces es incierta, se nota una tendencia hacia la disminución del tamaño del genoma cuando hay un incremento en la especialización de las especies. 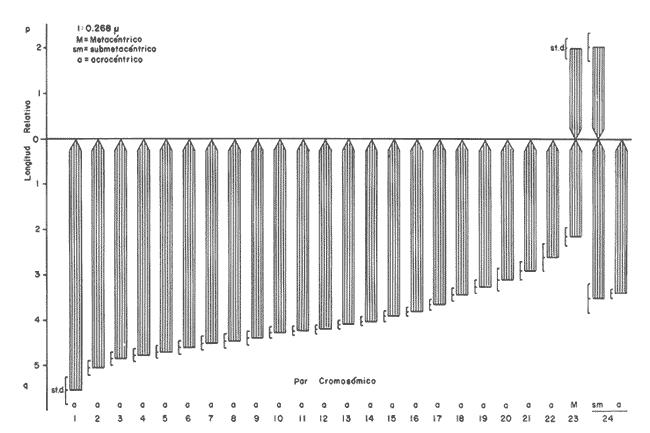 Figura 3. Idiograma del cariotipo de Terrapturus albidus Poey, 1860. La presencia de cromosomas sexuales se determinó por el heteromorfismo cromosómico manifestado en los machos de T. albidus, de acuerdo con el criterio propuesto por Ebeling y Chen (1979): la heterogametia citológica en peces se puede determinar por la evidencia invariable, en células mitóticas, de un par cromosómico heteromórfico en uno de los sexos. Desde el punto de vista citogenético, la confluencia de un par de autosomas metacéntricos, de cromosomas sexuales, de una longitud de 26.81 µm del complemento haploide y de un número fundamental de brazos cromosómicos de 52, denota, de modo considerable, el grado de evolución del genoma de T. albidus, especie que se considera altamente especializada. En la actualidad, las relaciones filogenéticas de la Familia Istiophoridae son tema de discusión amplia, en virtud de que taxonómicamente se encuentra ubicada en el Suborden Scombroidei (Nelson, 1984), lo que "formalmente implicaría una relación directa entre los picudos y los atunes y especies afines, con las consecuencias que conlleva esta consideración en los aspectos administrativos del manejo de estos dos grupos" (Polanco et al., 1988). Al respecto, Collete et al., (1984) tratan acerca de las relaciones filogenéticas de las seis familias y los 44 géneros de escómbridos vivientes y concluyen que Istiophoridae y Xiphiidae son grupos hermanos que a su vez se encuentran emparentados con Scombridae. Mientras que Johnson (1986) reafirma esto último, Fierstine (1989) no considera ciertas estas proposiciones de acuerdo con sus estudios paleontológicos realizados previamente (Fierstine 1974; Fierstine y Applegate, 1974; Fierstine y Welton, 1983), en los que señala la falta de evidencia que atribuya un ancestro común entre dichas familias. En este sentido, aun cuando los antecedentes cariotípicos son inexistentes para los miembros de la Familia Istiophoridae, serían de gran utilidad, pues por medio de estudios electroforéticos experimentados por Hartmann y Waldner (1989) en marlín blanco, marlín azul y pez vela del Océano Atlántico, se obtuvieron los patrones de bandas de las proteínas musculares respectivas, y concluyen que cada especie tiene un patrón característico aunque presenten similitudes considerables. Los estudios citogenéticos, así como los electroforéticos pueden coadyuvar a la solución del problema expuesto, ya que con ellos se pretende caracterizar a las poblaciones actuales con base en marcadores genéticos, los cuales a su vez pueden contribuir a determinar las decisiones deseables de regulación pesquera y protección de este recurso. ConclusionesDe los peces picudos, el marlín blanco Tetrapturus albidus tuvo mayor frecuencia de aparición en el Mar Caribe mexicano. Su cariotipo está conformado por un número diploide (2n) de 48 cromosomas, de los cuales 23 pares son autosómicos y un par sexual (XY). La fórmula cromosómica correspondiente es: 1m + 22t+ XY; con un número fundamental de 52 brazos cromosómicos, y una longitud del complemento haploide de 26.81 µm (A+X). Citogenéticamente, por el grado de reducción del tamaño del genoma y la presencia de formas cromosómicas birrámeas, es una especie altamente especializada. AgradecimientosLos autores agradecen todo el apoyo proporcionado por el personal académico de la Estación Puerto Morelos, del Instituto de Ciencias del Mar y Limnología, UNAM, para la realización del presente trabajo: al M. en C. Enrique Lozano A. (jefe de campaña) al M. en C. Felipe Flores A. (responsable del grupo) y a los Biólogos Fernando Negrete S., Marisa González V. y Miguel Ángel Martinez C. por la determinación del material biólogico. Así mismo, se agradece al personal técnico del Centro Regional de Investigaciones Pesqueras de Isla Mujeres que participó en las colectas de la Campaña Oceanográfica TUPICC-II. LITERATURAAL-AISH, M., I. Studies on normal chromosomes characterization and karyotyping. Human chromosome morphlogy. Can. J. Genet. Cytol. 1969 370-381 11 COLLETE, B. B., T. POTTHOFF, W. J. RICHARDS, S. UEYA-NAGI, J. L. RUSSO, and Y. NISHIKAWA, Ontogeny and systematic of fishes. Scombroidei: development and relationships, In: H. G. Moser et al. (Eds.). Supplement to Copeia, American Society of Ichthyologists and Herpetologists. Special Publication No. I, 1984 591-620. DENTON, T. E., Fish Chromosome Methodology. Charles C. Thomas Publisher, Illinois, USA. 166 DICKSON, H., and R. H. MOORE, Texas. Louisiana and adjacent waters. Fishes of Gulf of México Texas A & M Univ. Press USA. 1977 327 DURÁN-GONZÁLEZ, A. L., and A. LAGUARDA-FIGUERAS An. Inst. Cienc. del Mar y Limnol. Cytogenetic Study of Diplectrum eumelum Rosenblatt and Johnson, 1974 (Pisces: Serranidae). Univ. Nal. Autón. México, 1990 63-72 17 1 DURÁN-GONZÁLEZ, A., C. GARCÍA-RUELAS, and A. LAGUARDA-FIGUERAS An. Inst. Cienc. Mar y Limnol. The karyotype and "G" bands of Heamulon aurolineatum Cuvier, 1829 (Pisces: Haemulidae). Univ. Nal. Autón. México, 1990 299-307 17 2 EBELING, A. W., and T. R. CHEN, Heterogamety in teleostean fishes. Trans. Am. Fish Soc., 1970 131-138 1 FIERSTINE, H. L. Proceeding of the Internacional Billfish Symposium Kailua-Kona, Hawaii, Part. 2. The paleontology of billfish -The state of the art., pp. 34-44. In: R. S. Shomura y F. Williams (Eds.). Review and Contributed Papers, NOAA Tech. Rep. NMFS (Natl. Mar. Fish. Serv.) Spec. Sci. Rep, 1974 675 9-12 August 1972. FIERSTINE, H. L., and S. P. APPLEGATE Acad Sci. Bull., Xiphiorhynchus kimblalocki, a new billfish from the Eocene of Mississipi, with remarks on the systematics of xiphioid fishes. Southern Calif. 1974 14-22 73 FIERSTINE, H. L., and B. J. WELTON, Bull. Mar. Sci., A black marlin Makaira indica, from the Early Pleistocene of the Philippines and the zoogeography of Istiophorid billfishes. 1983 718-728 33 3 FIERSTINE, H. L., A paleontological review of three billfish families (Istiophoridae, Xiphiidae and Xiphiorhynchidae) In: Planning the future of billfishes. Research and management in the 90s and beyond Part. 2: 11-19Contributed paper Kailua-Kona, Hawaii. National Coalition for Marine Conservation,Sabanahh, Georgia 1989. GOLD, J. R., Fish Physiology VIII: 353-405. Cytogenetics. In: W. S. Hoar, D. J. Randall y J R. Brett (Eds.). Academic Press Inc. New York. 1979 HARTMANN, J. X., and R. E. WALDNER, In: Planning the future of billfishes. Resarch and management in the 90s and beyond Part. 2:231-254. Isolation of Biomarker Proteins from Blue Marlin, White Marlin and Sailfish from the Westem Atlantic Ocean: A Means of Identifying Billfish. Carcasses. Contributed paper Kailua-Kona, Hawaii. National Coalition for Marine Conservation, Sabanahh, Georgia. 1989 JOHNSON, G. D. Bull. Mar. Sci., Scombrid phylogeny: an alternative hypothesis. 1986 1-41 39 1 KLIGERMAN, A. D., and S. E. BLOMM, Rapid chromosome preparations from solid tissues of fishes. J. Fish. Res. Board Can., 1977 266-269 34 LEVAN, A., K. FREDGA, and A. A. SANDBERG Hereditas, Nomenclature for Centromeric Position of Chromosomes. 1964 201-220 52 LOZANO-ÁLVAREZ, E., y F. NEGRETE-SOTO Resúmenes de la Segunda Reunión de Intercomunicación Académica del Instituto de Ciencias del Mar y Limnología, UNAM. Prospección y evaluación de los peces pelágicos mayores del Golfo de México y Mar Caribe. 1989 26 MAGO, L. F., Bol. Inst. Oceanogr. Univ. Oriente, Osteología Comparada en Ocho Especies Pomadasydae (Pisces: Perciformes) del Golfo de Cariaco, Venezuela y Aguas Adyacentes. 1961 369-473 1 McPHAIL, J. D., and R. L. JONES A Simple Technique for Obtaming Chromosomes from Teleost Fishes. J Fish. Res Board Can., 1966 767-768 23 5 NAKAMURA, I., Proceeding of the International Billfish Symposium Kailua-Kona, Hawaii, Part. 2: 45-61. Some Aspects of the Systematics and Distribution of Billfishes. En: R.S. Shomura y F. Williams (Eds.). Review and Contributed Papers, NOAA Tech. Rep. NMFS (Natl. Mar. Fish. Serv.) Spec. Sci. Rep. 1974 675 9-12 August 1972 _____, FAO Species Catalogue. Billfishes of the World. An annoted and illustrated catalogue of Malins, Sailfishes, Spearfishes and Swordfishes known to date. FAO. Fish. Synop., 1985 65 5 125 NELSON, J. F., Fishes of the world Wiley Interscience, 2a. ed. USA. 1984 419 OHNO, S., U. WOLF, and N. B. ATKIN, Hereditas, Evolution from fish to Mammals by Gene Duplication. 1968 169-187 59 6 POLANCO, J., E. MIMBELA S., L. BELÉNDEZ M., N. A. FLORES, y A. L. REYNOSO A. Situación Actual de las Principales Pesquerías Mexicanas. Secretaría de Pesca. México. Cap. 1.4. Situación Actual de la Pesquería Mexicana de Picudos del Pacífico Oriental, POLANCO, J., E. MIMBELA S., L. BELÉNDEZ M., N. A. FLORES, y A. L. REYNOSO A. 1988 113-167 SCHULTZ, O., Ann. Naturhist. Mus. Wien. Taxonomische Neugrupiening der Überfamilie Xiphioidea (Pisces, Osteychthyes). Serie A. Für. Mineralogie und Petrographie, Geologie und Prähistorie, 1987 95-202 89 SOTA, E. R. de la Ser. Biología. Monografía 3. La Taxonomía y la Revolución en las Ciencias Biológicas. Secretaría Gral. de la Organización de los Estados Americanos. 3a ed Washington, D. C. 1982 90 THOMPSON, K. W., Copeia, Cytotaxonomy of 41 species of Neotropical Cichilidae, 1979 679-691 4 UYENO, T., R. R. MILLER, and J. M. FITZSIMONS, Copeia Karyology of Cyprinodontid fishes of the Mexican family Goodeidae. 1983 497-510 2
|