|
TASA DE FILTRACIÓN Y CULTIVO DE
Moina macrocopa (CRUSTACEA: CLADOCERA) ALIMENTADA CON Scenedesmus incrassatulus
(CHLOROPHYCEAE) Y ESTIÉRCOL VACUNO DIGERIDO
Trabajo recibido el 13
de febrero de 1992 y aceptado para su publicación el 21 de abril de
1992.
FILTRATION RATE AND CULTURE OF Moina macrocopa (CRUSTACEA: CLADOCERA) FED WITH Scenedesmus incrassatulas (CRLOROPHYCEAE) AND DIGESTED COW MANURE
Félix Espinosa-Chávez, Fernando Martínez-Jerónimo y Rodolfo Ramírez-Granados
Laboratorio de Ecología, Escuela Nacional de Ciencias
Biológicas, Instituto Politécnico Nacional, Apartado Postal
32-034, México, D.F. 06030, MÉXICO.
Se determinó el efecto de cantidades diferentes de la microalga
clorofícea Scenedesmus incrassatulus sobre la tasa de
filtración del cladócero dulceacuícola Moina
macrocopa. Así mismo, se evaluó el crecimiento en
biomasa de este crustáceo, alimentado con S.
incrassatulus en dosis inferiores a la que determina el 50% de su
capacidad filtrante, y una cantidad fija de infusión de estiércol
vacuno digerido aeróbicamente, como complemento alimenticio bacteriano.
La tasa de filtración con respecto a la concentración celular
describe una trayectoria curvilínea asintótica, que refleja
límites fisiológicos y mecánicos en el proceso de
filtración a medida que se incrementa el contenido de alimento. El
crecimiento en biomasa (peso en húmedo) de M.
macrocopa se ajusta al modelo logístico; se determinó
que tanto la densidad poblacional máxima alcanzable (k) como el
rendimiento máximo sostenible (rk/4) se incrementan a medida que aumenta
la concentración de microalgas. De M. macrocopa hay
gran potencialidad para cultivarse masivamente de manera controlada, para
suministro de alimento vivo y como organismo de prueba en bioensayos de
toxicología acuática.
The effect of
the concentration of the green microalgae Scenedesmus
incrassatulus on the filtration rate of the freshwater cladoceran
Moina macrocopa was assessed. Filtration rate under
different concentrations of microalgae followed an asymptotic path, denoting a
limit in the maximum filtering rate, imposed by mechanical and physiological
effects. The biomass growth of M. macrocopa fed with
S. incrassatulus concentrations below that determining the
50% of the maximum filtration rate, plus a constant dose of bacterial food
supply from cow manure infusion was also evaluated. Logistic population growth
models were fitted to the biomass data of M. macrocopa. The
maximum attainable biomass (k), and the maximum sustainable yield (rk/4)
increased with the concentration of S. incrassatulus. M.
macrocopa is quite suitable for controlled massive culture for use as
food supply and/or as a test organism for aquatic toxicological
assays.
En el género Moina están comprendidas especies conocidas comúnmente como pulgas de agua, que se distirbuyen en cuerpos de agua permanentes o temporales, ligeramente salinos, localizados en regiones generalmente áridas con fenómenos frecuentes de endorreísmo o arreísmo y pluviosidad variable (Ivleva, 1973; Margalef, 1983). En el Valle de México, Moina macrocopa se distribuye ampliamente en condiciones de eutroficidad, en ambientes con valores de pH de 9.17, y salinidad de hasta 2 g·l–1 (Espinosa-Chávez y Martínez-Jerónimo, inéditos). D'Agostino y Provasoli (1970), al estudiar la alimentación del zooplancton, definieron un medio mineral para el cultivo del cladócero Daphnia magna, empleando como alimento a las clorofíceas Chlamydomonas reinhardii y Scenedesmus obliquus. D' Abramo (1979), simulando la composición de ácidos grasos de distintos grupos de algas, encontró que la mezcla correspondiente a las clorofitas produjo los valores más altos de productividad en M. macrocopa, en comparación con los obtenidos con diatomeas y cianobacterias. Por su parte, Arnold (1971) demuestra que los valores de ingestión, asimilación, sobrevivencia y reproducción en D. pulex son mejores empleando Chlorella vulgaris y Ankistrodesmus falcatus, que utilizando cianobacterias. Conklin y Provasoli (1978) definen tres medios artificiales para el cultivo de M. macrocopa; D'Abramo y Baum (1981) desarrollan un medio artificial para esta misma especie e informan del primer caso de dependencia de colina en un crustáceo. Martínez-Jerónimo y Gutiérrez-Valdivia (1991) emplearon A. convolutus, S. incrassatulus y C. vulgaris como alimento para M. macrocopa, encontrando que todas satisfacen los requerimientos nutricionales de la especie. Proulx et al ., (1984), al demostrar que las bacterias son esenciales para el crecimiento de Daphnia magna, fundamentaron la importancia del componente bacteriano en cladóceros filtradores. Como se aprecia, la información relativa a esta especie es escasa, pues sólo recientemente ha sido objetivo de investigación en condiciones de laboratorio, en virtud de que se le considera importante no sólo como alimento vivo para especies acuáticas mayores (Espinosa, 1987; Villegas, 1990), sino también como organismo de prueba en bioensayos de ecotoxicología (Martínez-Jerónimo y Gutiérrez-Valdivia, 1991). Con el presente ensayo, se busca contribuir al conocimiento de la especie Moina macrocopa desde el punto de vista de su productividad y tasa de ingestión, empleando la microalga Scenedesmus incrassatulus como alimento principal. MATERIAL Y MÉTODOSSe dedujo la relación entre la cantidad de alimento y tasa de ingestión, aplicando básicamente la técnica empleada por Watts y Young (1980), quienes determinaron esta relación para Daphnia magna. De esta manera fue posible delimitar el intervalo de contenido alimentaro en el cual la tasa de filtración está en relación directa con la densidad celular del alga y la productividad en biomasa de Moina macrocopa. El procedimiento fue el siguiente: se separaron grupos de 12 hembras adultas de M. macrocopa cuya longitud total, medida desde el borde anterior de la región cefálica hasta el extremo posterior de las valvas del exoesqueleto, era de 500 a 800 micras; estos grupos se introdujeron al azar y por triplicado en tubos de ensaye con agua potable desclorada conteniendo las concentraciones de S. incrassatulus siguientes: 40,000; 80,000; 230,000; 270,000; 600,000; 1,360,000; 3,000,000 cel.·ml-¹. Estos tubos se mantuvieron en condiciones de oscuridad para evitar que las algas se multiplicaran; se permitía a los crustáceos alimentarse durante seis horas. Cada 60 minutos se les aplicó agitación mecánica durante cinco segundos, para evitar la sedimentación de las células. Al término del experimento se retiraron los cladóceros y se procedió a evaluar las algas remanentes mediante conteo directo en cámara de Neubauer; se determinó la cantidad de células consumidas en este intervalo por diferencia con la original. Para calcular los valores de productividad potencial se determinaron las curvas de crecimiento poblacional del crustáceo mediante el contenido algal siguiente: 404,000; 809,000; 1,618,000 y 3,236,000 cel.·ml-¹; estos guarismos se seleccionaron tomando como base los resultados del experimento precedente. De acuerdo con observaciones realizadas con anterioridad acerca del empleo de estiércol para el cultivo de M. macrocopa (Espinosa, 1986), además de las microalgas, en este experimento se empleó, a manera de complemento alimenticio bacteriano, una dosis constante de infusión de estiércol vacuno digerido aeróbicamente, en cantidad equivalente a 1. 114 g·l-¹ (peso en seco). Los cultivos iniciaron con un inóculo de 0.16 g·l-¹ en biomasa húmeda de M. macrocopa; los experimentos se realizaron por triplicado en frascos de fondo cónico de 3 litros de volumen total, que contenían 2.5 litros de agua potable desclorada. El crecimiento poblacional se registró diariamente como incremento de peso en húmedo, mediante filtración de 100 ml del medio de cultivo en una red de 50 micras; la biomasa se drenó completamente; con una balanza analítica se determinó el peso, en miligramos Posteriormente a esta operación, se regresó a los organismos a los depósitos de cultivo. En todos los casos, el contenido de oxígeno se mantuvo arriba de 4 mg·l-¹ suministro continuo de aire comprimido. La temperatura fue la ambiental, que en promedio tuvo un valor de 27°C. RESULTADOSEn la tabla 1 se muestran los datos cuantitativos de células de S. incrassatulus consumidas en las réplicas de las diferentes concentraciones introducidas. La figura 1 muestra los puntos observados y la curva teórica que describe la tasa de filtración de M. macrocopa como función de la concentración celular del alga, la cual, mediante un proceso iterativo, con el programa de estadística SystatR (versión 3.0) se ajustó a la ecuación siguiente: 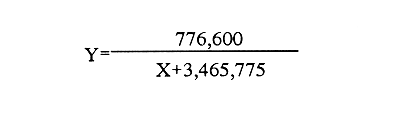 Y: cantidad de células filtradas X: concentración celular de S. incrassatulus En esta ecuación, que es semejante a la de la tasa de saturación de Michaelis-Menten, la constante del numerador representa la asíntota o cantidad máxima filtrable de células por el conjunto de 12 individuos de cada réplica durante seis horas; el coeficiente de correlación para este ajuste fue de r=0.981. 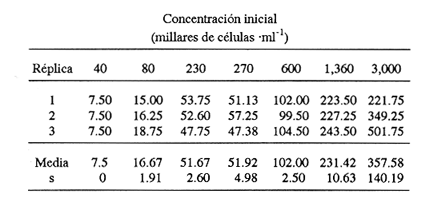 TABLA 1. Millares de células ml1 de Scenedesmus incrassatulus filtradas en un período de seis horas por grupos de doce hembras de Moina macrocopa, a concentraciones diversas del alga. Semuestran los valores promedio y los correspondientes de desviación estándar(s) 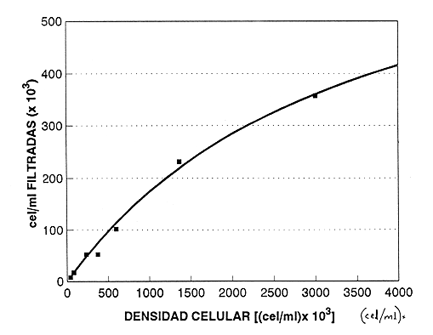 Figura 1. Tasa de filtración de Moina macrocopa según la concentración de Scenedesmus incrassatulus. 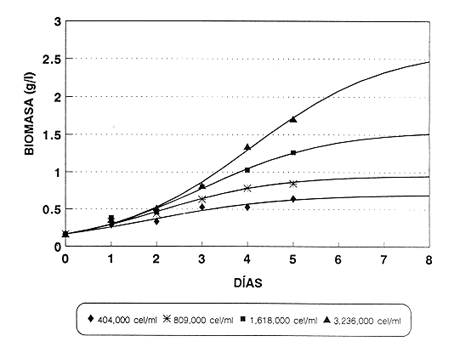 Figura 2. Datos obtenidos y curvas de crecimiento poblacional en biornasa (peso en húmedo) de Moina macrocopa alimentada con concentraciones diferentes de S. incrassatulus y una sola cantidad de estiércol vacuno digerido aeróbicamente. Con respecto al crecimiento poblacional de M. macrocopa, se debe mencionar que la concentración celular máxima empleada (3,236,000 cel. ·ml-¹ ) corresponde a la densidad en que teóricamente este cladócero filtra al 50% de su capacidad máxima calculada, de acuerdo con la ecuación determinada con anterioridad. Sin embargo, se debe considerar que ésta es una extrapolación, suponiendo que la tasa de filtración se ajusta al modelo que describe la curva mostrada en la figura 1, incluso fuera del intervalo en que se hicieron observaciones experimentales; de hecho, no se aplicaron concentraciones mayores debido a que aun en esta dosis máxima ya se observa obstrucción del aparato filtrador. En la figura 2 se muestran los datos promedio observados de biomasa (peso en húmedo) y las curvas teóricas correspondientes que, como se puede apreciar, se ajustan con bastante precisión al modelo de crecimiento logístico descrito por la ecuación siguiente: 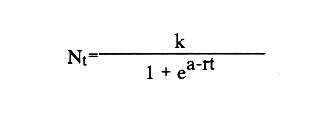 Nt: población al tiempo t k: capacidad de soporte del medio o densidad máxima poblacional alcanzable e: base de los logaritmos naturales a: constante relacionada con el tamaño de la población al tiempo cero r: tasa intrínseca de incremento poblacional t: tiempo Los ajustes de los datos experimentales se hicieron de manera iterativa, por medio del programa de estadística SystatR. En poblaciones cuyo crecimiento se ajusta a esta ecuación, es posible determinar el rendimiento máximo sostenible (RMS), descrito por la relación rk/4, expresado, en el presente caso, en g· 1-¹ día-¹. En la tabla 2 se muestran los valores calculados de productividad y los parámetros de las ecuaciones correspondientes obtenidos para cada dosis de S. incrassatulus aplicada. 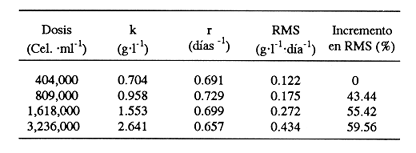 TABLA 2. Valores de productividad en poblaciones experimentales de Moina macrocopa alimentadas conconcentraciones diferentes de Scenedesmus incrassatulus y una sola cantidad de infusión de estiércol vacuno digerido aeróbicamente. Debe señalarse que aun cuando los experimentos no se llevaron hasta la fase de estabilización de las curvas de crecimiento, el ajuste de los datos observados al modelo es bastante bueno, lo que se evidencia en los valores de coeficiente de correlación rectilínea (lineal) y curvilineal determinados, que fluctuaron de -0.9881 a -0.9973, y de 0.9778 a 0.9958, respectivamente; este hecho permite establecer con buen grado de confianza la proyección de las curvas teóricas, como se aprecia en la figura 2. DISCUSIÓNDiferentes experimentos de laboratorio muestran que las especies del zooplancton son capaces de consumir alimento en cantidad mucho mayor que la revelada en estudios de contenido estomacal en condiciones naturales; así, en 24 h una hembra del copépodo Calanus finmarchicus ingiere hasta 373,000 células de la diatomea Skeletonema costatum, o bien 11,000 de Chaetoceros decipiens (Waterman, 1960). Por otra parte, el porcentaje de asimilación del alimento decrece cuando aumenta su disponibilidad. Observaciones de este tipo constituyen la base de la llamada hipótesis de la alimentación superflua, en la que se considera que estas especies consumen el alimento en exceso, lo que involucra cierto gasto energético sin provecho, implicado en la acción de filtrar. Ivleva (1973) señala que para Daphnia pulex la asimilación de Chlamydomonas disminuye entre dos y tres veces cuando la concentración cambia de 25,000 a 100,000 cel. ·ml-¹. Para el caso de adultos de Daphnia ambigua alimentados con la misma alga, Chi-Hsiang y Armitage (1980) informan que el cociente energía consumida/energía asimilada medido en calorías, varió de 4.7 a 57.3%; es decir, existe correlación negativa con la cantidad de alimento. Considerando estos hechos de manera simplista, esta hipótesis parece objetiva; sin embargo, Chi-Hsiang y Armitage (op. cit.) registraron también que con un crecimiento rápido del fitoplancton, Calanus finmarchicus incrementó la producción de huevos, con lo cual se mantiene en grado elevado la eficiencia de asimilación del alimento, y es más coherente con el criterio de aptitud biológica. Por su parte, Waterman (1960) indica que, en hembras maduras de Calanus, los niveles de alimentación se correlacionan de manera directa con la producción de huevos. Cuando se dice que existe alimentación superflua es porque el cociente energía asimilada/energía consumida es bajo, debido a que el tiempo de retención en el intestino es corto. Sin embargo, la cantidad total de energía captada para la reproducción es mucho mayor que cuando el consumo es bajo y el cociente anterior es alto. El comportamiento de M. macrocopa en las pruebas de crecimiento poblacional es consistente con estas observaciones en el sentido de que no existe lo que se podría considerar un fenómeno de sobrealimentación, en virtud de su capacidad para transmutar el exceso de alimento en incrementos significativos de productividad, evaluados en términos de rendimiento máximo sostenible y de crecimiento poblaciones en biomasa. Esto se observa en la figura 2 y en la tabla 2, que muestran el conjunto de curvas logísticas obtenidas y los valores de RMS. Es bastante notorio el incremento de éste al aumentar la dosis de algas; como se puede apreciar en la tabla 2, al incrementar el contenido suministrado de S. incrassatulus, en el RMS ocurren aumentos incluso superiores a 50%. El valor de este rendimiento correspondiente a la dosis mayor aplicada en este trabajo resulta 19.4 veces mayor que el determinado por Martínez-Jerónimo y Gutiérrez-Valdivia (1991). Tales resultados muestran congruencia con la teoría del nivel de pastoreo óptimo, que predice que un animal maximiza su ganancia neta de energía elevando su tasa de ingestión de alimento (Chi-Hsiang y Armitage, 1980). Así, un plan de acción evolutivamente estable sería captar tanta energía para la reproducción como sea posible, como se manifestó por medio de la productividad creciente de M. macrocopa al aumentar la concentración de algas. Como conclusiones importantes de este trabajo, se considera que la elevada capacidad de M. macrocopa en cuanto a su dinámica de ingestión de alimento y su transmutación efectiva en crecimiento poblacional, constituye un rasgo adaptativo al ambiente particularmente cambiante que representan las charcas temporales -su habitat típico-, en el sentido de poder aprovechar rápidamente los máximos de productividad primaria y completar su ciclo de vida, que culmina con la producción de huevos de resistencia o efipios. Por otra parte, también puede hacerse hincapié en lo inadecuado que en ecologia de poblaciones resulta el empleo de los términos alimentación superflua y sobrealimentación. Finalmente, M. macrocopa ha mostrado rendimiento elevado en biomasa y resistencia al manejo, por lo que se considera apropiada para cultivo a escala mayor, ya que, según los valores obtenidos de k, tolera densidades cuantiosas: el máximo potencial calculado es de 2.64 g·1-¹ (peso en húmedo). AgradecimientosLos autores expresan su agradecimiento a los revisores por sus valiosos comentarios, así como a las oficinas de COFAA y DEPI, del Instituto Politécnico Nacional, por los apoyos otorgados para la realización del presente trabajo. LITERATURAARNOLD, E. D., Limnol. Oceanogr. Ingestion, assimilation, survival, and reproduction by Daphnia pulex fed with seven species of blue-green algae. 1971 906-920 CONKLIN, E. D., and L. PROVASOLI, Biol Bull., Biphasic particulate media for the culture of filter-feeders. 1978 47-54 154 CHI-HSIANG, L., and B. K. ARMITAGE, Energy budget of Daphnia ambigua Scourfield. J. Plankton Res., 1980 261-281 2 D'ABRAMO, R. L., Biol. Bull., Dietary fatty acid and temperature effects on the productivity of the cladoceran Moina macrocopa. 1979 234-248 157 D'ABRAMO, R. L., and A. N. BAUM, Biol. Bull. Choline requirement of the microcrustacean Moina macrocopa: A purified diet for continuous culture. 1981 357-365 161 D'AGOSTINO, S. A., and L. PROVASOLI, Biol. Bull. Dixenic culture of Daphnia magna Strauss., 1970 485-494 139 ESPINOSA-CHÁVEZ, F., Memorias del IX Congreso Nacional de Zoología. Villahermosa, Tabasco, Cultivo de Moina macrocopa (Crustacea: Cladocera). México, 1988 207-213 IVLEVA, I. V., Biology and methods. Mass cultivation of invertebrates. Israel Program for Scientific Translations, Jerusalem. 1973 148 MARGALEF, R., Limnología. Ediciones Omega, Barcelona. 1983 1010 p. MARTÍNEZ-JERÓNIMO, F., and A. GUTIÉRREZ-VALDIVIA Hydrobiología, Fecundity, reproduction, and growth of Moina macrocopa fed with different algae. 1991 49-55 222 PROULX, D., R. LESEL, and J. DE LA NOÜE, Rev. Française Sci. de l'Eau Growth of Daphnia magna in axenic, monoxenic and holoxenic conditions. 1984 83-91 3 VILLEGAS, T. C., The effects on growth and survival of feeding water fleas (Moina macrocopa Strauss) and rotifers (Brachionus plicatilis) to milkfish (Chanos chanos Forsskal) fry. Bamidgeh 1990 10-17 42 1 WATERMAN, H. T. The physiology of Crustacea. Academic Press, Inc. New York. 1960 WATTS, E. Y., and S. YOUNG, Components of Daphnia feeding behaviour. J. Plankton Res., 1980 203-212 2
|