|
MOLUSCOS DEL SISTEMA LAGUNAR CHICA-GRANDE, VERACRUZ, MÉXICO:
SISTEMÁTICA Y ECOLOGÍA
Trabajo recibido el 14 de enero de
1992 y aceptado para su publicación el 5 de junio de 1992.
Antonio García-Cubas, Martha Reguero y Rafael Elizarrarás
Laboratorio de Malacología,
Instituto de Ciencias del Mary Limnología, UNAM. Apartado Postal 70-305,
México, D.F. 04510, México. Contribución No. 716 del
Instituto de Ciencias del Mar y Limnología.
Se registra la presencia de 36 especies de moluscos, repartidas
equitativamente en las Clases Gastropoda y Bivalvia, aunque predomina la
primera en número de individuos. En el bentos lagunar se observa un
porcentaje similar de infauna y epifauna, originaria principalmente de ambiente
marino salobre. La mayoría de los gasterópodos identificados en
el área se nutren de partículas orgánicas depositadas en
el sedimento, mientras que los bivalvos filtran las que se encuentran
suspendidas. El sistema es somero, de aguas cálidas oligohalinas y
turbias, con fondos limo-arcillosos y arenosos, en donde abundan las especies
Littoridina sphinctostoma, Cyrenoida floridana, Neritina reclivata,
Mytilopsis leucophaeata, Neritina virginea, Macoma tenta y
Mulinia lateralis. Los valores de diversidad son semejantes
para ambos taxa;aproximadamente la mitad de las especies
concurren en las distintas localidades de muestreo.
The presence of
thirty six species of mollusks, equitably distributed between Gastropoda and
Bivalvia Classes, was recorded. Similar percentages of infauna and epifauna,
originated mainly in marine-brackish environment, were observed in the lagoon
benthos. The majority of gastropods was deposit-feeders, meanwhile bivalves
were filter-feeders. The system is shallow, with warm, oligohaline and turbid
water. The bottom is constituted by mud, clay and sand, where
Littoridina sphitictostoma Cyrenoida floridana, Neritina virginea,
Macoma tenta and Mulinia lateralis are the most
important species because of their relative abundance and distribution.
Diversity values are similar for the two taxa and nearly
half of species are common to all sample locations.
Los cuerpos de agua costeros, como lagunas, albuferas y estuarios, son ecosistemas que constituyen una fuente de recursos bióticos de importancia pesquera, en cuyas aguas -en general salobres - se encuentran representantes de numerososPhyla zoológicos. Sus pobladores pueden ser permanentes, como en el caso de los moluscos; algunos, como el camarón, en ciertas etapas de su ciclo biológico requieren de estos ambientes para su crecimiento; otros (peces y algunos crustáceos) ocupan estos nichos como áreas de refugio transitorio. Por tanto, es de importancia primordial el estudio integral de cada uno de estos cuerpos de agua, con el fin de optimizar su aprovechamiento y detectar alteraciones hidrológicas ocasionadas por el aporte creciente de contaminantes procedentes de ríos y escurrimientos continentales menores, que normalmente drenan en estos sistemas y cuyos efectos sobre la biota resultan impredecibles debido a la carencia de información científica actual. ANTECEDENTESPara el área en estudio no existe ninguna referencia específica, aunque las lagunas costeras se mencionan en estudios generales, como los efectuados por Carranza Edwardset al., (1975), Lankford (1977) y Contreras (1985). DESCRIPCIÓN DEL ÁREA DE ESTUDIOEl sistema lagunar Chica-Grande se encuentra dentro de la llanura costera del Golfo de México, en la región central del litoral del estado de Veracruz. Se ubica entre los paralelos 20° 02' y 20° 06' de latitud septentrional y los meridianos 96° 36' y 96° 42' de longitud occidental (Fig. 1). El complejo lagunar es pequeño; está integrado por dos cuerpos de agua: Laguna Chica y Laguna Grande, comunicados entre sí por un canal estrecho, conocido como "El Caño". La Laguna Chica ocupa la porción noroccidental del sistema lagunar; sus dimensiones aproximadas son 3 km de longitud por 0.8 km deanchura en sus puntos extremos, y cubre una superficie 2 de 2.4 km² . La Laguna Grande tiene alrededor de 4.7 km de longitud por 1.5 km de anchura, con una área cercana a 5 km². La superficie total que abarca el sis tema lagunar es aproximadamente de 8 km². Con el propósito de estudiar la fauna malacológica, así como sus relaciones con los parámetros abióticos, que se considera influyen en su distribución, se establecieron 17 estaciones de muestreo, en torno a las cuales se ubicaron los bancos ostrícolas en producción, aun cuando en este estudio no se cuantificó la fauna que los integra (Fig. 2). 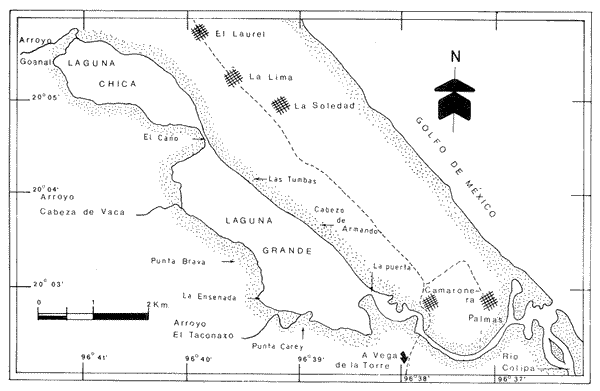 Figura 1. Ubicación y tiponimia del sistema lagunar Chica-Grande. 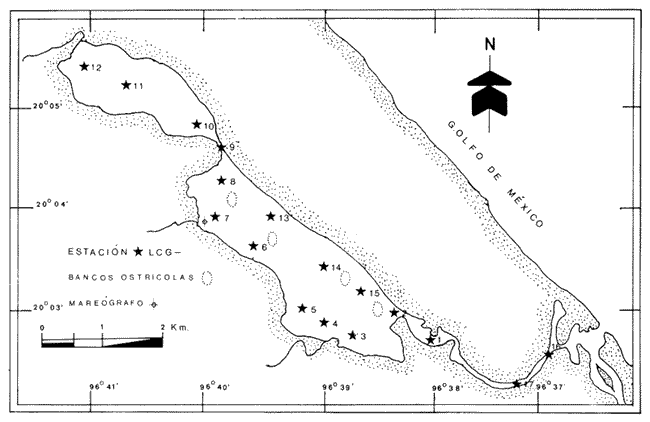 Figura 2. Distribución de las estaciones de muestreo y ubicación de bancos ostrícolas. CLIMATOLOGÍAEl clima de la zona geográfica donde se localizan las lagunas, de acuerdo con la clasificación de Köppen, modificada por García (1971) para la República Mexicana, es del tipo Aw1, (x') (e), el cual corresponde a un clima cálido subhúmedo con lluvias en verano, con dos épocas de secas, una marcada en invierno y otra más corta durante el verano, con lluvias poco frecuentes pero intensas durante todos los meses. El régimen climático, por la oscilación térmica, es extremoso, manifiesta un promedio de temperatura máxima de 27.5ºC a principios de verano y una mínima de 19.5ºC a mediados del invierno, con precipitaciones pluviales que alcanzan promedios de 22 mm como mínimo a 180 mm como máximo; los meses de enero-febrero son los de menor precipitación y julio-agosto los de máxima (García, 1971). Los datos de precipitación y temperatura que se emplearon para la caracterización del tipo de clima corresponden a los obtenidos en la estación climatológica "Vega de la Torre", Veracruz, localizada al sursureste del sistema lagunar, a 20° 01' de latitud norte y 96° 38' de longitud oeste. HIDROGRAFÍAAl sureste de la Laguna Grande desemboca el Río Colipa; integrados al sistema se encuentran tres arroyos: "El Goanal", que se conecta a la Laguna Chica en su extremo noroccidental, y los denominados "Cabeza de Vaca" y "El Taconazo", que vierten su contenido en la Laguna Grande en sus porciones noroeste y suroeste, respectivamente. HIDROLOGÍAEl sistema recibe el aporte de aguas marinas mediante una pequeña boca de aproximadamente 300 m de anchura, que comunica al sistema por un canal localizado en el extremo sureste de la Laguna Grande; mientras que el aporte de agua dulce es proporcionado por escurrimientos continentales, así como por la afluencia de los tres arroyos que desembocan en el sistema y en menor proporción por el Río Colipa, cuya influencia hacia el interior del sistema está limitada por la estrechez del canal de comunicación BATIMETRÍASe trata de un sistema somero, cuya profundidad media es de 0.70 m, con registros de más de 1 metro en el canal "El Caño", que comunica a las dos lagunas. TRANSPARENCIALos dos cuerpos de agua presentan turbiedad elevada, con transparencia promedio de 0.10 m. TEMPERATURA DEL AGUAEs un sistema de aguas cálidas; durante julio pueden registrarse valores medios de 32º C. SALINIDADDurante el verano, el sistema es predominantemente oligohalino (0.5-5.0 ‰), excepto en las áreas del canal próximas a la comunicación con el mar, en donde se comporta como polihalino (18-30‰). SEDIMENTOSEn el área abundan limos y arcillas, con cantidades variables de arena. VEGETACIÓNLos terrenos que circundan al sistema lagunar forman parte de la planicie costera del Golfo de México; en gran parte están cubiertos por Rhizophora mangle o mangle rojo y Avicenia germinans o mangle negro. MATERIAL Y MÉTODOSLa recolección del material biológico y el registro de los datos ambientales se llevaron a cabo durante una campaña realizada en la zona de estudio durante julio de 1980 por personal del Laboratorio de Malacología del Instituto de Ciencias del Mar y Limnología, de la UNAM. Se hizo un reconocimiento del área para excluir zonas con presencia de bancos de ostión y almeja y se estableció una red de 17 estaciones distribuidas en el sistema lagunar. En cada una de ellas se hicieron mediciones de- temperatura y salinidad, con un salinómetro de inducción Beckman; transparencia, con un disco de Secchi, y de profundidad con una sondaleza de mano. En cada estación se extrajo un' volumen constante de 5 litros de sedimento superficial húmedo, por medio de una dragavan Veen, con capacidad de 2 litros. Los sedimentos obtenidos se tamizaron in situ, con una malla de plástico, con abertura de 1.5 mm. Las muestras obtenidas se preservaron en bolsas de polietileno con formaldehído al 10%, neutralizado con borato de sodio y, debidamente etiquetados, se trasladaron al Laboratorio de Malacología, en donde se lavaron con agua corriente y se secaron a temperaturas inferiores a 70ºC. A los especímenes recolectados vivos se les separó de los exoesqueletos para proceder a su identificación y cuantificación a nivel específico. En el caso de los bivalvos con conchas desarticuladas se adoptó el criterio de García-Cubas (1963), quien considera cada valva como un organismo. Los ejemplares representativos se incorporaron a la colección malacológica del Instituto de Ciencias del Mar y Limnología, de la UNAM. Las comunidades de moluscos se caracterizaron mediante los componentes de abundancia, frecuencia y densidad de las especies (individuos/m²) , de donde se derivó el valor de importancia relativo a cada una de ellas y el índice de similitud entre las estaciones de muestreo del sistema (Krebs, 1972). Lo anterior se complementó con indicaciones de distribución geográfica y local, llevando a cabo una revisión biblio gráfica que permitiera determinar también, para cada una de las familias de moluscos identificados, el tipo de nutrición y la relación con el sustrato. 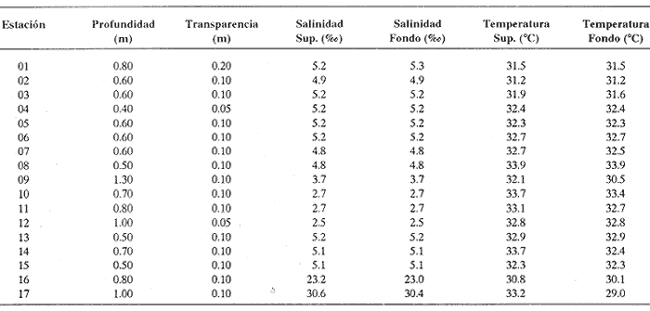 TABLA 1. Parámetros hidrológicos en las estaciones de muestreo Así mismo, se calcularon los índices de diversidad (H'n), diversidad máxima (Hmáx), equitatividad (J) y riqueza de especies (D) para cada una de las clases de moluscos, por estación, por ambiente halino y para el sistema en conjunto, mediante las expresiones matemáticas propuestas por Shannon-Wiener (1963), Pielou (1975) y Margalef (1958), respectivamente. El cálculo de estos índices se efectuó tanto para la comunidad total como para las poblaciones vivas. El análisis incluyó también la cuantificación de los coeficientes de determinación entre los valores de H'n vs. J y H'n vs. D, para establecer el factor dominante sobre la diversidad en cada taxón, trabajando por separado la comunidad viva y la total. RESULTADOS
HIDROLOGÍAEn el complejo Chica-Grande, la temperatura fluctuó entre 29 y 33.9ºC, la salinidad entre 2.5 y 30.°/oo y la profundidad entre 0.40 y 1.30 m. En la tabla 1 se concentran los datos detallados de estos parámetros para cada una de las estaciones de muestreo. CONSIDERACIONES A LA SISTEMÁTICAA continuación se presenta la sistemática del total de epecies encontradas en el sistema lagunar en estudio, cuya identificación se basó en caracteres conquiliológicos. El arreglo de la malacofauna se efectuó de acuerdo con el sistema de clasificación propuesto por Keen (1971), y las indicaciones acerca de distribución geográfica, habitat y tipo de nutrición de las especies se obtuvieron de obras de Purchon (1968), Andrews (1971), Abbott (1974), García Cubas (1981), Antolí y García-Cubas (1985), Reguero y García-Cubas (1989, 1991), García-Cubas et al., (1990), García-Cubas y Reguero (1990). Los organismos quedaron ubicados sistemátícamente en las Clases Gastropoda y Bivalvia. En la primera se reconocieron 4 órdenes, 11 familias, 14 géneros y 18 especies; mientras que en la Clase Bivalvia se identificaron 3 órdenes, 11 familias, 15 géneros y 18 especies. Por tanto, el registro integral lo conforman 2 clases, 7 órdenes, 22 familias, 29 géneros y 36 especies. CLASE GASTROPODASubclase Prosobranchia Orden Archaeogastropoda Superfamilia Neritacea Familia Neritidae Género Neritina Lamarck, 1816 1. Neritina reclivata(Say, 1822) Distribución local: estaciones 1, 2, 3, 4, 5, 9 y 15. Distribución geográfica: Florida (EE.UU. de A.), Golfo de México a Mar de las Antillas. Habitat y tipo de nutrición: es común en aguas salobres a dulces; frecuentemente se le encuentra sobre la vegetación sumergida, debido a sus hábitos nutritivos micrófagos.  Subgénero Vitta Mörch, 1852 2. Neritina (Vitta) virginea (Linnaeus, 1758) Distribución local: estaciones 1, 2, 3, 4, 5, 6, 7, 9, 14 y 17. Distribución geográfica: Florida, Golfo de México, las Antillas; de Islas Bermudas a Brasil. Habitat y tipo de nutrición: especie epifaunal entre la vegetación. Muy frecuente en zonas de intermareas y aguas salobres; nutrición micrófaga raspando la vegetación. 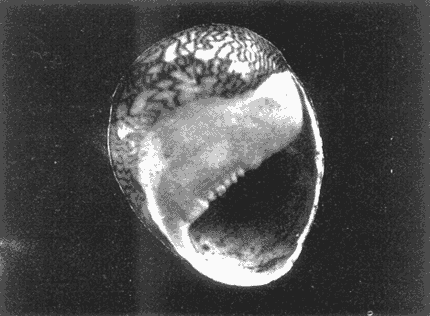 Orden Mesogastropoda Superfamilia Rissoacea Familia Rissoidae Subfamilia Cingulinidae Género Cingula Fleming, 1828 SubgéneroMicrodochus Rehder, 1943 3. Cingula (Microdochus) floridatia (Rehder, 1943) Distribución local: estación 1. Distribución geográfica: Florida, Golfo de México, parte de las Antillas; Puerto Rico. Habitat y tipo de nutrición: especie epifaunal, desde la zona de mareas hasta 50 m de profundidad. Nutrición micrófaga. 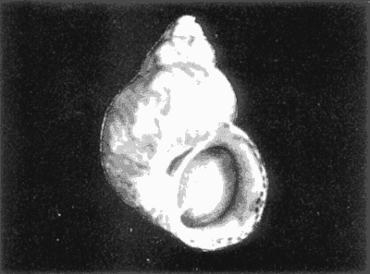 Familia Rissoinidae Subfamilia Rissoininae GéneroRissoina Orbigny, 1840 Subgénero Schwattziella Nevil, 1884 4. Rissoina (Schwartziella.) catesbyana Orbigny, 1842 Distribución local: estación 11. Distribución geográfica: Islas Bermudas a Florida (EE.UU. de A.); costas del Golfo de México; Mar Caribe hasta Brasil. Habitat y tipo de nutrición: se encuentra comúnmente en bahías y lagunas salobres y marinas; prosperan en praderas de Thalassia, sobre fondos arenosos; epifaunal micrófaga de materia orgánica depositada.  Familia Hydrobiidae Género: Hydrobia Hartman, 1821 5.Hydrobia totteni Morrison, 1954 Distribución local: estación 17. Distribución geográfica: Península de Labrador (Canadá) al Golfo de México. Habitat y tipo de nutrición: especie epifaunal, abun dante en aguas someras y estancadas. Microfagia de algas verdes. 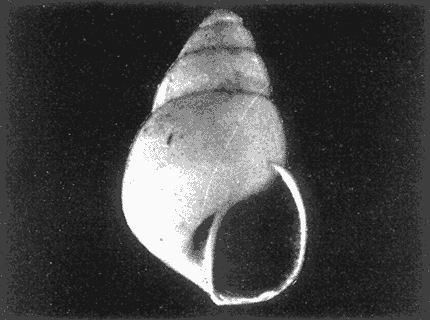 Género Littoridina Eydoux y Souleyet, 1852 SubgéneroTexadina Abbott y Ladd, 1951 6. Littoridina (Texaditia) sphinctostomaAbbott y Ladd, 1951 Distribución local: estaciones 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 131 14, 15 y 17. Distribución geográfica: de Mississippi a Texas (EE.UU. de A.) y Golfo de México. Habitat y tipo de nutrición: es muy común en aguas someras y salobres hasta casi dulces; viven en fondos arenolimosos, epifaunales; se nutren de materia orgánica depositada en el sustrato. 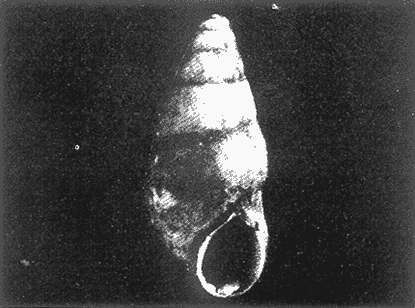 Familia Vitrinellidae Subfamilia Vitrinellinae Género Vitrinella C. B. Adams, 1850 SubgéneroVitrinella s. s. 7. Vitrinella (Vitrinella) floridana Plisbry y McGinty, 1946. Distribución local: estación 4. Distribución geográfica: desde Florida hasta el sur del Golfo de México (Laguna de Términos, Campeche, México). Habitat y tipo de nutrición: aguas someras; epifaunal entre rocas; micrófaga de materia orgánica depositada. 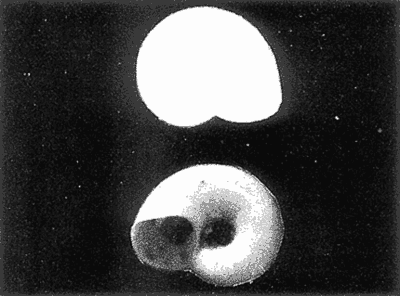 Subfamilia Teinostomatinae GéneroTeinostoma H. y A. Adams, 1854 8. Teinostoma biscaynense Pilsbry y McGinty, 1945 Distribución local: estación 16. Distribución geográfica: sur de Florida y costas del Golfo de México. Habitat y tipo de nutrición: se encuentra en bahías y lagunas costeras; epifaunal en fondos de arena; micrófaga de materia orgánica depositada. 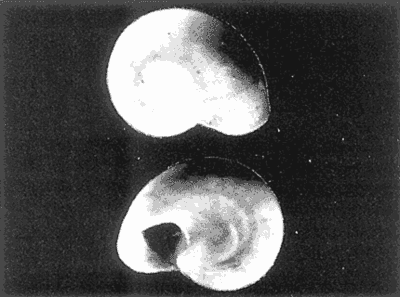 Superfamilia Turritellacea Familia Caecidae Género CaecumFleming, 1813 SubgéneroCaecum s. s. 9. Caecum (Caecum) pulchellum Stimpson, 1851 Distribución local: estaciones 1, 11 y 17. Distribución geográfica: Carolina del Norte, Golfo de México, Islas Bermudas hasta Brasil. Habitat y tipo de nutrición: es abundante en praderas de Thalassia y pastos marinos, en bahías y lagunas costeras; epifaunal sobre fondos de arena. Micrófaga de materia orgánica depositada. 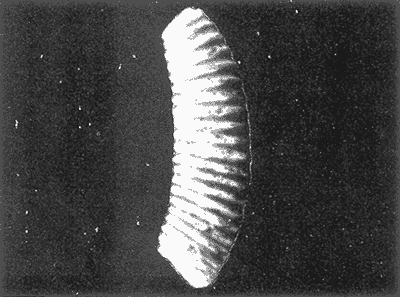 Subgénero Brochina Gray, 1857 10. Caecum (Brochina) vestitum De Fofin, 1870 Distribución local: estaciones 11, 16 y 17. Distribución geográfica: Cayos de Florida; sur del Golfo de México; Mar de las Antillas. Habitat y tipo de nutrición: epifaunales sobre arena o fragmentos de conchas. Se nutren de materia orgánica depositada en el fondo. 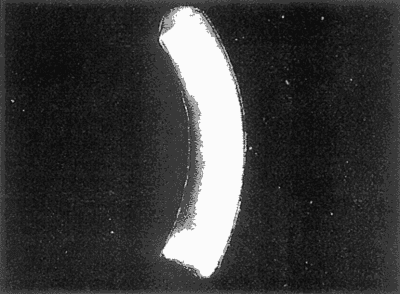 Superfamilia Cerithiacea Familia Cerithiidae Subfamilia Diastominae GéneroDiastoma Deshayes, 1850 11. Diastoma varium (Pfeiffer, 1840) Distribución local: estaciones 1 y 10. Distribución geográfica: desde Maryland (EE.UU. de A.), costas del Golfo de México hasta Brasil. Habitat y tipo de nutrición: epifaunal entre la vegetación sumergida en bahías y lagunas costeras. Hervivoros 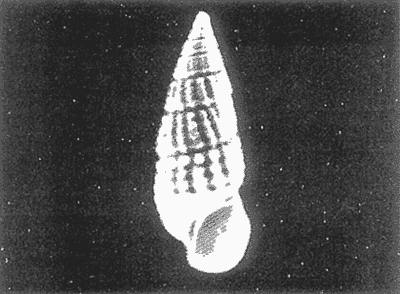 Familia Potamididae GéneroCerithidea Swainson, 1840 Subgénero Cerithideopsis Thiele, 1929 12. Cerithidea (Cerithideopsis) pliculosa (Menke, 1829) Distribución local: estaciones 1, 2, 3, 4, 5, 6, 11, 14 y 15. Distribución geográfica: desde Louisiana (EE.UU. de A.), Golfo de México a América Central. Habitat y tipo de nutrición: se encuentran en bahías y lagunas costeras donde predominan pastos marinos; seminfaunales en fondos fangosos; se nutren de materia orgánica depositada en el fondo. 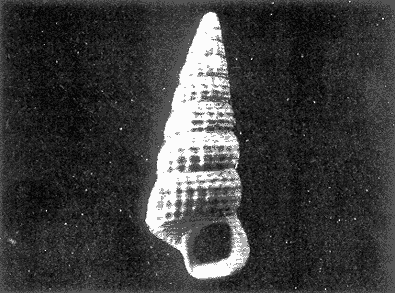 Superfamilia Volutacea Familia Olividae Subfamilia Olivellinae GéneroOlivella Swainson, 1831 Subgénero Niteoliva Olsson, 1956 13. Olivella (Niteoliva) ininuta (Link, 1807) Distribución local: estaciones 16 y 17. Distribución geográfica: Golfo de México; Caribe; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie infaunal en fondos arenosos, en aguas someras. Alimentación carnívora y carroñera. 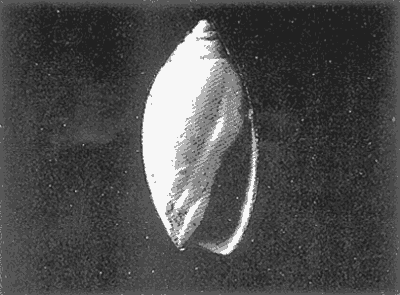 Subclase Opisthobranchia Orden Entomotaeniata Superfamilia Pyramidellacea Familia Pyramidellidae Subfamilia Odostomiinae Género Odostomia Fleming 1813 Subgénero Odostomias.s. 14. Odostomia (Odostomia) laevigata(d'Orbigny, 1842) Distribución local: estaciones 14 y 17. Distribución geográfica: Carolina del Norte a Florida, Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie epifaunal, en aguas marinas y salobres someras. Ectoparásita. 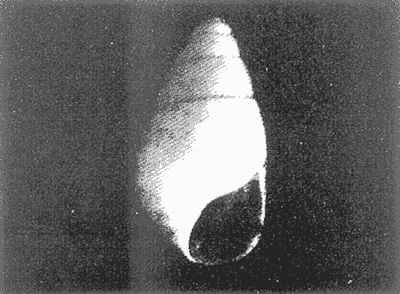 Subgénero Menestho Möller, 1842 15. Odostomia (Menestho) bisuturalis (Say, 1822) Distribución local: estaciones 1, 3, 13, 14 y 15. Distribución geográfica: Golfo de San Lorenzo (Canadá) a Delaware; costas del Golfo de México. Habitat y tipo de nutrición: común por debajo de la línea de la marea baja, adherida a las rocas, epifaunal. Se alimenta de ostiones y gasterópodos herbívoros. 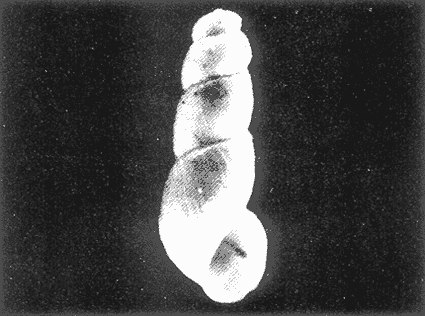 16. Odostomia(Menestho) impressa (Say, 1822) Distribución local: estaciones 1, 2, 3, 4, 5, 6, 8, 9, 11, 16 y 17. Distribución geográfica: Massachusetts a Florida y Golfo de México. Habitat y tipo de nutrición: epifaunal, de aguas salobres, especialmente en bancos ostrícolas, como ectoparásito. 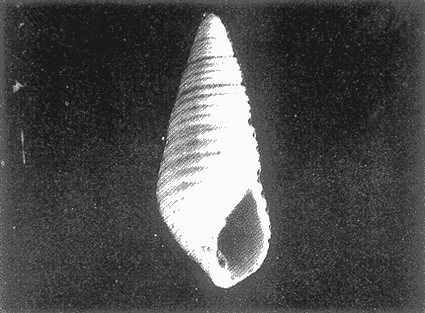 Subfamilia Turbonillinae GéneroTurbonilla Risso, 1826 17. Turbonilla abrupta Bush, 1899 Distribución local: estaciones 1, 2, 3, 15 y 16. Distribución geográfica: Golfo de San Lorenzo (Canadá) al Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie epifaunal en aguas someras marinas y salobres. Ectoparásita. 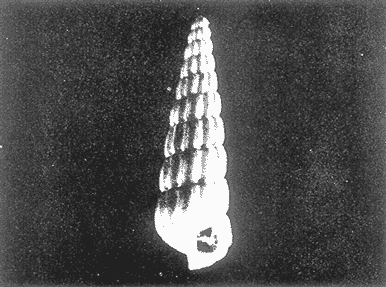 Orden Cephalaspidea Superfamilia Bullacea Familia Acteocinidae GéneroActeocina Gray, 1847 18. Acteocina canaliculata (Say, 1822) Distribución local: estaciones 1, 2, 3, 4, 11, 12, 15, 16 y 17. Distribución geográfica: Nueva Escocia (Canadá) a Florida, Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie epifaunal en bahías y lagunas costeras. Alimentación carnívora. 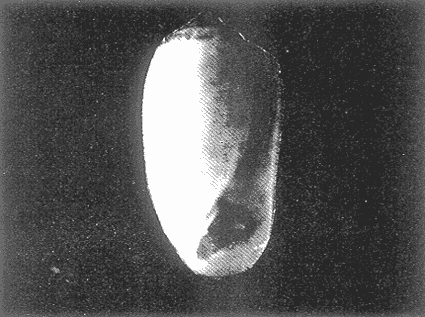 CLASE BIVALVIASubclase Pteriomorphia Orden Arcoida Superfamilia Arcacea Familia Arcidae Subfamilia Anadarinae Género Anadara Gray, 1847 Subgénero Cunearca Dall, 1898 19. Anadara (Cunearca) chemnitzii (Philippi, 1851) Distribución local: estación 16. Distribución geográfica: Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie poco común, epifaunal en fondos arenosos y aguas someras. Alimentación de partículas orgánicas en suspensión. 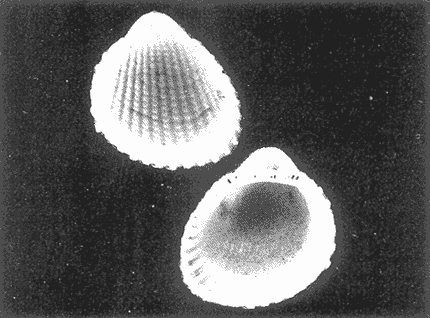 Subgénero Larkinia Reinhart, 1935 20. Anadara (Larkinia) transversa (Say, 1822) Distribución local: estación 15. Distribución geográfica: desde el sur de Cabo Cod, Massachusetts, Golfo de México a la Laguna de Términos, Campeche (México) hasta las Antillas. Habitat y tipo de nutrición: de la zona litoral hasta 11 m de profundidad; forma parte de la infauna; hábitos alimentarios filtrantes de partículas en suspensión.  Orden Mytiloida Superfamilia Mytilacea Familia Matilde Subfamilia Mytilinae Género Género BrachidontesSwainson, 1840 SubgéneroHormomya Mörch, 1835 21. Brachidontes (Hormomya) exustus (Linnaeus, 1758) Distribución local: estaciones 1, 2, 3, 5, 9, 14 y 15. Distribución geográfica: Carolina del Norte al Golfo de México; las Antillas hasta Uruguay. Habitat y tipo de nutrición: epifaunal sobre rocas y bancos de ostión, fijos mediante un biso; muy frecuente en aguas someras salobres. Nutrición suspensivora. 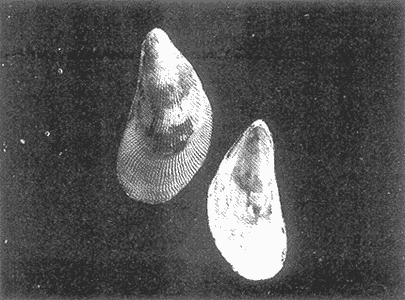 Género Ischadiuni Jukes-Brown, 1905 22. Ischadium recurvum(Rafinesque, 1820) Distribución local: estaciones 1, 2, 3, 5, 7, 9, 14 y 15. Distribución geográfica: Cabo Cod, Massachusetts, Golfo de México; las Antillas. Habitat y tipo de nutrición: aguas salobres y someras; forma parte de la epifauna; mediante un biso fibroso se adhieren a rocas y bancos de ostras; su nutrición es por filtración. 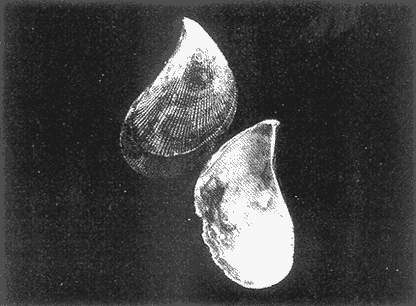 Subclase Heterodonta Orden Veneroida Superfamilia Dreisseniacea Familia Dreissenidae Género Mytilopsis Conrad, 1858 23. Mytilopsis leucophaeata (Conrad, 1831) Distribución local: estaciones 1, 2, 3, 4, 5, 9, 15 y 17. Distribución geográfica: Nueva York hasta Florida; Texas; costas del Golfo de México. Habitat y tipo de nutrición: especie epifaunal fija sobre rocas; de aguas dulces a salobres. Nutrición de partículas en suspensión. 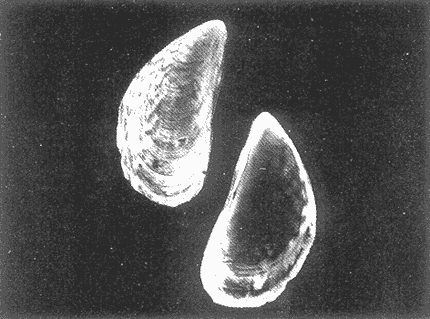 Superfamilia Cyrenoidacea Familia Cyrenoididae Género CyrenoidaJoannis, 1835 24. Cyreizoida floridana Dall, 1889 Distribución local: estaciones 1, 2, 3, 4, 5, 6, 7, 8, 9 y 14. Distribución geográfica: desde Georgia (EE.UU de A.) hasta el Golfo de México. Habitat y tipo de nutrición: comunes en aguas salobres a dulces; forman parte de la infauna superficial; filtradores suspensivoros.  Superfamilia Lucinacea Familia Lucinidae GéneroLucina Bruguiére, 1797 Subgénero Phacoides Blainville, 1825 25. Lucina (Phacoides) pectinata (Gmelin, 1791) Distribución local: estación 2. Distribución geográfica: Carolina del Norte a Florida; Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: forma parte de la infauna superficial en aguas someras de lagunas y estuarios. Se nutre de partículas orgánicas en suspensión.  Superfamilia Veneracea Familia Veneridae Subfamilia Chioninae Género ChioneMühlfeld, 1811 SubgéneroChione s.s. Chione (Chione) cancellata (Linaceus, 1767) Distribución local: estaciones 1, 10, 16 Distribución geográfica: de Carolina del Norte a Florida; Golfo de México; las Antillas hasta Brasil. Habitat y tipo de nutrición: especie infaunal en sustratos arenosos de aguas someras. Nutrición suspensívora.  Subgénero Lirophora Conrad, 1863 27. Chione (Lirophora) elenchi Pulley, 1952 Distribución local: estación 1. Distribución geográfica: Golfo de México. Habitat y tipo de nutricion: especie infaunal de sustratos arenosos y aguas marinas someras.Nutrición suspensívora.  Género Anomalocardia Schumacher, 1817 28. Anomalocardia auberiana (d'Orbigny, 1842) Distribución local: estación 2. Distribución geográfica: sur de Florida, Texas; Golfo de México; Centroamérica; las Antillas; Brasil hasta Uruguay. Habitat y tipo de nutrición: es común en aguas salobres; forma parte de la infauna en fondos de arena y fango. Se localiza en bahías abiertas, típicamente marinas, así como en lagunas y esteros; tolera amplias variaciones de salinidad. Su nutrición es suspensivora.  Superfamilia Mactracea Familia Mactridae Subfamilia Mactrinae Género MuliniaGray, 1837 29. Mulinia lateralis (Say, 1822) Distribución local: estaciones 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 13, 14, 15, 16 y 17. Distribución geográfica: desde Maine hasta el Golfo de México. Habitat y tipo de nutrición: especie infaunal de sustratos arenosos y fangosos, en aguas someras templadas y cálidas. Nutrición suspensívora.  GéneroRangia Desmoulis, 1832 Subgénero Rangianella Conrad, 1863 30. Rangia (Rangianella) flexuosa(Conrad, 1839) Distribución local: estaciones 1, 2, 3, 4, 5, 9, 10, 11 y 14. Distribución geográfica: desde Louisiana hasta el sur del Golfo de México. Habitat y tipo de nutrición: especie infaunalen sustratos blandos, en aguas salobres influidas por ríos. Nutrición de partículas orgánicas en suspención.  Superfamilia Tellininae Familia Tellinidae Género Tellina Linnaeus, 1758 Subgénero AngulusMegerle Von Mühlfeld, 1811 31.Tellina (Angulus) texana Dall, 1900 Distribución local: estación 7. Distribución geográfica: Costas del Golfo de México a Bahamas. Habitat y tipo de nutrición: en lagunas costeras y bahías, en bancos de arena, infaunal de nutrición filtrante.  Subfamilia Macominae Género Macona Leach, 1819 32. Macoma tenta(Say, 1834) Distribución local: estciones 8, 12, 13, 14, 15 y 17. Distribución geográfica: Cabo Cod a Florida; Bermudas; Golfo de México hasta Brasil. Habitat y tipo de nutrición: especie infaunal de aguas someras. Se nutre de partículas depositadas en el sustrto.  Subgénero Rexithaeruas Tryon, 1869 33. Macoma (Rexithaeruas) mitchelli Dall, 1895 Distribución local: estaciones 1 y 3. Distribución geográfica: de Carolina del Sur (EE.UU de A.) hasta el sur del Golfo de México. Habitat y tipo de nutrición: frecuente en lagunas y esteros con influencia de ríos, forma parte de la infauna. Nutrición de materia orgánica depositada en el fondo.  Familia Donacidae Género Donax Linnaeus, 1758 34. Donax variabilis Philippi, 1847 Distribución local: estaciones 16 y 17. Distribución geográfica: desde el norte de Texas hasta el sur del Golfo de México. Habitat y tipo de nutrición: especie infaunal en sustratos arenosos. Nutrición filtrante de partículas en suspensión.  Familia Solecurtidae Género GéneroTagelusGray, 1847 SubgéneroMesopleura Conrad, 1868 35. Tagelus (Mesopleura) plebeius(Lightfoot, 1786) Distribución local: estaciones 5 y 16. Distribución geográfica: Cabo Cod a Florida; Golfo de México; Brasil; Argentina. Habitat y tipo de nutrición: especie infaunal de aguas someras a moderadamente profundas. Se nutre de partículas depositadas en el sustrato.  Familia Semelidae Subfamilia Scrobiculariinae GéneroAbra Lamarck, 1818 36.Abra aequalis (Say, 1822) Distribución local: estación 1. Distribución geográfica: Carolina del Norte a Florida; Texas; Golfo de México; las Antillas; Brasil. Habitat y tipo de nutrición: bahías abiertas, áreas centrales de lagunas o próximas a la costa. Viven enterradas en sedimentos limosos y arenosos, como infauna superficial; nutrición suspensivora.  ANÁLISIS CUANTITATIVO Y PORCENTUALEn el complejo lagunar se fijó una red de 17 estaciones, en las que se capturaron 4,363 ejemplares; se cuantificaron 506 (11.6%) especímenes vivos y 3,857 (88.4%) remanentes esqueléticos. De esta área se identificaron 36 especies: 18 pertenecientes a la Clase Gastropoda y una cantidad igual a la Clase Bivalvia. Para el análisis cuantitativo se consideraron todas, debido a que no se realizaron arrastres, por ser una técnica de extracción que se utiliza preferentemente en estudios cualitativos. Respecto a su abundancia (Tabla 2), el 61.6% del total de individuos recolectados pertenecen a la Clase Gastropoda (2,688 ejemplares) y el 38.4% a la Clase Bivalvia (1,675 ejemplares). Las estaciones en las que se capturaron más individuos fueron la 3 y la 1, con 1,031 y 932 ejemplares, respectivamente; mientras que en las estaciones 7 y 13 se registraron los valores -más bajos, con 19 y 24 individuos, en esa secuencia. Por lo que se refiere a estaciones con la cantidad más elevada de especies, éstas fueron las 1, 2 y 3 (con 21 especies en la primera y 15 en las dos últimas); las que presentaron menor número fueron las 12 y 13, con cuatro especies por estación. En este sistema lagunar, 14 (38.9%) de las 36 especies que se analizaron cuantitativamente tuvieron representantes vivos: nueve incluidas en la Clase Gastropoda, con 449 individuos y un valor porcentual de 88.7 respecto al total de ejemplares vivos, y cinco especies de la Clase Bivalvia, con 57 organismos, que representaron el 11.3%. Las estaciones en las que se capturó un mayor número de individuos vivos fueron las 5 y 2 (Tabla 3). ESPECIES CARACTERÍSTICAS DEL ÀREA DE ESTUDIOEn los dos cuerpos de agua se reconocieron áreas en donde las especies sujetas a aprovechamiento comercial constituyen agregados en diversas etapas de producción. Así, en la figura 2 se observa la ubicación de bancos ostrícolas en el sistema Chica-Grande. Sin embargo, es importante hacer notar que no se extrajeron muestras de bancos. De manera que, al excluir estos agregados, el análisis de abundancia, frecuencia y densidad (Tablas 4 y 5) indica que las especies con mayor importancia relativa en el área son las siguientes: COMUNIDAD MALACOLÒGICA TOTAL:Littotidina sphinctostoma Cyrenoida floridana Neritina reclivata Mytilopsis leucophaeata Neritina virginea Mulinia lateralis con un porcentaje global correspondiente al 84.7%, en donde aproximadamente la mitad de los ejemplares capturados pertenecen a las dos primeras especies. COMUNIDAD MALACOLÒGICA VIVA:Cyrenoida floridana Neritina reclivata Macoma tenta Mulinia lateralis las cuales destacaron en la época de muestreo por constituir el 90.7% de especimenes capturados vivos; se hace notar que Cyrenoida floridana concentró al 70.9 %. ÍNDICES DE SIMILITUDSe evaluó la semejanza entre los componentes de las muestras o estaciones de cada sistema, mediante la aplicación porcentual del índice de similitud de Krebs (1972), el cual permite establecer una relación de distribución y asociaciones de las especies que integran la comunidad de moluscos. Con los valores obtenidos de comparar la presencia de especies comunes en las estaciones de muestreo se elaboraron diagramas de enrejado referentes a las comunidades malacológicas total y viva, se confrontó la afinidad relativa a las Clases Gastropoda y Bivalvia, así como la delplylum en conjunto. Las figuras 3 y 4 muestran gráficamente grupos de estaciones con porcentajes de similitud comprendidos en los intervalos de 0 a 24, 25 a 49, 50 a 74 y 75 a 100, así como el porcentaje que representan con respecto al número total de combinaciones posibles. 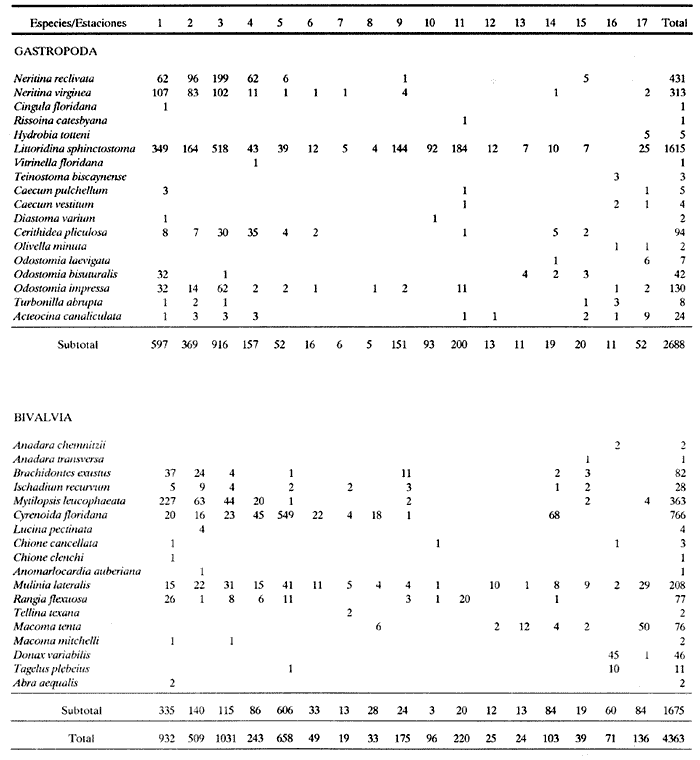 TABLA 2. Abundancia de la comunidad malacológica total  TABLA 3. Abundancia de la comunidad malacológica viva COMUNIDAD MALACOLÒGICA TOTALConsiderando al total de gasterópodos recolectados (Fig. 3A), la afinidad resultante indica que de las 136 combinaciones posibles, dadas por la cantidad de estaciones, 41.2% se ubica dentro del rango 25-49, con 56 combinaciones; 36% corresponde al intervalo 50-74, con 49; a los rangos de 0-24 y 75-100 corresponden porcentajes de 12.5% y 10.3%, con 17 y 14 combinaciones, respectivamente. La similitud promedio para todo el conjunto de estaciones es de 46.2%. Con base en las especies identificadas de bivalvos, la semejanza calculada para las estaciones de muestreo (Fig. 3B) indica que 37.5% ocupa el intervalo 25-49, con 51 combinaciones, de un total de 136; 30.9%, con 42 combinaciones, se encuentra en el rango de 50-74; 19.1% en el intervalo de 0-24, con 26 combinaciones, y 12.5%, con 17 combinaciones, está dentro del rango 75-100. El valor medio de afinidad para este taxón es de 44.2%. Para el total de moluscos, los valores porcentuales de similitud entre pares de estaciones, obtenidos a partir de 136 combinaciones posibles (Fig. 3C), -muestran que 52.2% se ubica dentro del rango 25-49, con 71 combinaciones; 28.7% en el intervalo de 50-74, con 39 combinaciones, y porcentajes de 9.6 para los intervalos 0-24 y 75-100, con 13 combinaciones cada uno, En este caso la semejanza media es de 45.7%. COMUNIDAD MALACOLÒGICA VIVALa similitud calculada para las estaciones de muestreo, con base en las especies de la Clase Gastropoda de las que se obtuvieron representantes vivos (Fig. 4A), indica que 40% ocupan los intervalos de 0-24 y 50-74, con cuatro combinaciones cada uno, respecto a las 10 posibles, y porcentajes de 10% se ubican en los intervalos 25-49 y 75-100, con una combinación en cada caso. El valor medio de afinidad correspondiente a este grupo es de 37.3%. 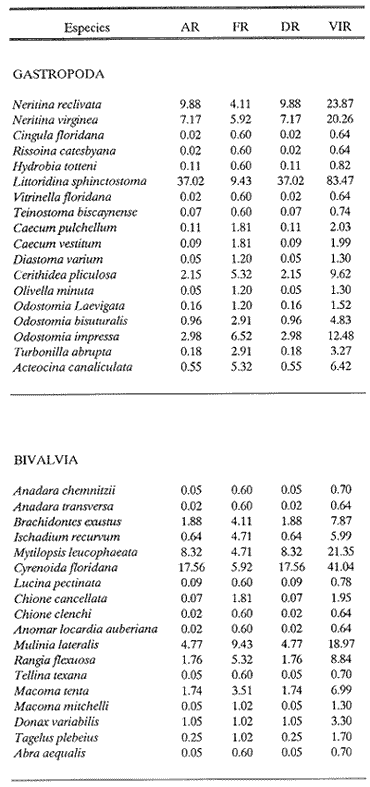 TABLA 4 Abundancia relativa (AR), frecuencia relativa (FR), densidad relativa (DR) y valor de importancia relativa (VIR) de la comunidad malacólogica total 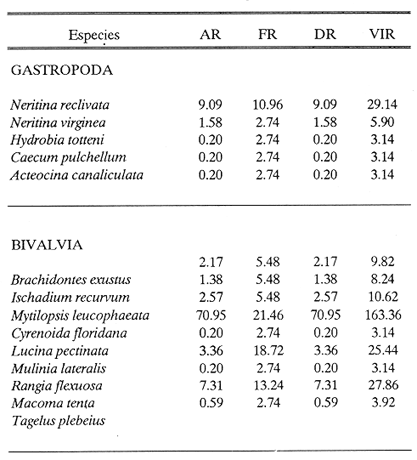 TABLA 5. Abundancia relativa (AR), frecuencia relativa (FR), densidad relativa (DR) y valor de importancia relativa (VIR) de la comunidad malacólogica viva La semejanza porcentual entre pares de estaciones, obtenida para la Clase Bivalvia, a partir de 91 combinaciones posibles (Fig. 4B), muestra que 42.8% se ubica dentro del rango 0-24, con 39 combinaciones; 30.8% en el intervalo de 50-74, con 28; 17.6% en el rango 25-49, con 16 combinaciones, y 8.8% se halla dentro del intervalo 75-100, con sólo ocho combinaciones. El promedio de similitud de este taxón es de 31.5%. La afinidad resultante de 91 combinaciones posibles, entre estaciones en las que se capturaron organismos vivos del Phylum Mollusca (Fig. 4C), indica que 50.5% corresponde al rango de 0-24, con 46 combinaciones; 26.4% se ubica en el intervalo 50-74, con 24; en el rango de 25-49 se presentan 16 combinaciones, con un porcentaje de 17.6%, y en el intervalo de 75-100 se registra 5.5%, con cinco combinaciones. La semejanza media relativa al conjunto es de 28%. 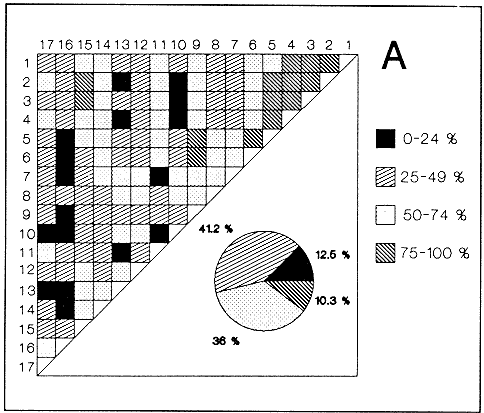 Figura 3A. Diagramas de enrejado de similitud entre estaciones de muestreo para la comunidad malacológica total. A) Clase Gastrópoda 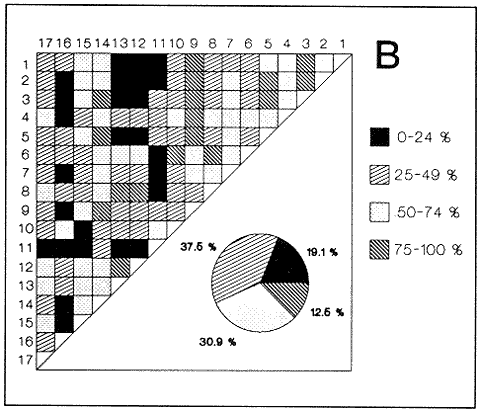 Figura 3B. Diagramas de enrejado de similitud entre estaciones de muestreo para la comunidad malacológica total. B) Clase Bivalvia 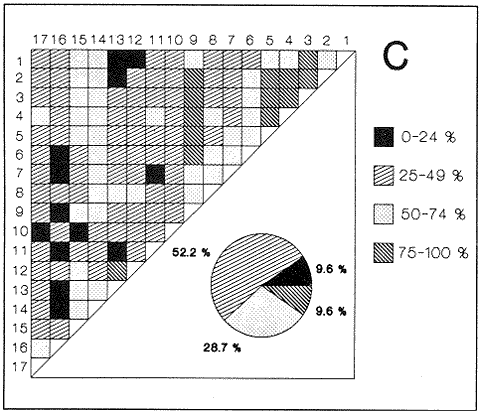 Figura 3c. Diagramas de enrejado de similitud entre estciones de muestreo para la comunidad malacológica total. C) Phylum Mollusca NUTRICIÓN Y HABITATDe acuerdo con la información bibliográfica, las especies de moluscos ocupan diversos habitat y presentan distintos tipos de nutrición, como se indica en la lista sistemática (Purchon, 1968; Andrews, 1971; Abbott, 1974; García-Cubas, 1981; Antolí y García-Cubas, 1985; Reguero y García-Cubas, 1989, 1991; García-Cubas et al., 1990; García-Cubas y Reguero, 1990). Además, tanto los gasterópodos como los bivalvos pueden utilizar diversas alternativas alimentarias y de relación con el entorno. Es por ello que, conforme a la literatura especializada, en las tablas 6 y 7 se enlistan las familias de moluscos presentes en este sistema lagunar, relacionándolas con su origen halino, posición respecto al sustrato y tipo de nutrición. En este complejo predominan las especies de habitat marino-salobre (Tabla 8). La mayor parte de los gasterópodos son epifaunales; entre los bivalvos destacan los organismos infaunales, y en el phylum hay porcentajes similares de epifauna e infauna. Las especies de la Clase Gastropoda son principalmente colectoras de depósitos, lo cual se manifiesta en valores altos también en el phylum, aunque estos últimos y los miembros de la Clase Bivalvia son en su mayoría suspensivoros. DIVERSIDADEn esta área, los índices de diversidad y sus componentes, tanto de la comunidad malacológica total como de la viva, muestran valores más altos si se considera el phylum (Tabla 9; Figs. 5 y 6). 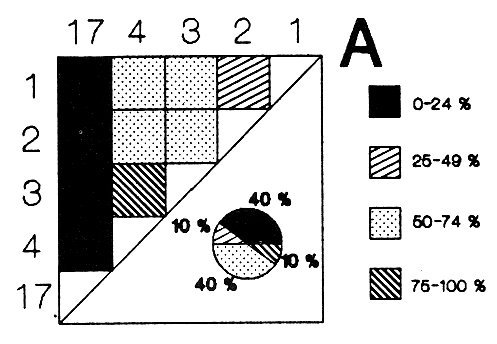 Figura 4A. Diagramas de enrejado de similitud entre estaciones de muestreo para la comunidad malacológica viva. A) Clase Gastropoda 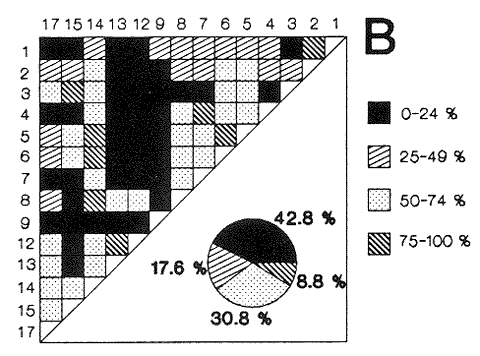 Figura 4B. Diagramas de enrejado de similitud entre estaciones de muestreo para la comunidad malacológica viva. B) Clase Bivalvia 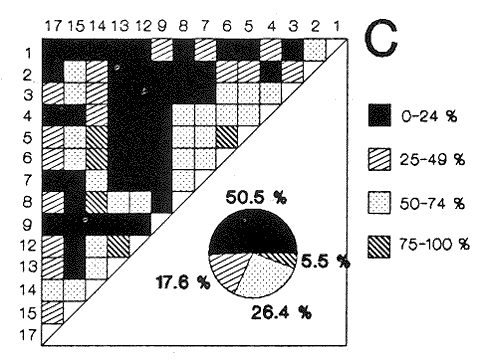 Figura 4C. Diagramas de enrejado de similitud entre estaciones de muestreo para la comunidad malacológica viva. C) Phylum Mollusca 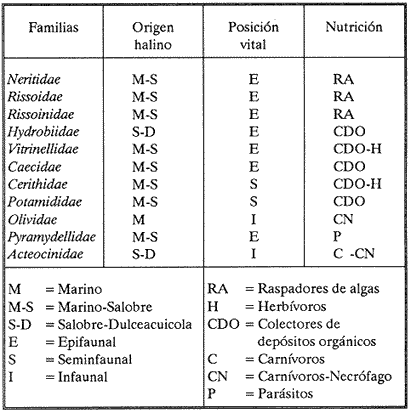 TABLA 6. Origen halino, relación con el sustrato y tipo de nutrición de las Familias de la Clase Gastropoda. 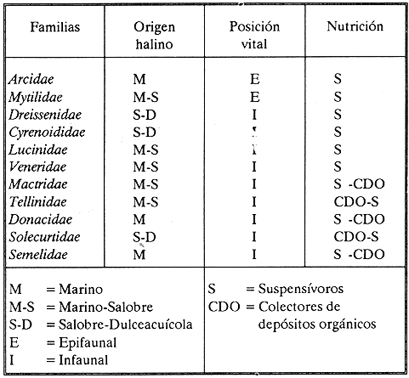 TABLA 7. Origen halino, relación con el sustrato y tipo de nutrición de las Familias de la Clase Bivalvia. 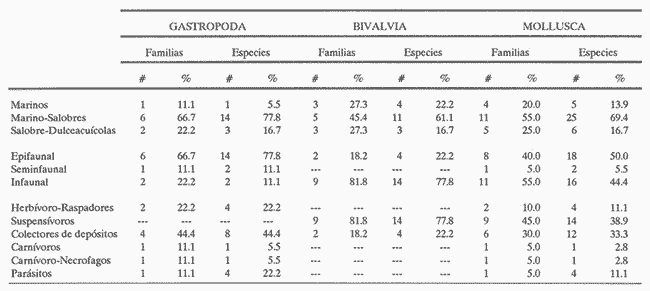 TABLA 8. Origen halino, relación con el sustrato y tipo de nutrición de las Familias y especies de las tres categorias taxonómicas. 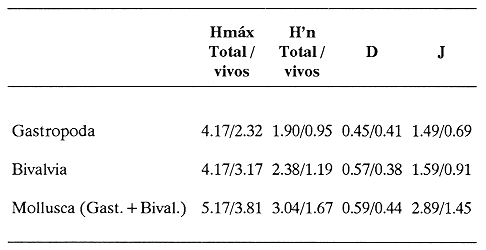 TABLA 9. Índices de diversidad máxima (Hmáx), diversidad (H'n) equitatividad (J) y riqueza de especies (D) por taxón. El análisis de resultados en cada una de las estaciones de muestreo indica que, entre las poblaciones totales de moluscos (vivos + muertos), la diversidad se registra en un intervalo de 0.09 a 2.41 bits/individuo en la Clase Gastropoda, de 0 a 2.23 para la Clase Bivalvia y de 0.33 a 3.25 el Phylum Mollusca, con medias de 1.30, 1.35 y 1.95, respectivamente. La equitatividad presenta fluctuaciones que van de 0.09 a 0.95, de 0 a 1 y de 0.14 a 0.93, de acuerdo con cada taxón. La riqueza de especies se encuentra entre los límites de 0.15 a 1.45 para gasterópodos, de 0 a 1.26 para bivalvos y de 0.61 a 2.08 para todo el conjunto (Tabla 10; Figs. 7 A-C). En las poblaciones vivas de gasterópodos, bivalvos y moluscos, el índice de diversidad varía de 0 a 1 bits/ind, 0 a 1.95 y 0 a 2.26, respectivamente, con promedios de 0. 14, 0.47 y 0.64 por grupo; el de equitatividad, de 0 a 1 en la Clase Gastropoda, 0 a 0.84 en Bivalvia y 0 a 0.80 en elphylúm; el de riqueza específica, de 0 a 1, 0 a 0.93 y 0 a 1.26 en los diferentes taxa (Tabla 11; Figs. 8 A-C). En este sistema, los cambios en la diversidad de las Clases Gastropoda y Bivalvia (vivos + muertos) se explican en mayor medida por la adición de especies a la comunidad; mientras que la uniformidad de abundancia de los individuos entre especies incide más sobre el phylum (Fig. 9). No obstante, en la comunidad viva de los bivalvos y moluscos analizados, la variabilidad de la diversidad se explica en un 97% por la riqueza de especies; para los gasterópodos hay mayor influencia de la equitatividad (Fig. 10). Por otra parte, la comunidad total de gasterópodos es más diversa en ambiente polihalino "A", en tanto que la de bivalvos y moluscos, en oligohalino. Las poblaciones vivas de los tres grupos presentan mayor diversidad en la zona oligohalina (Tabla 12; Figs. 11 y 12). 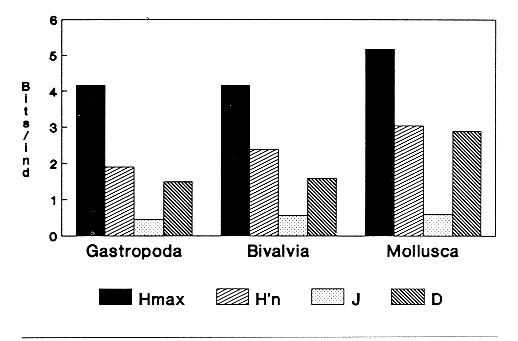 Figura 5. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D) para la comunidad malacológica total. 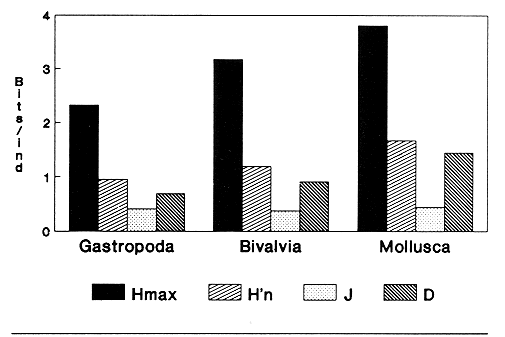 Figura 6. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D) para la comunidad malacológica viva. 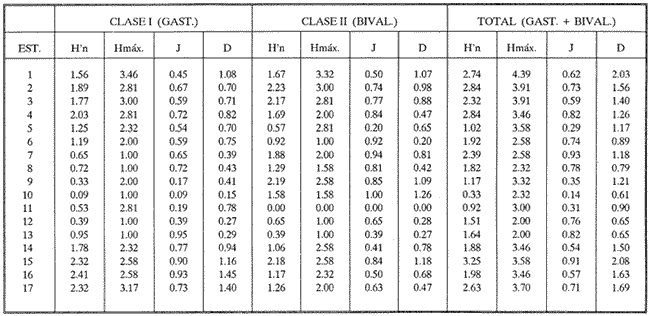 TABLA 10. Índices de diversidad y sus componentes, por estación, para la comunidad malacológica total. 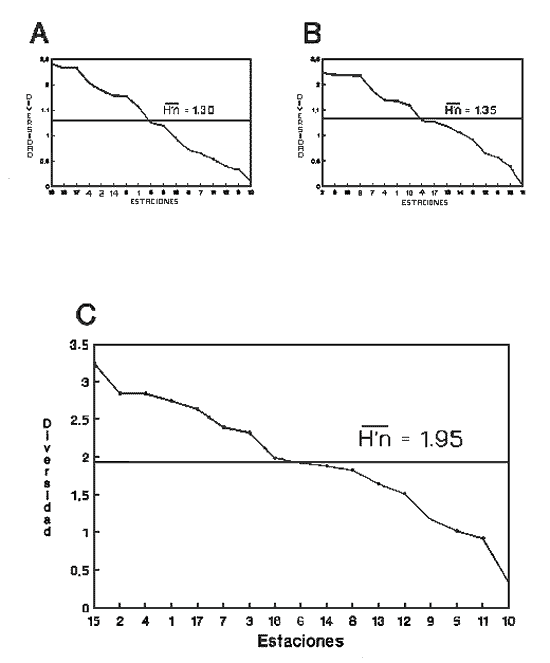 Figura 7. Curvas de valores de la diversidad en cada estación de muestreo, para comunidades totales. A) Clase Gastropoda; B) Clase Bivalvia; C) Phylum Mollusca. 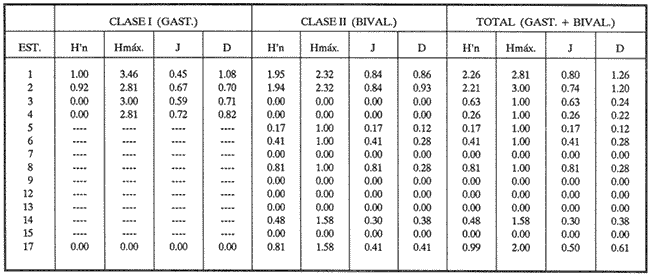 TABLA 11. Índices de diversidad y sus componentes, por estación, para la comunidad malacológica viva. 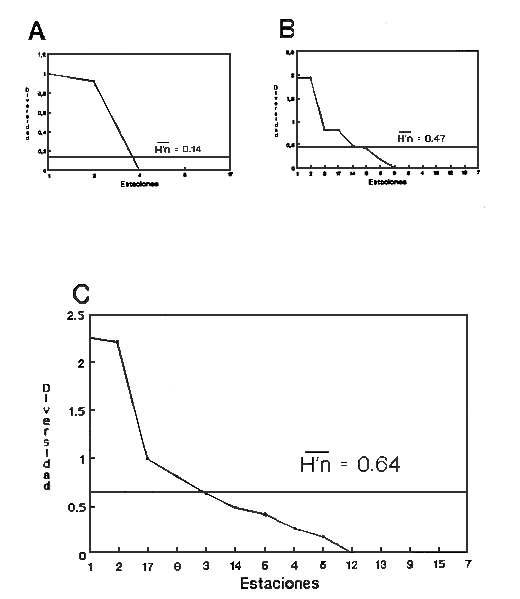 Figura 8. Curvas de valores de la diversidad en cada estación de muestreo, para comunidades vivas. A) Clase Gastropoda; B) Clase Bivalvia C Phylum Mollusca. 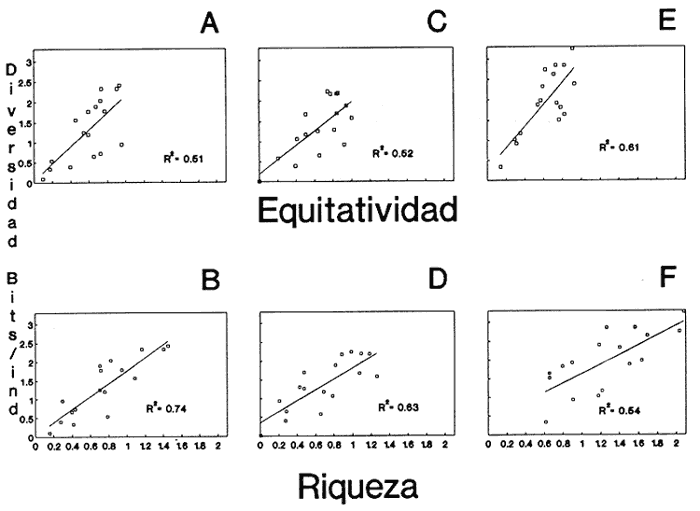 Figura 9. Coeficientes de determinación R2 entre la diversidad y los componentes de equitatividad y riqueza de especies, para comunidades totales. A-B) Clase Gastropoda; C-D) Clase Bivalvia; E-F) Phylum Mollusca. 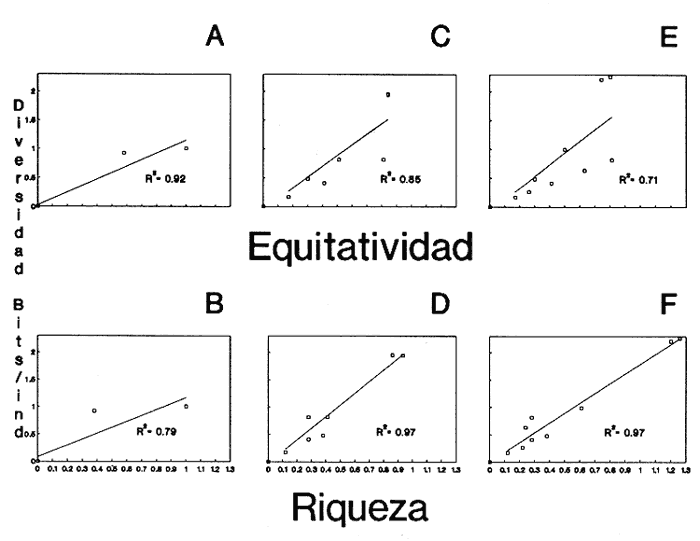 Figura 10. Coeficientes de detreminación (R2) entre la diversidad y los componentes de equitatividad y riqueza de especies, para comunidades vivas. A-B) Clase Gastropoda; C-D) Clase Bivalvia; E-F) Phylum Mollusca. 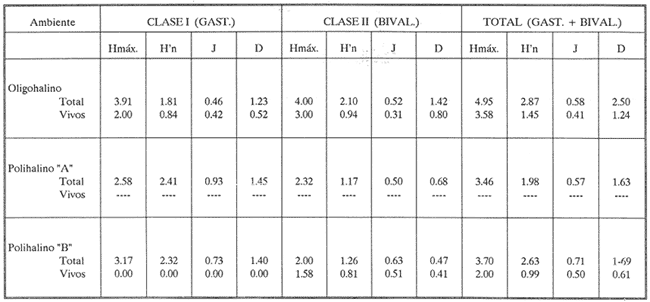 TABLA 12. Índices de diversidad y sus componentes, por ambiente halino. 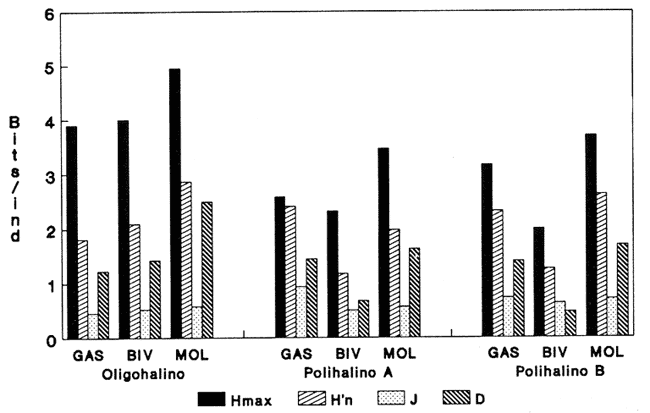 Figura 11. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D), por ambiente halino, para la comunidad malacológica total. 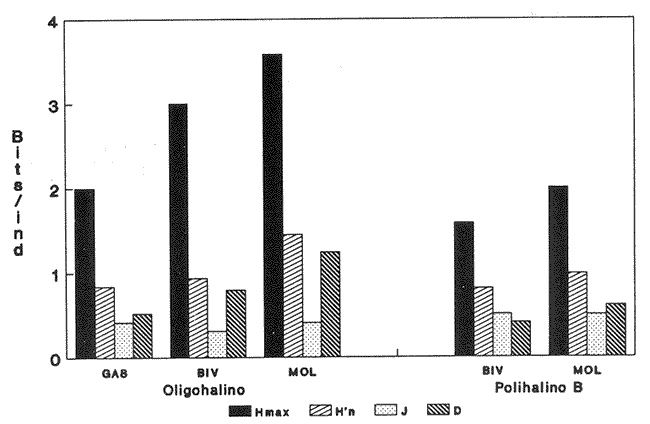 Figura 12. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D), por hambiente halino, para la comunidad malacológica viva. DISCUCIÓN Y CONCLUCIONESLos valores promedio de temperatura, registrados tanto en la superficie como en el fondo, son altos y semejantes (3232.5ºC), lo cual es atribuible principalmente a la estación del año en que se llevó a cabo el muestreo, que corresponde a la época más calurosa. Así mismo, la baja profundidad y el alto grado de insolación contribuyen, en conjunto, a mantener las aguas a temperaturas elevadas. También la salinidad está influida por lo somero del sistema, pues hay diferencias mínimas entre los valores de fondo y los de superficie. No obstante, la distancia con respecto al mar o a la desembocadura de arroyos provoca variaciones en la distribución horizontal, que va de ambientes oligohalinos en los dos cuerpos de agua hasta polihalinos de tipo "B" en las estaciones ubicadas en el canal de comunicación con el mar, con registros mínimos en la Laguna Chica. Por el promedio de profundidad en el área (0.70 m) se trata de dos lagunas someras. Las estaciones con valores más altos se localizan en sitios donde la influencia de las corrientes es más notoria, pues son los lugares más estrechos del sistema. El incremento en el caudal de los arroyos durante la temporada de lluvias provoca así mismo aumento en la cantidad de terrígenos en suspensión, lo cual se manifiesta en un promedio bajo de transparencia (0.10 m). En el área abundan los sedimentos limo-arcillosos, salvo en las localidades cercanas a la comunicación con el mar, en donde hay mayor cantidad de arena. El análisis de la comunidad malacológica indica predominio de la Clase Gastropoda en lo referente a cantidad de individuos, aun cuando el número de especies identificadas fue el mismo que para la Clase Bivalvia. Las estaciones que presentan la mayor abundancia se localizan en la parte austral de la Laguna Grande, en donde se mezclan sus aguas oligohalinas con las polihalinas que ingresan del mar. En el sistema abundan las especies marino-salobres. Entre los gasterópodos predominan los individuos epifaunales, colectores de depósitos; mientras que los bivalvos son preferentemete infaunales y suspensivoros. La diversidad promedio arroja valores similares para las poblaciones totales de las dos clases, con una diferencia de alrededor de 0.6 bits/ind respecto alphylum. En las comunidades vivas se observa mayor diversidad de miembros de la Clase Gastropoda. De acuerdo con los coeficientes de correlación entre la diversidad y sus componentes de equitatividad y riqueza, la adición de especies explica en mayor medida los cambios en la diversidad de las clases, y la equitatividad incide principalmente en el phylum. Finalmente, se observa que las poblaciones vivas de las tres categorías taxonómicas son más diversas en ambiente oligohalino, al igual que la comunidad total de bivalvos y moluscos en general; mientras que el índice de diversidad de gasterópodos totales es superior en la zona polihalina de tipo "A". AgradecimientosLos autores reconocen la participación de Francisco Escobar de la Llata y Vicente Antolí Figuerola en la recolección de las muestras y agradecen la valiosa ayuda de Gerardo Barrientos Mac Gregor en la elaboración de material gráfico. LITERATURAABBOTT, R. T. American seashells. The marine Mollusca of the Atlantic and Pacific coast of North America Van Nostrand Reinhold Co 2a. ed. New York, 1974. 666 p. ANDREWS, J., Seashells of the Texas coast. University of Texas Press, Austin, 1971. 298 p. AUTOLÍ V. y A. GARCIA-CUBAS, An. Inst. Cienc. del Mar y Limnol. Sistemática y ecología de los moluscos en las lagunas costeras Carmen y Machona, Tabasco, México. Univ. Nal. Auton. México, 1985. 145-198. (1). 12 CARRANZA-EDWARDS, A., M. GUTIÉRREZ-ESTRADA y R. RODRÍGUEZ T., An. Inst. Cienc. del Mar y Limnol. Unidades morfotectónicas continentales de las costas mexicanas. Univ. Nal. Autón México, 1975. 81-88. (1): 2 CONTRERAS, F., Las lagunas costeras mexicanas Centro de Ecodesarrollo, Secretaría de Pesca, D. F., 1985 253 p. GARCIA, E. An. Inst. Biol. Los climas del Estado de Veracruz (según el sistema de clasificación climática de Köppen modificado por la autora). Ser. Botánica, Univ. Nal. Autón. México 1971. 3-42. (1): 41 GARCIA-CUBAS, A., Bol. Inst. Geol. Sistemática y distribución de los micromoluscos recientes de la Laguna de Términos, Campeche, México. Univ. Nal Auton. México, 1963. 1-55, 24 figs., 4 láms. (4): 67 GARCIA-CUBAS, A., An. Inst. Cienc. del Mar y Limnol. Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de Términos, Campeche). Univ. Nal Autón. México 1981 1-182. Publ, Esp. 5: GARCÍA-CUBAS, A., F. ESCOBAR DE LA LLATA, L. V. GONZÁLEZ-ANIA y M. REGUERO. An. Inst. Cienc. del Mar y Limnol. Moluscos de la Laguna Mecoacán, Tabasco, México: sistemática y ecología. Univ. Nal. Autón. México, 1990 1-30. (1): 17 GARCIA-CUBAS, A. y M. REGUERO, , An. Inst. Cienc. del Mar y Limnol. Moluscos del sistema lagunar Tupilco Ostión, Tabasco, México. Univ. Nal Autón. México, 1990. 309-343. (2): 17 KEEN, M. A. Seashells of tropical west America. Marine mollusks from Baja California to Peru. Stanford University Press, 2a. ed., Stanford, 1971. 1065 p., 22 láms. KREBS, J. C. Ecology: The experimental analysis of distribution and abundance. Harper International Edition, New York, 1972. 694 p. LANKFORD, R. R., Estuarine Processes Coastal lagoons of Mexico: Their origin an classification. In: M. L. Wiley (Ed.). Academic Press Inc., New York 1977. 182-215. 2: MARGALEF, D. R., Information theory in ecology. General systems. Transl. from Mem. Real Acad Cienc. Arts. Barcelona, 1958. 373-449. 32: PIELOU, E. C., Ecological diversity. M. L. Wiley (Ed.), New York. 1975. 165p. PURCHON R. D., The Biology of the Mollusca. Pergamon Press, New York, 1968. 560 p. REGUERO, M. y A. GARCIA-CUBAS, An. Inst. Cienc. del Mar y Limnol. Moluscos de la Laguna de Alvarado, Veracruz: sistemática y ecología. Univ. Nal. Autón. México, 1989. 279-306. (2): 16 REGUERO, M. y A. GARCIA-CUBAS, An. Inst. Cienc. del Mar y Limnol Moluscos de la Laguna Camaronera, Veracruz, México: sistemática y ecología. Univ. Nal Autón. México, 1991. 1-23. (1): 18
|