|
ANÁLISIS EXPLORATORIO DE OTOLITOS
ASIMÉTRICOS EN EL PEZ PLANO Kareius bicoloratus EN LA BAHÍA DE
TOKIO
Trabajo recibido el 24 de abril de 1991 y aceptado para su
publicación el 9 de diciembre de 1991.
Isarias Hazarmabeth Salgado-Ugarte
Carrera de Biologia, Coordinación de Ciclo
Terminal, Escuela Nacional de Estudios Profesionales "Zaragoza", UNAM. Apartado
Postal 900, México D.F. 09230, México.
El pez plano Kareius bicoloratus
(ishigarei en japonés) es una especie abundante,
distribuida ampliamente en aguas costeras de Japón. Como
pleuronéctido, además de dimorfismo sexual, K.
bicoloratus muestra asimetría morfológica en grado
notable, la cual se manifiesta en los otolitos. El estudio de la
asimetría de éstos es importante, debido a que los
parámetros del crecimiento se calculan por medio de relacionales
estadísticas y matemáticas entre medidas de longitud del pez y de
los otolitos. Sin datos cuantitativos acerca de la asimetría de esta
estructura calcificada en el ishigarei y con el fin de
evaluar su utilización para la determinación de la edad y el
crecimiento, este estudio examina algunas medidas de los otolitos de machos y
hembras por medio de procedimientos estadísticos exploratorios y
convencionales. Los diagramas de tallo y hoja, resúmenes literales,
asícomo los diagramas de caja condujeron a la caracterización de
la diferencia entre los otolitos izquierdo y derecho de K
bicoloratus: el tamaño y el peso totales de estas estructuras
no difieren significativamente; sin embargo, el núcleo del otolito
derecho está localizado hacia el borde posterior, mientras que el
izquierdo tiene una posición más central. Esta diferencia parece
tener relación con los cambios estructurales y sensoriales en el
cráneo debidos al hábito particular de yacer en el fondo sobre un
lado del cuerpo. Los valores de longitud de los anillos de crecimiento
mostraron algunos casos de exclusión eviden ciados mediante
métodos exploratorios más confiables por lo que resultó
preferible utilizar como representativas las categorías
estadísticas mediana y pseudosigma F. El diagrama de
escalera junto con el análisis de correlación mostraron
correspondencia más clara entre la longitud delpez y el radio anterior
delotolito derecho, lo que se contrapone a la costumbre de utilizar el otolito
izquierdo. A la edad I, los machos mostraron mayor
tamaño que las hembras; sin embargo, a partir de la edad II este
comportamiento se invirtió. En general, las longitudes retrocalculadas
fueron mayores que las conocidas para otros sitios, lo cual indica que en la
Bahía de Tokio este pez plano parece crecer más que en otras
localidades. Sin embargo, para concluir acerca de su crecimiento en este lugar,
es necesario el análisis de una muestra mayor de
espéciménes que incluya peces de mayor edad.
The stone flounder Kareius
bicoloratus (Japanese name ishigarei) is one of
the most abundant and widely distributed species in Japanese coastal waters.
Besides sexual dimorphism, K. bicoloratus as a pleuronectid
shows a remarkable degree of morphological asymmetry, wich in turn is reflected
by its otoliths. Examination of the otoliths asymmetry is very important by the
fact mathematic and statistical relationships between fish and otoliths length
are often employed to estimate growth parameters. With no quantitative reports
on this calcified structures asymmetry, some male and female otoliths
measurements are examined by means of several exploratory and conventional
statistical procedures, in order to evaluate their utilization for age and
growth determination. Overall size and weight of ishigarei's left and right
otoliths do not differ significantly, nevertheless the right otolith nueleus
position is located toward the posterior edge, whereas the left one has a more
central position; stem and leaf displays, letter values resumes and boxplots
lead to the characterization of such difference, which seenis to be related to
the structural and sensorial cranial changes involved with the particular habit
of laying on bottom over one side of the body. Resistant exploratory methods
made evident the existence of outliers in the data set of radial distances
between nucleus and annuli; consequently the employment of the resistant
estimators median and F-pseudosigrna resulted advisable. The draftsman's plot
and the correlation analysis showed a clearer correspondence between fish
length and anterior radius of the right otolith than for the left one: this
fact distrust the employnient of the last one as a c ominon practice. With the
exception of age I, females showed greater sizes than males. The
back-calculated lengths for this species at Tokio Bay resulted generally
greater than the ones found for other locations; however, it is necessary to
analyze bigger samples including older fishes in order to achieve a right
conclusion.
The study of age and growth is essential for the proper assessment of fish populations. The knowledge of growth parameters and the population's age structure allows to estimate mortality, stock size and other values involved with its exploitation. The most usually employed methods to determine the age of fishes are the analysis of length-frequency distribution and the reading of calcified structures. Distribution of length frequencies in most temperate water fishes is polymodal (each mode representing fishes with the same age) and generaly is suitable to gaussian component decomposítion, with some exceptions (Harding, 1949; Cassie, 1954; Tanaka, 1962; Hasselblad, 1966; Bhattacharya, 1967). The study of calcified structures enclose the reading of scale, otolith, and some other structures' growth marks (Bagenal and Tesch, 1978; Pannella, 1971; Brothers, 1987). In some fishes (scaleless or tropical) the reading of calcified structures like otoliths, vertebrae or fin spines sections is a simple and practical manner to determine the age of caught samples. Furthermore, due to the problems associated with scale reading (asymptotic growth of scale in old fishes, scale resorption, etc.) in some instances the other calcified structures are the only reliable way to know the age. The otolith analysis may involve the enumeration of annual (or other periodic) marks (Williams and Bedford, 1974) or the counting of daily growth increments (Campana and Neilson, 1985; Brothers, 1987). The reading of annual marks in otoliths requires the identification of hyaline and opaque zones, which appearance is dependant on the kind of the used illumination (reflected or transmitted). To achieve this, the otoliths may be observed whole, broken and charred, grounded, or sectioned and grounded. The type of treatment depends on the characteristics of the otoliths, such as size, shape or thickness, which are related with the species and age of the fish. In order to accomplish a precise age estimation of fishes, it is important to analyze these otolith characteristics first. Pleuronectiformes are unique in that adult fishes have both eyes on the same side of the body, fact that implies a high degree of asymmetry (Eschmeyer et al., 1983; Prasada-Rao and Finger, 1984). When the fish hatches from. the egg is symmetrical, but shape and position of fins and other body structures change along development of larval stages; eventually one eye migrates across the top of the head and the fish takes up the habit of lying on bottom with both eyes facing up (Policansky, 1982). The typical asymmetrical architecture of this order is better manifested in the neurocranium, suspensorium and oral jaw apparatus. In Microchirus theophila (Pleuronectiformes, Soleidae), the otic region (neurocranium) shows an increasing degree of asymmetry in a rostral direction (Bürgin, 1986). The stone flounder Kareius bicoloratus (japanese name ishigarei) is an abundant and widely distributed species in Japanese coastal waters (Mito et al., 1969). This fish normally is dextral, having both eyes and coloration on the right side of the body, although some reversal cases have been reported (Fujita, 1980). K. bicoloratus is a scaleless fish with several rows of rugose plates on the eyed side and a maximal length around 60 cm (Ochiai and Tanaka, 1986). This species has been the subject of several studies, including: egg development and rearing of the larvae (Mito et al., 1969); carly life story (Minami, 1984); larval immigration (Tsuruta, 1978); distribution of fry according to environmental factors (Takagoshi et al., 1976). The analysis of age and growth has been undertaken by several authors: Hatanaka et al. (1952), who also studied the spawning in Sendal Bay; in the Fukushima Prefecture coastal zone, Takagoshi and Akimoto (1975) analyzed the growth and habitat of the fingerling group; Akimoto et al. (1978) carried out tagging experiments in young specimens; by means of otolith reading, Hirakawa (1980) studied the age and growth. In 1982, Shoji et al., analyzed the growth and maturity in Choshi Sea. Recent ly, masaki et al. (1986), with specimens from Suo-Nada of Seto Inland Sea, and using the otolith reading, studied the age and growth of the stone flounder. As a pleuronectid, K. bicoloratus shows a remarkable degree of morphological asymmetry, which in turn is reflected by the otoliths. The study of this otolith asym metry is important because the growth parameters are estimated by using statistical and mathematical relationships (de regressión, von Bertalanffy growth equation, etc.) between fish length and otolith length measures. With no quantitative reports on the asyminetry of this calcified structure of the stone flounder, and in order to assess its utilization for age and growth determination, the present study examines some otolith measures by means of some exploratory (Tukey, 1977; Hoaglin et al., 1983) and conventional statistical procedures (Sokal and Rohlf, 1981; Snedecor and Cochran, 1980). MATERIAL AND METHODSMonthly samples of stone flounder were obtained from commercial capture in Tokyo Bay from June to December 1989. Both sides otoliths (sagitta) were obtained from a total of 187 examined individuals. After thawing, specimens were measured to the nearest mm of standard and total lengths, weighed to the nearest g and dissected to determine sex by the observation of gonads. The otoliths were extracted by means of a cranial incisión, washed with water to remove membranes and other tissue remains, and kept dry in glass vials. The otoliths were observed under reflected light with black background by means of a dissection microscope and measured with the'included micrometer (K.K. Kougaku) to the nearest 0.01 mm. The left (L) and right (R) otoliths were observad and six measures were registered (Fig. 1): ANTERIOR OTOLITH RADIUS (AOR). The length from the otolith focus to the anterior margin along the central anteroposterior axis (APA). POSTERIOR OTOLITH RADIUS (POR). The length from the otolith focus to the posterior margin along the APA. ANTERIOR ANNULUS RADII (AAR). The length from the otolith focus to the anterior outer margin of the hyaline zone. POSTERIOR ANNULUS RADII (PAR). The length from the otolith focus to the posterior outer margin of the hyaline zone. OTOLITH DIAMETER (OD). The result of the addit- of AOR and POR. ANNULUS DIAMETER (AD). The addition of the corresponding AAR and PAR values per each annulus. Besides the above length measures, each otolith was weighed to the nearest 0.0001 g with an analytical balance (Inaba Seisakusho Ltd.). The quoted morphometric values were subjected to the following statistical procedures: STEM AND LEAF DISPLAYSThe stem and leaf display is an hybrid between a histogram and a frequency table, that enables to notice data frequency distribution features such as symmetry, spread, concentrations and gaps. To construct the simplest form of this display is necessary to choose a suitable pair of adjacent digits in the data. Further each data value is split between these two digits (for example 360 mm of total length can be split into 36 and 0). Then a separate line in the display is allocated for each pos- string of leading digits (the stem). lFinally, the first trailing digit (the leaf) of each data value is written down on the line corresponding to its leading digits (Tukey, 1977; Emerson and Hoaglin, 1983). The values 329,342, 357, 359, 360, 364, 365, 372, 376, 384, and 392 (total length in mm) can be represented as follows: 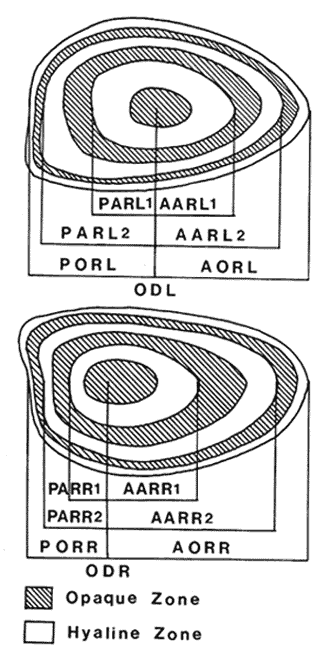 Figure 1. Left (upper) and right (lower) otoliths of Kareius bicoloratus with the measures considered: PAR(L, R)n = Posterior annulus radius of the left or right otolith. AAR(L, R)n = Anterior annulus radius of the left or right otolith. AOR(L, R) Anterior otolith radius of the left or right otolith. OD(L, R) Otolith diameter of the left or right otolith. The annulus diameter (AD) is not indicated but corresponds to the line obtained by addition of the respective PAR(L, R)n and AAR(L, R)n pairs. This display includes a reminder of the unit employed, as well as a column of depths (see below). There are several variations of this basic form (for details see Emerson and Hoaglin, 1983; Curts, 1986). 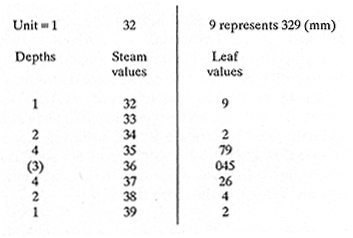 LETTER VALUES DISPLAYS. This display is a statistic summary based on statistics derived from the original data set after sorting and counting, which are labeled with letters like M (for median) and F (for "fourths", a form of the quartiles). These letter values are resistant, that is, an arbitrary change in a small part of the data can have only a small effect on the summary. On the other hand, the mean and variance can be affected substantially by a single wild data (Velleman and Hoaglin, 1981; Hoaglin, 1983). The first step to obtain the letter values, after ordering the data set (batch) is to determine the values position in the enumeration of values, starting at the nearest upper and lower ends of the batch. This position is the depth of each data value. If the number of data (n) is odd, there is a deepest data value, the median. Otherwise the median is the average of the two middle values. Fot the median´s depth evaluation the following expressión is employed (Velleman and Hoaglin, 1981): 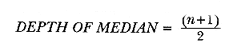 For the data set used above, n = 11, then the depth of median equals: (11 + 1)/2 = 6. The sixth value on the ordered sequence represents the median value (364). Another pair of summary values are the fourths; the fourths represent another pair of summary values with a depth, defined as follows: 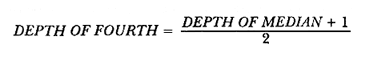 where numerator stands for the operation of finding the largest integer not exceeding the depth of median value. Using the batch of fish lengths, the depth of the fourths (one fourth for data lower than median and another fourth for higher data) is (6 + 1) / 2 = 3.5, where the fraction indicates that both fourths must be obtained from the average of the values with depth 3 and 4. In this case, for the lower half. (357 + 359)/2 = 358 = FL; and for the upper half (376 + 372)/2 = 374 = Fu. Extremes are the simplest summary values to evaluate. They are represented by those data with a depth. to 1 in the stem and leaf display; equal the highest and lowest values in the ordered data set are: 329 and 392 (Hoaglin, 1983). The letter values are displayed in the five-number summary form. For our data set, the five-number summary is: 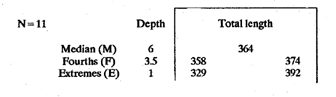 The difference between the upper fourth (FU) and lower fourth (FL) is defined as interquartific range, or fourth spread, which is a measurement of the data set dispersión analogue to standard deviation (374358 = 16). F-pseudosigma can be found according to the following expressión (Hoaglin, 1983): 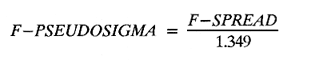 The use of the resistant F-pseudosigma in addition to, or instead of, the nonresistant standard devíation (s) has some advantages, particularly when data contain wild observations or outliers (Hoaglin, 1983). For the above lengths example, this resistant dispersión measure equals 16/1.349 = 11.86. On other hand the standard devíation is 17.03 and the mean is 363.64. When the two estimates differ substantially, it is better to choose the F-pseudosigma and look for those observations influencing s (Hoaglin, 1983; Iglewicz, 1983). In order to make more evident the effects of a wild value assume that the last of the length's values for our example is 500 mm. Median (364) and F-pseudosigina (11.86) remain unchanged, but mean (373.45) and standard devíation (42.56) are higher than the prev- ones obtained. In this case the median is more representative of the data batch than the mean. To obtain the best location estimates of the data sets, a comparison between the mean and standard devíation values with the median and F-pseudosigina values was carried on. BOXPLOTS. This kind of plots is based on the fivenumber summary and the fourth-spread of a batch of data. The boxplot shows structural distribution features such as location, spread, skewness, tail length and outdata points (Tukey, 1977; Emerson and Strenio, 1983). This plot consists of boxes drawn with both ends at the lower and upper fourths and a crossbar at the median. The next step is to draw a line from each of the box to the farest point that is not an outlier. The outlier cutoffs are defined as FL - 3/2 dF and Fu + 2/3 dF, where dF is the fourth spread Fu - FL (Emerson and Strenio, 1983). The values greater or smaller than these cutoff values are considered like possible outliers and are indicated by asterisks or separated points in the graphic. In this way, the location is indicated by the median (the crossbar inside the -bar); the length of the box shows the spread (by the fourth spread). The relative positions of the median, upper fourth and the lower fourth make possible to look at the symmetry (skewness); the tail length can be seen by the lines extending from the box and by the outlier points. A display of parallel boxplots makes easy the comparison of several batches of data. To have an objective judgtnent about the difference between the several measure data sets, the multiple notched boxplots were employed. This boxplot varíant uses the overlapping and non-overlapping regions of intervals defined by. median ± 1.58* dF/n placed symmetrically around the median, and marked by a notch in the side of the box (Mc Gill et al., 1978). The groups whose notched intervals do not overlap can be said to be significantly different at roughly the 5% individual level (Velleman and Hoaglin, 1981). DRAFTSMAN'S DISPLAY. One way to represent a number n of variables in a dimensiónal view is to use the draftsman's display that consist in an array of n paired scatter plots with the essential property that any adjacent pair of plots have an axis in caminan (Chambers et at, 1983; Curts et al., 1987). The resulting scatterplot matrix has the organization of a correlation matrix with the additional advantage of showing trends (like curvilinearity) and outliers that are no obvious just observing the numerical values of correlation matrix (Hamilton, 1990). In both correlation matrix and draftsman's display the cells above the diagonal are redundant. Therefore, only the lower half part was sidered. For confirmatory (conventional) statistical analysis, a correlation analysis between otolith radius measure and body length values was carried out. Regression analysis were performed based upon the highest correlation values between otolith and fish length values. The analysis of covariance (tests of equality of slopes and intercepts) was employed to verify the differences between male and female equations. The length-weight relationship was determined and the significance and difference between sexes were verified RESULTSFigure 2 contains the stem and leaf displays for the female otolith radius. It is easy to see that the anterior and posterior sides of left otolith are more or less equally distributed, whereas the anterior and posterior measurements of the right otolith show a remarkable difference. The anterior side radius of this otolith is larger than the left one. Values for male presented a similar behavior. The female otolith measurements show a clear multimodal or at least bimodal distribution pattern, with some gaps; males were more unimodal, with a shorter spread.  Figure 2a. Parallel stem and leaf displays for otolith radius of females. LEFT OTOLITH. About annulus radii (AR), most individuals presented one annulus; only 9 specimens had more than one ring (Table 2b). The stem. and leaf displays for the radii of the first annulus belonging to females are showed in Fig. 3. As expected, the values have a gaussian frecuency distribution and, like the otolith radú, the anterior-posterior measurements of the left otolith are more similar in length than those of the right otolith. In these radii, the female values are slightly smaller than those of the male.  Figure 2b. Parallel stem and Icaf displays for otolith radius of females. RIGHT OTOLITH. The median, fourths and the extremes values can be compared using the five number summaries (Tables 1 and 2a) and the multiple boxplots in Figure 4. The difference between the anterior and posterior measurements of right and left otoliths becomes quite evident both in males and females. According with the notched boxplots of Fig. 4, the anterior (AORL, median = 2.375 and 2.99 mm for males and females, respectively) and posterior (PORL, median = 2.22 and 2.79 for males and females) radii of the left otolith are not different (the notches overlap). On the other hand, the right otolith anterior-posterior measurements show significant differences, indicated by the non-overlapping notches; the medians for the anterior right side (AORR) are 2.95 and 3.545, whereas the posterior values (PORR) are 1.62 and 1.895, for males and females, respectively. In spite of the anterior-posterior differences found, the left and right otolith diameters (Fig. 4) and the otolith weights (Fig. 5) are very similar, with clearly overlapping notches. A similar patternis showed by the first annulus radii: the anterior and posterior radii have very similar lengths, but the corresponding measures for the right otolith are noticeably different, with non-overlapping notches (Fig. 6). In these measurements, the male values are slight1y larger than those for females (Table 2a). Except for AARL1, the female values show some outlier points. As in the otolith radius measures, the diameter of the first annulus is almost the same in both otoliths, in spite the anterior-posterior measurements differences.  Figure 3a. Parallel stem and leaf displays for the first annulus radius of females. LEFT OTOLITH. Due to the small number of individuals that presented two annuli, the results obtained are considered as preliminary, but in general the trend is like that of the first annuli: the left otolith anterior-posterior measurements are closer than the same values measured on the right otolith. In Table 3 is presented the comparison between the mean and standard devíation with the median and F-pseudosigma (FPS). For otolith radius (Table 3a) the SD values are smaller than FPS for male otolith measurements, except in the case of PORL; on the other hand, the female values show an opposite trend where SD values are bigger than FPS, except in the case of AORL. This fact remarks the influence of the outlier points in the female data. In the part b) of Table 3 where the first annulus radius measures are included, FPS is smaller than SD in: a) males, PARR1; and b) females, PARL1 and PARR1, in spite that the mean and median values are very similar. This indicates that these mean values are affected by extremes figures in the data and, according to Hoaglin (1983) and Iglewiez (1983), in this circumstance is better to choose the resistant values (median and F-pseudosigma) looking at the causes that increased the SD value. 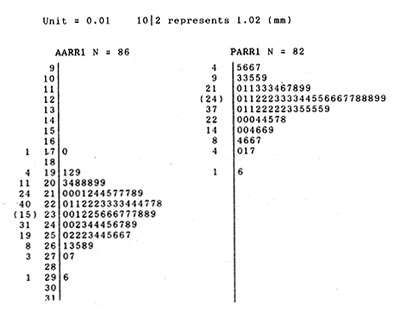 Figure 3b. Parallel stem and leaf displays for the first annulus radius of females. RIGHT OTOLITH. 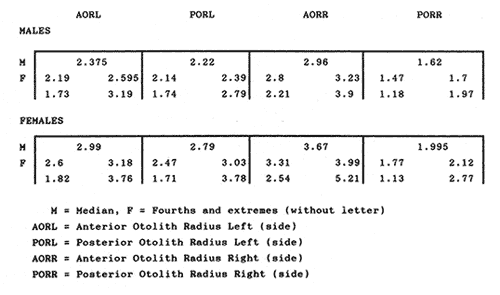 TABLE 1 Five-number summary for otolith radius measures 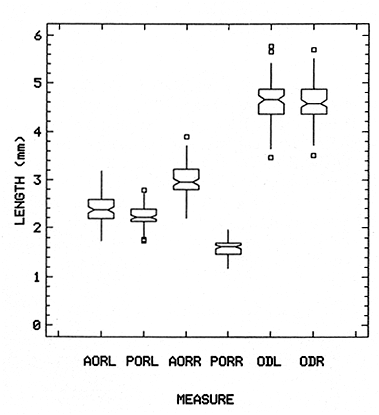 Figure 4a. Notched boxplots for otolith measures (males). 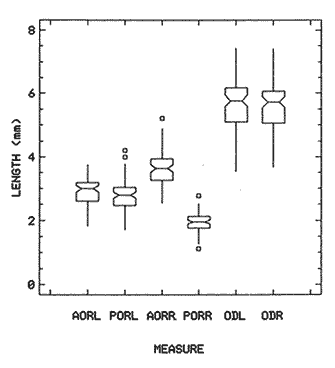 Figure 4b. Notched boxplots for otofith measures (females). The second annulus radius values, prefiminary considered because of the small number of specimens, present FPS values larger than SD, except in the females AARL2 (Table 3c). To have a global view of the relationship between the fish and otolith length measurements, a correlation analysis was carried out. The numerical results are dis played in Table 4 (males) and Table 5 (females). The draftsman's plot (Chambers et al., 1983) permits the graphical appreciatíon of the relationship between the variables, in this case the otolith measurements and fish length (Fig. 7). In the correlation matrix for the male values (Table 4a) the anterior measures of the left otolith (AORL) show the highest correlation coeffici- with total length and standard length (approx. 0.81, followed by PORL and AORL. For the females, the highest correlation coefficient with total length and standard fish length is shown by the posterior otolith radius values of the left otolith (approx. 0.93), fóllowed by AORR and AORL (Table 4b). In both, males and females, the smallest: correlation índices are shown by the posterior right otolith measurements. 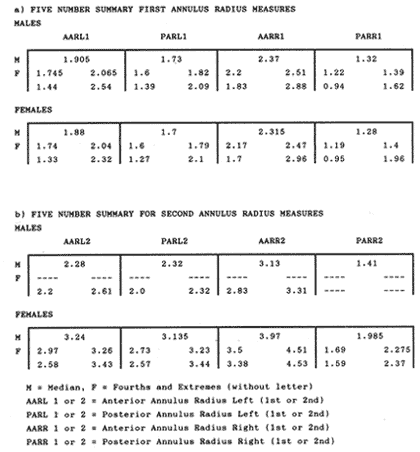 TABLE 2 Within the otolith radii measurements the highest correlation values are those presented by two of the longest otofith measures, that is, the correlation among the anterior right otolith radii with the posterior left otolith redii. The lowest correlation values occurs between the anterior and posterior radii of the right otoliths. From the draftsman's plot it can be easily seen that the male data appear more sparse than those of females, and one of the reasons for this is the shorter length size range of the male sample. The relationship between the otolith radius and the fish size was calculated considering the above highest correlation values. Two pairs of fish and otolith length measures were chosen: total fish length with anterior radius of the right otolith (TL-AORR), and standard length with posterior radius of the left otolith (SLPORL). The resulting regressións showed a high significance, with no defined trend of residuals. 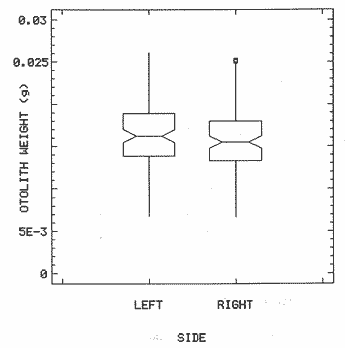 Figure 5a. Notched boxplots for otolith weight (males). 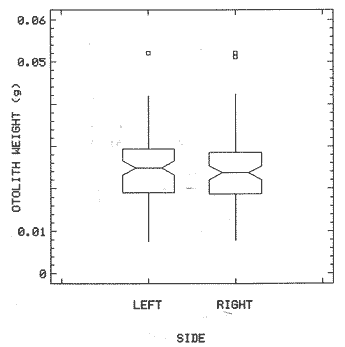 Figure 5b. Notched boxplots for otolith weight (females). 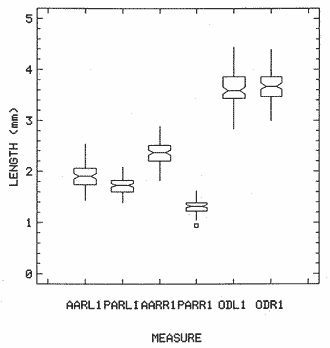 Figure 6a. Notched boxplots for the first annulus measures (males). 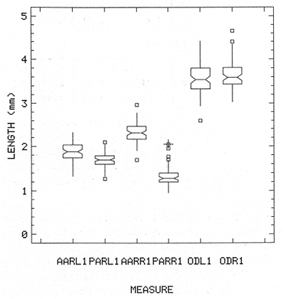 Figure 6b. Notched boxplots for the first annulus measures (fernales). From the covariance analysis results for male-female comparison, highly significant differences among the slope and intercept values were found. Therefore, the data were considered separately for each sex. The equations for the fish length-otolith radius regression data are: For total length (TL)-AORR: 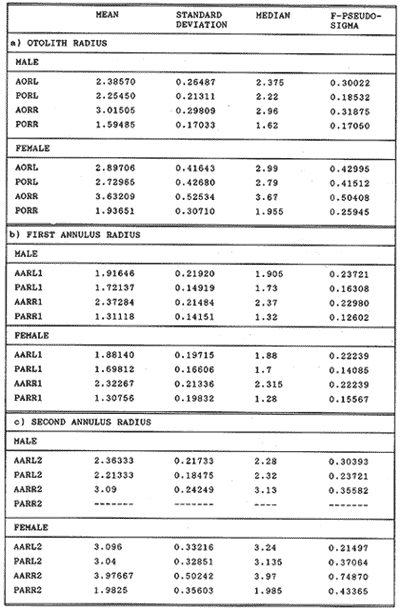 TABLE 3 Comparison between mean deviation with median an F-pseudosigma values of otolith measures for Kareius bicoloratus in Tokio Bay Males: L = 37.21 + 56.35 R ( r² = 66.17%) Females: L = -19.52 + 77.15 R (r² = 85.19%) where L is the total length of fish and R is the AORR (Fig. 8) For standar length (SL)-PORL Males: L = 24.38 +65.75 R (r² = 62.01%) Females L= - 7.81 + 83.12 R (r² = 86.51%) where L is the body length of fish and R is the PORL. Finally, the length-weight relationship with the log transformed data was analyzed. The regressións were highly significant and the covariance analysis indicates a rernarkably difference between sexes. The expressions (logarithmic) are as follows: Males log W = 2.96 L -4.83 (r² = 93.73%) Females log W = 3.20 L - 5.38 (r² = 98.80%) where W is total weight, and L is total length. The residual distribution (as in the otolith-length relationshiv) does not show a varticular tendency. 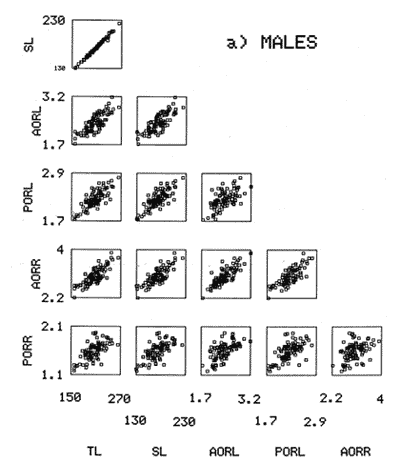 Figure 7a. Draftsman's plot for fish length / otolith radius measures of Karcius bicoloratus males in Tokyo Bay. DISCUSSIONThe stem and leaf and letter values displays, in addit- to the boxplots, lead to the characterization of the difference between the left and right otoliths of stone flounder. The trends and patterns exhibited by these exploratory procedures allow to state some generalities about the left-right otolith morphometry: The overall size and weight of these calcified structures do not differ significantly, but the localization of the focus and certainly the relative growth of the anterior and posterior sides of each otolith are different. The focus of the right otolith is located toward the posterior side, whereas the nucleus of the left otolith has a more central position. Therefore, the line measured from the focus to the posterior side of the right otolith becomes shorter than the radú measured toward the anterior side. By contrast, the anterior and posterior radii measurements of the left otolith are very similar (Fig. 1). 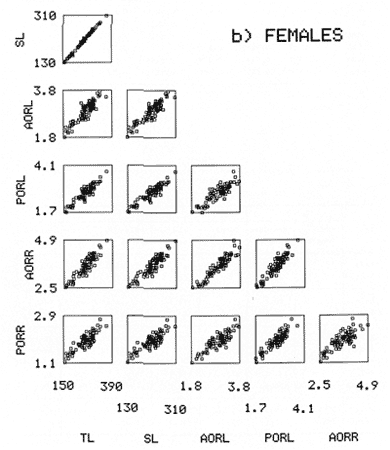 Figure 7b. Draftsman's plot for fish length / otolith radius measures of Kareius bicoloratus females in Tokyo Bay. 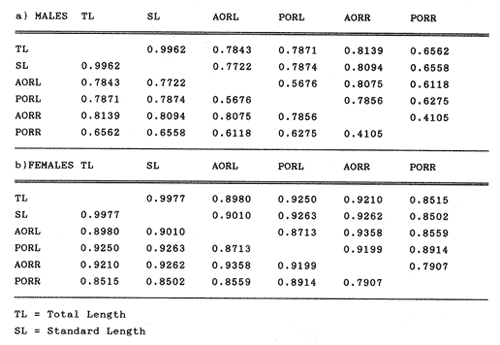 TABLE 4 Correlation matrix for fish tength and otolith raddii measures 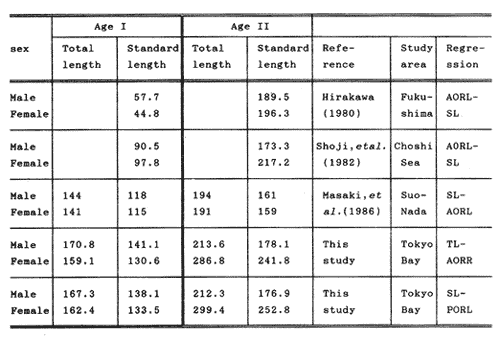 TABLE 5 Comparison of back-calculated lengths from several ageing studies of Kareius bicoloratus (length values in mm) 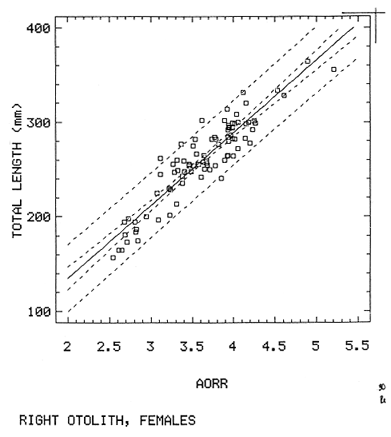 Figure 8a. Regression of total length on anterior otolith radius. 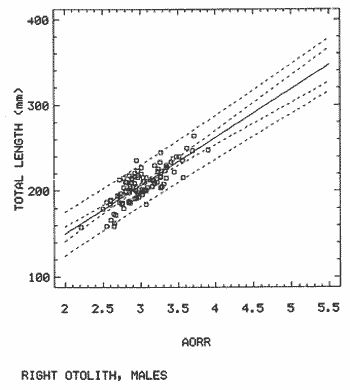 Figure 8b. Regression of total length on anterior otolith radius. Bürgin (1986) describes the syncranial asymmetry of the bastard sole (Microchirus theophila), which otic region gradually becomes asymmetric in a rostral direction. However, a description of the otoliths is not included. From other morphologic studies about the cranial elements of pleuronectids (Holmes and Gibson, 1983; Datta and Das, 1984) it is known that the osteologi- architecture of the blind sidde is best suited for feeding. Furthermore, in spite that in the winter flounder (Pseudopleuronectes americanus) no olfactory organ migration occurs, the olfactory system is asymmetric: the right olfactory organ, nerve and bulb are larger than the contralateral counterparts (Prasada-Rao and Finger, 1984). Brewster (1987) states that the asymmetry of the Pleuronectiformes cranium is caused by the relocation of the anterior blind side frontal to the ocular side, and is accentuated by the enlargement of the blind side lateral ethmoid. The above findings in relation with the asymmetry of the cranial structure and sensorial apparatus suggest that the asymmetry of the ototiths is involved with the structural and sensorial changes related with the particular habit of lying on bottom over one side of the body. In order to calculate the growth parameters for the species, it is important to have representative values for the-otolith ring radii. This means that the estimations are not biased or affected by extremes values. It is a custom to use a weighted average to represent each of the ring radius, but it is known that the mean is affected by spikes in the data. In several instances there were detected outlier values. Some of them probed to be error measurements, but those reported in the results represent the values of non-typical fishes too large or small respect to the other specímens. Thus, in spite of the similarity between the mean and median values, due to the occurrence of outliers, it is advisable to employ the resistant estimators. However, because the age of the sampled fishes is at most two years old, it can not be stated which one is better to estimate the growth parameters of the species. To achieve this it is necessary to obtain older individuals and to compare the growth parameters determined from each data set. It is a custom to employ the left otolith and to measure the anterior radius to calculate the fish lengthotolith radius relationship. In this study, however, the results indicate that the correlation with the anterior side of the right otolith and the one measured toward the posterior edge of the left otolith was higher than that of the pair of measurements usually employed. Certainly the left otolith is elearer under the microscope, and thus the measurements are easy to obtain. Nevertheless, the correlation analysis shows that at least it is preferable to explore several otolith radius measurements and to choose the one showing the closest relationship, with the fish length measures. One interesting characteristic found about the otolith measures is that the highest correlation is presented by the anterior radius of the right side with the posterior radius of the left otolith. This means that these otolith lengths show a closer covariation than those shown by the other measurements. It was expected that the anterior and posterior measures of the left otolith could shown the highest correlation. The length range difference between the male and female samples remarks the sexual dimorphisín and the necessity to obtain a larger size range for the male specimens. The back-calculated lengths from the two regressión equations show a good agreement (Table 5). At age I the males are larger than the females, but from age II this trend is clearly reversed. The comparison of the otolith measurements obtained with those reported in other studies is limited because in several reports different ways to measure the otolith radii were employed. Nevertheless, with the fish length-otolith radius regressión is possible to compare the backcalculated fish-length values (Table 5). In general the lengths calculated in This study are greater than the sizes from other places. This indicates that the stone flounder in Tokyo Bay seems to grow more than in other localities. Another difference between male and female is shown by the length-weight relationship. As in other localities, in Tokyo Bay the specimens of stone flounder with different sex do not have the same growth. However to conclude about the growth of K. bicoloratus in Tokyo Bay it is necessary to obtain a larger sample of both male and female that include older fishes. ConclusionesThe exploratory procedures (stem and leaf, letter values displays and boxplots) madepossible to establish that the focus location of the right otolith is toward the posterior edge of the structure, whereas that of the left one has a more central position, indicating a differential growth between right and left otoliths. These morphological differences and related findings in other flatfish species suggest that this cranial and sensorial structures asymmetry is due to the sideway lying habit of these fishes and the body metamorphosis involved. Due to the presence of some outliers and in order to have unbiased measures of otolith radii and body length measures, it was advisable to employ the resistant median and F-pseudosigma, instead of the mean and standard devíation. The draftsman's plot and correlation analysis showed that is preferable to explore several otolith radius - body length measurements before choosing the higher correlation pairs for growth analysis. Contrary to the custom to employ the anterior side otolith. measurements, the right otolith anterior radius and the left otolith posterior radius showed the highest correlation with the body length measurements and between otolith measures. The body length range difference, the back-calculated sizes and the length-weight relationship pointed out the sexual dimorphism and the growth differences between males and females. At young stages, males are larger, but later, females grow longer. Comparison with the back-calculated lengths from. other localitíes indic- that the stone flounder from Tokyo Bay attains larger sizes. Nevertheless, due to the limitations of available data, a larger male and female older fish sample must be analyzed before conciuding on K. bicoloratus growth in Tokyo Bay. AgradecimientosThe author is very grateful to Dr. Makoto Shimizu, and Dr. Toru Taniuchi, Department of Fisheries of the Faculty of Agriculture, the University of Tokyo, who allow to complete his training on Fisheries Science at the Laboratory of Fisheries Biology, and to Dr. Oscar Sosa-Nishizaki for his scientific and technimical advice. The graduate student Shinji Uehara and Dr. Jun Ohtomi helped a lot to the collection and treatment of samples. The author would like to express his gratefulnees to all the students and Professors of the Laboratory of Fisheries Biology, for their kindness and friendship. Finally, the writer expresses his gratitude to the Japan International Cooperation Agency for the economical support during his training. LITERATURAAKIMOTO, Y., K. ISOGAMI, and T. TAGOSHI, Bull. Fukushima Pref. Fish. Exp.. Stat., (in Japanese). Studies on the ecology of the stone flounder, Kareius bicoloratus III. Resuits on Tagging experiment of the young stone flounder. 1978. 71-79 5: BAGENAL, T. B., and F. W. TESCH, Methods for Assessment of Fish Production in Fresh Waters, Age and growth. In:T.B. Bagenal (Ed.). Blackwell Scientific Pub. Oxford, 1978. pp. 101-136. BHATTACHRYA, C. G., Biometrics, A simple method of resolution of a distribution into gaussian components. 1967. 115-135. 23: BREWSTER, B., J. Fish Biol, Eye migration and cranial development during flatfish metamorphosis: a reappraisal (Teleostei: Pleuronectiformes). 1987. 805-833. 31: BROTHERS, E. B., The Age and Growth of Fish. Methodological approaches to the examination of otoliths in aging studies. In: R. C. Summerfelt and G.E. Hali (Eds.). Iowa State University Press, Iowa, 1987. pp. 319-330. BÜRGIN, T., J. Zool, The syncranial morphology of the bastard sole Microchirus theopitila (Risso, 1810) (Pleuronectiformes, Soleidae). Netherlands, 1986. 117-161. 36 (1): CAMPANA, S. E. and J. D. NEILSON, J. Fish. Aquat. Sci., Mierostructure of fish otoliths. Can. 1985. 1014-1032. 42: CASSIE, R. M., Aust. J Mar. Freshw. Res., Some uses of probability paper for the graphical analysis of polymodal frequency distributions. 1954. 513-522. 5: CHAMBERS, J. M., W. S. CLEVELAND, B. KLEINER and P.A. TUKEY, Graphical Methods for Data Analysis, Wadsworth & Brooks/Cole Pub. Company, Pacific Grove. 1983. 395 p. CURTS, J. B., Biología, El diagrama de tallo y hoja. 1986. 7-12. 15 (1-4): CURTS, J. B., L. L. ALCÁNTARA and X. C. CHIAPPA, Ciencias, Introducción al análisis exploratorio de datos multidimensionales. 1987. 30-35. 11: DATTA, N. C. and M. DAS., Osteology. Cranial anatomy of some Indian Cynoglossids. A study in adaptation. Part I: Zoo. Jb. Anat., 1984. 33-42. 112: EMERSON, J. D. and D. C. HOAGLIN, Understanding Robust and Exploratory Data Analysis, Stem-and-leaf displays In: D. C. Hoaglin, F. Mosteller and J. W. Tukey (Eds.). John Wiley & Sons, New York, 1983. pp. 7-32. EMERSON, J. D. and J. STRENIO, 1983. Understanding Robust and Exploratory Data Analysis, Boxplots and batch comparison. In: D. C. Hoaglin, F. Mosteller and J.W. Tukey (Eds.). John Wiley & Sons, New York, 1983 pp. 58-96. ESCHMEYER, W. N., E. S. HERALD and H. HAMMANN, A Field Guide to Pacific Coast Fishes of North America, Houghton Mifflin Co., Boston, 1983. pp. 283-294. FUJITA, K. J Ichthyol., A reversal ambicolorate flounder, Kareius bicoloratus, caught from Tokyo Bay. Japanese. 1980. 175-178. 27 (2): HAMILTON, L. C., Statistics with Stata, Brooks/Cole Pub. Company, Pacific Grove 1990 172 HARDING, J. P., J. Marine Biol. Ass. UK, The use of probability paper for the graphical analysis of polymodal frequency distributions. 1949. 141-143. 28: HASSELBLAD, V., Technometrics, Estimation of parameters for a mixture of normal frequency distributions. 1966. 431-444. 8 (3): HATANAKA, M., K. SEKINO and A. OSUKA, Studies on the populations of the flatfishes in Sendal Bay. II. Age, growth and spawning of Karcius bicoloratus (Basilensky). J Agr. Res., Tohoku, 1952. 2. HIRAKAWA, H., Bull. Fukushima Pref Fish. Exp. Stat, (in Japanese). Studies on the fish populations of small nets traw1 in Fukushima - I. Age, and growth of stone flounder after recruitment. 1980. 1-10 6: HOAGLIN, D.C., Understanding Robust and Exploratory Data Analysis, Letter values: a set of selected order statistic In: D. C. Hoaglin, F. Mosteller and J.W. Tukey (Eds.). John Wiley & Sons, New York, 1983. pp. 33-57. HOAGLIN, D.C., F. MOSTELLER and J.W. TUKEY Understan ding Robust and Exploratory Data Analysis, HOAGLIN, D.C., F. MOSTELLER and J.W. TUKEY (Eds.), John Wiley & Sons, Inc. New York. 1983. 447 p. HOLMES, R. A. and R.N. GIBSON, Anim Behav., A comparison of predatory behavior in flatfish. 1983. 1244-1255. 31: IGLEWICZ, B., Understanding Robust and Eyploratory Data Analysis, Robust scale estimators and confidence intervais for location. In: D.C. Hoaglin, F. Mosteller and J.W. Tukey (Eds.). John Wiley & Sons, New York, 1983. pp. 404-431. MCGILL, R., J.W. TUKEY and W.A. LARSEN, The American Statistician, Variations of box plots. 1978. 12-16. 32: MINAMI, T. Bull. (in Japanese, with English summary). The carly life story of a flounder Kareius bicoloratus. Soc. Sci. Fish., Japan. 1984. 551-560 50 (4): MITO, S., M. UKAWA and M. HIGUCHI, Bull. Nansei Regional Fisheries Research Laboratory, (in Japanese, with English summary). On the egg development and rearing of the larvae of a flounder, Kareius bicoloratus (Basilewsky) with reference to its spawning in the culturing pond. 1969. 86-102 1: OCHIAI, A. and M, TANAKA, Shinpan Gyoruigaku Kouseisha Kousei kaku, (in Japanese). Japan 1986. pp. 1099-1102. PANNELLA, G., Science, Fish otoliths: daily growth layers and periodical pattems. 1971. 1124-1127. 173: POLICANSKY, D., Scientific American, The asymmetry of flounders. 1982. 96-102. 246 (5): PRASADA-RAO, P. D. and T. E. FINGER Journal Comp. Neurol., Asymmetry of the olfactory system in the brain of the winter flounder, Pseudoamericanus. 1984. 492-510. 225 (4): SHOJI, Y., K. MEGURO and M. ITO, Chiba Ken Suisan Shikenjyo Kenkyn Houkoku, (in Japanese). The growth and maturity of stone flounder in Choshi Sea. 1982. 67-74 40. SNEDECOR, G.W. and W.G. COCHRAN, Statistical Methods, Iowa State University Press, Iowa. 1980. SOKAL, R. R. and F. J. ROHLF, Biometry, W.H. Freeman, San Francisco. 1981. TAKAGOSHI, T. and Y. AKIMOTO, Bull. Fukushima Pref. Fish Exp. Stat.,(in Japanese). . Studies on the ecology of the flatfish Karcius bicoloratus - I. Growth and habitat of the fingerling group. 1975 41-50. 3: TAKAGOSHI, T., K. ISOGAMI, Y. AKIMOTO and A. TENJIN, Bull. Fukushima Pref. Fish. Exp. Stat., (in Japonese). Studies on the ecology of the flatfish Kareius bicoloratus II. On the distribution of fry and the environmental condition on the coast of Fukushima Prefecture. 1976. 33-40 4: TANAKA, S., Taius tumifrons A method of analysing a polymodal frecuency distribution and íts application to the length distribution of the porgy, (T. and S.). J Fish. Res. Bd Can., 1962. 1143-1159. 19. TSURUTA, Y., J. Agric. Res., Field observations on the inmigration of larval stone flounder into the nursery ground. Tohoku, 1978. 136-145. (3,4): 29 TUKEY, J.W., Exploratory Data Analysis, Addison-Wesley, Reading. 1977. 688 p. VELLEMAN, P. F. and D.C. HOAGLIN, Applícations, Basics, and Conpulting of Exploratory Data Analysis, Duxbury Press, Boston. 1981. 354 p. WILLIAMS, T. and B.C. BEDFORD, The Ageing of Fish. The use of otoliths for age deterinination. In: T. B. Bagenal (Ed.). Unwin Brothers Ltd. England 1974. pp. 114-123.
|