|
SINOPSIS DE LA DINÁMICA POBLACIONAL DE CINCO
ESPECIES DE LA FAMILIA LUTJANIDAE EN LA COSTA DE YUCATÁN
Trabajo
recibido el 1 de agosto de 1990 y aceptado para su publicación el 9 de
julio de 1991.
Ricardo Torres-Lara
Delegación Federal de Pesca en
Yucatán. Calle 65 No. 627 x 76 y 78. Mérida, Yucatán
97000, México.
Silvía Salas-Márquez, Guadalupe Mexicano-Cíntora y Miguel A. Cabrera-Vázquez.
CINVESTAV-INP-Unidad Mérida. Apartado Postal
73, Cordemex. Mérida, Yucatán 97310, México.
Se presenta una sinopsis de la dinámica
poblacional, realizada en la costa de Yucatán, de algunas especies de la
familia Lutjanidae. Se incluyen resultados preliminares en relación a
parámetros de crecimiento, tasas de mortalidad y evaluaciones generales
acerca del grado de explotación al que estuvieron sujetas durante la
temporada de pesca en que se realizaron los estudios. Los resultados indican
que las especies analizadas están sometidas a grados diversos de
aprovechamiento, que en algunos casos se relacióna con la inactividad de
la flota arrastrera de la pesca de escama en dicho período.
A synopsis of the population dynarnics aspects of
some Lutjanidae species in Yucatan coast is presentes. Preliminary results in
relation to growth parameters, mortality rates and general evaluations on the
magnitude of exploitation in these species through fishing season, are
included. In general, these studied species show different exploitation levels.
This behaviour can be associated, in some cases, to the period in which the
trawler fleet of the scale fishery does not operate.
El aprovechamiento de los recursos naturales es una de las actividades humanas más antiguas, aunque sólo recientemente se ha realizado con base en estudios que permiten conocer las fluctuaciones de la abundancia de tales recursos en la dimensión temporal. En relación a los productos pesqueros, el estudio de su dinámica poblacional es un medio primordial para conocer los cambios naturales y los introducidos por el hombre cuando están sujetos a extracción. La utilidad práctica de calcular las fluctuaciones de la población íctica con respecto al tiempo se basa en el hecho de poder realizar predicciónes acerca de posibles rendimientos pesqueros en temporadas subsecuentes de pesca. Estudios de dinámica poblacional son necesarios pero no suficientes para alcanzar este objetivo, pues el entendimiento de los procesos biológicos proporciona un conocimiento más amplio y consecuentemente mayor consistencia en las estimaciones que se realicen sobre los rendimientos de pesca. En el estado de Yucatán se han efectuado numerosos trabajos para conocer la dinámica poblacional de diversas especies de interés comercial. Dentro de éstas, destacan algunas pertenecientes a la Familia Lutjanidae, conocidas genéricamente como pargos. En el presente estudio se hace una revisión de los trabajos que en esa zona y acerca de ese tema se han realizado del pargo criollo (Lutjanus analis), el pargo mulato (Lutjanus griseus), el pargo canané (Ocyurus chrysurus), la rubia (Lutjanus synagris) y el huachinango (Lutjanus campechanus); esta sinopsis representa una diagnosis preliminar de estas especies en cuanto al conocimiento de sus parámetros de crecimiento, tasas de mortalidad y evaluación del nivel de explotación que soportaban durante la temporada de pesca en que se realizaron los muestreos respectivos. ANTECEDENTESLos peces de la Familia Lutjanidae se encuentran ampliamente distribuidos tanto en aguas tropicales como subtropicales; se les localiza desde las aguas protegidas someras hasta profundidades de 640 m. Se asignan a esta familia cerca de 30 géneros y 150 especies, la mayoría de las cuales viven asociadas al fondo y se encuentran confinadas a la plataforma y al talud continentales. La mayoría de las fases larvarias tempranas son todavía desconocidas (Druzhinin, 1970). Su distribución geográfica abarca desde el norte de Massachusetts hasta el sur de Brasil, incluidas las Islas Bahamas, Islas Bermudas y otras islas del Mar Caribe (Thompson y Munro, 1974). Entre los primeros trabajos conocidos sobre especies de esta familia destacan el de Crocker (1962), quien determinó la edad en L. griseus; el de Moseley (1966), quien realizó un estudio biológico de L. aya; se enfocó principalmente a temas como el crecimiento, la edad y la alimentación; de los trabajos más completos de este período, cabe menciónar el elaborado por Stark y Schröder (1971), quienes realizaron un estudio bastante detallado de L. griseus, y proporciónan información referente a otras especies de la misma familia. A partir de estas primeras investigaciones, surge una serie de estudios relativa a los lutjánidos de otras regiones, especialmente Cuba, Venezuela y Brasil. Sin embargo, no es sino hasta 1979 cuando Contreras realiza un estudio muy somero acerca de la rubia, L. synagris; inicia así el período actual de trabajos, por parte de investigadores mexicanos, relacionados con estos recursos en el litoral yucateco, tema central de este trabajo. Por otro lado, las principales pesquerías que en el estado de Yucatán han contribuido al desarrollo de la actividad pesquera, considerando las características de artes, equipo y método de captura, son: la de línea y la de arrastre. La primera es la que aporta los mayores volúmenes de captura por ser la más practicada, y porque el arte que utiliza comprende el palangre y las líneas de mano escamera y pulpera. Mediante estas artes se capturan el mero y el pulpo, las cuales aportan más del 60% de la captura total en el Estado (Departamento de Desarrollo Pesquero). La pesquería de arrastre trabaja con redes que operan sobre el sustrato suave, que puede o no tener vegetación. Esta pesquería fue muy importante durante el período de 1980-1983, ya que se incorporaron a la flota arrastrera embarcaciones mayores, con capacidad de bodega de 183 toneladas. Los registros de captura de especies de las Familias Serranie Lutjanidae y Sparidae aumentaron considerablemente; sin embargo, estas embarcaciones se desincorporaron de la flota yucateca en 1984, con el consiguiente descenso en la tasa de aprovechamiento de esos recursos (Torres, 1987). Los pargos mulato, criollo, canané y la rubia han sido tradiciónalmente extraídos mediante línea de mano escamera y palangre, aunque también lo fueron por los barcos arrastreros cuando éstos operaron en las costas yucatecas; por otro lado, al huachinango lo capturan embarcaciónes escameras, así como mereras y huachinangueras (Burgos y López, 1985, citados por González, 1988). Aunque se ha sugerido que las naves que están equipadas con bicicletas, que permiten operar una línea vertical hasta con cinco anzuelos en el extremo, pueden identificarse como unidades de pesca para la captura de huachinango, en general no es posible determinar una flota que ejerza un esfuerzo de pesca específico sobre cada uno de estos recursos. MÉTODOSEl procedimiento que se utilizó para recabar y analizar la información consistió básicamente en la investigación documental. El enfoque se orientó hacia los estudios que acerca de dinámica de poblaciones se han realizado de los desembarques de la captura comercial que se lleva a cabo en puertos del litoral yucateco. La suposición a priori que se deriva de estos elementos es que la flota del Estado opera en la misma área (Banco de Campeche) y que ejerce su actividad sobre una sola población de cada una de las especies capturadas, independientemente de las relaciones que existan entre tales especies. Se utilizó la información prevíamente generada por varios autores en relación a la estimación de los parámetros poblacionales y a su variación temporal en los casos en que fue posible hacerlo. Cabe mencionar, sin embargo, que algunos de ellos consideraron someramente otros temas del ciclo de vida de las especies, especialmente la fecundidad y la alimentación, como son los trabajos de Torres (1987), Torres y Salas (1989) para la rubia, y Garduño y López (1985) para el pargo canané. Dentro de este marco de trabajo, se recopiló la información referente a: 1) parámetros de crecimiento en longitud: L∞, K y to; 2) parámetros de la relación peso-longitud y determinación del peso asintótico (W∞; 3) tasas de mortalidad total (Z), natural (M) y por pesca (F); 4) tasa de extracción (E); 5) relación F/Z, como evaluación práctica de la intensidad de pesca; y 6) en los casos en que se aplicó el Modelo de Beverton y Holt (1957), cálculo del rendimiento por recluta. Se hace referencia a los casos en que se calcularon las tasas de mortalidad a partir de la información contenida en estos estudios mediante aplicación de métodos adecuados a tal información, con el fin de contar con mayores elementos para efectuar el análisis entre las especies consideradas. Por último, se incluye la comparación de los valores de dichos parámetros entre las cinco especies mencionadas para más de una temporada, cuando fue el caso. Esta investigación finaliza con una serie de conclusiónes y recomendaciónes como aportación para la elaboración de trabajos futuros acerca de dinámica de poblaciones, especialmente de los recursos pesqueros de las costas de Yucatán. RESULTADOS Y DISCUSIÓNCon base en los estudios revisados, la información recabada se concentró en tablas, donde se observa el valor de los parámetros por especie y se indican los autores. CRECIMIENTOEn la tabla 1 se presentan los párametros de crecimiento L∞, K y to. La relación más evidente se da entre los valores de la tasa de crecimiento y la longitud asíntótica; esta relación es inversa, pues tanto a L. campechanus como a L. analis, que son las especies que alcanzan las tallas mayores, corresponden los valores más bajos de K; por otro lado, las especies de tamaños asintóticos menores, como O. chrysurus y L. synagris, tuvieron los valores más altos de K. Considerando el patrón antes mencionado, los casos atípicos son los del pargo mulato (L. griseus) y el canané, (O. chrysurus), reportados por Cantarell (1982). Para el primero, se obtuvo el valor L∞ bajo, si se toma en cuenta que esta especie es semejante a L. analis y a L. campechanus, las cuales han sido registradas con tallas mayores en las capturas comerciales. Por otro lado, para O. chrysurus se tiene un valor bajo de K si se compara con los valores obtenidos por los otros autores para la misma especie. Las causas posibles que provocaron esta aparente discrepancia, con relación al patrón general, se pueden resumir como sigue: a) La muestra no fue representativa de la población, ya sea porque el tamaño de la muestra no haya sido adecuado o porque se obtuvieron muestras sesgadas, que no corresponden a la distribución real de los organismos en la integración poblacional, es decir dentro de un intervalo limitado de tallas, al depender de la captura comercial. b) Errores de procedimiento en el cálculo de los parámetros. Esto se da en el caso de que los métodos utilizados son muy subjetivos y, en consecuencia, un mismo procedimiento puede aportar resultados distintos al aplicarlo personas diferentes. Además, el hecho de que algunas veces se aplican métodos sin considerar sí el recurso analizado cumple a suposiciones pertinentes. 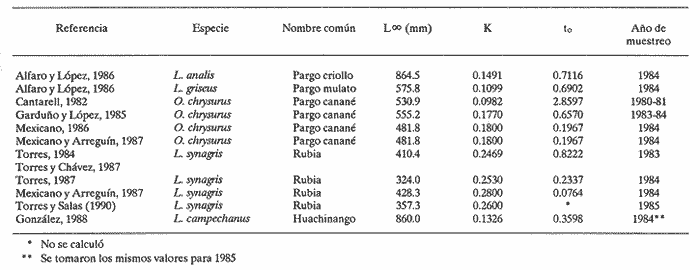 TABLA 1 Parámetros de crecimiento de las especies analizadas de la familia Lutjanidae: longitud asintótica (L8), tasa de crecimiento (K) y longitud al tiempo cero (to) En cualquier caso, lo importante es señalar que, en términos generales, se cumple la relación que Pitcher y Hart (1982) habían señalado entre estos dos parámetros, en el sentido de que el valor de K puede tomarse como medida del tiempo en el que se alcanza la longitud asintótica, de tal manera que, con una tasa de crecimiento acelerado, la talla máxima se alcanza rápidamente y, por consiguiente, ésta es menor que la lograda por especies de crecimiento más lento. Observando los valores de K aquí tabulados, es posible afirmar que en estas especies el gradiente de velocidad de crecimiento, de lento a rápido, se ordena como sigue: pargo mulato, huachinango, pargo criollo, canané y rubia. Por otra parte, este gradiente tiene un significado desde el punto de vista ecológico: especies con valor alto de la tasa de crecimiento se caracterizan por ser depredadores muy activos, que requieren un gasto energético fuerte para capturar otros peces; este hecho los coloca en un nivel trófico elevado; tales podrían ser los casos de la rubia (Torres, 1987) y del canané (Garduño y López, 1985). En el otro extremo, especies bentónicas de nivel trófico menor son de crecimiento lento, lo cual sugiere especies de poca movilidad cuyas presas son preferentemente crustáceos y moluscos, como el pargo mulato (Alfaro y López, 1986). Por último, no se discuten con mayor amplitud las diferencias en los valores del parámetro to, puesto que representa sólo un factor de ajuste de la curva generada por el Modelo de von Bertalanffy. En relación a ésto, Moreau (1987) explica que no es necesario calcular un valor exacto de dicho parámetro para algunas evaluaciones: rendimiento por recluta, uso de curvas de captura (en longitud) y análisis de cohortes (en longitud). Sin embargo, es importante señalar que algunos autores lo consideran indicador de la variación de la temporada de desove (Basson et al., 1988). RELACIÓN PESO-LONGITUDUna de las derivaciónes de los estudios de crecimiento es la relación peso-longitud (W = aLb ), a partir de la cual es posible definir el crecimiento individual en el peso asintótico (W ∞. Este último es importante debido a que se incorpora al Modelo de Beverton y Holt (1957) para calcular el rendimiento por recluta en biomasa. El valor de a puede variar estacionalmente según la alimentación, la época reproductiva, diariamente o el habitat. En cuanto al exponente b, si éste es igual a 3, el crecimiento es isométrico, es decir, las proporciones geométricas del cuerpo no cambian. Si la forma del cuerpo cambia durante el crecimiento de modo tal que la relación de las medidas lineales con respecto al peso decrece, entonces b es mayor que 3; en caso contrario, b es menor que 3. Esto sucede en la mayoría de las especies de peces y se afirma que su crecimiento es alométrico.(Bagenal, 1978). Como se observa en la tabla 2, los valores encontrados para b en los trabajos revisados son muy cercanos a 3 en L. campechanus, L. analis y L. griseus, por lo que es posible afirmar que el crecimiento de estas tres especies es isométrico. En los otros casos el valor de b es menor, excepto en uno reportado por Garduño y López (1985) para O. chrysurus, que es el más cercano a 3. Una consideración sumamente importante cuando se trabaja con esta relación es la necesidad de especificar la época de toma de los datos (peso y longitud), y el intervalo de ambas variables dentro del cual se calcularon los valores de a y de b, por lo que estos valores son válidos sólo para la época y los intervalos de las menciónadas variables en que fueron tomados, puesto que esta relación es particular para cada estadio durante el desarrollo ontogenético de las especies, para cada sexo, y para cada estación del año, debido principalmente a los ritmos y cambios en la alimentación y a las épocas de reproducción, todo lo cual influye en. las variaciones temporales de la biomasa de la población en estudio. No es válido asignar los valores de a y b de un momento particular a las condiciones de otro instante (Winberg, 1971; Bagenal, op. cit.). Dado que estos parámetros se utilizaron para determinar el valor del peso asintótico y posteriormente el rendimiento por recluta para el canané (Mexicano-Cintora y Arreguín-Sánchez, 1987), para la rubia (Mexicano-Cíntora y Arreguín-Sánchez, op. cit.; Torres, 1987 y Torres y Chávez, 1987), y para el huachinango en 1985 (González, 1988), estos cálculos deben tomarse con reserva. MORTALIDADDentro del conjunto de conocimientos necesarios acerca de la dinámica de poblaciones para gestión adecuada de una pesquería, el cálculo de la mortalidad es de importancia primordial, ya que si no es apropiado, la evaluación del rendimiento no es confiable y tampoco las proposiciónes de administración pesquera derivadas de ésta. En todos los casos la determinación del valor de Z corresponde a la composición por grupos de edad de los peces capturados (curvas de captura). Como se observa en la tabla 3, la especie de mayor tasa de mortalidad total fue L. synagris, en 1983 (Torres, 1984) y en 1984 (Mexicano- Cíntora y Arreguín-Sánchez, 1987), con valores de Z = 1.1545 y Z = 1.2450, respectivamente. Las determinaciones de Z realizadas por el primer autor revelan un descenso paulatino de 1983 a 1985, pero señala que esto se atribuye a una reducción en la intensidad de operación de la flota arrastrera yucateca (Torres y Salas, 1990). En la misma tabla se observan los valores conocidos de la mortalidad por pesca de esta especie. 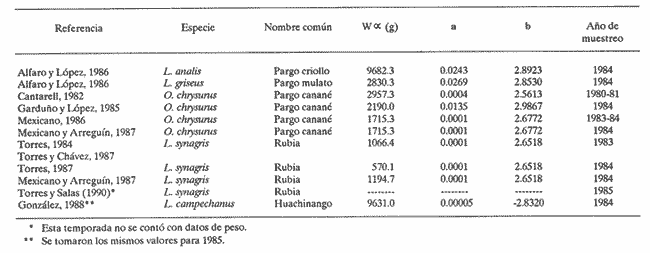 TABLA 2 Parámetros de la relación peso-longitud (a y b) y peso asintático (W (infinito) de las especies analizadas de la familia Lutjanidae 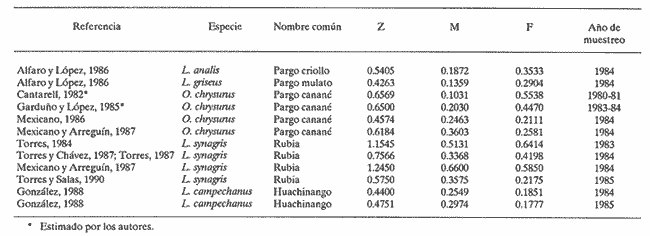 TABLA 3 Tasas de mortalidad total (Z), natural (M) y por pesca (F) de las especies analizadas de la familia Lutjanidae El mismo panorama se observa para el canané, ya que en 1980-1981 su mortalidad total era de Z = 0.6184 (Mexicano-Cíntora y Arreguín-Sánchez, 1987) y Z = 0.4574 (Mexicano, 1986). Este hecho puede deberse a la misma reducción en el esfuerzo pesquero explicado para la rubia, pues ambas especies se capturan con el mismo arte de pesca, lo cual se confirma observando los valores de la mortalidad por pesca para O. chrysurus en la tabla 3. En el caso del huachinango, la mortalidad total no varió en las dos temporadas estudiadas por González (1988). Análisis comparativos de esta índole no se pueden realizar para los pargos criollo y mulato, pues sólo se tiene la referencia de un solo año para cada especie; sin embargo, es posible observar en la misma tabla que los valores de sus respectivas tasas de mortalidad total y por pesca se sitúan entre los correspondientes a la rubia y al canané en 1984 (Mexicano, 1986; Torres y Salas, 1990), ya que ese año, al menos para el caso de F, estas cuatro especies soportaron la misma intensidad de extracción. Los valores de la mortalidad natural en todos los casos, salvo para los pargos mulato y criollo (Alfaro y López, 1986), se obtuvieron mediante la ecuación empírica propuesta por Pauly (1980), en la que se consideran los parámetros de crecimiento de cada especie, así como la temperatura promedio de su habitat de residencia. Sin embargo, González (op. cit.) también aplicó el método ANPOVIR (Arreguín- Sánchez y Chávez, 1985) y aceptó como válidas las cifras calculadas por medio de este modelo, el cual se basa en la abundancia de los reclutas y del último grupo de edad para realizar tal cálculo. Como se observa en la tabla 3, el valor de M para L. campechanus en 1985 es mayor (M= 0.2974) que el correspondiente para 1984 (M = 0.2549). González (op. cit.) mencióna que las diferencias en los valores de M entre los años son consecuencia de la diferencia cuantitativa de individuos en la última edad registrada en peces capturados. Al respecto, se encontró relación directa entre la tasa de crecimiento y la mortalidad natural; es decir, valores pequeños de K se asocian con valores pequeños de M, por ejemplo en el caso del pargo canané (Cantarell, 1982): K= 0.0982 y M= 0. 1031, ya que si a una especie corresponden valores bajos en la tasa de crecimiento, indican longevidad comparativamente mayor. Por otro lado, especies con cifras altas de K y M indican longevidad menor, como en el caso de la rubia (Mexicano- Cíntora y Arreguín-Sánchez, 1987), con K = 0.28 y M = 0.66. En cuanto a la mortalidad por pesca, ésta se determinó por diferencia de las dos anteriores (F=Z-M). Sólo para el caso de la rubia se mencionó que variaciones en el modo de operación de intensidad de la flota provocaron valores distintos, que se reflejaron en los valores correspondientes de la mortalidad total (Torres y Salas, 1990). Puesto que la mortalidad por pesca depende tanto de la abundancia del recurso como del esfuerzo de pesca aplicado, este último está supeditado a varios factores, como: eficiencia o poder de pesca del equipo y artes empleadas. Cualquier recomendación que se haga acerca de cambios en intensidad extractiva debe estar precedida por un estudio que permita conocer: el intervalo de tallas sometido a la selección del arte, la vulnerabilidad de los organismos, la proporción de peces retenida por grupo de talla, la talla y la edad de primera captura (Pereiro, 1982, citado por Garduño y López, 1985), la intensidad extractiva a la que está sometida la especie, y la capacidad de pesca (Caddy, 1979). RENDIMIENTOSe puede obtener una evaluación preliminar del grado de explotación del recurso, mediante el conocimiento de sus respectivas tasas de mortalidad y de extracción [E = F/Z (1-S)] y proporción de la mortalidad total atribuida a la pesca (F/Z). Doi (1976) mencióna que cuando ésta es aproximadamente igual a la mortalidad natural, el valor de la relación F/Z es cercano a 0.5, lo que significa que aún no se ha afectado la tasa de renovación natural de la población, y se encuentra en su nivel óptimo de explotación (la misma conclusión se encuentra implícita en el desarrollo del Modelo ANPOVIR, propuesto por Arreguín-Sánchez y Chávez en 1985). Bajo el criterio antes expuesto, los datos puntuales referentes a los pargos criollo y mulato (Alfaro y López, 1986) revelaron que en 1984 (año en que se colectaron los datos con los que se realizó el estudio) las poblaciones de estas especies estaban sometidas a extracción excesiva. Para los casos del canané y de la rubia se observa una tendencia descendente (1980-1981) en el valor de F/Z, es decir, se llegó desde un estado en el que la extracción es excesiva hasta otro en que tiende a que sea óptima, lo que podría explicar la disminución en la operación de la flota arrastera, pues ambas especies se capturan con el mismo método de pesca. Con respecto al huachinango, se obtuvo la misma tendencia en los valores de dicha relación, es decir, en el transcurso de las temporadas de pesca se redujo la proporción de la mortalidad total debida a esta actividad. Así, de F/Z = 0.4207 en 1984, se reduce a un valor de F/Z = 0.3740 en 1985 (González, 1988). Un método más elaborado para conocer el grado de explotación real e identificar las condiciones de pesca bajo las cuales se logra el rendimiento óptimo (expresado en gramos por recluta), es el Modelo de Beverton y Holt (B-H). Este modelo considera constante la cantidad anual de reclutas, de tal manera que el rendimiento resulta como interacción de la mortalidad natural y por pesca, y del crecimiento. Así mismo, es importante hacer notar que la existencia de este rendimiento máximo como función de la mortalidad por pesca depende de la edad de reclutamiento, y esto a su vez de las propiedades selectivas del arte de pesca utilizado y de las características de los individuos. En la tabla 4, para el pargo criollo el Modelo de B-H predice un rendimiento óptimo de 1,100 gramos por recluta en 1984, equivalente a 22.4 toneladas en la población, con F = 0.6500, y longitud de reclutamiento de 495 mm (tr = 5); sin embargo, bajo las condiciones de explotación de ese mismo año (F = 0.35 y tr = 4) la captura fue de 26.6 toneladas. Es decir, con F y tr mayores se obtendrían rendimientos menores a los obtenidos ese año. Si se considera la relación F/Z antes expuesta, que indica condiciones superiores al nivel óptimo para esta especie, coincide con el planteamiento de este modelo, lo cual indicaría que el Modelo de B-H puede predecir un rendimiento óptimo, aun a costa de elevar el nivel de explotación del recurso. En el caso del pargo mulato, los cálculos indicaron un rendimiento óptimo de 22.11 toneladas (400 g/recluta), con F = 0.95 y Ir = 377 (tr = 8); en realidad, en el año de referencia (1984) la captura fue de 17.78 toneladas, con F = 0.3 y tr = 4. En este caso se obtuvo el 80% del nivel óptimo determinado mediante el modelo, lo cual motivaría un análisis más detenido para decidir si convendría incrementar la actividad pesquera en 200% para lograr una ganancia del 20%, que agregaría el riesgo de agudizar la extracción excesiva pronosticada desde la determinación del valor de F/Z para ambas especies. Para el pargo canané se debe apuntar que tanto Cantarell (1982) como Garduño y López (1985) no realizaron cálculos del rendimiento. Por otro lado, Mexicano (1986) encontró que aumentando la mortalidad por pesca de F = 0.21 a F = 0.56, así como la longitud de reclutamiento, lr = 256.1 mm (tr = 4) a lr = 284 mm (tr = 5) se obtendría, para 1984, un rendimiento de 140 g por individuo; mientras que Mexicano Cíntora y Arreguín-Sánchez (1987) calcularon que con lr = 260 mm y elevando F = 0.21 a F = 0.40, el rendimiento por recluta sería de 50 g. Por otro lado, estos resultados conducen a la conclusión de que en ambos casos el cálculo del grado de extracción del recurso corresponde a una ligera sobre explotación, pero al mismo tiempo permiten hacer diferentes cálculos que se pueden utilizar para definir qué métodos se usaron para el procesamiento de tales datos. Como ya se había mencionado, también es importante no tomar los valores de parámetros de un año determinado para aplicarlos en la misma especie en otra temporada de pesca. 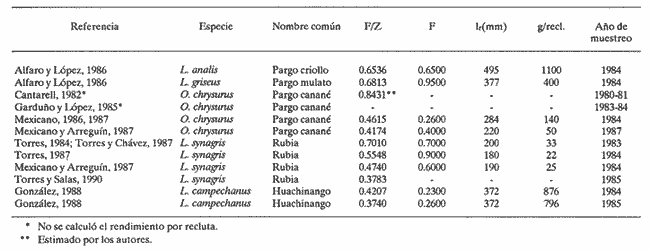 TABLA 4 Pronóstico de las condiciones de explotación, mortalidad por pesca (F) y longitud de primera captura (Ir) del rendimiento óptimo, expresado en gramos por recluta, según el Modelo de Beverton y Holt Ahora bien, en 1983 los datos reales relativos a la rubia fueron lr = 247 mm (tr = 3) y F = 0.6114, y se logró un rendimiento de 28 g por recluta. Las condiciones óptimas de explotación habrían sido lr = 200 mm y F = 0.7 para un rendimiento de 33 g. Como se puede apreciar, ese año el nivel se encontraba cercano al óptimo. Para 1983 la situación cambió, ya que se redujo la intensidad en el modo de operación de la flota arrastrera y por lo tanto la mortalidad por pesca bajó a F=0.4198; al mismo tiempo, la longitud de primera captura registrada fue de lr=230 mm; mediante la aplicación del modelo se determinaron F = 0.9 y lr = 180 mm, con lo cual se lograría un rendimiento de 22 g/recluta. Ese año la mortalidad por pesca fue menor al valor calculado en condiciones óptimas por el modelo, con longitud de reclutamiento lr = 180 mm, igualmente menor, lo cual sugiere subutilización del recurso. Por otro lado, Mexicano-Cíntora y Arreguín-Sánchez (1987) encontraron que, para la misma especie, durante 1984 las condiciones bajo las cuales se alcanzaría el rendimiento máximo, de 25 g por recluta, corresponden a F = 0.6 y lr= 240 mm, que comparadas a las condiciones reales (F = 0.66, lr = 240 mm y rendimiento de 23 g) no se obtendrían grandes beneficios reduciendo en 50 mm las tallas de primera captura. De cualquier modo, el nivel sería de subexplotación. El estudio realizado por Torres y Salas (1990) no incluye cálculo del rendimiento pesquero correspondiente a 1985. En el caso del huachinango, se había detectado desde la determinación de los valores de F/Z que en 1984 y 1985 el recurso se encontraba subexplotado. Referente a 1985, el Modelo de B-H pronosticaba un rendimiento óptimo de 876 g/recluta, con F = 0.2600 y tr = 4; bajo estas condiciones, el recurso se encontraría cercano a la condición de aprovechamiento óptimo. Es conveniente hacer dos consideraciones importantes en relación con el cálculo del rendimiento óptimo mediante el Modelo de Berverton y Holt. La primera se refiere básicamente al tipo de pesquería a la que se aplica: el modelo tiene limitaciónes, pues las poblaciones de las especies con las cuales se trabajó pertenecen a una comunidad que interactúa entre sí y con su medio, y se extraen en conjunto por un arte de pesca, lo cual hace difícil la aplicación de las recomendaciones derivadas de los resultados para una especie porque necesariamente ejercen algún efecto sobre todas y cada una de las poblaciones restantes. Las pesquerías que se desarrollan en las costas del estado de Yucatán son en escencia multiespecíficas; aunque en algunos casos existe una especie-objetivo para el arte empleado, éste no es totalmente selectivo y no discrimina entre esta especie y las que cohabitan con ella. Por esta razón la reglamentación que se proponga para cada especie debe basarse en estudios que incluyan las interacciones interespecíficas a las que se ha hecho referencia. La segunda consideración de importancia se refiere a las propias hipótesis del modelo. Se reconoce que considera el nivel de reclutamiento y la mortalidad natural como constantes; empero, es muy probable que estos factores varíen en las diferentes especies y con el trancurso del tiempo, o de acuerdo con los cambios en la intensidad de pesca que pueden conducir a niveles de sobre-explotación, aunque se logren rendimientos mayores que los obtenidos mediante las condiciones reales de cada temporada. ConclusionesLa eficiencia de una pesquería depende en gran medida del mayor conocimiento que se tenga de los recursos que se están explotando. Este conocimiento involucra los procesos biológicos que se desarrollan de manera natural en las poblaciones ícticas que, en condición de equilibrio, permiten aprovechar determinada biomasa de la población mediante la actividad pesquera, sin que se le afecte. Así mismo, de alguna manera el cálculo de los parámetros poblacionales refleja dichos procesos y éstos a su vez muestran el estado de aprovechamiento de los recursos en el momento de la extracción. En este sentido se han orientado los trabajos aquí revisados, pues a partir de ello se generó información valiosa que podrá utilizarse como base para el planteamiento de nuevos planes de investigación a futuro. Eventos como el crecimiento y la mortalidad han servido para caracterizar a las diferentes especies estudiadas, de acuerdo con los valores de los parámetros poblacionales incorporados en los modelos matemáticos que describen y representan la dinámica del recurso. Por otra parte, es necesario tomar en cuenta las limitaciones y suposiciones de los modelos en los cálculos que se deriven de éstos, sobre todo en el momento de hacer inferencias y proponer planes de aprovechamiento de un recurso dado. Por lo tanto, los resultados deben tomarse como preliminares hasta tener conocimiento más amplio y profundo del comportamiento de la dinámica poblacional de las especies estudiadas, lo cual se logrará en la medida en que se establezca una continuidad en las investigaciones que de ellas se estén realizando y de esta manera asegurar que no queden como hechos aislados, ya que para algunos casos no se cuenta con información más que para sólo un año; en otros se han obtenido distintos análisis de diferentes autores a partir de los mismos datos y en general no se han continuado estudios acerca de estas especies, excepto los aquí presentados. Por ello, se considera necesario contar con un programa sistemático y continuo de trabajos relativos a dinámica poblacional y a biología básica de éstas y otras especies, que por su importancia económica y por su papel dentro de la estructura y la función del ecosistema en que habitan, ocupen una situación relevante dentro del esquema multiespecífico de las pesquerías que se practican en las costas de Yucatán, que se puedan complementar con información socioeconómica para tener un esquema más completo de éstas. Otro asunto de suma importancia es la necesidad de difundir los resultados de cada investigación en el menor tiempo posible, pues considerando la dinámica poblacional, que se potencia en función de la intervención del hombre, las proposiciones de regulación pesquera van perdiendo, validez a medida que pasa el tiempo, debido al cambio constante de las condiciones naturales con referencia a las que había cuando se tomaron los datos originales. Así se observa que estudios realizados determinado año se publican varios años más tarde, lo cual obliga a tomar sus resultados con reserva en el caso de que se quieran tomar como base para proponer acciónes regulatorias de un recurso, e incorpora además un factor de incertidumbre respecto de las capturas proyectadas. Finalmente, es necesario lograr una vinculación mayor entre el quehacer científico y el proceso productivo pesquero, con el fin de aplicar de manera directa los conocimientos biológicos que permitan rendimientos mayores de esta actividad económica, pues el alejamiento que existe entre ambos ha influido para que la pesca no se consolide en el estado de Yucatán en la medida de su riqueza íctica y de su infraestructura humana y material. LITERATURAALFARO, G. L. A. y M. LÓPEZ, Tesis Profesional, Contribución al estudio de la pesquería del pargo criollo (Lutjanus analis, Cuvier) y pargo mulato (Lutjanus griseus, Linneo) en el litoral del Estado de Yucatán. ENEP-Iztacala, UNAM, México. 1986. ARREGUÍN-SÁNCHEZ, F. y E. A. CHÁVEZ, Internacional Symposium on Age and Growth of Fish. A method for fish stock assessment based upon age structure. Desmoines, Iowa (M.S.). 1985. BAGENAL, T., Methods for assessment of fish production in fresh-waters. I.B.P. Handbook No. 3, Blackwell Sci., Pubs., Oxford. 1978. 365 p. BASSON, M., A. ROSENBERG y J. R. BEDDINGTON, J. Cons. Int Explor. Mer, The accuracy and reliability of two new methods for estimating growth parameters from length-frequency data. 1988. 277-285. 44: CADDY, J., Fish Mar. Ser., Report Some considerations underlaying definitions of catchability and fishing effort in shellfish and their relevance for stock assessment purpose. 1979. 1-9. 1489: CANTARELL, E., Tesis Licenciatura, Determinación de la edad y el ritmo de crecimiento del pargo canané (Ocyurus chrysurus Bloch, 1791) en el Estado de Yucatán. ENEP-Iztacala, UNAM, México. 1982. CONTRERAS, M., Curso de Investigación Pesquera, Programa de pesca experimental para la costa de Yucatán. CRIP Yucalpetén. 1979. CROCKER, R. A., Growth and food of the gray snapper Lutjanus griseus in Everglades National Park. Trans. Amer. Fish. Soc., 1962. 379-383. 91 (4): DOI, T., Curso matemático sobre dinámica de poblaciones. Instituto Nacional de Pesca, SEPESCA, México. 1976. DRUZHININ, A. D., J. Ichthyol, The range and biology of snappers (Fam. Lutjanidae). All Union Reserch Institute for Sea Fisheries and Oceanographic (UNIRO). Moscú. 1970. 715-735. 10: GARDUÑO, D. M. y C. A. LÓPEZ, Tesis de Licenciatura, Análisis preliminar de la población del pargo canané Ocyurus clirysurus (Bloch) (Pisces: Lutjanidae), en el Banco de Campeche. UAM, México. 1985. GONZÁLEZ, M. E., Tesis de Maestría. Estudio de la pesquería de huachinango (Lutjanus campechanus Poey) del Banco de Campeche. CINVESTAV-INP-Unidad Mérida. Yucatán, México. 1988. MEXICANO, G., Tesis Profesional, Contribución al conocimiento de la pesquería del pargo canané (Ocyurus chrysurus Bloch, 1971) en el litoral del Estado de Yucatán. ENEP-Iztacala, UNAM, México. 1986. MEXICANO-CÍNTORA, G. y F. ARREGUÍN-SÁNCHEZ, Congreso de Ciencias del Mar. Dinámica de las poblaciones de-rubia (Lutjanus synagris) y canané (Ocyurus chrysurus) de las costas de Yucatán. La Habana, Cuba. 1987. MOREAU, J., The age and Growth of Fish. Mathematical and biological expressión of growth in fishes: recent trends and further development. In: R.C. Summerfelt and G. E. Hall (Eds.). The Iowa State University Press. Ames, Iowa. 1987. pp. 81-113. MOSELEY, F. N., Biology of the red snapper Lutjanus aya (Bloch) of the North Western Gulf of Mexico. Inst. Mar. Sci., 1966. 90-101 11:. PAULY, D., J. Cons. Ins. Explor. Mer., On the interrelationships between natural morgrowth parameters and mean environmental temperature in 175 fish stocks. 1980. 175-192. 30 (3): PITCHER, T. J. y P. J. HART, Fisheries Ecology. Crom. Helm, Londres 1982. 360 p. STARCK, W. A. y R. E. SCHROEDER, Studies in Tropical Oceanography, Investigations on the gray snapper, Luijanus griscus. Univ. of Miami Press, Florida. 1971. (10). THOMPSON, R. y J. MUNRO, The biology, ecology, eploitation and management of Caribbean reef fishes. The biology and dynamics of Caribbean reef fushes Lutjanus (snappers). In: R- Thompson y J. Munro (Eds.). Univ. of the West Indies,Part 5d. Departament of Zoology. Research Report 3. Kingston, Jamaica. 1974. TORRES, R., Tesis Profesional Diagnosis de la pesquería de la rubia (Lutjanus synagris Linnaeus, 1758) en las costas de Yucatán. ENEP-Iztacala, UNAM, México. 1984. , TORRES, R. y E. CHÁVEZ, Ciencias Marinas, Evaluación y diagnóstico de la pesquería de la rubia (Lutjanus synagris L. 1758) en el Estado de Yucatán. 1987. 7-29. 13 (1): TORRES, R., Tesis de Maestría, Análisis biológico-pesquero de dos temporadas de pesca de la rubia (Lutjanus synagris L. 1758) en las costas de Yucatán. CINVESTAV-INP-Unidad Mérida. Yucatán, México. 1987. TORRES, R. y S. SALAS An.Inst Cienc. del Mar.y Limnol, Univ. Nal. Autón México, Crecimiento y mortalidad de la rubia Lutjanus synagris de las costas de Yucatán durante las temporadas 1983-1985. (1990). 157-161. 17 (2): WINBERG, G. C., Methodsfor the estimation of productions of aquatic animals. Academic Press, Londres. 1971. 175 p.
|