|
ALGUNAS CARACTERÍSTICAS ECOLÓGICAS DEL
FITOPLANCTON DE LA REGIÓN CENTRAL DE LA BAHÍA DE CAMPECHE
Trabajo recibido el 15 de noviembre de 1990 y aceptado para su
publicación el 8 de abril 1991.
Sergio Licea
Laboratorio de Fitoplancton y Productividad,
Instituto de Ciencias del Mar y Limnología, UNAM. Apartado Postal
70-305, México, D.F. 04510. México.
Hermilo Santoyo
Departamento de Biología Marina, Universidad
Autónoma de Baja Califotnia Sur. Apartado Postal 19-B, La Paz 23080.
B.C.S. Contribución No. 705 del Instituto de Ciencias del Mar y
Limnología. UNAM.
Se analizan los nutrimentos, parte de la estructura
del fitoplancton y su productividad en un transecto de cinco estaciones en el
verano de 1977. Los resultados indican oligotrofía acentuada con
estabilidad de las masas de agua y la presencia de una termoclina a 30 m de
profundidad. El contenido de clorofila a y la producción primaria fueron
relativamente bajos. La producción del nanofitopiancton y del
microfitoplancton fue de 37.2 y 102.0 mg C/m /día, respectivamente;
dinoflageladas, diatomeas y cianofitas constituyen los grupos dominantes,
así como los productores primarios Principales. El análisis
taxonómico reveló la presencia de 112 especies, de las cuales 72
correspondieron a diatomeas y 31 a dinofiageladas. Los taxa dominantes fueron
los géneros: Gymnodinium, Glenodinium, Gyrodinium y
Amphidinium, así como las especies Nitzschia
pungens y Oscillatoria thiebautii. El
nanofitoplancton estuvo integrado fundamentalmente por dinoflageladas y
fitoflageladas del tipo desnudas. Se registró un incremento en la
diversidad de especies, relaciónado con el incremento de la biomasa, que
se inicia en la zona costera y continúa hacia la porción
pelágica.
Nutrients, a portion of phytoplankton and its
productivity, in a five stations transect in the summer of 1977 were analyzed.
Observations showed a strong oligotrophic condition, with water masses
stability and thermocline presence at 30 m depth. Chlorophyll a concentration
and primary production valueres were relatively low. Nanophytoplankton and
microphytoplankton production were 37.2 and 1O2.O mg c/m²/day
respectively, being dinoflagellates, diatoms and cyanophytes dominant groups,
as well as main primary producers. On systematic examination, it was found to
contain 112 species, including 73 diatoins, and 31 species of dinoflagellates.
Prevailing genera were: Gymnodinium, Glenodinium, Gyrodinium,
Amphinium, as well as the species Nitzschia pungens
and Oscillatoria thiebautii . Nanophytoplankton
was primarily composed of dinoflagellates and phytofiagellates, both naked
type. An increase of species diversity was registered from the coastal zone to
the pelagic region, which was related to phytoplankton biomass
increase.
La Bahía de Campeche se ha convertido en una zona importante desde el punto de vista ecológico, económico y social, en virtud de su reconocido valor como zona pesquera de las más importantes de México (Secretaría de Pesca, 1990). Recientemente sobresale su importancia por contener yacimientos petroleros considerables, cuyas exploración y extracción han generado conflicto de intereses entre los recursos naturales renovables con los no renovables (Nehring, 1978; Stewart-Gordon, 1979), cuyo análisis escapa a los objetivos del presente estudio, que solamente se reduce a informar acerca de algunas características ecológicas del fitoplancton en la porción central de la Bahía de Campeche, como: distribución horizontal y vertical del grupo de vegetales planctónicos, su composición, diversidad de especies y productividad primaria. ÁREA DE ESTUDIOLa Bahía de Campeche está situada en la región suroriental del Golfo de México; la característica dominante de la circulación ciclónica se debe principalmente a variaciones del transporte en el Canal de Yucatán (Nowlin, 1972; Molinari et al., 1978; Merrell y Morrison, 1981). Sin embargo, este giro desaparece completamente en mayo; en julio, comienza a desarrollarse nuevamente sobre la costa occidental de Yucatán, y su efecto se percibe en la bahía al inicio del otoño (Monreal-Gómez y Salas de León, 1985; Monreal-Gómez 1986). De esta manera es de esperarse que en esta época de muestreo el área de estudio tenga la mayor estabilidad de las aguas. Más información al respecto se encuentra en Cruz (1971); Bulanienkov y García-Díaz (1975); Ruiz-Rentería y Merino-Ibarra (1989). Desde el punto de vista biológico, la Bahía de Campeche es una región de productividad biológica relativamente alta, en comparación con el resto del Golfo y el Mar Caribe adyacente (Bogdanov et al., 1968; Khromov, 1969; Bessonov et al., 1971; Cruz, 1971; Zernova, 1969, 1974; Krylov, 1974; Villalobos y Zamora, 1975; Licea et al., 1982). Con base en el trabajo de Santoyo y Signoret (1973) se distribuyeron cinco estaciones de muestreo en un transecto perpendicular a la costa, frente a la Laguna de Términos, hasta la isobata de 183 m, profundidad que en este sitio marca el límite de la plataforma continental (Fig.1). 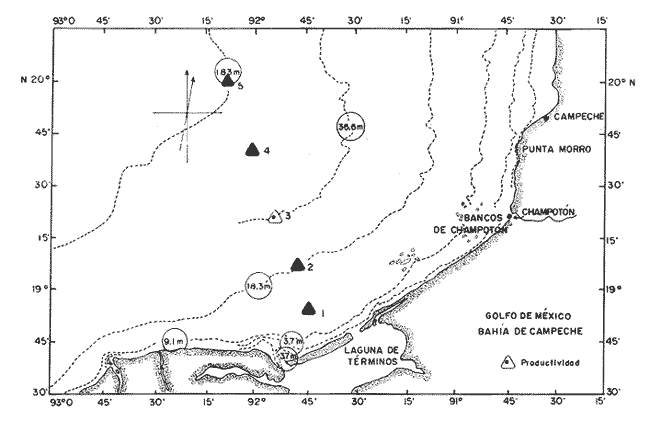 Figura 1. Área de estudio en la Bahía de Campeche. Localización de las estaciones. MÉTODOSLa obtención de muestras y los experimentos para la realización de este estudio se efectuaron en julio de 1977, durante la campaña oceanográfica EN-010, de la Universidad de Rhode Island, en el buque oceanográfico (B. O.) "Endeavor". La temperatura y la salinidad del agua se obtuvieron por medio de un equipo S.T.D. (salinidad, temperatura y densidad). Los nutrimentos inorgánicos (nitratos, amonio, fosfatos y silicatos) se cuantificaron con un autoanalizador, según las recomendaciones de Smayda (1975). Las determinaciones de clorofila a se efectuaron por fluorometría, por el método de Holm-Hansen et al. (1965). El cálculo de la productividad primaria se realizó siguiendo el método de asimilación de carbono radioactivo (C 14) , de acuerdo con lo propuesto por Steemann Nielsen (1964). Las incubaciones se hicieron in situ simulado durante 24 horas, con iluminación natural; la valoración de la asimilación del isótopo radioactivo se efectuó por medio de un analizador de centelleo líquido. Los experimentos para determinar la productividad primaria se hicieron en microfitoplancton (de 54 µm), nanofitoplancton (de 10 µm) y fitoplancton total. La separación por tallas se efectuó mediante prefiltración con red de nilón. El fitoplancton se analizó cualitativa y cuantitativamente por el método de Utermöhl (1958). Los análisis se hicieron con cámaras de sedimentación de 50 ml, a 250 y 400 aumentos cada muestra, en volúmenes de 5 a 50 ml, según la abundancia y el tipo de comunidad. RESULTADOS
NUTRIMENTOSLas variaciones de contenido de nutrimentos fueron relativamente amplias, como se encuentra en el cuadro siguiente, en el que se dan los valores máximo y mínimo de cada componente químico.  La distribución de la concentración de salesnutrimentos tuvo como patrón general que los valores bajos se localizaron cerca de la costa, y los mayores en la zona del talud continental, tipificación que señala gradientes más o menos acentuados desde el talud hacia la margen costera. La distribución vertical de los nutrimentos tuvo, así mismo, un patrón que se manifestó con las concentraciónes más altas en profundidades mayores de 50 metros; resultan notables los valores elevados de nitratos (13.5 µ-at/l) y de silicatos (6.33 µg-at/l) con respecto al nitrógeno del amonio y el fósforo de los fosfatos. El nitrógeno del amonio estuvo en proporción bastante baja comparativamente con el de los nitratos. En la zona de 70-80 metros de profundidad se identificó una "lengua" de mayor concentración (0.8 µg-at/l), semejante ala distribución de los fosfatos en ese mismo intervalo (Fig. 2). FITOPLANCTONLa característica del plancton vegetal se particularizó por deficiencia de constituyentes del fitoplancton. La escasez fitoplanctónica se hizo más evidente hacia la región pelágica, con respecto a la zona más cercana a la costa; en la distribución vertical hubo disminución proporciónal con la profundidad (Tabla 1), excepto en las cotas de 40 y 100 metros (estaciones 4 y 5), en las cuales ocurrieron incrementos notables de individuos. Los valores de células por litro fluctuaron entre los límites impuestos de 138,000, como registro máximo, y 1,000, como registro mínimo (estaciones 1 y 5, respectivamente). Las diatomeas constituyeron el grupo dominante en las áreas aledañas a la costa (estaciones 1 y 2); llegan a representar hasta el cien por ciento. La proporción de diatomeas fue disminuyendo conforme hubo un alejamiento respecto a la línea litoral, hasta alcanzar porcentaje tan bajo como 1.0 en la estación 5, patrón que se vio alterado sólo por incrementos ligeros debidos a núcleos poblacionales de Hemiaulus sinensis y Hemiaulus membranaceus (Lám. 1, Figs. 1 y 2), coincidentes con los incrementos en la densidad fitoplanctónica ya citados. La relación de los taxa determinados se refiere en la tabla 2. 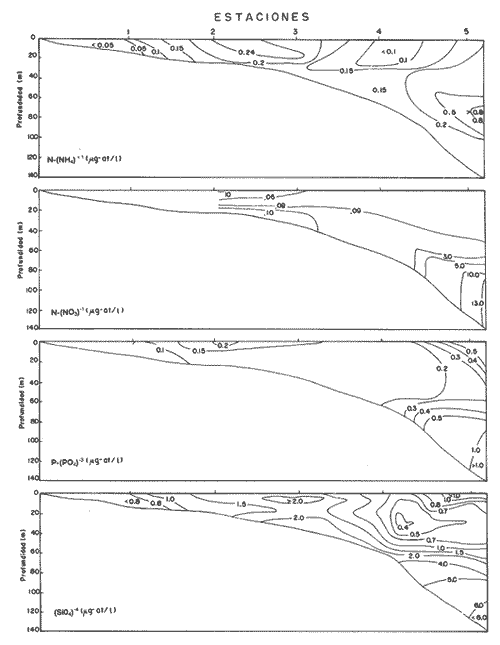 Figura 2. Perfiles de la distribución de los nutrimentos: amonio, nitratos, fostatos y silicatos, en ug-at/l. Las dinoflageladas desnudas (Lám. 2, Figs. 4, 6, 7), las fitoflageladas (Lám. 2, Figs. 10-12), junto con las cianofitas y las cocolitofóridas sustituyeron a las diatomeas en las estaciones cercanas al límite de la plataforma continental. Es importante destacar que en la zona central de ésta hubo un dominio de Oscillatoria thiebautii (estación 3) restringido a la capa superficial, pues en ninguna otra región a lo largo del transecto se encontró esta población (Lám. 2, Fig.14).  TABLA 1 Densidad del fitoplancton, expresada en células/litro DIVERSIDAD DE ESPECIESLa diversidad de especies se calculó por medio de la ecuación de Shannon y Weaver, tomado de Pielou (1966). Los valores de este factor tuvieron gradientes, cuya dirección fue de la costa al mar abierto y de la superficie hacia el fondo (Fig. 3). Sin embargo, existió un incremento ligero en el borde de la plataforma continental en la profundidad de 30 a 50 metros, región donde dominó con varias especies el género Hemiaulus, una de las cuales (H. sinensis) registró un promedio del 55%. Por otro lado, se evidencia una relación notable entre la diversidad de especies y la clorofila a, de tal modo que el incremento de la biomasa estuvo relacionado con el aumento de la diversidad. CLOROFILA aLa distribución de la clorofila a se correlacionó con la distribución horizontal y vertical de la densidad fitoplanctónica (Fig. 3) con la presencia del grupo de las diatomeas, pues este pigmento fue abundante en las estaciones cercanas a la costa, y disminuyó en las restantes, en las cuales se definieron sólo incrementos ligeros hacia profundidades de 30 a 70 metros. El contenido de este pigmento fluctuó entre 0.49 µg/l como valor máximo y 0.03, µg/l como valor mínimo. PRODUCCIÓN PRIMARIALa producción primaria medida en la zona central de la plataforma continental enfrente de la Laguna de Términos fue de 40.4 mgC/m²/día, tomando en consideración toda la gama de tallas del fitoplancton, en tanto que el microfitoplancton (10-54 µm) tuvo un registro de 101.9 mgC/m²/día, mientras que el nanofitoplancton (< de 10 µm) produjo 37.2 mgC/m²/día. De esta manera, el microfitoplancton fue el grupo más importante como productor primario, el cual estuvo dominado por la diatomea Hemiaulus sinensis, Glenodinium spp y Gymnodinium spp, en tanto que el nanofitoplancton estuvo compuesto principalmente por Amphidinium spp y fitofiageladas desnudas no identificadas, así como la cianofita Oscillatoria thiebautii. Esta última sólo en la parte superficial, como se anotó con anterioridad.  TABLA 2 RELACIÓN DE ESPECIES DETERMINADAS 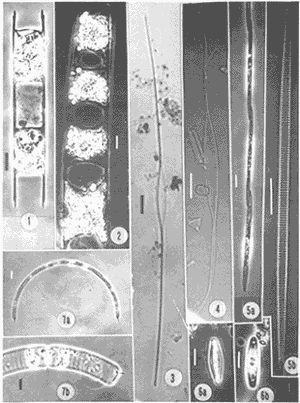 Lámina 1. Explicación de la lámina. La barra de la escala equivale a 10 µm; en cada figura se indica la procedencia del material. Las ilustraciones diferentes que pertenecen a un mismo espécimen se denotan por el mismo número arábigo con letras sucesivas. Todas las ilustraciones se hicieron con microscopio de contraste de fase. Fig. 1. Hemiaulus sinensis- E-2, dos células en cadena; Fig. 2. Hemiaulus membranaceus- E -1, cuatro células en cadena; Fig. 3. Nitzschia longissima.- E-3, valva libre de materia orgánica; Fig. 4. Nitzschia closterium,- Ibidem; Fig. 5a. Nitzschia pungens E-3, tres células en cadena; Fig. 5b. Ibidem, valva libre de materia orgánica, Fig. 6a. Navicula sp- E-5, en vista valvar; Fig. 6b. Ibidem, en vista cingular; Fig. 7a. Rhizosolenia stolterfothii.- E-1, dos células en colonia circular; Fig. 7b. Ibidem, dos células en colonia semicircular. 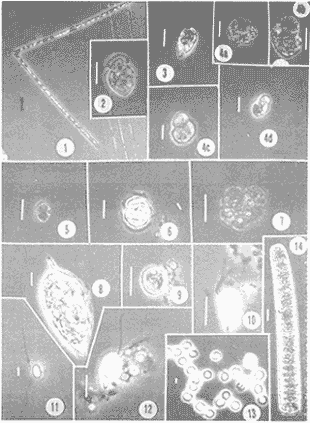 Lámina 2. Fig. 1. Thalassiothrix frauenfeldii E-1, dos células en cadena: Fig. 2.Amphidinium sp a.- E-5; Fig. 3.Amphidinium sp b, E-4; Figs. 4a-d. Gymnodinium sp a, E-5, mismo espécimen, en posición diferente; Fig. 5.Peridinium sp.- E-3; Fig. 6. Gymnodinium sp b.- E-4; Fig. 7. Ptychodiscus brevis- E-4; Fig, 8. Prorocentrum micans- E-3; Fig. 9. Prorocentrum minimum - E-5; Figs 10-12. Fitoflageladas desnudas (Fam. Prasinopllyceae).- E-3, 4,5 respectivamente Fig. 13. Cianofita (Fam. Chroococcaceae), E-2; Fig. 14. Oscillatoria thiebauthii. -E-3. 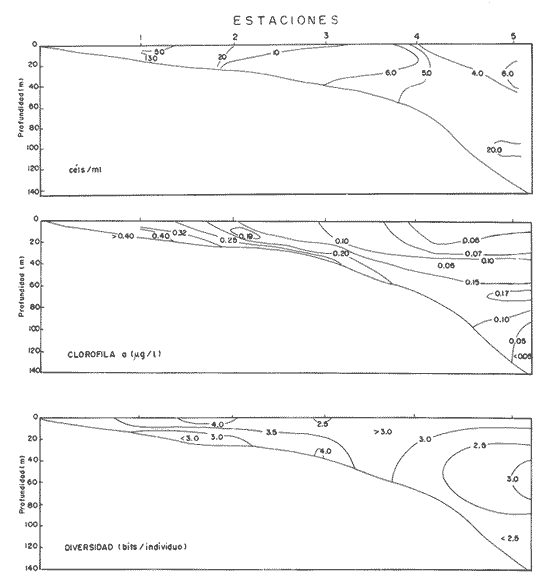 Figura 3. Perfiles de isopletas de fitoplancton, clorofila a y diversidad de especies DISCUSIÓN Y CONCLUSIONESLas características ecológicas del área estudiada determinan una oligotrofía acentuada en esta época (verano), a causa de la cantidad baja de sustancias nutritivas disueltas y de estabilidad de las masas de agua; estabilidad indicada por la presencia de una termoclina en la profundidad de 30 metros y por el caldeamiento de la masa acuosa en la zona litoral, menos profunda. Las características oligotróficas registradas en este estudio coinciden con la aseveración de El-Sayed et al. (1972), consistente en que los valores más bajos de producción y biomasa en el Golfo de México se generan durante primavera y verano. Sin embargo, en esta misma época se registraron valores de producción del orden de 422 mgC/m²/día en la región septentrional del banco de Campeche, zona con propiedades eutróficas (Smayda, 1977), lo cual significa que en el mismo período estacional existen áreas cuya fertilidad difiere notablemente en el propio Banco de Campeche. Los valores de contenido de clorofila a y de producción primaria quedan, en términos generales, por debajo del promedio de las registradas en el Golfo de México por El-Sayed et al. (op. cit.), cuyos valores son los siguientes:  Los datos anteriores revelan que, en general, el Golfo de México es un área oligotrófica. Sin embargo, en esta época, la región de la plataforma continental frente a la Laguna de Términos presentó una oligotrofía más acentuada que la promedio para la región. La escasez de elementos nutritivos inorganicos está aunada a una fase de la sucesión fitoplanctónica en la que las dinoflageladas y las cianofitas son los grupos dominantes, en tanto que las diatomeas han sido relegadas a un término secundario; su prosperidad se fundamenta en eficiencia mayor en la asimilación de los nutrimentos del medio acuático y su consecuente repercusión en la productividad primaria. Los valores más altos de productividad correspondieron a las dinoflageladas, lo cual indica eficiencia mayor que las diatomeas (Hemiaulus sinensis, principalmente); posiblemente esta característica se debe a que los fosfatos fueron los compuestos limitantes para el desarrollo del fitoplancton (Smayda, 1977). Estudios realizados en esta misma área por Santoyo y Signoret (1973, 1975) y Licea (1977, 1982) señalan que las características cualitativas y cuantitativas del fitoplancton fueron semejantes a las registradas en este estudio, en las cuales aquél fue escaso; la dominancia de dinoflageladas fanerógamas y fitoflageladas en la región central del banco de Campeche, frente a la Laguna de Términos, induce a plantear que las características ecológicas de la taxocenosis fitoplanctónica se repiten cíclicamente. AgradecimientosLos autores hacen patente su reconocimiento al Dr. T. Smayda por las facilidades brindadas para la realización de este estudio a bordo del buque oceanográfico (B/O) Endeavor, de la Universidad de Rhode Island; así como al M. en C. Roberto Cortés por la revisión crítica de la versión inicial. A la Biol. Ruth Luna por haber transcrito ésta. LITERATURABESSONOV, N., O. GONZÁLEZ y A. ELIZAROV, Coloquio sobre investigaciones y recursos del Mar Caribe y regiones adyacentes. Curasao, Antillas Holandesas, Resultados de las investigaciones Cubano-Soviéticas en el Banco de Campeche. En: UNESCO (Ed.). París 1971. pp. 375-383. 18-26 nov. 1968, BOGDANOV, D.V., V.A. SOKOLOV y N.S. KHROMOV, Oceanol., Regions of high biological and commercial productivity in the Gulf of Mexico and Caribbean Sea. 1968. 371-381. 8: BULANIENKOV, S. K y C. GARCÍA-DÍAZ, Influencia de los procesos atmosféricos en el afloramiento del Banco de Campeche, 1975. 99-140. 1 (2): CRUZ, A. de la, Coloquio sobre investigaciones y recursos del Mar Caribe y regiones adyacentes. Curasao, Antillas Holandesas, Estudios de plancton en el Banco de Campeche. En: UNESCO (Ed.). Paris 1971. pp. 375-383. 18-26 nov. 1968, EL-SAYED, S. Z., W. M. SACKETT, L. M. JEFREY, A. D. FREDERICKS, R. P. SAUNDERS, P. S. CONGER, G. A. FRYXELL, K. A. STEIDINGER y S. A. EARLE, Serial Atlas of the Marine Environment, Chemistry Primary Productivity and Benthic Algae of the Gulf of Mexico. Amer. Geogr. Soc., 1972. 22.26p. HOLM-HANSEN, O., C. J. LORENZEN, R.W. HOLMES y J.D.H. STRICKLAND, J. Cons. Perm. Intern. Explor. Mer., Fluorometric determination of chlorophyll. 1965. 5 -15. 30: KHROMOV, N. S., Soviet Cubban Fishery Research, Distribution of plankton in the Gulf of México and some aspects of its seasonal dynamics. In: D.V. Bogdanov (Ed.). 1969. 36-56 KRYLOV, V.V., Distribución del fitoplancton y de las biocenosis planctónicas en el Banco de Campeche. Cuba Res. Inv., 1974. 75-79. 1: LICEA, S., Symposium on Progress in Marine Research in the Caribbean and adjacent Regions. Variación estacional del fitoplancton de la Bahía de Campeche, México (1971-1972) In H.B. Stewart (Ed.). FAO Fish. Rep., Caracas, Venezuela, 1977. 253-273. 200: 2-16 Julio 1976. LICEA, S., R. LUNA y P. TORRES, Informe Final (inédito), . Evaluación de los posibles efectos del derrame del Pozo Ixtoc-I sobre las comunidades del fitoplancton y la producción primaria en la Sonda de Campeche. UNAM/Progr. Coord. Est. Ecol. Sonda Campeche 1982. 44 p. MERREL, W. J. Jr. y J. M. MORRISON, On the circulation of the westem Gulf of Mexico with observations from April 1978. J. Geophys. Res., 1981. 4181-4185. 86: MOLINARI, R. L., J. F. FIESTA y D. W. BEHRINGER, The circulation in the Gulf of Mexico derived from estimated dynamic heigth fields. J. Phys. Oceanogr., 1978. 987-996. 8: MONREAL-GÓMEZ, A. y D. A. SALAS DE LEÓN, Proceedings "Progress in Belgian Oceanographic Research". Barotropic and baroclinic modes in the Gulf of Mexico. In: V. Grieken and R. Wollast (Eds.). Brussels, 1985. pp. 81-91. 3-5 March, 1985, MONREAL-GÓMEZ, A., Memoria Reunión Anual 1986, Unión Geofísica Mexicana. Análisis del primer modo baroclínico en la Bahía de Campeche. En: Urrutia Fucugauchi (Ed.). Univ. Mich. San Nicolás Hidalgo, México, 1986. pp. 441-448. NEHRING, R., Ciencia y Desarrollo, Campos petroleros gigantes y recursos mundiales de petróleo. CONACyT. 1978. 199 p. NOWLIN, W. D., Contributions on the physical Oceanography of the Gulf of Mexico. Winter circulation patterns and property distributions. In: L. R. A. Capurro and J. L. Reid (Eds.). Gulf Publ. Co., Houston, 1972. pp. 3-53 PIELOU, E. C. Amer Natur., Shannon's formula as a measurement of specific diversity, its use and misuse. 1966. 463-465. 100 (914): RUIZ-RENTERÍA F. y M. MERINO-IBARRA, Upwelling off the north coast of Yucatan peninsula. In: E. Chávez (Ed.). Proc. Worshop Mexico-Australia Mar. Sci., Merida, Yuc. and Puerto Morelos, Q. Roo, Mexico. 1989. pp. 149-160 July 6-17, 1987, SANTOYO, H. y M. SIGNORET, Rev. Lat-amer. Microbiol., Hidrología y Fitoplancton en un transecto en la plataforma continental de la Bahía de Campeche, México (agosto de 1972). 1973. 207-215. 15 (4): SANTOYO, H. y M. SIGNORET, Rev. Lat-amer. Microbiol., Variación nictemeral del fitoplancton en la Bahía de Campeche, México. 1975. 161-167. 17 (3): SECRETARÍA DE PESCA, Anuario estadístico de pesca 1988. Dirección General de Informática y Estadística, México. 1990. 351 p. SMAYDA, T., Gorilla Manual. Univ. Rhode Island, Grad. Sch. Oceanogr., (unpublished). Kingston, USA 1975. 106 p. SMAYDA, T. Endeavor 010 Cruise Report. Univ. Rhode Island, Grad. Sch. Oceanogr., (unpublished). Kingston, USA 1977. 10p. STEEMANN. E., J. Ecol, Recent advances in measuring and understanding marine primary production. 1964. 119-130. 52: UTERMÖHL, H., Mitt. int. Ver. theor. angeb. Zur Vervollkommnung der Quantitativen Phytoplankton-Methodik. Limnol., 1958. 1-38. 9: STEWART-GORDON, J., El petróleo en México y en el mundo, El petróleo mexicano: mitos, realidades y futuro. En: CONACyT (Ed.). CONACyT 1979. pp. 259-268. VILLALOBOS-FIGUEROA y M. E. ZAMORA-SÁNCHEZ, Mem. 1er Simp. Lat. Ocean. Biól. Importancia biológica de la Bahía de Campeche. En: R. Pérez Rodríguez (Ed.). México D.F., 1975. pp. 375-394. 25-29 nov. 1974, ZERNOVA, V.V, Oceanology, The horizontal distribution of phytoplankton in the Gulf of México. 1969. 565-574. 9 (4): ZERNOVA, V.V., Oceanology, Distribution of the biomass of phytoplankton in the tropical Atlantic ocean. 1974. 1070-1076. 14:
|