|
POSITION PREFERENCES IN A SCHOOL OF ATLANTIC HERRING
(Clupea harengus): THE EFFECT OF HUNGER
Trabajo recibido el 26 de agosto
de 1991 y aceptado para su publicación el 2 de diciembre de
1991.
Carlos J. Robinson
Instituto de Ciencias del Mar y Limnología.
UNAM. Estación "Mazatlán" Explanada de la Alzada y Cerro del
Crestón S/N Apartado Postal 811, CP 82240 Mazatlán, Sinaloa.
México.
Virgilio Arenas
Instituto de Ciencias del Mar y Liminología.
UNAM. Apartado Postal 70-305. México 04510 D.F. MÉXICO.
Contribución No. 703 del Instituto de Ciencias del Mar y
Limnología. UNAM.
Un grupo formado por diez arenques (Clupea
harengus) fué mantenido en condiciones experimentales y la
posición de cuatro de ellos fué registrada mediante
video-fotografía en diferentes condiciones de hambre. Los resultados
demuestran que en condiciones de hambruna y bien alimentados, los peces no
nadan azarosamente; pero se observaron posiciones específicas. Cuando un
pez se mantuvo bien alimentado y el resto hambriento, su presencia al frente
del grupo se incrementa significativamente. Una revisión de la
literatura indica que nadar al frente del cardúmen proveé una
mejor posición para el encuentro con el alimento y una seguridad
relativa contra el ataque de predadores.
The swimming locations of four herring
(Clupea harengus) in a shoal of ten fish were investigated
in experimental conditions by video photography. Observations were performed
with the fish in different hunger states. Results shows that when all fish
where well-fed or all hungry, fish did not swim randomly, but particular
positions were preferred. When a single fish was well- fed and the rest hungry
its appearance at the front of the shoal increases significantly. A literature
review indicates that swimming at the front of the shoal appears to provide a
better position for an encounter with food and relative security against
predator attack.
Perhaps at first sight one of the most noticeable features of individuals in a single group of fish is the relative uniformity in size, shape and colour. However, recent work has demonstrated that individuals continually re-estimate the cost and benefits of remaining in a shoal (Pitcher, 1986) suggesting that variation in behaviour may be found even in structured and polarised groups (Magurran, 1986). Differences between members of a group of fish have been investigated from at least two aspects. First, the individuals have been shown to respond to different conditions, but the behaviour of particular individuals is not followed (e.g. Seghers, 1974; Giles and Huntingford, 1984; Street et al., 1984 Magurran, 1986). The second, more elaborate procedure, is to recognise specific individuals, usually by means of artificial marks or guided by particular features in the fish. Within this category we can recognise two different applications: i) behavioural aspects of shoaling without any implications of structure and ii) behaviour of individuals within a structured and polarised group. Using the former approach, interesting aspects of individual behaviour of shoaling fish have been investigated (Kennedy, 1981; Helfman, 1984; Bayliff, 1988). However, because of its inherent methodological difficulty, together with the assumption of 'equality' within members of a school of fish, this latter approach has rarely been utilised. Consequently, the particular aspects of shoaling behaviour concerning individuals within a structured group have been poorly investigated. Only a handful of investigations have been developed, all of them under assumed constant motivational states (McFarland, 1985) (e.g. Healey and Prieston, 1973; Partridge, 1978; Pitcher, et al., 1982; Pitcher et al, 1985). POSSIBLE IMPLICATIONS OF DIFFERENT BEHAVIOURS IN A SCHOOLAs the costs and benefits of belonging to a group change, shoaling tendency may vary within individuals (Pitcher, 1986). Species of open water environments that aggregate in large numbers should expect to present lower phenotype and behavioural differences than those which inhabit heterogeneous habitats. However, even within the former species with highly polarised and synchronised manoeuvres, behavioural differences may exist, though they are difficult to find. If differences between members of a school do appear, the question to ask is what are the costs or benefits of being different in a shoal of fish? The answer could be closely related to the survival value of being in a shoal as a mechanism to avoid predation, increase the efficiency of gathering food and possibly to some hydrodynamical advantages. The present study aims to find out whether in a two-dimensional plane, some members of a shoal of herring (Clupea harengus) occupy randomly positions within the group or whether there is preference for specific locations. Furthermore, and for the first time in the literature, we intend to associate the cost or benefits in position preference (if any) to the level of hunger. MATERIAL AND METHODSTen herring (mean total length = 15.7 cm, s.d. = 0.91; mean wet body weight = 25.8 g, s.d.= 5.1) were maintained in a circular tank (2.8 m diam., 1 m deep) under a 13% wet body weight fish -¹day-¹ diet of mysid shrimps. Using black plastic the shoal was herded to one side of the tank where four fish were randomly selected and carefully netted and transferred to a 20 1 plastic container (30 x 25 cm) aerated to saturation. Fish were anaesthetised with MS 222 (tricaine methane sulphonate) (1: 15,000) and marked through an inoculator using a standard stain of methylene blue on the fishes' right flank below the lateral line. A combination of number of spots, together with position on the fish was used in order to recognise each of the four individuals. The fish labelled "A" was 14.3 cm in length; fish "B" was 15.7 cm in length; fish "C" 16.7 cm in length and fish "D" 17.1 cm. The fish were swimming continuously around the tank in a clock-wise direction, completing a circuit in a mean time of 28.7 seconds (s.d. = 1.42) and following a track rarely exceeding 1 m away from the tank wall. A video-recorder system (Hitachi CCV camera and a Panasonic video cassette recorder model NV-788d) mounted beside the tank was used to film the fish. The camera was pointed in a direction which allowed the recognition of the marked fish as the shoal passed through a single point in the tank. A 17.5-105 mm zoom lens was set at 75 mm focal length giving a field of view of approximately 4.8 by 3.7 fish body length. The subject distance was 3.1 m from the camera at an incident angle of 35 °. From this viewpoint it was possible to follow a fish for a mean time of 3.1 s (s.d. 0.34). Since the shoal was being filmed at 25 frames per second it was possible to analyse about 77 frames in each pass. Every time fish passed by the filming area the position of each marked fish was identified and scored. CRITERION TO DEFINE POSITIONUsing a system of two dimensions (front-back-side), four positions of the fish in the shoal were selected in order to recognise possible preferences. 1) front: defined here as the situation where a fish did not have a companion in front of it and no other fish were further forward than its middle body. 2) back: no fish behind and the nearest in front at least half a body length forwards. 3) side: no other fish came between it and the tank wall (left side) or not other fish between it and the centre of the tank (right). 4) middle: companions to the front, back and both sides of the fish. CONDITION 1 ALL FISH WELL-FED COMPARED TO ALL FISH HUNGRYAfter tagging, the fish were undisturbed and provided with the daily diet of 13% fish body weight day¹ of food given at 1200 hours. Filming sessions which lasted three hours from 1400 hours, started one week after the marking process (day 1), observations were repeated the following day (day 2). Overall, the results of this session will be referred to as 'all fish well-fed'. During the following 48 h, all the fish were deprived of food and recording started again at 1400 hours. The same procedure of position recording was performed as with the previous diet, although this time fish were fed the complete diet after the first recording session (day 1) and maintained for a recuperation period of five days. Fish were again deprived of food for 48 h in order to obtain the second set of observations under this diet (day 2). Overall these two results will be referred to as 'all fish hungry'.
CONDITION 2 ONE PARTICULAR FISH WELL-FED; THE REST HUNGRY AND VICE VERSAOn the fifth day of recuperation period and one hour before the light went off, a curtain of black plastic was used to partition the tank along its diameter. Fish "A" was carefully herded into one half while the rest remained in the other half. Five herring (mean total length = 14.6 cm, s.d. = 0.91) who were maintained in an adjacent circular tank, were carefully transferred to the experimental tank and placed together with the separated fish. Fish were left in this condition overnight and the following day a ration of 13% body weight was provided at 1000 hours to the fish "A" and its companions. Once finishing their ration, which took about one hour, the spare fish were netted and returned to their tank, the curtain was carefully removed and fish were allowed to swim in a single shoal. Prior to the observations, a series of trials were carried out to evaluate the effect of the division on the overall behaviour of the shoal. They showed that minutes after the spare fish were returned to their tank and the curtain removed, experimental fish were swimming in a compacted shoal showing no visible alteration in their swimming performance. The next day, the entire procedure of partitioning and feeding was repeated in order to have a register of nine fish with a hunger level of 48 h and fish "A" well-fed (day 1). Video- recording started at 1400 hours lasted three hours and position of fish "A" was then scored as before. After this session all fish were fed complete diet during five days in order to normalise hunger. Then, the procedure of separation was repeated again using each marked fish (i.e. fish B,C,D). Each one with the same period of isolation and proportion of food. Next, all the operation was repeated again starting with fish "A" in order to have a second observation for this condition for all four marked fish (day 2). Overall the results of this procedure will becalled as 'this fish well-fed'. Finally, after a further five days of recuperation period, marked fish were again systematically isolated, but this time however, the separated fish (and its companions) were left without food whereas the rest of the experimental shoal was provided with the normal ration. The same procedure of separation, deprivation length and scoring was performed as with the above section. Overall this last procedure will be called as 'this fish hungry'. Mean water temperature of the tank during all the experiments was 15.2°C (14.8-15.7 95% confidence interval) RESULTSA total of 11,174 scores of position were obtained. In both conditions no significant interaction between day of observation, position in the shoal and fish was found (Tables 1 and 2). Results were then pooled by day and the histograms of frequencies are presented in Figs. 1 and 2. CONDITION 1Under a complete diet, scores of all fish marked over the four shoal positions were significantly different from a random pattern (Table 3). From Fig. 1 it is noticeable that either wellfed or hungry, fish "A" preferred a back-side position. Fish "B" was often swimming between neighbours and although fish "C" did not present an evident position when all well-fed, when all-hungry a back position was preferred. Finally, under complete diet, fish "D" clearly presented a strong forward swimming position with an increment of back positions when hungry. These presumptions of position-preference are supported by the results of the multi-factorial ANOVA, where more than 65% of total variability in the data is explained by the interaction between fish and position. When all fish well-fed, front position was positively correlated with fish length (r² = 0.75). However the slope was not significantly different from zero (analysis of covariance (ANCOVA), P>0.05). When all-hungry, the correlation was also positive (r² = 0.87), but the slope was again not different from zero (P>0.05). Back position was inversely correlated with fish length in both conditions (r² = 0.80 and r²= 0.73) and the slope was different from zero P<0.05) only when all fish were well-fed. Others positions were not significantly correlated with fish length (P<0.05). 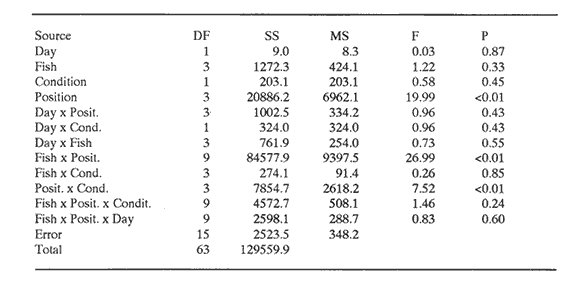 TABLE 1 MULTI-FACTORIAL ANOVA TESTING THE EFFECT OF INDIVIDUAL FISH, DAY OF OBSERVATION AND HUNGER CONDITION ('ALL FISH WEILL- FED' AND 'ALL FISH HUNGRY') ON NUMBER OF OBSERVATIONS REGISTERED FOR EACH POSITION Although multi-factorial ANOVA shows a significant interaction between position and condition, this was not the case all four fish, i.e. the interaction between fish, position and condition was not significant (P>0.05). CONDITION 2Under these two opposite situations, all but fish "A" when well-fed, presented a distribution significantly different from a random pattern (Table 3). Results of the multi-factorial ANOVA show a significant interaction between fish, position and condition (Table 2). A comparison between distribution of frequencies presented in Fig. 2 suggests that fish change location within the shoal accordingly to their hunger situation, and this is particularly noticeable for front and back positions. When a fish was well-fed and the rest hungry, front and back positions were correlated with fish length the first positively (r² = 0.71), the second negatively (r²= 0.98) (the slope was significantly different from zero (P<0.05 only for rear position). However, when a single fish was hungry, no significant correlation was found for either of these two positions. 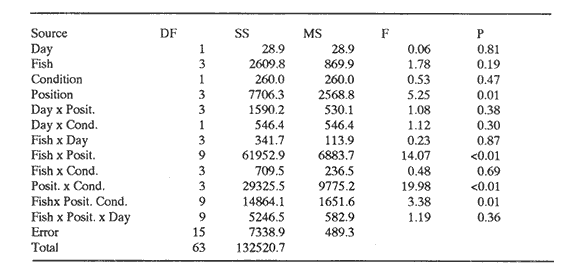 TABLE 2 MULTI-FACTORIAL ANOVA TESTING THE EFFECT OF INDIVIDUAL FISH, DAY OF OBSERVATION AND HUNGER CONDITION ('THIS WELL-FED' AND 'THIS HUNGRY') ON NUMBER OF OBSERVATIONS REGISTERED FOR EACH POSITION  TABLE 3 RESULTS A R X C TEST OF INDEPENDENCE USING G - TEST, VALUES OBSERVED WERE COMPARED AGAINST AN EQUAAL PROBABILITY ALONG POSITIONS, WF=ALL WELL FED, AH=ALL HUNGRRY, TW THIS WELL FED, TH=THIS HUNGRY. X²0.05[3]=7.81 DISCUSSIONAlthough observations were limited by the two-dimensional representation of the three-dimensional structure of the shoal (Partridge et al., 1980), the results indicate that the fish did not swim randomly, but particular positions were preferred. In contrast to former studies on individual position preference in fish shoaling, the present work shows that when all fish had the same hunger motivation, differences of position preference between the smaller and the larger fish were evident, in spite of the fact that the difference in length was only 19.5% (compare for instance the range of size analysed by Healey and Prieston, 1973; Partridge, 1978; Pitcher, et al., 1982). Size sorting behaviour in fish shoals, has been reported in European cyprinids. Pitcher et al., (1986) found that, although small and large European minnow respond very similar to the exposure of a predator threat, undisturbed small fish were found more often in their own. Theodorakis (1989) presents evidence that bluntnose minnow (Pimephales notatus), fathead minnow (P. promelas) and stroneroller minnow (Campostoma anomalus) segregate into groups of similar-sized individuals. Results in the present work demonstrate that when an individual fish was differently hunger motivated from the others significant differences of movements were presented regardless of the fish's length. However, due to the particular shortage of experiments done in this area, it remains a matter of speculation whether more frequent appearances in the front or back positions in a similarly hunger-motivated shoal may be explained only by a faster cruising speed by virtue of greater size or vice-versa. Results of the first, experiment in the present work support the sorting hypothesis. However, this may achieved only under certain circumstances (see below). EXISTENCE OF BETTER POSITION WITHIN A SHOALWhatever the hunger state of the individuals, but all fish under similar hunger motivation, fish swam more or less in the same positions. However, when a fish was well-fed and the rest hungry, the preference was clear, its frequency of appearance at the front of the shoal increased. Although it may be argued that this movement could be passive, i.e. fish appear to swim in front positions only because the others reduce their swimming speed, possibly for energy-saving reasons (Robinson and Pitcher, 1989), the fact that a well-fed fish is capable of reducing its swimming speed according to its own decision makes this hypothesis unlikely. Whether an 'optimal' location within a school does exist and whether individuals within the group gain advantage of its existence has not been proved (Parrish, 1989). According to the 'selfish herd' theory (Hamilton, 1971), a fish should be positioned in the best location of the group regardless of what the others are doing. In a shoal of fish, unless all positions within the group are equally advantageous, a situation which is particularly questionable (e.g. Parrish, 1989; Major, 1978; 0'Connell, 1972; Eggers, 1976), a fish would locate in the best position. The question is what position and why? Guided by the results of work on fish shoaling behaviour, together with theoreotical observations from some authors, it is possible to associate possible costs and benefits for an individual fish when swimming in some regions within the shoal. FRONTFront positions have been mentioned as 'advantageous' for planktivorous fish as they are the first to encounter particles of food (O'Connell, 1972; Eggers, 1976). Parrish (1989) reported that in a shoal of silverside fish (Menidia menidia), members at the front of the shoal were at significantly lower risk than at any other position within the shoal when attacked by the black seabass (Centropristis striata). Although no valid evidence of hydrodynamical advantage in travelling schools has yet been found (e.g. Partridge and Pitcher, 1979), the idea remains attractive and probable (Pitcher,1986). However, any advantage attributed to hydrodynamic aspects in a schools (Breder, 1 965; Belyayev and Zuyev, 1969; Weihs, 1973) may be exploited by fish swimming in any other but front position. 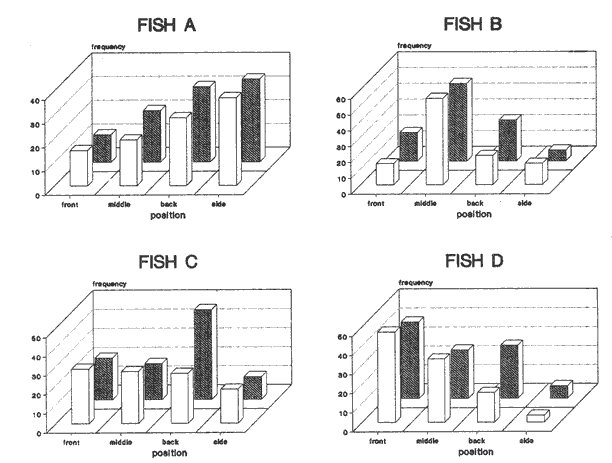 Figure 1. Frequency distribution of scores obtained in four positions during two different hunger conditions. White bars: all well-fed. Black bars: all hungry. Observations were obtained from four different fish in a school of ten herring. MIDDLEThe 'selfish herd' theory (Hamilton, 1971) suggests that individuals towards the centre positions are safer from an predator's attack, compared with those in the periphery. However, the hypothesis was recently re-examined by Parrish (1989) in a shoal of silverside fish. She found that within the body of the shoal (i.e. omitting stragglers, and fish in the periphery) individuals at the centre incurred the highest percentage of risk. Moreover, observations made by Hobson (1963) on the interaction between the gafftopsail pompano (Trachinotus rhodopus) and a shoal of flatiron herring (Harengula thrissina) showed that predators tended to attack the densest part or the main body of the group. Milinski (1977a) investigated whether different regions of water fleas (Daphnia magna) were differently preferred when attacked by the three-spined stickleback (Gasterosteus aculealus). His observations demonstrated that hungry sticklebacks preferentially struck the densest region of the swarm. However, when less hungry, the predator preferred stragglers or individuais at the periphery. Furthermore, due to the competition for food from surrounding companions, central positions would not be the best location to be when food is scarce. However, individuals swimming there may be rewarded by the hydrodynamical advantages produced by the vortices emitted by preceding fish (Breder, 1965; Belyayev and Zuyev, 1969; Weihs, 1973). 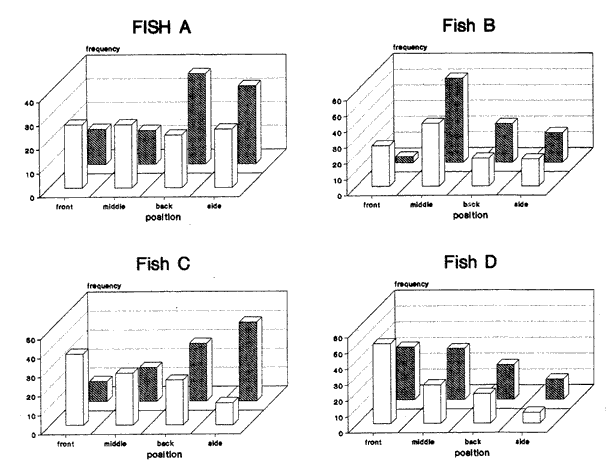 Figure 2. Frequency distribution of scores obtained in four positions during two different condifions. Black bars: when this particular fish was well-fed and the rest hungry. White bars: when this particular fish was hungry and the rest weIl-fed. Observations were obtained from four different fish in a school of ten herring. SIDESwimming in the side positions may be advantageous as a way to reduce the hazard of internal competition as the necessity of finding food increases. Individuals may detect a possible predator earlier and have an advantage for it (Charnov and Krebs, 1975). However, results of several experiments and observations (e.g. Neill and Cullen, 1974; Major, 1978) have demonstrated that being in the periphery of a group highly increases risk of predation. Furthermore the probabilities of being a 'straggler' as the fish swims in this position may increases, condition which is particularly vulnerable when a predator strikes (Milinski, 1977a,b). Additionally, advantages from hydrodynamical factors in these positions are reduced (Weihs, 1973). BACKFinally, the rear position, where according to the observations made by Eggers (1976), the probability of finding particles of food, compared to those fish swimming at the front is significantly lower. Parrish (1989) found that silverside herring located at the back of the shoal where at highest risk from predation compared with those in the front or in the middle. Pitcher and Wyche (1983) described how a 'confusion zone' was created when two schools of sand-eels (Ammodytes sp.) joined. This area was mainly at the rear of the union. The authors suggested that the lack of co-ordination in the 'confusion zone' may put individuals swimmig there in a high risk of predation. However, if hydrodynamical advantages do exist, fish will certainly benefit in this position. COSTS/BENEFITS ANALYSIS AND HUNGERHaving presented the possible cost and benefits for each region of the shoal, it is possible to assume that front position in a travelling shoal should be highly desirable for both a better position for an encounter with food and a relatively safe position. However, at least two constraints may impede an individual for occupying this position, one is the competition from other members, the other is the resistance of the water which should be higher relative to other parts of the shoal. Therefore, is possible that in order to benefit from swimming in this position, an individual should be among the fittest in the group in order to be able to present a swimming speed relatively faster than the others. However, this does not necessarily means the largest fish. Although the optimal cruising speed of fish increases in a shallow curve with body length (Wardle, 1975), constraints imposed when food resources are depleted may alter this pattern (Priede, 1985; Robinson and Pitcher, 1989). Accordingly, in the present work, when all fish were well-fed the benefits of swimming in front position may be achieved by the largest fish only for hydrodynamical aspects, condition that did not change when all fish were hungry i.e. the benefits of swimming at the front remain unchanged. Results when individual fish were differently motivated from the rest of the shoal support the hypothesis that whatever the length and as long as its fitness allowed it, a fish will tend to swim in a front positions. Other less advantageous positions as side and mainly back were relegated to the hungriest and consequently less fit fish. ConclusionesFrom the results of the observations presented in this work the following conclusions can be presented: Fish in a structured and polarized group may not swim randomly, but specific positions may be chosen. According to observations and results of other experiments, it seems that front positions provide further benefits to a shoal's members. Results in this work support the hypothesis. The supposed benefits of swimming at the front may be achieved if the fish is fit enough to maintain swimming in this position. However, it is evident that fish are able to control their swimming performance. If not, fish swimming at the front would be easily become a stragler, for instance, if the rest of the group slow down. Competition from other members and the energy necessary to maintain a stable swimming speed may constrain a fish to swim there. Accordingly, any fish within the shoal may be a candidate regardless of its length (within certain limits). Consequently other positions within the shoal may be occupied by any other fish. AgradecimientosWe thank Dr. T. J. Pitcher for early criticism on the manuscript. This work forms part of the requirements for CJ.R`s doctoral degree and was supported by UNAM, Mexico. The experimental work was done at the Univeristy of Wales, Bangor. Animal Biology Department. LITERATURABAYLIFF, W., Fish. Bull. U.S., Integrity of schools of skipjack tuna, Katsuwonus pelamis, in the eastern pacific ocean, as determined from tagging data. 1988. 631-643. 86. BELYAYEV, V.V. AND ZUYEV, G.V., J. Icthyol. Hydrodynamic hypothesis of school formation in fishes 1969. 589-594. 9: BREDER, C.M., JR., Zoologica Vortices and fish schools. 1965. 97-114. 50. CHARNOV, E.L. AND KREBS, J.R., Am. Nat., The evolution of alarm calls: altruism or manipulation. 1975. 107-112. 109: EGGERS, D.M., J. Fish. Res. Bd Can, Theoretical effect of schooling by planktivorous fish predators on rate of prey consumption. 1976. 1964-1971. 33: GILES, N. AND HUNTINGFORD, FA., L. Anim. Behav., Predation risk and interpopulation variation in anti-predator behaviour in the three-spined stickleback, Gasterosteus aculeatus 1984. 264-275. 32: HAMILTON, W.D., J. Theor. Biol., Geometry for the selfish herd. 1971. 295-311. 31: HEALEY, M.C. AND PRIESTON, R., Fish. Res. Bd Can. Tech. Report., The interrelationships among individuals in a fish school. 1973. 1-15. 389: HELFMAN,G.S., Anim. Behav., School fidelity in fishes: the yellow perch paltern. 1984. 663-672. 32: HOBSON, E.S. Copeia Selective feeding by the gafftopsail pompano Trachinotus rhodopus (Gill), in mixed school of herring and anchovies in the gulf of California. 1963. 595-596. KENNEDY, G.J.A., Anim. Behav., Individual variation in homing tendency in the European minnow, Phoxinus phoxinus (L). 1981. 621-625. 29: MAGURRAN, A.E., The behaviour of teleost fishes. Individual differences in fish behaviour. In: (Ed. by TJ. Pitcher), London:Croom Helm. 1986. 553 pp. 338-365. MAJOR, P.E, Anim. Behav. Predator-prey interactions in two schooling fishes Caranx ignobilis and Stolephorus purpureus. 1978. 760-777. 26: MCFARLAND, D., Animal behaviour. Psychobiology, Ethology and Evolution. Pitman. London: 1985. 576 p. MILINSKI, M. Z. Tierpsychol., Experiments on the selection by predators against spatial oddity of their prey. 1977a. 311- 325. 43: MILINSKI, M., Z. Tierpsychol., Do all members of a swarm suffer the same predation? 1977b. 373-388. 45: NEILL, S.R. AND CULLEN, J.M., J. Zool. Lond, Experiments on whether schooling by their prey affects the hunting behaviour of cephalopods and fish predators. 1974. 549-569. 172: O'CONNELL, C.P., J. Fish. Res. Bd. Can., The interrelation of biting and filtering in the feeding activity of the northern anchovy (Engraulis mordax). 1972. 285-293. 29: PARRISH, J.K., Anim. Behav., Re-examining the selfish herd: are central fish safer? 1989. 1048-1053. 38: PARTRIDGE, B.L., Ph.D. Thesis, Sensory aspects of schooling. University of Oxford. U.K. 1978. 450 p. PARTRIDGE, B.L. AND PITCHER, T.J., Nature, Evidence against a hydrodynamic function for fish schools. 1979. 418-419. 279: PARTRIDGE,B.L., PITCHER,T.J.,CULLEN,J.M. AND WILSON,J., Behav Ecol. Sociobiol., The three-dimensional structure of fish schools. 1980. 277-288. 6: PITCHER, T.J., Anim. Behav. Heuristic definitions of fish shoaling behaviour. 1983. 611-612. 31: PITCHER, TJ. The behaviour of teleost fishes., Functions of shoaling behaviour in teleosts. In: (Ed. by TJ. Pitcher), London: Croom-Helm. 1986. pp. 294-337. 553 p. PITCHER, TJ., WYCHIE,C.J. AND MAGURRAN, A.E., Anim. Behav., Evidence for position preferences in schooling mackerel. 1982. 932-934. 30: PITCHER, TJ. AND WYCHISCJ. Predator and prey in fishes Predator-avoidance behaviors of sand-eel schools: why schools seldom split In: (Ed. by D.L.G. Noakes,D.G. Lindquist,G.S. Helfman and J.A.Ward), The Hague:Dr. W Junk. 1983. pp. 193- 204. PITCHER, TJ., MAGURRAN,A.E. AND EDUARDS, J..I., Mar. Biol., Schooling mackerel and herring choose neighbours of similar size. 1985. 319-322. 86: PITCHER, T.J., MAGURRAN, A.E. AND ALLAN, J.R., J. Fish. Biol., Sizesegregative behaviour in minnow shoals. 1986. 83-95. 29: PRIEDE, I.G., Fish energetics new perspectives Metabolic scope in fishes. In: (Ed. by P. Tyler and P. Calow), London:Croom-Helm. 1985. pp. 33-64. 352 p. ROBINSON,CJ. AND PITCHER,T..J., L. J. Fish. Biol, The influence of hunger and ration level on shoal density, polarization and swimming speed of herring, Clupea harengus 1989. 631-633. 34: SEGHERS, B.H., Evolution, Schooling behaviour in the guppy (Poecilia reticulata): an evolutionary response to predation. 1974. 486-489. 28: STREET,N.E., MAGURRAN,A.E. AND PITCHER,T.J., J. Fish Biol., The effecls of increasing shoal size on handling time in goldfish, Carassius auratus L. 1984. 561-566. 25: THEODORAKIS, C. W., Anim. Behav., Size segregation and the effects of oddity on predation risk in minnow schools. 1989. 496-502. 38: WARDLE, C.S., Nature, Limit of fish swimming speed. London, 1975. 255; 725-727. WEIHS, D., Nature, Hydromechanics of fish schooling. London, 1973. 241,290-291.
|