|
ANÁLISIS DE LOS HÁBITOS ALIMENTARIOS Y DEL
APORTE DEL CARBONO DE LAS PRESAS A LA DIETA DE LA SARDINA CRINUDA, Opisthonema
libertate, GUNTHER 1866 (PISCES: CLUPEIDAE) EN EL GOLFO DE CALIFORNIA
Recibido el 11 de diciembre de 1989 y aceptado para su publicación
el 19 de junio de 1991
Manuel Gallardo-Cabello
Mercedes Jacob-Cervantes
Xavier Chiappa-Carrara
Instituto de Ciencias del Mar y Limnología,
Universidad Nacional Autónoma de México. Apartado Postal 70-305,
México, D. F. 04510 MÉXICO Contribución No. 707 del
Instituto de Ciencias del Mar y Limnología.
En este estudio se analizan los hábitos y el
régimen alimentarios de la sardina crinuda Opisthonema
libertate en el Golfo de California. Asimismo se reportan datos sobre
la relación entre el tamaño del depredador y el de la presa, la
intensidad de la alimentación en relación a los períodos
diurnos y nocturnos y la contribución del carbono a la dieta. Las
diatomeas, los crustáceos, los huevecillos de los peces, los
dinoflagelados y los protozoarios constituyen los principales componentes en
número. El tamaño del alimento que esta especie filtra
corresponde a un intervalo entre 0.10 mm y 0.75 mm, mientras que el que ataca a
un rango entre 0.50 mm y 1.40 mm de la longitud de la presa. 0.
libertate se alimenta con mayor intensidad en las horas del atardecer
y del alba. La relación entre la longitud del intestino y la del pez
mostró un crecimiento alométrico positivo. Los huevecillos de los
peces y los crustáceos representan los principales contribuyentes del
carbono a la dicta de esta especie, mostrando diferencias en relación a
las clases de tallas. Conforme se incrementa la longitud de la sardina crinuda,
aumenta la longitud de sus presas y varía la proporción de los
grupos de los organismos de los que se alimenta.
In the present paper, feeding behaviour and dietary
composition of thread herring Opisthonoma libertate along
the coasts of Gulf of Califonia were analyzed. Predator-prey relationship,
feeding hours of mayor intensity and carbon contribution of the dietary
components were also studied. Diatoms, crustaceans, fish eggs, dinofiagellates
and protozoans were the mayor -by number- contributors to the 0.
libertate diet. Filtering mechanisms acting over a 0.10 mm lo 0.75 mm
range of prey length, while selective particulate feeding occurs through 0.50
mm to 1.40 prey size spectrum. Major feeding intensity occurs during dusk and
dawn. The morphometric relationship between intestine length and fish standard
length showed an alornetric positive growth. Fish eggs and crustaceans were the
mayor carbon contributors to the thread herring diet. Prey size increases as
predator growths.
Las distintas especies de peces pelágicos menores -clupeidos y engraúlidos, principalmente- constituyen tanto desde el punto de vista ecológico como del pesquero organismos de gran importancia. Ello es debido a que en el ambiente marino representan el alimento de un gran número de especies de peces, moluscos, aves y mamíferos marinos y en el medio pesquero constituyen algunos de los recursos de mayor importancia económica (Holt, 1975). En México, la sardina representa, de acuerdo con las cifras estadísticas de la Secretaría de Pesca, aproximadamente el 30% de la captura total anual. No obstante, el número de trabajos publicados acerca del ciclo de vida y de la dinámica de las poblaciones de las distintas especies de sardinas es, en términos generales, escaso. (Lewis, 1929; Cadet y Berner, 1959; García y Molina, 1986 y Gallardo-Cabello et al., en prensa). En base a lo anteriormente expuesto, en el presente estudio, se llevó a cabo un análisis sobre los hábitos y la dieta alimentarios de la sardina crinuda Opisthonema libertate en el Golfo de California. Asimismo, es importante mencionar que en este trabajo se tratan, por primera vez, temas con respecto a los hábitos alimentarios de esta especie, la relación entre el tamaño del depredador y el de la presa, la digestión del alimento y el contenido del carbono que las presas aportan a la dieta de O. libertate. Asimismo, es importante señalar, que los estudios relacionados con los porcentajes del carbono que las presas proveen en el régimen alimentario de un depredador, proporcionan una visión de conjunto que no es posible obtener por medio de la aplicación exclusiva de los métodos tradicionales (porcentaje numérico y frecuencia de ocurrencia) para el análisis de la dieta de un organismo. En este estudio, se observó, entre otras cosas, que los organismos que 0. libertate consume en mayor cantidad y con mas frecuencia, no son los que aportan un mayor porcentaje de carbono a su dieta, sino que existen otros, que aunque menos abundantes en el contenido estomacal - como es el caso de los copépodos, los anfípodos, los ostrácodos y los eufásidos -representan una mayor contribución del carbono necesario para procesos tan importantes como son el crecimiento y la reproducción, principalmente. MATERIAL Y MÉTODOSSe obtuvieron un total de 140 ejemplares de la sardina crinuda Opisthonema libertate en el Golfo de California, desde la boca del río Mayo hasta la localidad denominada Cabeza de Caballo (Fig. 1). La identificación de los ejemplares se llevó a cabo de acuerdo con los criterios establecidos por Berry y Barret (1963). Se realizaron muestreos mensuales de la captura comercial, en cada una de las localidades, durante el período comprendido entre febrero y julio de 1987. De los ejemplares capturados se registraron la longitud estándar y el peso entero. Asimismo, se obtuvieron los tractos digestivos de los ejemplares que fueron conservados individualmente, primero, en formol neutralizado al 4% (Steedman, 1976) y posteriormente en alcohol al 70%. Se analizaron los organismos encontrados en el esófago y en el estómago. No se consideraron los intestinos para evitar errores relacionados con las diferentes tasas de la digestión de las presas (Tomas y Davidson, 1962; Gannon, 1976; Berg, 1979 y Hyslop, 1980). La identificación de los organismos se llevó a cabo mediante el empleo de una lupa binocular estereoscópica y un microscopio óptico. Todos los organismos encontrados en el contenido estomacal se identificaron hasta el menor nivel taxonómico posible, empleando las claves de Cupp (1943 y 1950), Tregouboff y Rose (1957) y Rose (1975). Se contaron las presas identificadas en el contenido estomacal de la sardina crinuda y se calcularon los índices de la frecuencia de ocurrencia de las especies (Hynes, 1950 y Hyslop, 1980) expresándose como el porcentaje de los estómagos que contienen dichas presas con respecto del total. Asimismo, se obtuvieron los porcentajes numéricos de las presas (Wihndell y Bowen, 1978). El grado de llenado de los estómagos de la sardina crinuda se determinó mediante la aplicación de un índice empírico de repleción gástrica de cuatro componentes, que considera el volumen del bolo alimenticio. La escala empleada es la siguiente: I Lleno 3/4 Capacidad total II Medio lleno 1/2 Capacidad total 3/4 III Medio vacío 1/4 Capacidad total 1/2 IV Vacío Capacidad total 1/4 Asimismo, se obtuvo el coeficiente de repleción gástrica, definido como el porcentaje del número de estómagos vacíos con respecto al total de estómagos. Las fases de la digestión del contenido estomacal se obtuvieron por medio de la aplicación de la escala propuesta por Carranza (1969) quien considera cuatro fases. Fase 1. Material recién ingerido. La digestión aún no se inicia, las presas pueden ser fácilmente identificadas. Fase 2. Inicio de la digestión. El proceso digestivo ha comenzado paro no está muy avanzado. El contenido puede ser determinado sin gran dificultad.  Figura 1. Localización Márea de estudios. Fase 3. Digestión avanzada. Muchas de las estructuras de las presas han sido destruidas, por lo que su identificación es solo parcial. Fase 4. Digestión concluida. El contenido estomacal está digerido y es irreconocible. Por otra parte, se obtuvieron los datos sobre la longitud (L) y el diámetro (D) de los cuerpos de las presas, a partir de los cuales se calculó el cociente D/L, que expresa la forma geométrica del organismo (Angelescu, 1982). También se obtuvo la longitud del intestino de cada uno de los ejemplares en relación a las clases de tallas, analizándose la relación entre la longitud estándar del pez y la longitud total del intestino por medio de una función potencial, calculada por el método de la regresión resistente (Tukey, 1977). Por último, se obtuvieron los porcentajes promedio del aporte del carbono de las presas a la dieta aplicando las relaciones propuestas por diversos autores. (Beers, 1966; Omori, 1969; Parsons et al, 1984; Verity y Langdon, 1984 y James, 1987). RESULTADOS Y DISCUSIÓN
INGESTIÓN DE ALIMENTOEn las tablas 1 y 2, se observan, respectivamente, los porcentajes numéricos de la ocurrencia y las frecuencias porcentuales de las ocurrencias de las especies encontradas en el contenido estomacal de O. libertate en relación a las clases de tallas. Se identificaron un total de 2 357 presas de las cuales 1 681 (el 71.32%) fueron reconocidas a nivel de especie; 263 (el 11.16%) a nivel de género; 216 (el 9.16%) a nivel de orden y 197 (el 8.3%) a niveles de grupos superiores al de orden. De acuerdo con lo anterior, se observa, que la mayor parte de los organismos del fitoplancton pudieron ser reconocidos a nivel de especie, mientras que los ejemplares del zooplancton fueron identificados a nivel de grandes grupos. Ello fue debido a que la mayoría de las diatomeas presentaron una mayor preservación a la acción de los ácidos gástricos con respecto a los grupos de los crustáceos, lo que hizo posible la identificación del fitoplancton a un menor nivel taxonómico en relación al zooplancton. Asimismo, en el caso de los crustáceos, se observó que su identificación a nivel del género y especie, resultaba dudosa, por lo que se prefirió reportar estos ejemplares a nivel de subclase y de orden según fue su caso.  TABLA 1 PORCENTAJE NUMÉRICO DE OCURRENCIA DE LAS ESPECIES ENCONTRADAS EN El CONTENIDO ESTOMACAL DE LA SARDINA CRINUDA, POR CLASES DE TALLA 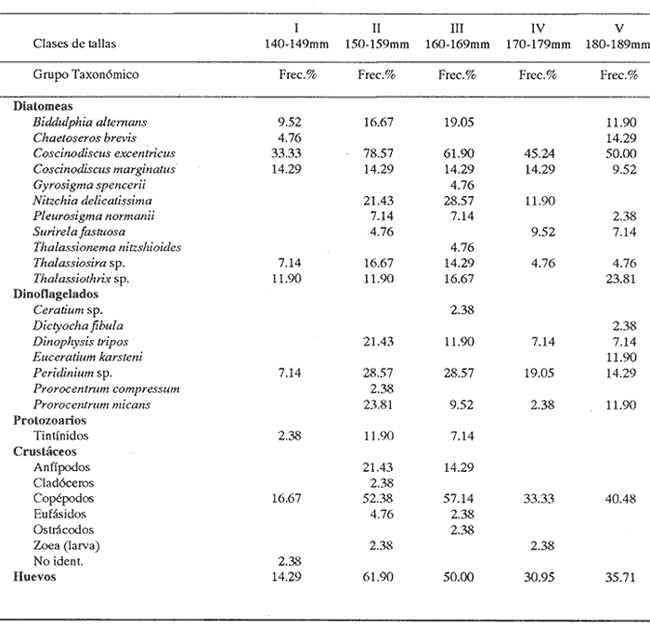 TABLA 2 FRECUENCIA DE OCURRENCIA DE LAS ESPECIES ENCONTRADAS EN El CONTENIDO ESTOMACAL DE LA SARDINA CRINUDA, POR CLASES DE TALLA Por otra parte, es importante mencionar, que el estado de la digestión en que se presenta una presa en el contenido estomacal, se debe tanto a su tiempo de permanencia en el estómago del depredador como a su propia naturaleza. Así organismos que presentan exoesqueletos - compuestos por quitina o substancias calcáreas y silíceas - ofrecen un mayor tiempo de conservación - y por lo tanto una mayor posibilidad de identificación - aunque su ingestión no haya sido reciente. Por el contrario, los organismos que no poseen estructuras duras son digeridas con una mayor velocidad en relación a los mencionados anteriormente (Gual-Frau y Gallardo-Cabello, 1988). Tal es el caso de los poliquetos, los tunicados larváceos y los quetognatos, que no se encontraron en el contenido estomacal de 0. libertate. Por lo tanto, es importante mencionar, que aunque los quetognatos alcanzan una gran abundancia en el medio marino (Feigenbaum y Maris, 1984) su digestión en el contenido estomacal del depredador es muy rápida, preservándose, solamente y en ocasiones, las mandíbulas, y considerándose el resto del cuerpo del animal en una categoría indeterminada (Rice, 1963). Un fenómeno semejante ocurre con los poliquetos y los tunicados larváceos. En relación a estos organismos, Rice (1963) encuentra en el contenido estomacal de Clupea harengus L., en el Mar del Norte, que los tunicados ocurren en un 4.6%, los poliquetos en un 0.2% (solo un organismo del género Phyllodoce) y los quetognatos en 0.1%. Este autor considera que los quetognatos debieron representar, por lo menos, el 1% del total de los organismos encontrados en el contenido estomal de C. harengus. De acuerdo con lo anteriormente expuesto, cabe esperar que en los resultados obtenidos en el presente estudio, los organismos más lábiles no estén debidamente representados, o bien, se presenten subestimados en cuanto a su proporción numérica en el contenido estomacal O. libertate. En la figura 2, se observa que las diatomeas ocupan el mayor porcentaje numérico en relación a los principales grupos de presas encontradas en el contenido estomacal de la sardina crinuda. Le siguen en orden de importancia los crustáceos, los huevecillos de los peces y los dinoflagelados. Los protozoarios representan valores bastante menores con respecto al total de las presas encontradas para los otros grupos. 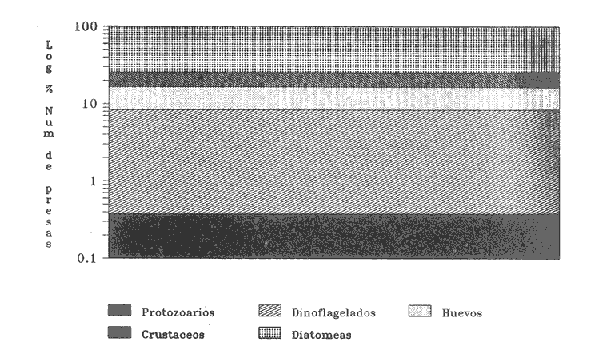 Figura 2. Valores M porcentaje numérico de ocurrencia de los grupos de presas encontradas es el contenido estomacal de O. libertae. De acuerdo con lo anterior, es importante notar (Tablas 1 y 2) que tan solo Coscinodiscus excentricus ocupa los mayores porcentajes (entre el 39.77% y el 53.22%, para las longitudes consideradas) del total de las presas que constituyen la dieta de la sardina crinuda. Similarmente, los valores de la frecuencia de ocurrencia de C. excentricus en el contenido estomacal de O. libertate son los mayores: entre el 33.33% y el 78.57% del total del presas ingeridas. En términos generales y considerando exclusivamente sus valores numéricos, también son importantes en la dieta alimentaria otras especies de diatomeas como son: Coscinodiscus marginatus, Nitzschia delicatissima, y el género Thalassiothrix. Los copépodos muestran importantes valores tanto del porcentaje numérico de ocurrencia (entre el 4.1% y el 8.6%) como la frecuencia porcentual de ocurrencia (entre el 16.67% y el 57.14%) con respecto del total de las presas identificadas. Similarmente, los huevecillos de los peces presentan valores significativos: entre el 5.26% y el 10.98% en relación al porcentaje numérico, y entre el 14.29% y el 61.90% con respecto a la frecuencia de ocurrencia del total de las especies encontradas. En relación a los dinoflagelados, destaca la importancia del género peridinium en la dicta de O. libertate. Con respecto a los protozoarios, se observa que aunque no ocupan importantes porcentajes numéricos en la dicta, muestran valores no del todo despreciables, con respecto a los índices de la frecuencia de ocurrencia en los ejemplares de la sardina crinuda de algunas clases de tallas, lo que puede indicar que no son tan abundantes en el medio marino como los organismos de otros grupos. RELACIÓN ENTRE El TAMAÑO DEL DEPREDADOR Y EL DE LA PRESAEn la tabla 3, se muestran los datos obtenidos sobre la longitud (L) y el diámetro (D) del cuerpo de las presas, así como la forma de las especies ingeridas (relación D/L) por la sardina crinuda. El análisis de las presas encontradas en el contenido estomacal de O. libertate mostró, en términos generales, que la mayor parte de ¡os organismos cuya longitud correspondía a un intervalo entre 0.10 mm y 0.75 mm, se encontraban enteros en el estómago de la sardina crinuda, mientras que las especies cuya talla oscilaba entre 0.50 mm y 1.40 mm se presentaban fragmentadas en el tracto digestivo de este depredador, mostrando signos (distintos de aquellos producidos por la acción de las contracciones musculares del tubo digestivo) de haber sido atacadas y fraccionadas por medio de mordidas para ser ingeridas, en todas sus partes, posteriormente. 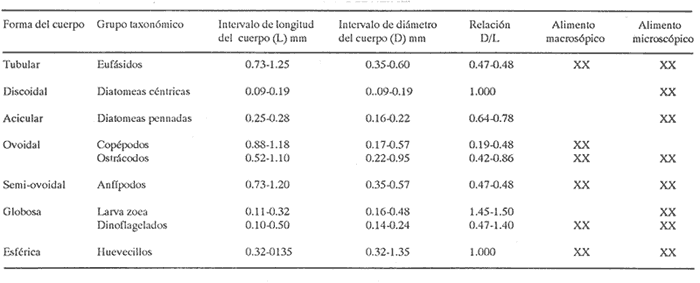 TABLA 3 LONGITUD, DIÁMETRO Y RELACIÓN D/L DEL CUERPO DE LOS GRUPOS DE LAS PRESAS DE Opisthonema libertate. ASI MISMO, SE MUESTRA LA CLASIFICACIÓN DEL ALIMENTO DE ACUERDO CON SU TAMAÑO De acuerdo con lo anterior, puede considerarse que O. libertate utiliza dos procesos para la captura de su alimento: la filtración y el ataque a las presas por medio del mordisqueo. Por lo tanto se estableció que el tamaño del alimento que se filtra, corresponde a un intervalo entre 0.10 mm y 0.75 mm y el que se ataca a un rango entre 0.50 mm y 1.40 mm de la longitud de la presa. Así, se observa que para la captura de las diatomeas y de las larvas de los crustáceos, la sardina crinuda emplea, como tipo de alimentación: la filtración. Cuando se trata de los dinoflagelados, los huevecillos de los peces y de los crustáceos en general, O. libertate puede capturarlos tanto por medio de la filtración como del mordisqueo, dependiendo del tamaño de la presa. Resultados similares han sido encontrados en el estudio de la alimentación de otras especies de peces pelágicos menores, como son los engraúlidos por diversos autores (Leong y O'Connell, 1969; O'Connell, 1972 y Chiappa-Carrara, 1988). Asimismo, es importante señalar, que los procesos para la captura del alimento -filtración y mordisqueo- de estas especies pelágicas están en relación, tanto con la disponibilidad del alimento como con la relación entre el tamaño del depredador y el de la presa (Leong y O'Connell, 1969 y O'Connell, 1972). INDICES DE LA REPLECIÓN GÁSTRICA Y DE LAS FASES DE LA DIGESTIÓN DE LA PRESAS EN EL CONTENIDO ESTOMACAL DE O. libertateLa relación encontrada entre el índice empírico de la repleción gástrica y el peso del material ingerido es la siguiente: I Lleno 5.67 g <= Peso del contenido II Medio lleno 3.78 g <= Peso del contenido = 5.66 g III Medio vacío 1.89 g <= Peso del contenido = 3.77 g IV Vacío Peso del contenido = 1.88 g Asimismo, se obtuvo el peso del alimento en relación al porcentaje del peso del pez (Hureau, 1.969): I lleno 9% = Peso total II Medio lleno 6% = Peso total s 8.9% III Medio vacío 3% = Peso total s 5.9% IV Vacío Peso total = 2.9% De acuerdo con lo anterior, se observó que el 43.4% de los estómagos analizados se presentaron en el estadio I, es decir, que el peso del contenido fue igual o mayor al 9%. En el estadio II, se observaron el 9.42% de los estómagos, con una relación entre el peso del pez y el del alimento entre el 6.0% y el 8.9%. En el estadio III se encontraron el 32,6% de los estómagos, con una relación entre los pesos del 3.0% al 5.9%, y por último, el 11.59% de los estómagos se observaron en el estadio IV, con una relación entre los pesos inferior al 2.9%. El coeficiente de repleción gástrica calculado fue de 0.107, por lo tanto, se observó que el número de estómagos vacíos no fue considerable. En relación a los índices de las fases de la digestión del contenido estomacal se encontraron los siguientes resultados: para la fase 2 el porcentaje de los estómagos fue del 45%, para la fase 3 del 53% y del 2% para la fase 4. No se encontraron organismos recién ingeridos, es decir, en la fase 1, en el contenido estomacal de 0. libertate, debido a que la digestión se inicia en el momento en que el alimento llega a la cavidad estomacal. INTENSIDAD DE LA ALIMENTACIÓN EN RELACIÓN A LOS PERIODOS DIURNOS O NOCTURNOSDe acuerdo con lo anteriormente expuesto, se observa que la mayor parte de los estómagos se encontraban llenos y que las presas que contenían se presentaron en el inicio de las fases de la digestión. Ello es debido, a que las capturas comerciales de 0. libertate se llevan a cabo al atardecer y durante el alba, horas en las que la sardina crinuda está cerca de la superficie para alimentarse. Resultados similares son reportados para otros organismos pelágicos por diversos autores (Baxter, 1967; Loukashkin, 1970; Rojas de Mendiola, 1971; Angelescu y Angnuzzi, 1986; James, 1987 y Chiappa-Carrara, 1988) quienes establecen que las diferentes especies de anchovetas se alimentan durante el alba. Asimismo, es importante señalar que durante el análisis de los contenidos estomacales de la sardina crinuda, se observó que las presas microscópicas se disponían por capas alternadas de acuerdo con su tamaño en el bolo alimenticio, lo que puede indicar la existencia de una secuencia durante los procesos de la captura del alimento, es decir, entre las presas que se filtran y las que se atacan por medio del mordisqueo. Por lo que es muy probable que la ingestión de la mayor parte de los organismos macroscópicos como son: los copépodos, los anfipodos, los ostrácodos y los eufásidos por O. libertate ocurra durante las horas de mediana intensidad luminosa, que es cuando los organismos del zooplancton realizan migraciones verticales a las capas superficiales para alimentarse del fitoplancton. Dicho fenómeno ha sido observado en otros peces pelágicos que realizan una activa búsqueda y captura de presas macroscópicas durante las horas del alba (Leong y O'Connell, 1969; Rojas de Mendiola, 1971; Angelescu, 1982 y Chiappa-Carrara, 1988). Asimismo, Blaxter (1966) establece, a través de observaciones tanto en acuarios como en el mar, que Clupea harengus (al igual que otros peces pelágicos) es un depredador visual, en el cual el sentido de la visión juega un papel determinante para la detección de las presas. Por lo que la alimentación de estos organismos pelágicos, alcanza su mayor intensidad durante las horas del alba y del atardecer. DESARROLLO MORFOMÉTRICO DEL INTESTINOLa relación alométrica encontrada entre la longitud del intestino (Li) y la longitud estándar de la sardina crinuda (Lt) es la expresada por la siguiente ecuación: Li = 0.00057 Lt 2,4149 El valor del índice de regresión b = 2.4149 muestra un crecimiento alométrico positivo. De acuerdo con lo anterior, a continuación se muestran los valores de la longitud del intestino (Li) en relación a las clases de tallas de O. libertate. DIGESTIÓN DE LAS PRESASFitoplancton. En el estómago cardíaco de 0. libertate se observó que el fitoplancion se presenta como un conglomerado en el que aparece otro tipo de material orgánico no identificable. Las especies de las diatomeas que resisten mas el ataque de los jugos gástricos son: Biddulphia alternans, Coscinodiscus excentricus, C. marginatus, Nitzschia delicatissima y Surirella fastuosa, así como los géneros Thalassiosira y Thalassiothrix. En estos casos, se observó que la mayoría de estos organismos se encontraban enteros, sin que hubiese separación o fragmentación de la epiteca e hipoteca ni disolución del contenido citoplasmático. Por otra parte, las especies: Chaetoseros brevis, Gyrosigina spencerii, Pleurosigma normanii y Thalassionema nitzschiodes mostraron una menor resistencia a los procesos digestivos en el contenido estomacal de O. libertale. En relación a los dinoflagelados se observó que el género Peridinium es el mas resistente a la descomposición producida por los ácidos grasos. Las especies como Dinophysis tripos y Prorocentrum micans aparecen generalmente fragmentadas en el contenido estomacal. En el estómago pilórico de la sardina crinuda, la mayoría de las diatomeas presentaron un rompimiento de las valvas y la dispersión del citoplasma y de los cloroplastos, que en la mayor parte de los casos, solo se localizó en el extremo de la hipoteca. Ello es debido fundamentalmente, a la acción del tejido muscular de las paredes del estómago que fragmenta las valvas de estos organismos, no obstante su dureza debida a la depositación del sílice en sus fústulas Tregouboff (1957). Asimismo, se observan, en el estómago pilórico de 0. libertate, las células rotas de algunos dinoflagelados como es el caso del género Peridinium y de Prorocentrum micans. En el intestino se finalizan los procesos digestivos, por lo que resulta imposible identificar la mayor parte de las especies del fitoplancton. Solamente pueden observarse los fragmentos de las valvas de algunas diatomeas. Zooplancton. La mayor parte de las presas del zooplancton encontradas en el contenido estomacal de la sardina crinuda correspondían a los crustáceos. Los copépodos y los anfípodos fueron los organismos que - aunque fragmentados por los ataques del depredador - se mostraron mejor preservados con respecto al total de las presas de este grupo. Durante el principio de la digestión (zona cardíaca) se observó que -debido a la acción mecánica de los pliegues musculares de las paredes del estómago - se producía una ruptura del cefalotórax de estas presas, que provocaba el que se vaciase su contenido somático casi en su totalidad. De esta manera, estos organismos se observaron completamente transparentes. Conforme continúan los procesos digestivos en las partes posteriores del estómago (zona pilórica) se desprenden del cefalotórax de estos crustáceos tanto los segmentos abdominales como los apéndices, debido, principalmente, a la acción enzimática de los ácidos digestivos. En el intestino termina la digestión pero aún es posible reconocer los restos de algunos organismos, como es el caso de las mandíbulas de los quetognatos (Feigenbaum y Maris, 1984) algunas estructuras de los tunicados larváceos (Shelborne, 1962) y las mandíbulas con proyecciones de siliceo de los copépodos (Sullivan et al., 1975). En términos generales, se observó que el zooplancton (crustáceos) a diferencia del fitoplancton (diatomeas) es susceptible a ser desintegrado mas rápidamente por los procesos de la digestión. Ello es debido, fundamentalmente, al mayor tamaño de los crustáceos y a que su exoesqueleto, aunque es duro debido a la presencia de las sales de calcio, es menos resistente que las valvas silíceas de las diatomeas a la acción de los jugos digestivos en el estómago. CONTENIDO DEL CARBONO PROVISTO POR LAS PRESAS A LA DIETA DE O. libertateEn la figura 3, se muestra la relación entre las clases de tallas de la sardina crinuda y los logaritmos de los porcentajes promedio de los aportes del carbono de las presas a la dieta. 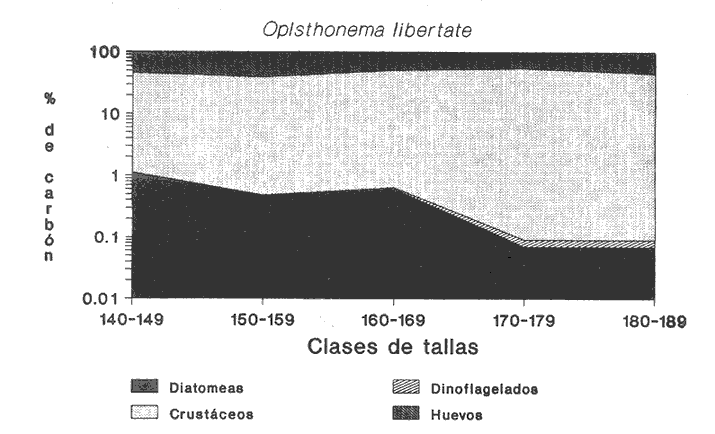 Figura 3. Relación entre el porcentaje del carbono de los principales grupos de presas y las clases de tallas de la sardina crinuda O. libertate. Los huevecillos de los peces constituyen uno de los principales contribuyentes del carbono a la dieta de la sardina crinuda, en ejemplares de 140 mm y de 150 mm de longitud estándar. Conforme a la talla de O. libertate aumenta, se incrementa la importancia de los crustáceos en los individuos de 160 mm y 170 mm de longitud estándar, en los cuales estos organismos aportan los mayores valores del total del carbono provisto a la dieta. La importancia de los crustáceos disminuye en ejemplares de 180 mm de longitud estándar siendo reemplazados por los huevecillos de los peces. Aunque las diatomeas constituyen el mayor aporte en número de individuos y de especies a la dieta alimentaria de la sardina crinuda (Tabla 1 y figura 2) son las que con menos cantidad de carbono contribuyen a la dieta de O. libertate en relación a los crustáceos y a los huevecillos de los peces. En la figura 4, se presenta la relación entre las clases de tallas de O. libertate y los logaritmos de los porcentajes promedio de los aportes del carbono de los grupos de presas de acuerdo con su tamaño. En relación a lo anterior, se observa que conforme aumenta la longitud de la sardina crinuda aumenta el tamaño de su presas. Por lo que la importancia de los huevecillos de los peces es reemplazada, gradualmente, por los copépodos, los anfipodos, los ostrácodos y los eufásidos. Las diatomeas y los dinoflagelados son mas abundantes en los ejemplares de menor talla. 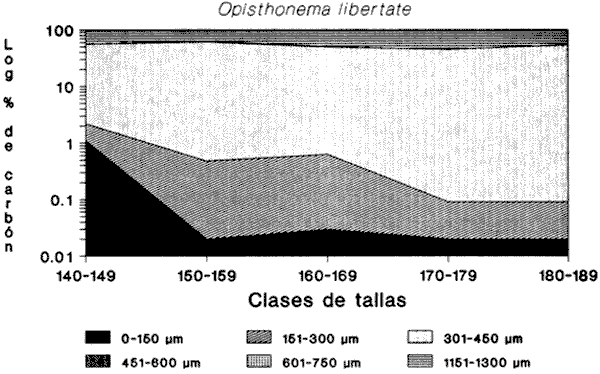 Figura 4. Relación entre el porcentaje del carbono de los distintos tamaños de las presas y las clases de tallas de la sardina crinuda O. libertate. ConclusionesConsiderando exclusivamente el porcentaje numérico, las diatomeas ocupan la mayor proporción en la dieta de O. libertate, estando representadas en un 47% por una sola especie: Coscinodiscus excentricus. Los crustáceos, los huevecillos de los peces, los dinoflagelados y los protozoarios, constituyen en orden de importancia (con respecto al porcentaje numérico) el resto de los componentes en el régimen alimentario de la sardina crinuda. El tamaño del alimento que esta especie filtra, corresponde a un intervalo entre 0.10 mm y 0.75 mm y el que este organismo ataca a un rango entre 0.50 mm y 1.40 mm de la longitud de la presa. La sardina crinuda emplea dos procesos para la captura de sus presas: la filtración y el ataque por medio del mordisqueo, por medio del primer mecanismo ingiere a las diatomeas y a las larvas de los crustáceos. Cuando se trata de los dinoflagelados, los huevecillos de los peces, los copépodos, los anfípodos, los ostrácodos y los eufásidos emplea tanto la filtración como el mordisqueo dependiendo del tamaño de la presa. Es muy probable que O. libertate se alimente con mayor intensidad durante las horas del atardecer y del alba. La relación entre la longitud del intestino y la del pez mostró un crecimiento alométrico positivo. Los huevecillos de los peces representan el principal contribuyente del carbono a la dieta de O. libertate en ejemplares de 140 mm y 1.50 mm de longitud estándar. En individuos de esta especie de 160 mm y 170 mm de longitud estándar, el mayor aporte del carbono al régimen alimentario es provisto por los crustáceos. Aunque las diatomeas representen el mayor aporte en número de individuos y de especies a la dieta de O. libertate, son las que con menos cantidad de carbono contribuyen al régimen alimentario de la sardina crinuda, en relación a los huevecillos de los peces y a los crustáceos. Conforme incrementa la longitud de O. libertate se incrementa la longitud de sus presas. las diatomeas y los dinoflagelados son mas abundantes en los ejemplares de menor talla. Conforme la sardina crinuda aumenta su longitud son mas abundantes en el contenido estomacal: los copépodos, los anfípodos, los ostrácodos y los eufásidos. LITERATURAAngelescu; V, Ecología del mar Argentino (Engraulidae: Engraulis Anchoita ). Parte II : Alimentación, comportammiento y relaciones troficas del ecosistema. Contrib. Inst. Nac. Invest. Des. Pesq. Mar del Plata. 1982, 1-83 409: Angelescu, V. y A. Anganuzzi, Ecología trófica de la anchoita (Engraulidae: Engraulis anchoita) del mar Argentino Parte III: Requerimiento trófico individual en relación con el crecimiento, ciclo sexual y las migraciones estacionales. Rev. Invest. Des. Pesq. 1986. 194-223 5: Baxter, J.L., Fish. Invest. Rept. Summary of biological information on the northern anchovy Engraulis mordax Girard. Calif. Coop. Ocean. 1967. 110-116 11: Beers, J.R., Limnol. Oceanogr. Studies on the chemical composition of the mayor zooplankton groups in the Sargasso Seas off Bermuda 1966. 520-528 11 (4) Berg, J., Mar. Biol. Discussion of methods of investigating the food of fishes with reference to a preliminary study of the prey of Gobiusculus flavescens (Gobilidae) 1979. 263-273 50 (3): Berry, F.H. y I. Barret, Inter. Am. Trop. Tuna Comm.Bull. Análisis de las branquispinas y denominación de las especies del arenque de hebra género Opisthonema. 1963. 1-153 7 (2): Blaxter, J.H.S., Light as an ecological factor Brit. Ecol. Soc. Symp. The effect of light intensity on the feeding ecology of herring. In: . Bainbridge, R. el al (Eds.). 1966. 393-409 6: Cadet, H.H. y L. Berner Jr., Fish. Bull. Fish. and Wildlife Ser. Food on the pacific sardine (Sardinops caerulea) 1959. 175-184 60: Carranza, J., 3er. Informe del Contrato de estudios EI-69- 51. Informe preliminar sobre la alimentación y hábitos alimenticios de las principales especies de peces de las zonas de los planos piloto de Yávaros y Esquinapa. U.N.A.M. Inst. Biol. y SRH. 1969. 28 p. Cupp, E.E., Bull. Scripps Marine plankton diatoms of the west coast of North America. Inst. Oceanog. 1943. 1-238 5 (1): Cupp, E.E., Bull. Scripps Marine plankton of the westem coast of North America Inst. Oceanog. 1950. 1-237 5: Chiappa-Carrara, X., Tesis Consideraciones bioecológicas sobre la alimentación, crecimiento, hábitos alimenticios y contenido calórico de la dieta de las poblaciones de la anchoveta Engraulis mordax Girard localizadas en las costas occidentales de Baja California. UACPyP-CCH-UNAM, 1988. 131 p. Feigenbaum, D. and R.C. Maris, Mar. Biol. Ann. Rev. Feeding in the chaetognatha Oceanogr. 1984. 343-392 22: Gannon, J.E., Trans. Am. Fish. Soc., The effects of differential digestion tales of zooplancton by alewife, Alosa pseudoharengus, on determinations of selective feeding. 1976. 89-95 105 (1): Gallardo-Cabello, M., A. Laguarda-Figueras y R. Corrales-Urrea. Cienc. Pesq. Análisis de la edad, crecimiento y mortalidad natural de la sardina crinuda Opisthonema libertate (Günther, 1866) de las aguas del sur del Golfo de California. (Pisces-Culípeidae). En prensa. García G.M. y D. Molina, Cienc. Pesq. Edad y crecimiento de la sardina crinuda de la zona de Guaymas. 1986. 17-31 5: Gual-Frau, A. y M. Gallardo-Cabello. An. Inst. Cienc. del Mar y Limnol. Análisis del régimen y hábitos alimenticios de la cigala Nephrops morvegicus (Linneo, 1758) en el Mediterráneo Occidental (Crustacea:Nephropsidae) Univ.Auton. México. 1988. 151-166 15 (1): Holt, S.J. Oceanografía. Selecciones del Scientific American. Los recursos alimenticios del océano. Edit. Blume, Madrid: 1975 400-414. Hureau, J.C., Bull. Inst.Oceanog. Monaco Biologie comparée des quelques poissons antartiques (Nothotheniidae). Inst.Oceanog. Monaco, 1969. 1-44 68: Hynes, H.B.N., J. Anim. Ecol. The food of freshwater stieklebacks (Gasterosteus aculeatus and Pygosteus pungitius) with a review of methods used in studies of the food of fishes. 1950. 35-38 19: Hyslop, EJ., J. Fish. Biol Stomach contents analysis. A review of methods and their applications. 1980. 411-429 17. James, A.G., Engraulis capensis Gilchrist. S. Afr. J. Mar. Sci. Feeding ecology, diet and field-based studies on feeding selectivity of the Cape Anchovy 1987. 673-692 5: Leong, R.C. y C.P. O'Connell, J. Fish. Res. Bd A laboratory study of particule and filter feeding to the northem anchovy (Engraulis mordax). Canada, 1969. 557-582 26: Lewis, R.C. Bull. of the Scripps Inst. Oceanogr. Food habits of the California Sardine 1929. 156-180 2 (3): Loukashkin, AS., On the diet and feeding behaviour of the northern anchovy Engraulis mordax Girard. Proc. of the Calif. Acad. of Sciences, XXXVI 1970. 419-458 (13): O'Connell, C.P., J. Fish. Res. Bd. The interrelation of biting and filtering in the feeding activity of the northem anchovy (Engraulis mordax). Canada, 1972. 285-293 29 (3): Omori, M., Mar. Biol Weight and chemical composition of some important oceanic zooplankton in the Northem Pacific Ocean. 1969. 4-10 3: Parsons, T.R., Y. Mieta y C.M. Lalli, A manual of chemical and biological methods for seawater analysis. New York, Pergamon Press, 1984. 173 p. Rice, A.L., J. Cons. Perm. Int. Explor. Mer, The food of the Irish Sea Herring in 1961 and 1962. 1963. 188-200 26: Rojas de Mendiola, B., Fertility of the sea. Science Publish. Some observations on the feeding of the peruvian anchovy Engraulis ringens J. in two regions of the peruvian coast. In: Gordon y Breach (Eds.) 1971. 417-440. 2: Rose, M., Centre National de la Rechérche Scientifique, Manuel de planctonologie Mediterranée. París. 1975. Tomos I y II Shelborne, J.E., J. Mar. Biol. Ass. UX. A predator-prey relationship for plaice larvae feeding on Oikopleura. 1962. 243-252 42: Steedman, H.F., Monographs on Oceanographic Methodology, Zooplancton fixation and preservation. UNESCO. 1976. 4, 350 p. Sullivan, B.K., C.B., Miller, W.T. Petersen and A.H. Soeldner, Mar. Biol. A scanning electron microscope study of the mandibular morphology of boreal copepods. 1975 175-182 30: Thomas, H.J. y C. Davidson, Mar. Res. The food of the Norway lobster. 1962. 515 3: Tregouboff, G. y M. Rose, Manuel de planctonologie Mediterranée. Centre National de la Rechérche Scientifique, París, 1957. Tomos I y II. Tukey, J., Exploratory Data Analysis. Reading, MA: Addison Wesley Publ. Co., New York, 1977. 412 p. Verity, P.G. y C. Langdon, Bay. J. Plankt. Res., Relationships between lorica volume, carbon, nitrogen and ATP content of tintinnids in Narragansett 1984. 857-869 6 (5): Windell, J.T. y S.H. Bowen, An. Inst. Cienc. del Mar y Limnol. Methods for study the fish diets based on analysis of stomach contents. In: Methods for the Assessmente of Fish Production in Fresh Waters, 3a ed. Oxford Blackwell Scientific Publ:219226 T. Bagned (ed.) Univ. Nal. Autón. México, (1991) 1978. 49-58 18 (1):
|