|
EFECTO DE LA MAREA EN EL PASO DE LARVAS DE SCIAENIDOS (PISCES) EN BOCA DEL CARMEN, LAGUNA DE TÉRMINOS, CAMPECHE Recibido el 15 de noviembre de 1990 y aceptado para su publicación el 8 de mayo de 1991.
César Flores Coto
Margarita Pérez Argudín
Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. Apartado Postal 70-305, México, D. F. 04510 MÉXICO Contribución No. 706 del Instituto de Ciencias del Mar y Limnología, UNAM.
RESUMENSe estudió el transporte de larvas de sciaenidos a través de la Boca del Carmen, en la Laguna de Términos, Campeche, en relación a la marea y fisiografía de la boca. Se establecieron tres estaciones de muestreo en la parte más angosta de la boca. La colecta de ictioplancton se hizo durante ocho meses, en tres distintas profundidades, cubriendo en cada ocasión un ciclo de marea. Las condiciones hidrográficas en la Boca del Carmen son reguladas por la marca. Siete sciaenidos fueron identificados, de los cuales sólo Bairdiella chrysoura y Cynoscion nebulosus estuvieron sujetos a la marea en su paso a través de la boca y fueron capturados principalmente de día. Stellifer lanceolatus, Micropogonias furnieri/undulatus y Cynoscion arenarius/nothus estuvieron sujetos a la marea sólo parcialmente, en virtud de que la mayoría de los especímenes alcanzaron la boca a tallas relativamente grandes, entrando a la laguna por áreas de baja salinidad, principalmente en la noche. Menticirrhus saxatilis y M. americanus fueron colectados sólo ocasionalmente. ABSTRACTThe sciaenid larvae migration through the El Carmen Inlet at Terminos lagoon, Campeche; was studied with regard to the tidal and physiographical conditions of the inlet. Three sampling stations were established at the narrowest portion of the inlet. Ichthyoplankton samples were collected during eight months at three different depths complying in each case with a complete tidal cycle. Tides regulate the hydrographic conditions of the inlet. Seven sciaenid species were identified, and from them, only Bairdiella chrysoura and Cynoscion nebulosus were subject to tide forces on their passage through the inlet and were captured mainly during the day, while Stellifer lanceolatus, Micropogonias furnieri/undulatus and Cynoscion arenarius/nothus were only partially subject to the tidal effect, considering that most specimens reached the inlet with a considerable larger size and penetrated the lagoon through low salinity areas, especially by night. Menticirrhus saxatilis and M. americanus were only ocasionally collected.
INTRODUCCIÓNLa mayoría de las corrientes en los estuarios y lagunas costeras, principalmente en las bocas y canales son producidas por las mareas, que son la fuente de energía más importante para mezclar las aguas. El volumen de agua que fluye a través de una boca está directamente relacionado a el área de la laguna y al ciclo y amplitud de las mareas (Phleger 1969). Las corrientes también pueden influenciar el número y tipo de organismos que usan las lagunas y estuarios como áreas de crianza. La inmigración de larvas de camarones y larvas y juveniles de peces a estuarios y lagunas costeras, es influenciado por la marea también como por el viento, el flujo del río, la topografía y las características biológicas de los organismos. Previo al presente trabajo, se estudió también el paso de larvas de Clupeidae en la Boca del Carmen (Flores-Coto et al. 1987) y existe como el único antecedente de estudios tendientes a conocer la inmigración de larvas y juveniles de peces en las lagunas costeras del país. El principal objetivo de este estudio fue determinar a través de un ciclo anual, como influye la marea en la inmigración de larvas de sciaenidos en la Boca del Carmen. MATERIAL Y MÉTODOSLa ubicación de la Boca del Carmen, así como una descripción de su batimetría y la metodología empleada en este estudio se mencionan ya en el trabajo de Flores-Coto et al. (1987), sin embargo se referirán aquí los aspectos más importantes, indispensables para el entendimiento de este escrito. Se establecieron tres estaciones de muestreo en la Boca del Carmen: estación A, sobre el canal más profundo, cerca de la Isla del Carmen, estación B, en la porción media de la boca y estación C, sobre la porción oeste, cerca de punta Zacatal (Figuras 1 y 2). El muestreo se llevó al cabo mensualmente desde abril de 1979 a enero de 1980, excepto en mayo y noviembre, durante los días de máxima amplitud de marea, determinada de acuerdo a las tablas de predicción de marea de Ciudad del Carmen (Anónimo 1978). El ciclo de marca fue dividido en siete fases iguales (Figura 3). las fases 2-3 y 5-6 corresponden a los periodos de máxima velocidad de corriente. Fase uno es el inicio del flujo, fase cuatro es la máxima altura de marea y fase siete es el final del ciclo. Las fases uno, cuatro y siete corresponden a bajas velocidades de corriente e incluyen la estoa. 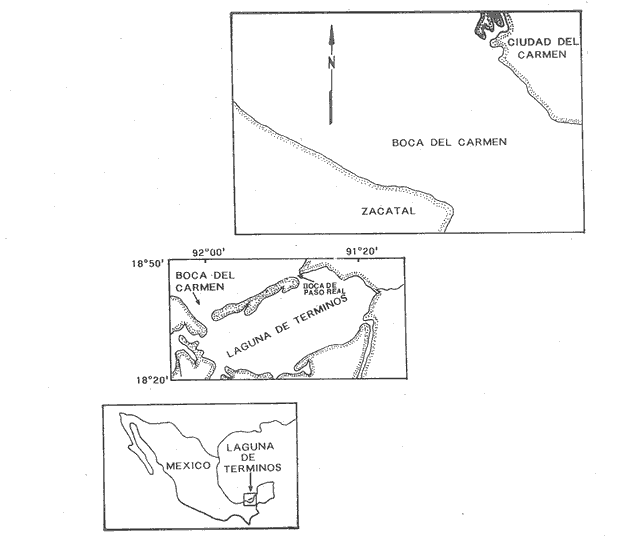 Figura 1. Localización geográfia de la boca del Carmen, laguna de Términos, Campeche, México. 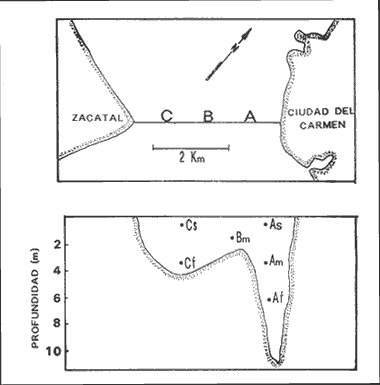 Figura 2. Estaciones (A. B, C) y profundidades (superficie, S; media agua, M; fondo, F) de muestreo en Boca M; Carmen. Para las colectas se utilizaron redes de plancton tipo trapecio de 50 cm de diámetro en la boca y 1.5 m de largo y luz de malla de 500 micras, con un contador en la boca de cada red. En cada estación se hicieron arrastres horizontales cada tres horas, con duración de tres minutos, simultáneos en las diferentes profundidades. Se muestreó primero la estación A seguida de la B y la C. Se midió la salinidad y temperatura por medio de un refractómetro y termómetro de cubeta, respectivamente. 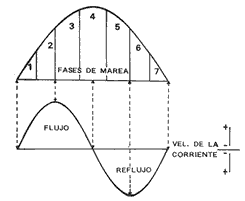 Figura 3. División de la curva de marca y velocidad relativa de las corrientes Fases 1,4, 7=Baja velocidad; Fases 2,3,5,6= Alta velocidad. Los organismos se preservaron en formalina al 4%. Se midió la longitud patrón (LP) de cada larva. RESULTADOS
HIDROLOGÍALos resultados de la HIDROLOGÍA han sido ya publicados por Flores Coto et al. (1987) y se presentan aquí gráficamente (Figura 4), como parte de lo necesario para entender este trabajo. ICTIOPLACTONUn total de 2,157 larvas de sciaenidos fueron capturadas. La densidad anual de los taxa, expresada como larvas/100 m³ fue: Stellifer lanceolatus 3.2, Bairdiella chrysoura 1.8, Micropogonias furnieri/undulatus 1.3, Cynosc¡on nebulosus 0.2, Cynoscion aranarius/nothus 0.2, Menticirrhus saxatilis 0.04 y Menticirrhus americanus 0.007. No pudieron distinguirse las larvas de M. furnieri de las de M. undulatus, ni tampoco las de C. arenarius de las de C. nothus, por lo que las tratamos en conjunto. Stellifer lanceratusDurante este estudio las larvas de esta especie fueron encontradas principalmente en abril y octubre y estuvieron ausentes el resto del año, excepto por un espécimen capturado en agosto y dos en diciembre. Los especímenes fueron muy abundantes en la porción oeste de la boca (estación C), principalmente en aguas del fondo durante el reflujo o estoa, con larvas grandes (5 a 20 mm LP) y pequeños juveniles (>20 mm LP) presentes (Tabla 1). La mayor frecuencia de captura y densidades más altas ocurrieron durante la noche (Figura 5). 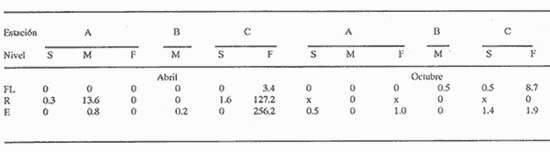 TABLA 1 DENSIDAD MEDIA DE LARVAS DE Stellifer lanceolatus POR 100 m 3 DURANTE LOS PERIODOS DE FLUJO (FL), REFLUJO (R) Y ESTOA (E) EN AGUAS SUPERFICIALES (S), MEDIAS (M) Y DE FONDO (F). X= NO HUBO LARVAS. JUNIO, JULIO, SEPTIEMBRE Y ENERO, NO SE CAPTURARON LARVAS. AGOSTO Y DICIEMBRE SÓLO UNA LARVA EN CADA MES. BOCA DEL CARMEN, LAGUNA DE TÉRMINOS, CAMPECHE En la porción este (estación A) la especie estuvo virtualmente ausente en octubre y pocos especímenes fueron capturados en abril, excepto durante el reflujo. Larvas < 5 mm LP fueron escasas (Figura 6). Bairdiella chrysouraEsta especie estuvo presente en todos los meses muestreados, excepto diciembre. La mayor abundancia fue registrada en abril y junio (Tabla 2). La densidad de las larvas decreció durante el periodo de lluvias (julio-octubre) y se incrementó en enero, pero siempre con densidades promedio menores de 1.5 larvas/100 m³ La mayor frecuencia de ocurrencia y densidad de larvas generalmente se presentó en la porción este (estación A) principalmente en media agua y fondo, durante el flujo (Figura 7). En junio y agosto se observaron altas densidades en muestras tomadas en reflujo. En las estaciones central y oeste la especie fue menos abundante. Mayores densidades y frecuencia de ocurrencia se registraron en capturas diurnas. 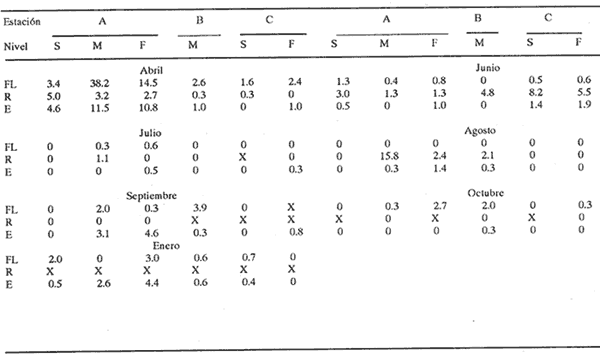 TABLA 2 DENSIDAD MEDIA DE LARVAS DE Bairdiella chrysoura POR 100 m 3 DURANTE LOS PERIODOS DE FLUJO (FL), REFLUJO (R) Y ESTOA (E) EN AGUAS SUPERFICIALES (S), MEDIAS (M) Y DE FONDO (F). X= NO HUBO LARVAS. JUNIO, JULIO, SEPTIEMBRE Y ENERO, NO SE CAPTURARON LARVAS. AGOSTO Y DICIEMBRE SÓLO UNA LARVA CADA MES. BOCA DEL CARMEN, LAGUNA DE TÉRMINOS, CAMPECHE 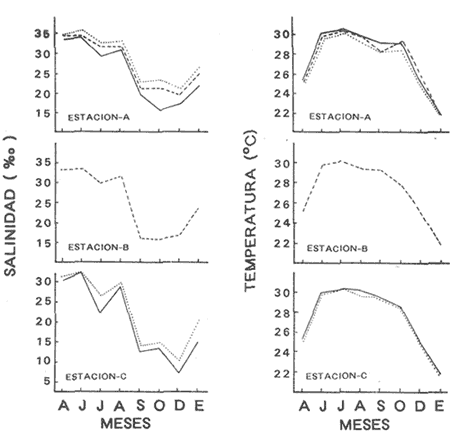 Figura 4. Valores medios de salinidad y temperatura en cada estación (A, B, C,) y profundidad (,-- superficie -------- media agua y ...............fondo) de muestreo. Boca M Carmen, laguna de Términos, Campeche. 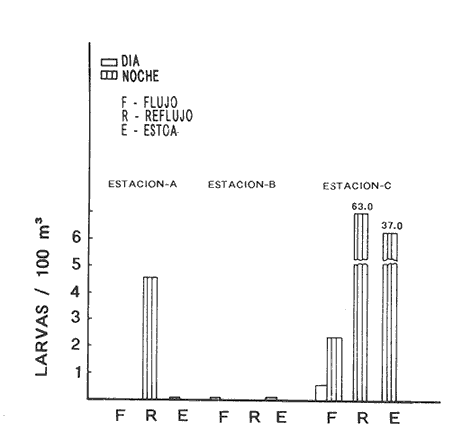 Figura 5. Densidad media anual de Stellifer lanceolatus durante el flujo (F), reflujo (R) y estoa (E). La mayoría de las larvas fueron pequeñas (Figura 6). En abril y junio sus tallas fluctuaron entre 1.5 y 6.0 mm LP durante el flujo, y sólo de 1.0 a 2.5 mm LP durante el reflujo. Micropogonias Furnieri/undulatusLarvas de estos taxa fueron capturadas a través del año, también como algunos juveniles en abril. La densidad mensual promedio fue de 6.5 especímenes/100 m³ en abril y 1.0 espécimen/100 m³ en los otros meses (Tabla 3). Las mayores densidades, particularmente evidentes en abril ocurrieron en el fondo en la estación oeste (C). Esta especie fue registrada principalmente durante el flujo y estoa (Tabla 3). La talla promedio de las larvas fue mayor en la estación oeste que aquellas capturadas en las estaciones media y este. La mayoría de los especímenes fueron >5 mm LP con alguna proporción de juveniles (>20 mm LP) más bien que las larvas (Figura 6). La mayoría de los especímenes fueron capturados durante la noche (Figura 8). Cynoscion nebulosusLas larvas de esta especie ocurrieron en bajas densidades a través del ciclo de muestreo, excepto en invierno. La mayor frecuencia de ocurrencia y abundancia se registró en las zonas de canales (estaciones A y C) en media agua y fondo durante el flujo y en superficie en el reflujo (Tabla 4). En la estación C también hubo altas densidades en el fondo durante el reflujo (Figura 9a). Las larvas fueron menores de 3 mm LP excepto tres y la mayoría fue capturada durante muestreos diurnos (Figura 6). 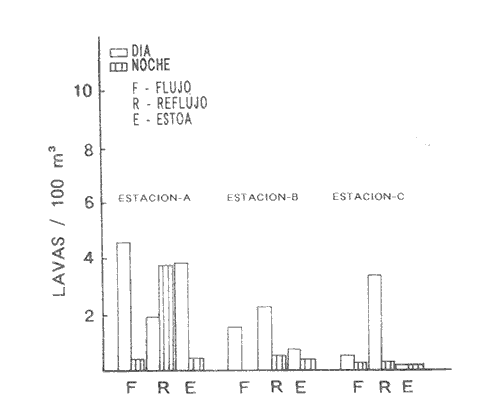 Figura 6. Porcentaje de ha clases de talla de larvas de de capturados en la Boca del Carmen. 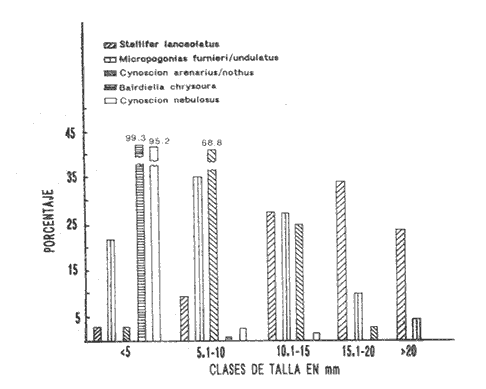 Figura 7. Densidad media anual de Bairdiella chrysoura, durante el flujo (F) reflujo (R) y estoa (E). 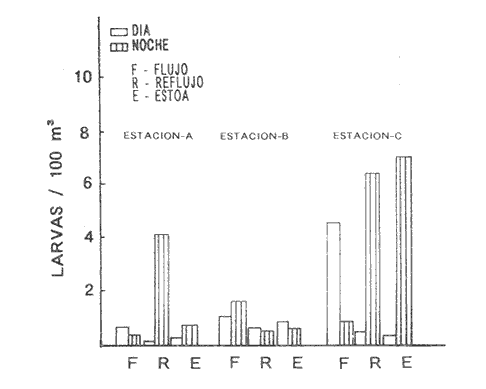 Figura 8. Densidad media anual de Micropogonias furnieri/undulatus durante el flujo (F), reflujo (R) y estoa (E). Cynoscion arenarius/nothusLa mayoría de las larvas de estos taxa fueron capturadas en abril, unas pocas el resto del año, pero ninguna en diciembre y enero. No hubo una gran diferencia en la abundancia de larvas entre estaciones, pero la densidad total promedio es ligeramente mayor en la estación A. La mayor abundancia y frecuencia de ocurrencia se registraron en muestras de fondo y media agua durante periodos de flujo y reflujo (Tabla 4, Figura 9b ). Las larvas fueron mas frecuentes en capturas nocturnas y con una excepción fueron mas frecuentes en capturas nocturnas y con una excepción fueron mayores de 5 mm LP (Figura 6). Menticirrhus saxatilis y M. AmericanusEstas especies estuvieron pobremente representadas. Sólo unos cuantos especímenes de la primera fueron capturados en julio y agosto y sólo tres de la segunda se obtuvieron en abril. 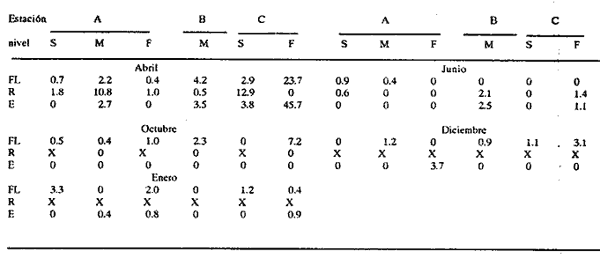 TABLA 3 DENSIDAD MEDIA DE LARVAS DE Micropogonias furnieri/undulatus POR 100 m3 DURANTE LOS PERIODOS DE FLUJO (FL) REFLUJO (R), Y ESTOA (E) EN AGUAS SUPERFICIALES (S), MEDIAS (M) Y DE FONDO (F). X= NO LARVAS. JUNIO JUlIO:SEPTIEMBRE Y ENERO, NO SE CAPTURARON LARVAS. AGOSTO Y DICIEMBRE SÓLO UNA LARVA EN CADA MES. BOCA DEL CARMEN, LAGUNA DE TÉRMINOS, CAMPECHE 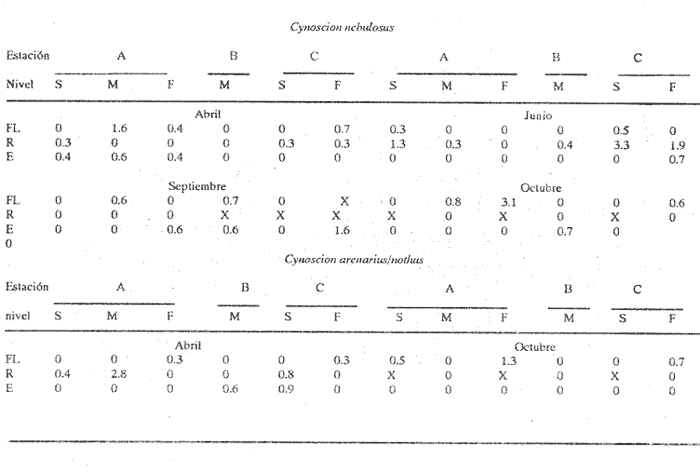 TABLA 4 DENSIDAD MEDIA DE LARVAS DE Cynoscion nebulosus Y Cynoscion arenarius/nothus POR 100 M 3 DURANTE LOS PERIODOS DE FLUJO (FL), REFLUJO (R) Y ESTOA (E) EN AGUAS SUPERFICIALES (S), MEDIAS (M) Y DE FONDO (F). X= NO HUBO LARVAS. NO SE CAPTURARON LARVAS DE C. nebulosus EN AGOSTO, DICIEMBRE Y ENERO Y SÓLO UN ESPÉCIMEN EN JULIO. DURANTE SEPTIEMBRE, DICIEMBRE Y ENERO NO HUBO LARVAS DE C. arenarius/nothus Y SÓLO UNA EN JUNIO Y JULIO Y DOS EN AGOSTO. BOCA DEL CARMEN. LAGUNA DE TÉRMINOS, CAMPECHE 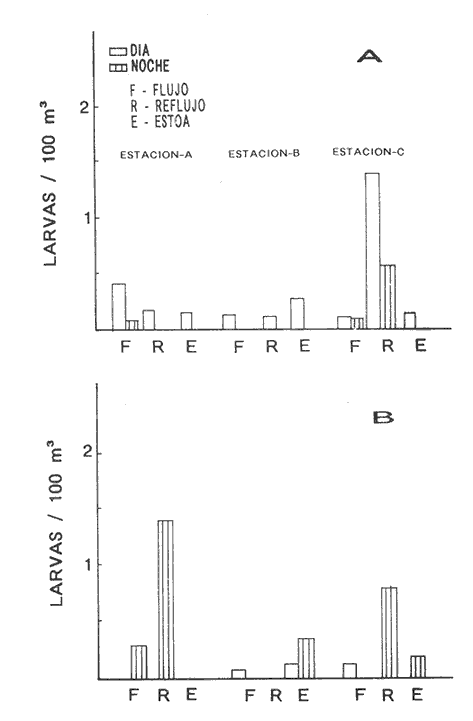 Figura 9. Densidad media anual de Cynoscion nebulosus (A) y Cynoscion arenarius (B) durante el flujo (F), reflujo (R) y estoa (E). DISCUSIÓNEste patrón de circulación fue observado principalmente en la porción este de la boca (estación A) durante la mayor parte del año y fue menos evidente en primavera, cuando la diferencia en salinidad entre las aguas de la laguna y las de la zona costera es mínima. La ocurrencia de mayores salinidades y menores temperaturas en la porción este (estación A) que en la oeste (estación C) es causada por el patrón general de circulación en la laguna de Términos, de este a oeste. La estación de enmedio (estación B) tuvo condiciones hídricas similares a la estación C durante el periodo de lluvias y a la A durante el periodo de secas. HIDROLOGÍALa variación de la salinidad y en menor grado la temperatura, mostraron que la hidrodinámica de la Boca del Carmen es esencialmente controlada por la marea (Flores-Coto et al. 1987); pero la descarga de varios ríos particularmente aquellos localizados en porción oeste de la laguna también es importante. La boca del Carmen se comporta como un estuario estratificado, en el cual durante el flujo, en agua marina entra sobre o cerca del fondo. Las aguas superficiales son menos salinas. Durante el reflujo la mayor parte del agua sale por la superficie. ICTIOPLACTON
Abundancia y periodo de desoveLos resultados muestran que la abundancia relativa de las larvas registradas en este trabajo, corresponde con la referida para la laguna de Términos y área marina adyacente. Larvas y adultos de S. lanceolatus, M. furnieri/undulatus y C. arenarius/nothus son los sciaenidos mas abundantes en el área marina (Sánchez-Gil et. al 1986; Rivera-Elizalde 1988). Larvas de C. arenarius han sido registradas como las más abundantes entre los sciaenidos en las aguas de la plataforma de Florida (Cowan and Shaw 1988). Por otro lado larvas de B. chrysoura y C. nebulosus son las más abundantes entre los scianidos en la laguna de Términos (Pérez-Argudín 1985, Sánchez-Iturbe y Flores-Coto 1986 ). La única especie capturada a través del año fue C. nebulosus; sin embargo esta especie junto con los demás taxa, excepto Menticirrhus pueden considerarse como desovantes de todo el año, en el sur del Golfo de México, como puede evidenciarse por la ocurrencia de larvas en la laguna de Términos y área marina adyacente (Sánchez-Iturbe y Flores-Coto 1986; Rivera-Elizalde 1988). En la Boca del Carmen S. lanceolatus y C. Arenarius/nothus tuvieron dos picos estacionales de abundancia de larvas, el más importante a principios de primavera y el otro en verano o principios de otoño; M. furnieri/undulatus sólo tuvo uno a principio de primavera. Los picos de abundancia estacional de estas especies ocurrieron entre uno y dos meses mas tarde que los periodos de alta abundancia de larvas registradas en el área marina adyacente por Rivera-Elizalde (1988) (Tabla 5). Micropogonias furnieri/undulatus puede ser considerado el único taxon entre los sciaenidos cuyo principal periodo de desove es el invierno, y que es el mismo período al cual restringe su desove en latitudes mas altas (Ditty et al. 1988). Influencia de la mareaEste estudio muestra que todas las especies registradas estuvieron en algún grado sujetas a la marea en su paso a través de la Boca del Carmen. Generalmente las mayores densidades de larvas fueron capturadas durante los momentos de flujo y reflujo. Considerando la hidrodinámica de la boca puede esperarse que las larvas de sciaenidos sujetas a la marea inmigren a la laguna principalmente en aguas medias o de fondo durante el flujo de marea y pueden salir durante el reflujo, en aguas superficiales. Sin embargo sólo B. chrysoura y C. nebulosus mostraron tal patrón teórico de distribución, quizá debido a que la mayoría de sus larvas fueron menores a 3 mm Lp y por tanto con capacidades natatorias muy reducidas. La pequeña talla de estas larvas corresponde al hecho de ser las únicas especies de sciaenidos que desovan en la laguna o área litoral adyacente (Ferreira-González y Acal-Sánchez 1984; Pérez-Argudín 1985; Sánchez-Iturbe y Flores-Coto 1986). Stellifer lanceolatus, M. furnieri/indulatus y C. arenarius/nothus estuvieron sujetas a la marea sólo parcialmente. Sus mayores densidades de larvas en aguas medias y de fondo y su talla fue generalmente mayor de 10 mm Lp, con sólo unos pocos especímenes menores de 5 mm Lp. Las mayores densidades en aguas medias y de fondo pueden ser consecuencia de que las larvas avanzadas y pequeños juveniles tengan mayores capacidades natatorias para moverse independientemente de las corrientes y estén fuertemente orientadas hacia el fondo. Hoese (1965) registra que las postlarvas de M. undulatus son preferentemente organismos del fondo y que gran número de ellos entra a las bahías durante los flujos de marea. La misma tendencia es registrada para S. lanceolatus y otras especies de sciaenidos (Weintein et al. 1980; Powles 1981; Miller et al. 1984). No hay información disponible sobre C. arenarius, pero los resultados sugieren que esta especie se comporta de la misma manera. La relativa alta abundancia de larvas avanzadas y pequeños juveniles y la relativa escasez de larvas menores de 5 mm LP de S. Lanceolatus, M. furnieri/undulatus y C. arenarius/nothus en la Boca del Carmen es resultado, de que estos taxa habiendo desovado en las zonas marinas emigran hacia los estuarios, los que alcanzan a tallas generalmente mayores de 10 mm Lp. La gran capacidad natatoria de larvas y juveniles al alcanzar el estuario les permite moverse independientemente de la marea y explica las altas densidades registradas durante los periodos de estoa. El desove de muchos sciaenidos en áreas oceánicas y la emigración de sus larvas hacia los sistemas estuarinos ha sido ampliamente registrada por diversos autores (Hoese 1965; Moffet 1970; Weintein 1980; Warlen 1982; Miller et al. 1984; Ruple 1984) para las costas atlánticas de los Estados Unidos. Powles (1980) considera que S. lanceolatus desova más en los estuarios que en el mar. Sin embargo la presencia abundante de larvas de esta especie, así como también de M. furnieri/undulatus y C. arenarius/nothus en el sur del Golfo de México (Rivera-Elizalde 1988) sugiere que el desove ocurre ahí y que sus larvas emigran hacia los sistemas estuarinos como la laguna de Términos.  TABLA 5 PICOS DE MAYOR ABUNDANCIA DE LARVAS EN LA BOCA DEL CARMEN Y ÁREA MARINA ADYACENTE Stellifer lanceolatus y M. furnieri/undularus fueron más abundantes en la porción oeste de la boca, la cual tuvo las salinidades mas bajas a través del año, lo que sugiere que el paso a través de la boca este orientado por un fuerte gradiente salino. Stellifer lanceolatus y M. furnieri/undularus y C. arenarius/nothus fueron capturados principalmente durante la noche. Cowan y Shaw (1988) también registraron las mayores capturas de C. arenarius y M. furnieri/undulatus durante la noche. Weinstein et al. (1980) sugieren que las larvas de M. undulatus tienden a moverse hacia arriba en la columna de agua durante la noche. La mayor actividad nocturna de los planctontes parece un hecho bastante generalizado, sin embargo en la Boca del Carmen B. chrysoura y C. nebulosus fueron especies capturadas principalmente en muestreos diurnos. AgradecimientosLos autores agradecen a S. Warlen, S. Holt y L. Soto por la revisión del manuscrito; así como al CONACYT y DGAPA-UNAM por el apoyo económico para la estancia sabática-posdoctoral, durante la cual se terminó el presente trabajo. LITERATURAANÓNIMO. Tablas de predicción de marea. Instituto de Geofísica. Univ. Nal. Autón. México 1979. 48 p. COWAN, J. Y R. F. SHAW. Fish. Bull. The distribution, abundance and transport of larval Sciaenids collected during winter and early spring from the continental shelf water off Louisiana. 1988. 129-142. 86 (1): DITTY, J. G., G.G. ZIESKE Y R. SHAW. Fish. Bull. Seasonality and depth distribution of larval fish in the northern Gulf of Mexico above latitudes 26° 00' N. 1988. 811-822. 68 (4): FERREIRA-GONZÁLEZ, R, Y D. ACAL-SÁNCHEZ. Tesis profesional. Biología. Estudio de la comunidad ictioplanetónica de la laguna de Términos, Campeche. Escuela Nacional de Estudios Profesionales. Iztacala. UNAM 1984. 93 p. FLORES-COTO, C. V. DUCOING-CHAHO, F. ZAVALA-GARCÍA, A. VELARDE-MÉNDEZ Y S. MÉNDEZ-VELARDE. An. Inst. Cienc. del Mar y Linmol. Efecto de la marca en el paso de larvas de algunas especies de la familia Clupeidae (Pisces), en la Boca del Carmen, laguna de Términos, Campeche. México Univ. Nal. Autón. México. 1987. 14 53-68. (1): GIERlOFF-EMDEM, H. G. In: Orbital Remote Sensing of Coast and offshore environments. A manual of interpretalion. Laguna de Terminos and Campeche Bay, Gulf of Mexico, water mass interaction lagoonal oceanic visible due to sediment landed maters. Water Gawfyter, Ed. Berlin 1977. 78-79. GRAHAM, D.S., J.F. DANIEL, J.M. HILL Y J. W. DAY JR. An. Inst. Cienc. del Mar y Limnol. A preliminary model of circulation of Terminos lagoon, Camp. Mexico. Univ. Nal. Auton. Mexico 1981. 51-61. 8 (2): HOESE, H. D. Spawning of marine fishes in Port Aransas, Texas, area as determined by distribution of young and larvae. Ph. D. Dissertation. Univ. of Texas, Austin 1965. 150 p. MANCILLA-PERAZA, M. Y M. VARGAS-FlORES. An. Centro Cienc. del Mar y Limnol. Los primeros estudios sobre la circulación y flujo neto de agua en la laguna de Términos, Camp. Univ. Nal. Autón México. 1980. 1-14. 7 (2): MILLER, J.M., J.P. REED, Y CJ. PIETRAFESA. Mechanisms of migrations in fishes Patterns, mechanisms and approaches lo the study of migrations of estuarine-dependent fish larvae and juveniles. J.D. McCleave, G.P. Arnold, JJ. Dodson, and W.11. Neil (editors), Plenum Publ. Corp. N.Y. 1984 p. 209-225. MOFFET, A. W., L.W. MCEACHRON Y J. G. KEY. Texas Contributions in Marine Science. Observations on the biology of sand seatrout (Cynascion arenarius) in Galveston and Trinity Bays. 1979. 163-172. 22: PÉREZ-ARGUDÍN, M. Tesis profesional Contribución al conocimiento de los primeros estudios del desarrollo de las especies de Sciacnidos encontrados en la laguna de Términos, Campeche. E.N.E.P. Iztacala U.N.A.M. 1985. 140 p. PHLEGER, F. B. Lagunas costeras. Some general features of costal lagoons. In: Lagunas costeras un Simposio. Mem. Simp. Inter. Ayala-Castañares, A y F. B. Phleger (eds.) UNAM-UNESCO, Nov. 28-30,1967: 1969. 5-25. PHLEGER, F. B. Y A. AYALA-CASTAÑARES. The American Association of Petroleum Geologist Bulletin. Process and history of Terminos lagoon, Mexico. 1971. 2130-2140. 55 (12): POWLES, H. Fisheries Symposium. Eggs and larvae of North American sciaenid fishes Proceedings of the sixth Annual Marine Recreational Sport Fish. Int. Washington. D. C. Houston, Texas, March 19-20, 1981. 1981. 99-109. RIVERA-ELIZALDE, J. Tesis profesional. Contribución al conocimiento de los primeros estadios de vida de las especies de la familia Sciaenidae (pisces) en el sur del Golfo de México. Fac. de Ciencias. U.N.A.M. 1988. 42 p. RUPLE, D. L. Estuarine, Costal and Shelf Science Occurrence of larval fishes in the surf zone of the Northern Gulf of Mexico Barrier Island. 1984. 191-208. 18: SÁNCHEZ-GIL, P.A. YAÑEZ-ARANCIBIA Y F. AMEZCUA-LINARES. An. Inst. Cienc. del Mar y Limnol. Diversidad, distribución y abundancia de las especies y poblaciones de peces demersales de la Sonda de Campeche (Verano, 1978) Univ. Nal. Autón. de México. 1981. 209-240. 8: SÁNCHEZ-ITURBE, A. Y C. FLORES COTO. An. Inst. Cienc. del Mar y Limnol. Estimación de la biomasa de Bairdiella clirysoura por medio del censo de huevos y algunos aspectos de sus primeros estadios, laguna de Términos, Campeche (Pisces: Sciaenidae.) Univ. Nal. Autón. México. 1986. 133-148. 13 (2): WARLEN, S. M. Proc. Ann. Conf. S. E. Assoc. Fish and Wild Agencies Age and growth of larvae and spawning time of atlantic croaker in North Carolina. 1982. 204. 34: WESTEIN, M. P., S. L. WEIS, R. G. HODSON Y L. GERRY. Fish. Bull. Retention of three taxa of postlarval fishes in an intensively flushed tidal estuary. Cape Fear, North Carolina. 1980. 410-436. 78 (2): WEYL, P.K. 1970 Oceanography. An introduction to Marine Environment. Willey and Sons Inc. New York. 534 p. YÁÑEZ-CORREA, A. Bol. Inst. Geol. Univ. Nal. Autón. México. Batimetría, salinidad, temperatura y distribución de sedimentos recientes en la laguna de Términos, Campeche, México. Univ. Nal. Autón. México. 1963. 147. 67 (1):
|