|
THE KARYOTYPE AND "G" BANDS OF Haemulon
aurolineatum CUVIER, 1829 (PISCES: HAEMULIDAE)
Trabajo recibido el 18 de
mayo de 1990 y aceptado para su publicación el 12 de julio de
1990.
A. Durán - González
C. E. García - Rucias
A. Laguarda-Figueras
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología.
Contribución No. 687 del Instituto de Ciencias del Mar y
Limnología, UNAM. Trabajo presentado en el VII Congreso Nacional de
Oceanografía, efectuado del 27 al 31 de julio de 1997 en Ensenada Baja
California México
Los autores presentan el
análisis citogenético de la especie Haemulon aurolineaum
(Pisces: Haemulidae), cuya variable abundancia muestra amplia
distribución en el Océano Atlántico incluyendo el Golfo de
México y el Mar Caribe. La especie estudiada ha caído en
sinonimia con los génerosBathystoma y
Brachygenys. El cariotipo, obtenido de 63 organismos
colectados en el banco de Yucatán, México, muestra un
número diploide de 2n=48 cromosomas acrocéntricos, por lo que su
número de brazos cromosómicos es FN = 48. No fueron identificados
cromosomas sexuales. Asimismo, con el propósito de reconocer y
clasificar mejor los cromosomas homólogos de esta especie fueron
estudiadas las bandas "G" de sus cromosomas.
The authors
report the cytogenetic analysis ofHaemulon aurolineatum
(Pisces: Haemulidae) species, which variable abundance shows a wide
distribution in the Atlantic Ocean including the Gulf of Mexico and the
Caribbean Sea. Sometimes it falls in synonymities with the
Bathystoma and Brachygenysgenera. The
karyolype, obtained from 63 organisms collected in the bank of Yucatán
México, shows a diploid number of 2n=48 acrocentric chromosomes, with a
major chromosome arm number of FN-48 Sexual Chromosomes were not identified.
Moreover, "G" bands in the karyotype of this species were studied, in order to
get a batter recognition and classification of the homologous
chromosomes.
The particular dynamic of the sea makes difficult to establish clearly the geographic location and isolation degree of the different fishes populations. For this reason, the cytogenetic studies whose purpose is the cytotaxonomy of these organisms, are very important, because they help to locate populations, races or species in space and time, and allow us to stablish, by means of chromosome analysis, the philogenetic distances of the taxonomic entities which form this important group of living beings. That is the case of the populations of Haemulon aurolineatum which are very abundant and widely distributed in the reefs and coastal areas of the Gulf of Mexico and the Caribbean Sea. This species form a dominant part in the structure of the demersal fish communities in its wide geographic distribution (Fischer, 1978, Darcy, 1983, Yañez-Arancibia et al., 1985) which cover and extense area from Cape Cod, Massachussetts (USA) to Brazil, including the Gulf of Mexico and the Bermuda Islands (Courtenay, 1961, Fischer, 1978, Díaz -Ruiz et al., 1982, Darcy, 1983). Some authors have made diverse taxonomic considerations about the "status" of the Haemulidae family (Konchina, 1976, Robins et al., 1980) and their genera and species (Courtenay, 1961, Hong, 1977, Fischer, 1978, Díaz Ruiz et al., 1982, Darcy, 1983, Nelson, 1984, Corona García, 1989). The genus and species have fallen into synonymities with the genera Brachygenys and Bathystoma in a general level and at specific level with Bathystoma aurolineatum, a scientific name which is still used in naming the species studied in the present study (Fischer, 1978). Knowing that the Haemulidae shows, large morphology and color variations which have made very difficult to delimit its populations, races or species and to recognize youths, adolescents and adults of the same species, the present work establishes the karyotype and the "G" bands of the chromosomes of Haemulon aurolineatum in order to obtain a better characterization of this taxonomic entity. COLLECTING AREAThe studied specimens were collected and processed on board of the B/O " Justo Sierra " vessel, during the oceanographic campaigns " Prospección Caribe Mexicano PROIBE II, III, IV y V ", which took place in July of 1984, April and October of 1985 and February of 1986 respectively. The organisms were caught in the area located between the 21° 45' - 22° 20' latitude north and the 86° 45' - 87° 10' longitude west (Fig. 1) at variable depths between 30 and 60 meters. 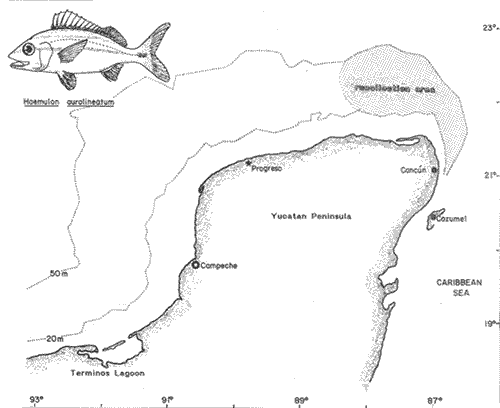 Figure 1. Collecting arca of Haemulon aurolineatum. MATERIAL AND METHODSFifty one live organisms (which were maintained in aquaria) and twenty that died during the dragging and liberation of the catch, were processed. The sex of all the organisms was determined, by means of direct observation of fresh gonads and by microscopic examination. Twenty five females, ten males and twenty eight undifferentiated juveniles were processed. Some treatment based on the general methodology proposed by McPhall and Jones (1966), Lieppman and Hubbs (1969). Denton (1973), Gold and Gall (1975) and Kligerman and Bloom (1977), were adapted in order to obtain mitotic fields from the gill's epithelia and lymphocytes of the studied organisms. The slides with tissue were treated by the air drying technique. A number of 463 mitotic fields, at metaphase were obtained. A range of 4 to 22 metaphases for each one slide was reached. A number of eleven metaphase fields from different organism was selected in order to analyze the Karyotype of the species The statistical analysis of the mitotic fields was made according to Levan et al., (1964) and Al-Aish (1969) criteria. The "G" bands were obtained in accordance with the model and the techniques of Makay (1973), for chromosomes at prometaphase from three organisms. RESULTSThe karyotype presents a diploid number (2n) of 48 chromosomes, arranged in 24 acrocentric pairs (Fig.2). Chromosomic heteromorphism or negative heteropygnosis which make evident the presence of sexual chromosomes, was not found. Table 1 shows the results of the results of the statistical analysis of the karyotypes. According with the classification of Levan et al., (1964) and Al-Aish (1969) all the chromosomes were telocentric (acrocentric) on account of the strict terminal position of the centromere. As it can be appreciated in this table, the values corresponding to the short (p) and long (q) amr's proportion, the centromeric index (I.C.) and the difference between arms, are omited because the chromosomes lack of short arms. According with the European School - that define de Fundamental Number (FN) as the total number of arms which correspond to the haploid complement, without taking in to account the sexual chromosomes - the species in study has a FN of 48 chromosomes, because all of them are monoarmed. For this reason the idiogram (Fig.3) was made according with the decreasing lenght of the haploid complement. Table 1 also shows the number of the "G" bands in each one of the chromosomes. Figures 3 and 4 show the position of the "G" bands along each chromosome, taking the centromere as a reference point according to their relative lenght, in order to facilitate the identification of the achrocentric homologous pairs (Comings, 1978). DISCUSSIONThe descriptions of various authors about several species of the Haemulidae Family are incomplete and it is not coincident. The descriptions for the species determination, using meristic and morphometric characteristics, have limited precision. Sometimes, criteria such as coloring, are used as determinant (Hong, 1977). Therefore it has generated a high number of synonymities which make uncertain the taxonomic location of the species. Other studies have been made trying to clarify phylogenetic relations at genus level in the Haemulidae family among which Cequea and Pérez (1971) studied the proteins variations in hemoglobines and esterases. However, the proteins used (Hb and Es) did not permit differentiate various species of Haemulon genus but showed to be useful for separation at genus level. Authors mention that possibly the separation of species is a recent fact. 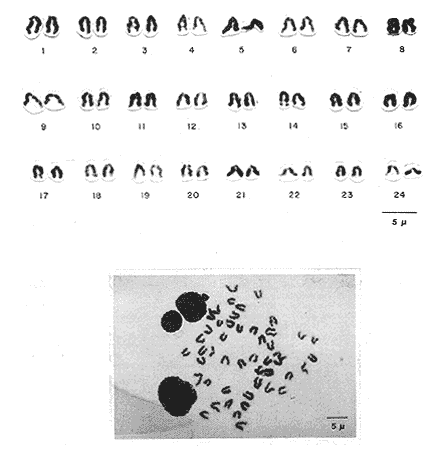 Figure 2. Karyotype of Hammulon aurolineatum Cuvier, 1829. 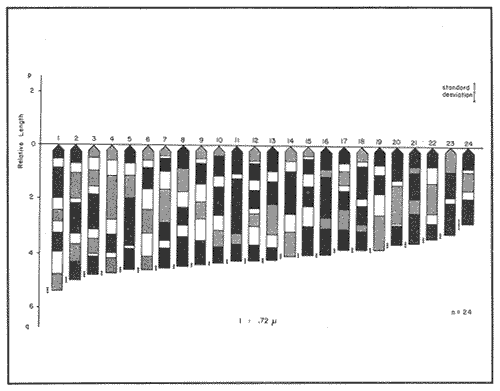 Figure 3. Idiogram with G Bands of Haemulon aurolineatum showing "G" Bands. Mago in 1961, performs comparative osteological studies on some species of the Haemulidae family. He mentions that representatives of same are modern perciformes in full stage of evolution. Evolution referred to by Mago is morphoanatomic as cariologically speaking, the two studied species, H. sciurus, (Regan et al., 1961) and H. aurolineatum (present study), do not show variations in regards to the ancestral karyotype. Therefrom the conventional Idiograms (size and morphology) and the "G" band Idiograms help to establish the correct taxonomic localization of the species and the rank of kindred among them. Moreover, the Cytogenetic studies help us learn the mechanisms of the adaptative dynamics of this kind of fish. Several caryologic works-among which stand out those of Ohno et al. (1968), Denton (1973) and Uyeno et al. (1983) consider that the ancestral karyotype, from which the evolution of the chromosomic complements of the fish are generated, is composed of 48 achrocentric (telocentric) chromosomes. Therefrom, the importance and need that all those works should be complemented by chromosomic bands studies. The 48 chromosomes which form the diploid number of Haemulon aurolineatum are telocentric according to the Levan et al. (1964) classification. Nevertheless, Darlington (1936), Rhoades (1940) and White (1954), consider that the telocentric are acrocentric because the short arm is always present, nevertheless his size must be smaller than the optic microscope resolution limits. Therefore, the cromosomic complement of the studied species coincide with the ancestral karyotype in the number and morphology of its chromosomes. According with the previous considerations, the "G" band pattem of the studied population ofHaemulon aurolineatum is very important because it will be compared with "G" band patterns that are distributed along the coast of the Atlantic Ocean from Cape Cod, Massachussets (USA) to the coast of Brazil, which will be studied in further works. 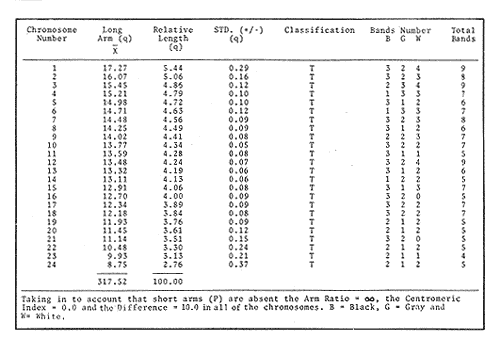 TABLE 1 RESULTS OF STATISCAL ANALYSIS OF THE KARYOTYPE OF Haemulon aurolineatum CUVIER, 1829 CONCLUSIONSThe species determination of fishes, using meristic and morphometric characteristics, have limited precision and therefore, it has generated a high number of synonymities which make not precise the taxonomic location of the species. The conventional idiograms and the "G" bands, studies help to stablish the correct taxonomic localization of the species and the adaptative dynamics, of the fishes. Several caryologic: works consider that the ancestral karyotype from which all the evolution of the chromosomic: complements of the fishes are generated, is composed. by 48 achrocentric (telocentric) chromosomes. The 48 chromosomes which form the karyotype of Haemulon aurolineatum (Pisces: Haemulidae) obtained from organisms collected in the bank of Yucatán, México, are telocentric (FN = 48) and sexual chromosomes where not identified. Therefore, the chromosomic complement, of the studied species coincide with the ancestral karyotype in the number and morphology of its chromosomes. 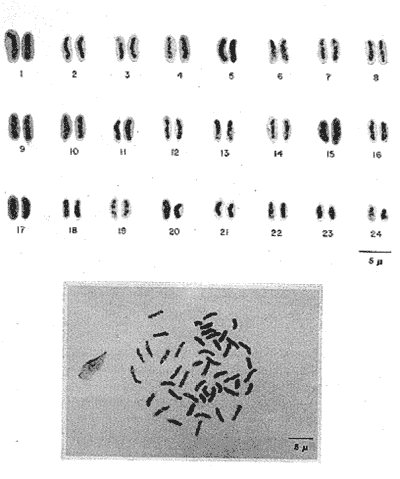 Figure 4. Karyotype of Haemulon aurolineatum showing "G" Bands. AgradecimientosThe authors are grateful to E. Lozano-Alvarez antl to F. Flores-Andolais from the "Estación de Investigaciones Marinas Puerto Morelos" of the "Instituto de Ciencias del Mar y Limnología" of the "Universidad Nacional Autónoma de México" (UNAM) for their help in order to participate in the oceanographic campaigns "PROIBE". Likewise we would like to thank A. Corona-García and P. Fuentes-Mata from the "Instituto de Biología de la UNAM" for the classification of the Biologic Material. LITERATURAAL-AISH, M. Can. Jours. Gen. and Cytol. Human Chromosome Morphology. I. Studies on normal (chromosome) caracterization, Classification and karyotyping. 1969. 370 - 381.11: CEQUEA, R.H. Y J.E. PEREZ.Bol. Inst Oceanogr. Univ. Oriente Variación Intra e Interespecíficas de Hemoglobina y proteínas del plasma en algunas especies del género Haemulon. 1971.79-85.10 (2) COMINGS, D.E. Ann. Rev. Genet. Mechanism of Chromosome Banding and implications for chromosome structure.1978. 25 - 46. 12: CORONA-GARCIA, M.A. Tesis Profesional.Contribución a la Taxonomía y Biología de los géneros Pomadasys y Haemulon (Pices: Haemulidae) de la Costa del Pacífico Mexicano. Fac. de Ciencias UNAM. 1989.121 p. COURTENAY, W.R. Jr.Bull. Mar. Sci. Gulf Caribb. Western Atlantic fishes of the genus Haemulon (Pomadasyidae): Systematic status and juvenile pigmentation. 1961.66 - 149. 11 DARCY, G.H. Synopsis of Biological Data of the Grunts Haemulon aurolineatum and H. plumieri (Pisces: Haemulidae)U.S. Dep. Commer. NOAA Tech. Rep. NMFS Circ. 1983. 1 - 37. 448: DARLINGTON, D.E. Jour. of Genet Crossing Over and its Mechanical Relation in Chortippus and Stauradarus.1936. 440 - 465. 33: DENTON, T.F. Fish Chromosome Methodology.Charles C. Thomas PublisherIllinois U.S.A. 1973. 166 p. DIAZ-RUIZ, S.A. YAÑEZ-ARANCIBIA y F. AMEZCUA LINARES.An. Inst Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Taxonomía, Diversidad, Distribución y Abundancia de los Pomadasidos de la Laguna de Términos, Campeche (Pisces: Pomadasyidae)1982. 251 - 278. 9(1): Species Identification Sheets for Fishery Purposes. Western Central Atlantic (Fishing Area 31) FISCHER, W. (Ed.) FAO Rome1976 1-7. GOLD, J.R. and G.A.E. GALL.Can. J. Genet. Cytol. Chromosome Cytology and Polymorphism in the California High Sierra Golden Trout (Salmo). 1975. 41-53. 17: HONG, S.L. Copeia Review of Eastem Pacific Haemulon with notes on Juvenile Pigmentation.1977. 493 - 501. 3: KLIGERMAN, A.D. and S.F. BLOOM. J. Fish. Res. Board Can. Rapid Chromosome Preparations from solid tissues of fishes. 1977.266-269. 34 KONCHINA, Y.V. J. Ichthyol. The Systematics and Distribution of the Grunt family (Pomadasyidae).1976. 883 - 900. 16: LEVAN, A., K. FREDGA and A.A. SANDERBERG. Hereditas. Nomenclature for Centromeric Position of Chromosomes. 1964.201-220. 52 LIEPPMAN, M. and C. HUBBS. Texas Rep. Biol. Med. A karyologycal analysis of two cyprinid fishes, Notemigonus crysoleucas and Notropis lutrensis.1969. 427 - 435. 27 (2): MAGO, L.F. Bol. Inst. Oceanogr. Univ. Oriente. Osteología Comparada en ocho especies Pomadasyidae (Pisces: Perciformes) del Golfo de Cariaco Venezuela y áreas adyacentes. 1961. 369 - 473. 1: McKAY, R.D.G. The Mechanism of G and C Banding in Mammalian Mataphase Chromosomes. Chromosoma (Berl.) 1973.1-14. 44: McPHAIL., J.D. and R.L. JONES.A Simple Technique for obtaining Chromosomes from Teleost Fishes. J. Fish. Res. Board Can. 1966. 767 -768. 23(5) NELSON, J.S. Fishes of the World. Wiley Interscience 2a. Ed. U.S.A. 1984. 419 p. OHNO, S., U. WOLF and N.B. ATKIN. Hereditas Evolution from Fish to Mammals by Gene Duplication. 1968. 169 - 187. (6)59 REGAN, J.D., M.M. SIGEL, W.H. LEE, K.A. LLAMAS, A.R. BEASLEY. Can. J. Genet. Cytol.Chromosomal Alterations in Marine Fish Cells in Vitro. 1978. 448 - 453. 10: RHOADES, M.M. Genetics Studies of Telocentric Chromosome in maize with reference to the Stability of its Centromeres. 1940.483 -520. 25: ROBINS, C.R., M. BAILEY, C.R BOND, J.R. BROOKER, E.A. LACHNER, R.N. LEA and W.B. SCOTT. A List of Common and Scientific Names of Fishes from the United States and Canada. Am. Fish. Soc. Spec. Publ. 4th. ed. 1980. 174 p. 12. UYENO T., R.R. MILLER and J.M. FITZSIMONS. CopeiaKaryology of the Cyprinodontoid Fishes of the Mexican Family Goodeidae. 1983407-510. (2): WHITIE, M.J. Animal C. and Evolution. Cambridge Univ. Press.LONDON.1954. 188p. YAÑEZ-ARANCIBIA A., SANCHEZ-GIL G.L VILLA LOBOS ZAPATA y R. RODRIGUEZ-CAPETILLO. Recursos Pesqueros Potenciales de México: La Pesca Acompañante del Camarón. Distribución y Abundancia de las Especies Dominantes en las Poblaciones. Peces demersales de la Plataforma Continental Mexicana del Golfo de México. In: Yañez-Arancibia, A. (Ed.) Prog. Univ. de Alimentos, Inst. Cienc. del Mar y Limnol., Inst. Nal. de la Pesca. UNAM. D.F.México1985. Cap. 8: 315-398. 748 p.
|