|
INTERACCIÓN TRÓFICA ENTRE LOS BAGRES Arius melanopus
(AGASSIZ 1129) Y Arius felis (LINNAEUS, 1766), EN LAS COSTAS DE
CELESTÚN, YUCATÁN, MEXICO
Trabajo recibido el 20 de marzo
de 1990 y aceptado para su publicación el 20 de septiembre de
1990.
Ma. Eugenia Vega Cendejas
Centro de Investigación y Estudios
Avanzados del IPN. Apartado Postal 73 CORDEMEX 97310. Mérida,
Yucatán, México.
En
Arius felis y Arius melanopus se
determinaron los patrones tróficos con su variación temporal al
fin devaluar el translape de recursos alimenticios que existen entre estas dos
especies y determinar el tipo de adaptaciones ecológicas que les
permiten coexistir. El análisis trófico se realizó en 463
ejemplarers juveniles y adultos capturados con chinchorro en las costas de
Celestún, Yucatán. Ambas especies usan una amplia variedad de
artículos alimenticios en su crecimiento, pero fundamentalmente
crustáceos y peces constituyen los principales componentes de su dieta.
En la ontogenia de ambos peces se presentan tres etapas tróficas: 1) Una
especialización en el consumo de pequeños crustáceos, 2)
Una generalización trófica y 3) la tendencia a una
especialización en el consumo de peces durante las últimas etapas
de crecimiento. Este patrón presenta variaciones temporales. En la
estación de secas los peces adultos de ambas especies consumen
sólo peces lo que resulta en un mayor traslape entre ellas (1.0) al
incidir prácticamente sobre los mismos recursos alimenticios. Mientras
que en los periodos de lluvias y nortes otros componentes como peneidos y
braquiuros son agregados como componentes en su dieta . La alta diversidad
trófica que se presenta para ambas espécies en el período
de secas, indica que en esta estación climática ambas son
más generalistas en sus hábitos alimenticios. Se concluye
queArius felis y A. melanopus coexisten
por diferencias tróficas ontogenéticas que están en
sincronía con patrones estacionales de productividad. Ambas especies de
bagre influyen directamente en las pesquerías y en el sistema
multiespecífico del que son parte al constituir los decápodos y
peces de importancia comercial su fuente alimenticia
principal.
The trophic ontogenetic patterns with its seasonal
variation were determinated inArius felis y Arius
melanopus in order to evaluate the overlap of food resources that
exist between these two, species and to determinate the kind of ecological
adaptations which allow them to coexist. Trophic analyses were carried out in
463 juvenile and adult fishes that were caught with trawling operations in the
coast of Celestun, Yucatán. Both species took a wide variety of food
items over the growth period, but fundamentally crustaceans and fishes
constituted the main components of their diet. In their ontogeny three trophic
stages exist for both fishes: 1) an especialization on small crustaceans, 2) a
trophic generalization and 3) the tendency to be trophic specialists in fish
ingestion during later growth stages. This pattern presents temporal variation.
In dry season, the adult stages in both species consume only fishes, resulting
in the higher dietary over lap between them (1.0) because of the ingestion of
almost the same resources. Meanwhile in rainy and northperiods other components
like pennaids and brachiurans are added as components in their diet. The high
trophic diversity determinated for both fines in dry period indicates that both
are more generalist in their feeding habits. It is concluded that A.
felis and A. melanopus coexist because of
differences, in their trophic ontogenetic paltem which are in syncrony with
seasonal progressions of productivity. Both species have a direct influence in
fisheries and in the multiespecific system from which they from a part because
decapods and commercial fishes constitute their principal food
resources.
En América tropical y subtropical los bagres se encuentran entre los peces más abundantes en las lagunas costeras y estuarios. dentro de este gran grupo, Arius felis y Arius melanopus revisten gran importancia económica y ecológica al presentar una alta frecuencia de pesca y una amplia distribución en el Golfo de México y el Mar Caribe (Yáñez -Arancibia y Lara, 1988; Lara et al., 1981). El conocimiento de los hábitos alimenticios de estas especies es importante debido a que por su abundancia desempeñan una función importante en la conducción, intercambio, transferencia y almacenamiento de energía dentro y fuera de los sistemas costeros (Yáñez-Arancibia y Nugent, 1977). Así también, a partir de esta información es posible realizar un manejo óptimo de los recursos pesqueros, ya que forman parte de un sistema multiespecifico, en donde las interacciones predador-presa influyen directamente en las pesquerías (Pauly, 1986). A pesar de la importancia en el conocimiento de los hábitos alimenticios de esta familia de peces, poco han sido los estudios sobre este tópico (Darnell, 1958; Harris y Rosse, 1968; Odum, 1971; Day, 1973; Odum y Heald, 1972; Yáñez-Arancibia, et al., 1976) y mucho menos, los que analicen y discutan el espectro trófico de estas dos especies (Yáñez-Arancibia y Lara, 1988; Lara, et al., 1981). La revisión de la literatura refleja el poco conocimiento que actualmente existe sobre la trofodinámica y la interacción ecotrófica entre Arius felis y Arius melanopus y con otras especies que se encuentran dentro del sistema del que son parte. Dada la importancia que tiene este tipo de investigaciones para un manejo adecuado a los recursos pesqueros, se pretende aquí determinar en una base estacional el espectro trófico de A. felis yA. melanopus en el crecimiento. Así también estimar el grado de interacción trófica que existe entre estas dos especies y discutir el nivel real del translape y las estrategias ecológico-adaptativas que les han permitido coexistir. MATERIALES Y MÉTODOSEl material para el presente trabajo fue obtenido de las capturas comerciales que se realizan con chinchorro playero en las costas del Puerto de Celestún. Dicho Puerto está ubicado en el litoral occidental de la Península de Yucatán a los 2º 52' latitud norte y 90º 24' longitud oeste (Fig.1). Del total de la captura se tomó mensualmente una muestra de ambas especies para los períodos octubre-diciembre de 1986 y Febrero-Julio de 1987. Se analizaron un total de 463 ejemplares juveniles y adultos, a los que se les determinó, individualmente, el peso húmedo total, peso eviscerado y longitud patrón. Inmediatamente después de realizar la disección a cada uno de los peces, los estómagos se separaron del resto de las visceras y se preservaron en formalina comercial al 10%.  Figura 1. Localización del Puerto de Celestùn en el Estado de Yucatán, México. Las dietas se determinaron por el análisis cuantitativo del contenido estomacal con la metodología ya descrita por Yáñez - Arancibia (1976) y Lara (1981). En la tabla 1 se enlistan las categorias tróficas registradas de las presas que se identificaron al nivel taxonómico más bajo posible, dependiéndo del grado de digestión en que se encontraban los organismos. Los datos obtenidos fueron agrupados en clase de longitud para ambas especies y la composición de cada grupo trófico se evaluó como el porciento del área total del contenido estomacal. El valor relativo de cada tipo de alimento, junto con la frecuencia de ocurrencia (Bowen, 1983; Overstreet y Heard, 1984), se emplearon para determinar el espectro alimenticio y sus variaciónes con respecto a la época del año (secas, lluvias y nortes). El índice de frecuencia F (Albertine, 1973; Yáñez-Arancibia, 1975) se obtuvo para categorizar los tipos alimenticios como -preferencial (F> 0.5), secundario (0.1> F< 0.5) y accidental (F<0.1). Se estimó la diversidad trófica (Shannon y Weaver, 1963) y la equidad (Pielou, 1966) para comparar el grado de especifidad en la dicta durante la ontogenia de ambas especies y en las tres épocas del año. La evaluación de las diferencias y semejanzas tróficas a través del crecimiento y entre las dos especies, se realizó utilizando el índice de Morisita (1959). Mediante el mismo procedimiento se estimó el grado de interacción trófica de ambas especies: 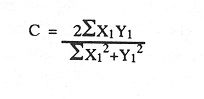 Donde X1 y Y1 son las proporciones de la categoría alimenticia en la especie X y Y respectivamente. El Indice varía de 0 cuando no hay alimentos en común hasta 1 cuando todos los artículos son comunes para ambas especies y se encuentran en las mismas proporciones.  TABLA 3 LISTA DE CATEGORÍAS TRÓFICAS GENERALES REGISTRADAS EN LOS ESTÓMAGOS DE Arius melanopus y Arius felis, CON LAS CLAVES EMPLEADAS EN LOS HISTOGRAMAS QUE REPRESENTAN LOS HÁBITOS ALIMENTICIOS DE CADA ESPECIE RESULTADOS
COMPOSICIÓN TRÓFICAAl seguir el criterio del Indice de Albertine (1973) que toma en cuenta la frecuencia y representatividad de un determinado grupo trófico en la dicta de cada especie, se observó que ambos bagres dentro de su estado carnívoro, consumen preferentemente a los crustáceos y peces. En Arius melanopus los crustáceos constituyen el 41% del total de la dieta. Dentro de este grupo los decápodos, ostrácodos, los tanaidaceos representados por la Familia Apseudidae y anfípodos del géneroAmpelisca se presentan con una ocurrencia del 2.4 al 20%. Mientras que los peces constituyen un 25.2% del total y son consumidos con un 17% de ocurrencia. En este componente, las sardinas Opisthonema oglinum y Harengula jaguana son las presas más abundantes, con una frecuencia siete veces mayor que la deAnchoa hepsetus. Las otras categorías que se incluyen dentro de su alimentación son los poliquetos, moluscos y detritus, los cuales son considerados de naturaleza secundaria, mientras que los moluscos y materia vegetal como fuentes accidentales (Tabla 2). Arius felis presenta un aspectro trófico muy similar al de A. melanopus en donde los crustáceos, siguen siendo las presas consumidas con mayor frecuencia (57.2%). Sin embargo, para esta especie, los peces constituyen una fuente alimenticia de gran importancia en su dieta al constituir un 38% del contenido estomacal. Dentro de los crustáceos, los decápodos son consumidos preferentemente (17.6%), mientras que de los peces, Anchoa hepsetus y Opisthonema oglinum son las presas de mayor importancia (18.9 y 13.5% respectivamente). Se considera el consumo de poliquetos y moluscos de naturaleza secundaria, y el de detritus y materia vegetal accidental por los bajos valores de ocurrencia que presentan (Tabla 2). Al analizar los espectros tróficos de ambos bagres se observa que ambas especies consumen básicamente los mismos tipos alimenticios. Sin embargo, se encuentran diferencias en las proporciones a nivel de grandes grupos. En Arius melanopus, los crustáceos son el grupo trófico más abundante (41%) y el consumido con mayor frecuencia (37%) mientras que en A. felis los crustáceos y peces se presentan en la misma proporción del total del contenido estomacal (38%). 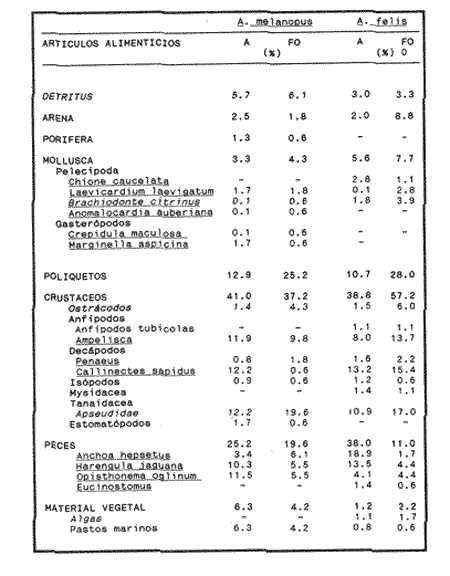 TABLA 2 FRECUENCIA DE OCIRRENCIA (FO) Y PORCENTAJE PROMEDIO ANUAL POR ÁREA (A) DE LOS DIFERENTES ARTÍCULOS ALIMEMTICIOS PRESENTES EN EL CONTENIDO ESTOMACAL DE Arius melanoptus y Arius felis ONTOGENIA TROFODINÁMICALas proporciones relativas de los artículos alimenticios que son consumidas por estas especies varían con la talla del pez. Esta variación trófica se denota claramente en el análisis de similitud en donde se observa paraArius felis la formación de tres grupos (Fig.3). El primero con ejemplares entre 121-140 mm, cuya alimentación se basa principalmente en tanaidaceos, anfípodos, decápodos y poliquetos. Con el crecimiento (141 - 160 mm) esta especie incrementa el consumo de anfípodos e inicia el de peces, mientras que al inicio de la madurez gonádica (170 mm) que corresponde a la segunda etapa trófica, se presenta la mayor diversidad en su alimentación (3.0 bits) al agregar detritus, ostrácodos, mysidaceos y moluscos en su dieta e incrementar el consumo de peces como Harengula jaguana y Anchoa hepsetus. En la tercera unidad trófica, el papel que desempeñan los peces se incrementa hasta alcanzar el 100% de la dieta en ejemplares mayores de 300 mm, siendo este componente la única fuente de energía. Por el contrario, con el aumento de talla el consumo de invertebrados bentónicos se reduce gradualmente y se registra la diversidad más baja. (Fig. 3). En Arius melanopus esta variación ontogenética en la alimentación no es tan marcada. Sin embargo, también se denotan tres etapas. En el primer grupo, los juveniles (80-140 mm L.P.) basan su alimentación en el bentos, en donde los anfípodos, tanaidaceos y poliquetos constituyen su fuente de alimento principal. En esta etapa de crecimiento, se inicia el consumo de peces, el cual se incrementa con la talla del pez para conformar la segunda etapa trófica (161-260 mm) que se caracteriza por presentar la más alta diversidad (3.0 bits) al incluir en su dieta el mayor número de grupos tróficos. La tercera etapa la conforman ejemplares de más de 261 mm los cuales disminuyen la amplitud de su espectro trófico al basar su alimentación principalmente en el consumo de materia vegetal, poliquetos y peces. (Fig. 4). Mientras que esta progresión ontogenética representa una continuidad, se observa para ambas especies un patrón trófico estable que va desde una especifidad en invertebrados bentónicos durante la etapa juvenil a una especialización de los ejemplares adultos en el consumo de peces, pasando por una etapa intermedia de generalización en donde se incluyen el detritus y la materia vegetal. Este comportamiento trófico se observa claramente en los valores de diversidad registrado en las dos especies para cada intervalo de talla. Los valores máximos se presentan en las etapas intermedias y los mínimos en los ejemplares juveniles y adultos. (Figs. 3 y 4). 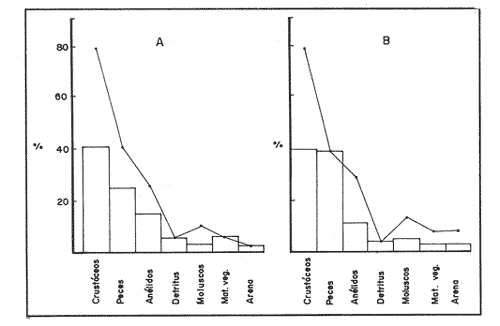 Figura 2. Hábitos alimenticios de Artus melanopus (A) y Arius felis (B). La barra representa la importancia relativa de cada tipo alimenticio y el polígono la frecuencia relativa respectiva. A pesar de que ambas especies presentan en su ontogenia un patrón trófico muy similar. Se observan diferencias entre ambas especies respecto al grado de generalización y especificidad de su alimentación. Los juveniles de Arius melanopus son más específicos en el consumo de pequeños invertebrados bentónicos, mientras que los adultos de Arius felis en el consumo de peces. Este análisis realizado en una base anual, indica que las variaciones temporales que originan los cambios en abundancia y biomasa de las presas, se refleja en la ontogenia trófica de estas especies durante las épocas de secas, lluvias y nortes. TEMPORALIDAD DE LA UNIDAD TRÓFICADe la misma manera que en el análisis trófico anual, temporalmente ambos bagres presentan una progresión ontogenética en su alimentación, que se observa en los valores de diversidad trófica registrados para las dos especies y para las tres épocas del año. Independientemente de la estación anual y de la especie de bagre, la diversidad trófica más baja se registra en los ejemplares juveniles al consumir preferentemente pequeños invertebrados y en los adultos al constituir los peces su alimento principal. Los ejemplares de tallas intermedias se consideran generalistas tróficos al ampliar su espectro y en consecuencia presentan la más alta diversidad (3 bits). (Fig. 5 y Tabla 3). Sin embargo, de las tres épocas del año, es en la estación de secas, en donde se observa más claramente este patrón trófico al alimentarse los adultos de ambas especies de bagre exclusivamente de peces y los juveniles principalmente de poliquetos, anfípodos y decápodos (Fig.6). En lluvias soloA. felis tiende a la especialización en el consumo de peces, mientras que A. melanopus los consume principalmente en etapa juvenil (Fig.7). Por el contrario en la época de nortes ambos bagres presentan una menor tendencia a la especialización al incluir en su alimentación además de esta categoria trófica a los peneidos y branquiuros como sus presas principales. (Fig.8). Estas diferencias temporales en el espectro trófico de ambas especies, originan que en la época de lluvias y nortes, el translape en la dieta entre ambas especies sea menor que en la época de secas, en donde los dos bagres consumen básicamente los mismos tipos alimenticios y ocurre la mayor diversidad trófica. (Tabla 3). 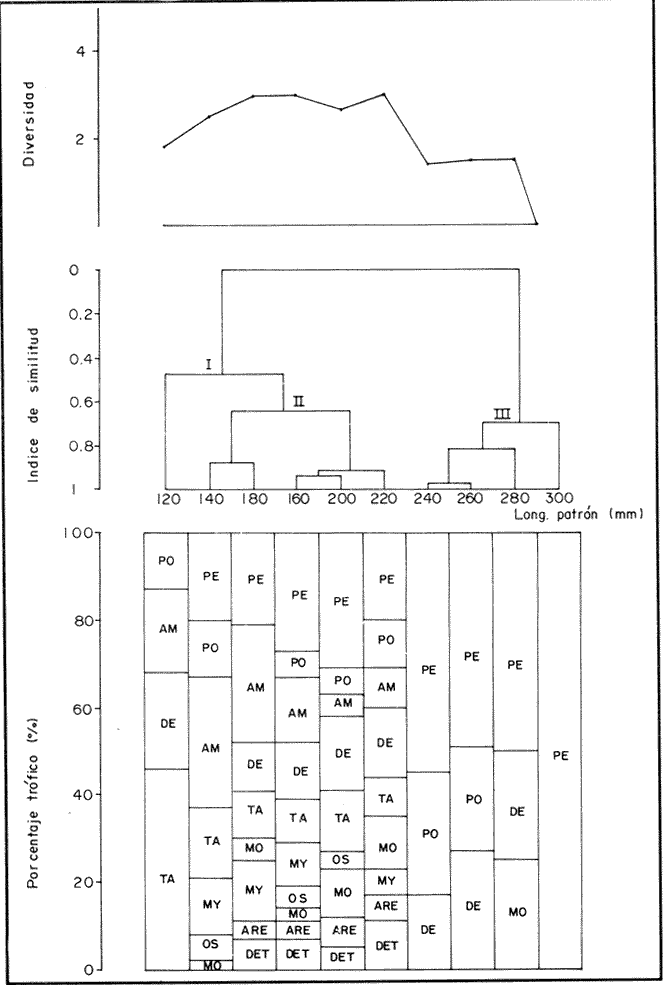 Fig 3. Cambios ontogonéticos en la dicta de Arius felis. El histograma representa ha propordones relativas de los principalea componentes tróficos y el dendograma la similitud trófica entre las clases de talla. Se represen la diversidad trófica a través de la ontogenia. El código de ha categorías tróficas se representa en la tabla 1. 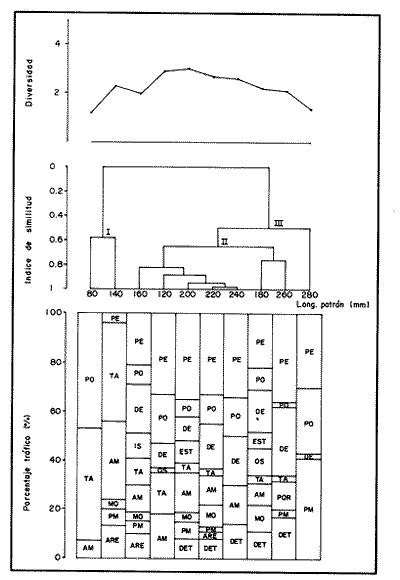 Figura 4. Cambios ontogenéticos en la dicta de Arius melanopeus. El histograma representa las proporciones relativas de los principales componentes tròficos y el dendograma la similitud trófica entre las de talla. Se representa la diversidad trófica a través de la ontogenia. El código de las categorias tròficas se representa en la tabla 1. 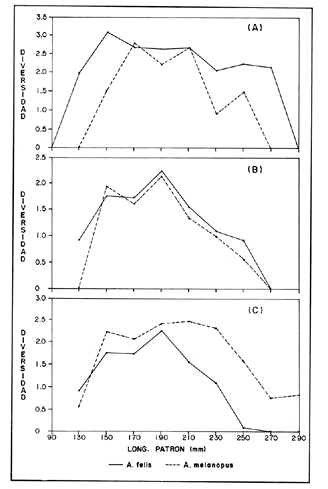 Figura 5. Representación comparativa de la diversidad trófica registrada en las épocas de secas (A), lluvias (B) y nortes (C) entre Arius felis (---) y Arius melanopus ( ---- ). 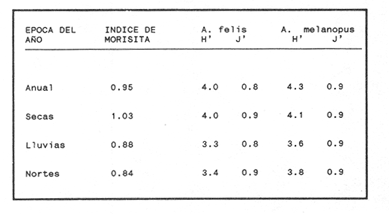 TABLA 3 VARIACIÓN ANUAL Y ESTACIONAL DEL TRANSLAPE Y AMPLITUD DE LA DIETA DE Arius felis Y Arius melanopus SE UTILIZARON LOS ÍNDICES DE MORISITA (1959), DE DIVERSIDAD (H'), (SHANNON Y WEAVER, 1963) Y EQUIDAD (J') DE PIELOU (1966) En el análisis trófico temporal, se observa para ambas especies una relación inversa en el consumo de peces y de crustáceos. En la época de secas ambas categorías tróficas constituyen el alimento principal. Sin embargo, al llegar los nortes, se registra un decremmento de hasta 3 veces en la abundancia y ocurrencia de los peces e inversamente se duplica el consumo de crustáceos. (Tablas 4 y 5). Las otras categorías alimenticias presentan ligeras variaciones temporales. En A. felis los anélidos se incrementan gradualmente desde la época de secas a los nortes, mientras que enA. melanopus disminuyen en la época de lluvias y se incrementan en nortes. Los moluscos y la materia vegetal se mantienen estables en los das bagres y continúan siendo de naturaleza accidental. DISCUSIONEl análisis trofodinámico Arius felis y Arius melanopus presenta a los crustáceos y peces como su fuente principal de alimento. Las dos especies de bagre utilizan los mismos recursos. Sin embargo, es un hecho que los explotan en tiempos y situaciones diferentes, reduciendo así el nivel real de translape entre ellas. Las migraciones estuarinas que realizan como estrategias alimenticias y reproductivas les permiten variar sus patrones de distribución espacial y temporal (Yáñez Arancibia y Lara 1988; Lara, et al., 1981). Al respecto, Margalef (1978) señala que los peces ocupan varios nichos simultáneamente al cambiar su alimentación con el crecimiento y al presentar patrones espaciales o temporales de repartición de recursos. Los resultados indican que el patrón trófico y ontogenético de ambas especies de bagre, comprende tres etapas: 1) una especialización en el consumo de pequeños crustáceos como anfípodos y tanaidaceos; 2) una generalización trófica que incluye la utilización de diversos crustáceos, poliquetos y peces, así como el consumo de pequeñas cantidades de moluscos y materia vegetal; 3) finalmente, una etapa que presenta la tendencia hacia una especificidad en el consumo de peces, acompañada por un decremento en la ingestión de crustáceos. Estos resultados están de acuerdo con los encontrados por Livingston (1982) quien indica que las poblaciones con progresiones alimenticias complejas, tienden a ser especialistas tróficos durante las últimas etapas de crecimiento. Este patrón presenta diferencias estacionales que se relacionan con las condiciones climáticas que imperan y en consecuencia con la abundancia de las presas. En el período de secas no existen diferencias estacionales interespecíficas y el nivel real de translape entre ambas especies es el más elevado (1.0) al compartir básicamente los mismos recursos alimenticias y predominar el consumo de peces. Los altos valores de translape: no una evidencia suficiente de competencia, para ello es necesario considerar el suministro alimenticio debido a que sólo la escases de recursos originaría que especies tróficamente similares lleguen a competir por el alimento (Pianka, 1976). Al respecto, en estudios anteriores realizado en el Sur del Golfo de México, Yáñez-Arancibia y Sánchez-Gil (1986, 1988) detectaron para la época de secas las máximas capturas de pesca de escama (Yáñez-Arancibia y Sanchez-Gil, 1986, 1988). Así también, en los análisis tróficos realizados, se encontró que las bagres consumen el mayor porcentaje de peces en esta época, lo que probablemente esta relacionado con la máxima abundancia de estas especies (Tabla 3 y 4). Estas antecedentes y resultados hacen considerar que independientemente del alto nivel del translape que se presenta entre estas dos especies de bagre (1.0), no se puede hablar de competencia por el alimento, al ser los peces tomados como presa un recurso que se encuentra en abundancia en esta época del año. Esta conclusión está de acuerdo con lo mencionado por Krast (1978) y Livingston (1982) Werner y Hall (1976), quienes indican que el translape ocurre entre especies muy relacionadas y que tiende a ser mayor durante períodos de un incremento en la disponibilidad del alimento al ser las dos especies atraídas hacia el mismo recurso. 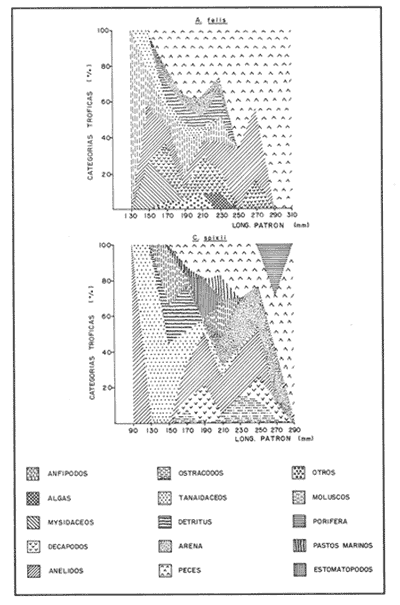 Figura 6. Variación del espectro trófico de Arius felis y Arius melanopus durante la época desecas. 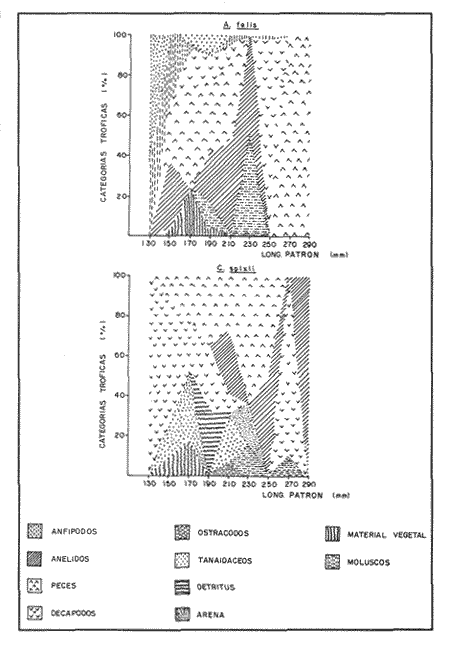 Figura 7. Variación del espectro trófico de Arius felis y Arius melanopus durante la época de lluvias. 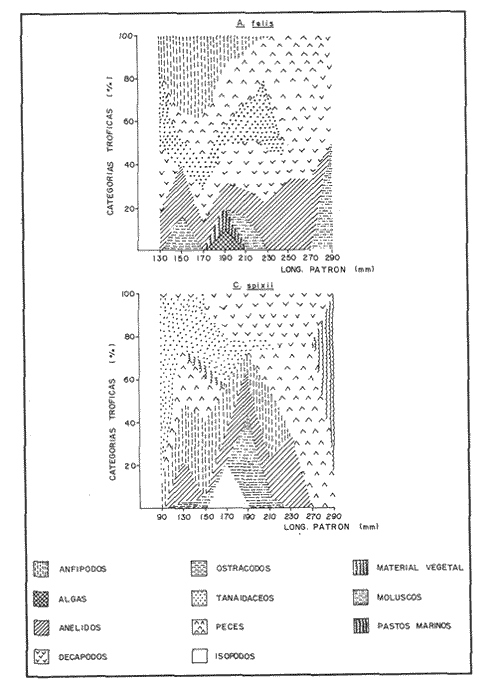 Figura 8. Variación da espectro trófico de Arius felis y Arius melanopus durante la época de nortes. 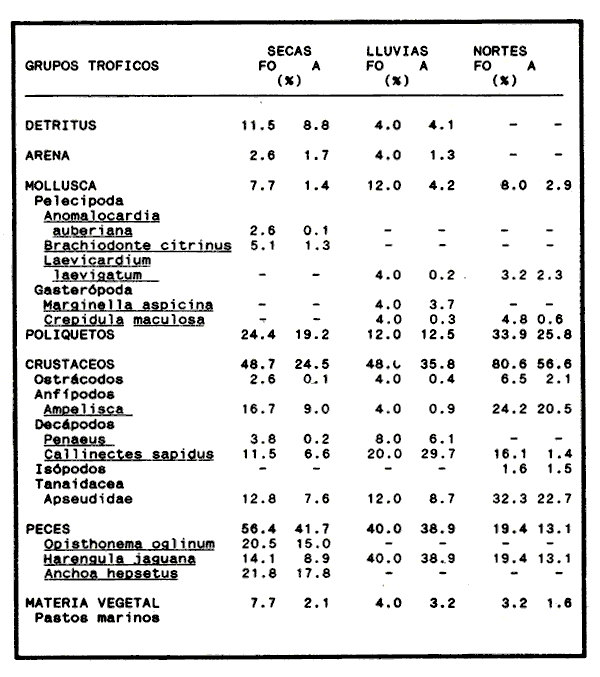 TABLA 4 FRECUENCIA DE OCURRENCIA (FO) Y PORCENTAJE PROMEDIO ANUAL POR ÁREA (A) DE LOS GRUPOS TRÓFICOS REGISTRADOS EN Arius melanopus DURANTE LAS ÉPOCAS DE SECAS, LLUVIAS Y NORTES Por el contrario, en la época de "nortes" la disminución en el consumo de peces y la incorporación de los peneidos, braquiuros y poliquetos en la alimentación de ambos bagres, es originado por las condiciones meteorológicas que se originan en esa época que provocan la migración de los peces a zonas portegidas o de mayor profundidad (Leggett, 1984; Yáñez-Arancibia y Sanchez Gil, 1988). Lo anterior demuestra que la fluctuación temporal de los recursos parece estar asociada con las progresiones estacionales de las preferencias alimenticias dentro de las dos especies. Este cambio estacional en la dieta de los peces en relación con la variación temporal en la distribución y abundancia de las presas ha sido reportada y discutida en estudios similares sobre la alimentación de diversas especies de climas templados y trópicales (Yáñez-Arancibia, 1976; Stoner, 1980: Livingston, 1984; Collie, 1987; Hopkins y Baird, 1977; Young y Blaber, 1986). Por otro lado, la variación trofidinámica con el crecimiento, reduce la competencia intraespecífica y permite, como menciona Sheldon (1969), una utilización más eficiente de los recursos del hábitat dependiendo de su disponibilidad. Esta variación intraespecífica en la dieta de estas dos especies de bagre dentro de estados ontogenéticos individuales, indica que estos peces tienen una gran flexibilidad en su conducta alimenticia. Asimismo, es la utilización de un gran número de recursos los que probablemente explica la habilidad deA. felis y A melanopus para adaptarse exitosamente a diferentes hábitats. Al respecto, Sierra (1986) considera que la mayoría de los peces tropicales son oportunistas tróficos en virtud de la gran diversidad de habitos alimentarios que presentan a través de su ontogenía. Sin embargo, dentro de este marco de generalización trófica los ejemplares adultos de ambas especies de bagre tienden a la especialización en el consumo de peces y son selectivos en cuanto a las especies que son tomadas como presa. Exclusivamente se alimentan de dos clupeidos (Opisthonema oglinumy Harengula jaguana), un engraulido(Anchoa hepsetus) y representantes del géneroEucinostomus. El grado de especifidad en su consumo presenta variaciones interespecificas. Comparativamente en A. felis se observa una tendencia mayor a la especifidad en el consumo de peces, mientras que en A. melanopus la generalización trófica es más evidente (Fig. 3 y 4). Esta observación se manifiesta más claramente los resultados de diversidad obtenidos para ambas especies en donde A. felis presenta los valores más bajos (Tabla 5). 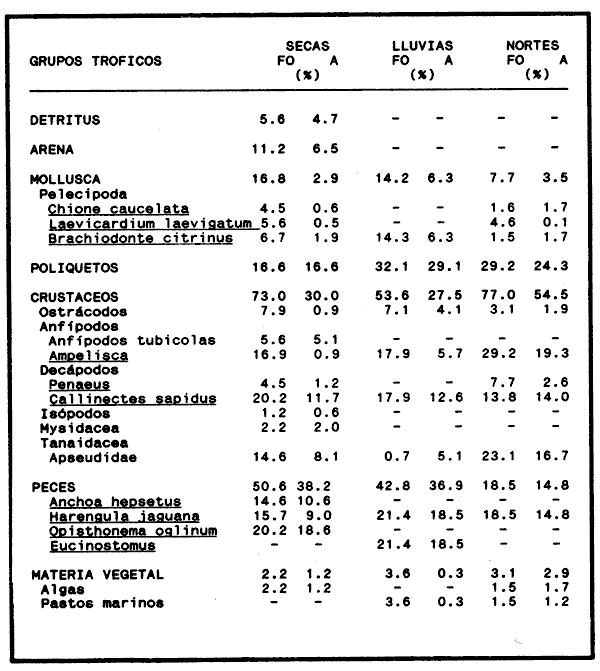 TABLA 5 FRECUENCIA DE OCURRENCIA (FO) Y PORCENTAJE PROMEDIO ANUAL POR ÁREA (A) DE LOS GRUPOS TRÓFICOS REGISTRADOS EN Arius felis DURANTE LAS ÉPOCAS DE SECAS, LLUVIAS Y NORTES Los resultados de este estudio indican que los bagres son especies carnívoras cuyos patrones alimenticios ontogenéticos comprenden varios niveles tróficos Su trofodinámica está en sincronía con la disponibilidad del alimento, las características del hábitat, las condiciones estacionales que influyen directamente en la productividad del sistema, el estado de madurez y el estado de desarrollo. Esta sincronía específica, junto con diferencias en sus patrones espaciales y temporales de distribución, son las que permiten la coexistencia de estas dos especies tan similares en su trofódinámica. El estudio ecotrófico de A. felis yA. melanopus realizado en una base estacional y en su ontogénia, revela que ambas especies son los predadores principales de crustáceos de importancia comercial como peneidos y branquiuros(Callinectes sapidus) y de pequeños cardúmenes en la zona costera del Golfo de México. La dominancia de los clupeidos y engraulidos dentro de los componentes tróficos de estas dos especies apoyan esta observación. Se concluye que la sincronía en la ontogenia de las especies con las variaciones estacionales en la distribución y abundancia de las presas, es el mecanismo por el cual, se optimiza la repartición de los recursos disponibles, disminuyendo la competencia inter-intraespecífica y permitiendo la coexistencia de especies tróficamente similares en un mismo ambiente. AgradecimientosSe agradece a la compañera Mireya Hernández de Santillan su colaboración en la colecta y análisis de las muestras. Al M. en C. Gustavo de la Cruz por la guía y revisión de manuscrito y al CINVESTAV-IPN, UNIDAD MÉRIDA por el apoyo brindado para el desarrollo de este trabajo. Los resultados de esta Investigación forman parte del Proyecto "Investigación para el manejo y administración racional de las pesquerías tropicales". Proyecto apoyado por COSNET. LITERATURAALBERTINE, B.J. Aquaculture Biologic des stade juveniles de téleostésne mugilidae Mugil auratus. Risso 1810. Mugil capito, Cuvier 1829 et Mugil saliens Risso 1810. J. Régime alimentaire. 1973. 251 - 266.2: BORUVSKI, E. V. Manual para el estudio de la alimentación y relaciones alimentarias de los peces en condiciones naturales. Editorial Nauka Moscú1974. 254 p. BOWEN, S.H. Fisheries Techniqques. Quantitative description of She diet In: Nielsen LA. y O.L. Johnson (Eds). American Fisheries Society Bethesda.1983.325-336. COLLIE, SJ. Food consumption by yelowtail flounder in relation to production of its benthic prey. Mar. Ecol. Prog. Ser.1987.205-213. 36: DARNEL, R.M. Publ. Inst. Mar. Sci. U. of Texas Food habitas of fishes and larger invertebrates of Lake Ponchartrain, Louisiana, an estuarine community.1958. 353 - 416. 5: DAY, J.W. W.G. SMITH y C.S. HOPKINSON Proc. Coastal Marsh and Estuary Symposium. Some trophic relationships of marsh and estuarine areas. Lousiana State University Baton Rouge, La. 1973. 115 - 135. Jul. 1972 HARRIS, A.H. y C.D. ROSSE Shrimps predation by the sea catfish Galeichthy felis Trans. Am. Fishc. Soc. 1968.503-504.97 (4): HOPKINS, T.L. y R.C. BAIRD Oceanic sound scattering prediction. Aspects of the feeding ecology of oceanic midwater fishes. In.: Anderson W.R. y B.J. Zaburance (Eds) Plenum Press New York 1977.325-360. KEAST, A. Biol. Fish. Trophic and spatial interrelationships in the fish species of an Ontario temperature lake. Environ. 1978. 7 - 31. 3: LARA A.L. A. YAÑEZ-ARANCIBIA, F. AMEZCUA An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Biología y ecología del bagre Arius melanopus Gunther, en la Laguna de Términos. Sur del Golfo de México (Pisces: Ariidae). 1981. 267 - 304. 8 (1) LEGGETT, W.C. Mechanisms of Migration in Fishes. Nato Conference Series. Fish migration and coastal and estuarine enviroments: A callo for new approaches to the study of an old problem. Series IV: Marine Sciences In: McCleave, J.D. G.P. Arnold, J.J. Dodson y W.H. Neill (EDS). 1984. 159 - 178. 14: LIVINGSTON, J.R. Trophic organization of fishes in a coastal seagrass system. Mar Ecol. Prog. Ser. 1982. 1 - 12.7 LIVINGSTON J.R.Ecology Trophic response of fishes to habitat variability in coastal seagrass systems.1984. 1258 - 1265. 65(4) MARGALEF, R. Marine Ecology General concepts of population dynamics and food links. In: Kinne, O. (ED). John Wiley y Sons. Chichester, Toronto.1978. 617 - 704. vol. 4. MORISITA, M. Measuring interespecific association and similiatity between communities. Ser. Ed. (Biol.). Mem. Fac. Sci. Kyushu. Univ. 1959. 65 - 0. 3 ODUM, E.W. Tech. Bull. Pathways of energy flow in a South Florida estuary Ph. D. Thesis.Univ. of Miami. Sea Grant Program.1971. 162p. No. 7 ODUM, W.E. y E.J. HEALD Bull. of Marine Science.Trophic analyses of an estuarine mangrove community.1972. 671 - 738. 22(3): OVERSTREET, M.R. y R.W. HEARDGulf Researcis reports Food contents of six commercial fishes from Mississipi sound.1982. 137 -149. 7 (2): PAULY, D. Living Resources. Problems of Tropical Inshore Fisheries: Fishery Research on tropical Sift Bottom Communities and the evolution of its Conceptual Base. In: E. Mann Borgese y N. Ginsburg (Eds). University of Chicago Press. 1986. 29 - 37. PIANKA, E.R. Competition and niche theory.Theorical Ecology, principies and aplications. W.B. Saunders, Philadelphia and toronto In: May, R.M. (Ed) 1976.114 - 141. PIELOV, E.C. J. Theort. biol. The measurement of diversity indifferent types of biological collections. 1966.131 -144. 13: SALOMAN, H.C. y S.P. NAUGHTON, Food of crevalle jack (Caranx hippos) from Florida NOAA. Tech. Mem. NMFS - 59 FC Louisiana and Texas. 1984. 34 p.134 SIERRA, L.N. y J. DIAZ ZABALLA. Reporte de Inv. del Inst. de OceanologíaAlimentación de dos especies de sardina, Harengula humeralis (Cuvier, 1829) y Harengula clupeola (Cuvier, 1829) en la costa Norte de la Ciudad de la Habana (rada del Instituto de Oceanografía).1984. 18 p. 25 SIERRA, L.M. Tesis Doctorado. Ecología de la alimentación de algunos peces herbívoros de Cuba. Univ. de la Habana Cuba.1986. 23 p. SHANNON, E.C. y WEAVERThe Mathematical Theory of communication.Univ. of Illinois Press Urbana 1963. 119 p. SHELDON, A.L.Hydrobiology Size relationship of Acroneuria california (perlidae, Plecoptera) and its prey. 1969. 85 - 94. 34 STONER. W.A. Fishery Bulll. Feeding ecology of Lagodon rhomboides (Pisces: Sparidae): variation and functional responses. 1980 337-352. 78(2): WERNER. E.E. y DJ. HALL. Science, Niche shifts in sunfishes: experimental evidence and significance.N.Y. 1976. 404 - 406. 191: YAÑEZ-ARANCIBIA. A. In: Informe final de la "Segunda etapa del programa de uso de la zona costera de Michoacán y Guerrero". Relaciones tróficas de la Fauna ictiológica del sistema lagunar costero de Guerrero y aspectos parciales de dinámica de poblaciones de los peces de importancia comercial 230 p Conv. Com. del Río Balsas, SRN y Centro de Ciencias del Mar y Limnología. UNAMMéxico 1975. 750 p. YAÑEZ-ARANCIBIA, A.An. Centro. Cienc. del Mar. y Limnol Univ. Nal Autón. México Observaciones sobre Mugil curema Valenciennes. en áreas naturales de crianza, México. Alimentación, madurez, crecimiento y relaciones ecológicas. 1976. 92 - 124.3: YAÑEZ-ARANCIBIA, A. J. CURIEL, V. LEYTON An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México Prospección biológica y ecológica del bagre marino Galeichthys caerulescens (Gunter) en el sistema lagunar costero de Guerrero, México (Pisces: Ariidae). 1976. 125 - 180. 3 (1): YAÑEZ-ARANCIBIA, A. y R.S. NUGENT An. Centro Cienc. del Mar y Limnol. Univ. Nal. Auton. México. El papel ecológico de los peces estatuarios y lagunas costeras. 1977. 107 -114 p.4: YAÑEZ-ARANCIBIA, A. y P. SANCHEZ-GIL Los peces demersales de la plataforma continental del Sur del Golfo de México, caracterización del ecosistema y ecología de las especies, poblaciones y comunidades. Inst. Cienc. Del Mar y Limnol. Univ. Nal. Autón. México Pub. Esp.1986. 2-30 p. 9: vol. 1. YAÑEZ-ARANCIBIA, A. y P. SANCHEZ-GIL Ecología de los recursos demersales marinos, fundamentos en costas tropicales. A.G.T. editor. S.A. 1988.228p YAÑEZ-ARANCIBIA, A. y A.L. LARA-DOMINGUEZ Ecology of three sea ea tfishes (Ariidae) in a tropicalo coastal ecosystem-Southern Gulf of Mexico.Mar. Ecol. Prog. Ser. 1988.215-230. 49: YOUNG WJ. y SJ.M. BLABER Marine Biology Feeding ecology of three species of midwater fishes associated with continental slope of eastern Tasmania. Australia.1986. 147 - 156. 93:
|