|
CRECIMIENTO Y MORTALIDAD DE
LA RUBIA Lutjanus synagris DE LAS COSTAS DE YUCATÁN DURANTE LAS
TEMPORADAS DE PESCA 1983-1985
Trabajo recibido el 17 de octubre de 1990 y
aceptado para su publicación el 20 de septiembre de
1990.
Ricardo Torres-Lara
Delegación Federal de Pesca en
Yucatán
Silvia Salas-Márquez
Centro de Investigación y
Estudios Avanzados del IPN-Unidad Mérida.
Se determinaron
los parámetros de crecimiento de la rubiaLutjanus
synagris de las costas de Yucatán durante las temporadas de
pesca 1983, 1984 y 1985, realizándose un análisis comparativo de
los parámetros de crecimiento k y Loo, obtenidos mediante la
aplicación de retrocálculo, del paquete ELEFAN I (Brey y Pauly,
1986) y del Método de Harding (Cassie, 1954). Considerando los
resultados obtenidos con el Programa ELEFAN I, se hizo la comparación
entre las diferentes temporadas, obteniéndose valores de k que
fluctuaron de k = 0.250 en 1983, a k = 0.275 en 1985. Se determinó
asimismo, a través de la relación radio de la escama-longitud
furcal del pez, la longitud teórica de inicio de formación de las
escamas que es de 8.9 mm. El valor de la mortalidad natural no mostró
grandes variaciones, estimándose valores de M = 0.3045, M = 0.3368 y M =
0.3575 en 1983, 1984 y 1985 respectivamente; por lo que las variaciones en la
mortalidad total se atribuyen a los diferentes valores de la mortalidad por
pesca: F = 1.3157 en 1983, F = 0.4198 en 1984, y F = 0.2175 en 1985. Los
patrones de reclutamiento y las capturas mensuales exhibieron un perfil muy
similar en las tres temporadas analizadas.
During the
fishing seasons 1983, 1984 and 1985, the growth characieristics of the lane
snapper Lutjanus synagris were determined, carrying out a
comparative analysis of the growth coefficient k, and the asymptotic length
Loo, by means of the Program ELEFAN I (Brey and Pauly, 1986), and Harding's
Metbod (Cassie, 1954). k values ranged from k = 0.247 in 1983 to k = 0.275 in
1985. Through the scale diameter-fork length ratio, the theoretical lenght at
the beginning of the scale formation was 8.9 mm. The natural mortality was
almost the same in the three seasons: M = 0.3045 in 1983, M = 0.3368 in 1984,
and M = 0.3575 in 1985; then, the total mortality values were different from
year to year due to the variation in the fishing mortality values: F = 3.157 in
1983, F = 0.4198 in 1984, and F = 0.2175 in 1985. The recruitment patterns and
monthly captures were similar in the analyzed period.
La descripción cuantitativa de ciertas variables poblacionales vitales referidas con respecto al tiempo, es una parte necesaria en la mayoría de los modelos de explotación pesquera. En muchos casos, estas variables se miden sobre el ciclo de vida de los organismos; es por ello que la edad de los peces se utiliza como una medida de tiempo necesaria para la estimación de tales variables, como la mortalidad y el crecimiento. En la adquisición de la información necesaria para determinar los parámetros de crecimiento de los peces se presentan problemas de distinta índole, tales como la obtención de estructuras duras en algunas especies, de tal suerte que se han seguido diferentes enfoques para abordar tales problemas. Dado que algunos presentan ciertas ventajas sobre otros, debe realizarse una elección cuidadosa del método que mejor se adapte a la biología de la especie en estudio. Sin embargo, una práctica común es la aplicación de dos o más métodos, con el fin de lograr bases más sólidas en la caracterización del crecimiento de los individuos (Beamish y McFarlane, 1983). En el presente trabajo se hace un análisis comparativo de los resultados obtenidos a través de retrocálculo del Método de Harding (Cassie, 1954), y de la aplicación del Programa ELEFAN I (Pauly y David, 1981; Brey y Pauly, 1986) para definir el crecimiento individual en la población de la rubia Lutjanus synagris, de la costa de Yucatán durante las temporadas de pesca 1983, 1984 y 1985. Adicionalmente, se estimaron las tasas de mortalidad y los patrones de reclutamiento que exhibió esta especie en el periodo mencionado. MÉTODOSLos datos utilizados en el presente estudio provienen de muestras tomadas de los puertos donde se desembarca la captura comercial de la flota arrastrera yucateca que operó en 1983, y la información de las dos temporadas siguientes (1984-1985), de la flota menor que captura con línea este recurso. Con base en un enfoque anatómico, haciendo uso de escamas, en las temporadas 1984 y 1985 se aplicó el método de retrocálculo cuya fórmula es la siguiente: Ln= (Sn/S) (L-c) + c donde Ln es la longitud a la cual se forma cada anillo, L es la longitud furcal (mm) a la que se capturó el pez, Sn es la distancia del foco a cada anillo (mm), S es la distancia del foco al borde de la escama (mm) y c es el factor de corrección (ordenada al origen cuando la relación no pasa por el cruce de ambos ejes). Con este método se obtuvieron las longitudes promedio de los distintos grupos de edad, y posteriormente los parámetros de crecimiento a partir de la Ecuación de Von Bertalanffy. Para la temporada de 1983 se aplicó un enfoque estadístico (haciendo uso de frecuencias de longitud), utilizando el Método de Harding (Cassie, 1954) para obtener las longitudes promedio en esta temporada. Bajo este mismo enfoque, se contó con el registro de la longitud furcal de 1,954 en organismos en 1983, 2,974 en 1984 y 4,124 en 1985. Estos datos fueron analizados mediante los Programas ELEFAN I y II (Brey y Pauly, 1986), de tal manera que además de obtener los valores de los parámetros de la ecuación de crecimiento de Von Bertalanffy, se realizaron estimaciones de la mortalidad total (Z) para cada temporada, mediante las curvas de captura modificadas que elaboran estos programas, los cuales permitieron también obtener los patrones de reclutamiento del periodo analizado. La mortalidad natural fue estimada para cada temporada mediante la ecuación empírica propuesta por Pauly (1980): Log M =0.0066 - 0.279* log L∞ + 0.6543* log k + 0.463* log T en la cual se relacionan los parámetros de crecimiento (Loo y k), y la temperatura ambiente promedio (T) del área donde habita esta especie utilizando 23ºC como valor promedio (García, 1980). La mortalidad por pesca se obtuvo como resultado de la diferencia de las dos anteriores (F=Z-M) y que incidieron sobre la población durante el lapso en cuestión. Por último, se estimó la longitud teórica a la cual comienzan a formarse las escamas, utilizando información de la temporada 1985 con base en la relación del radio de la escama y la longitud furcal del pez. RESULTADOSMediante la aplicación del Método de Harding (Cassie, op. cit.), se construyó la curva en papel de probabilidad que se observa en la figura 1 y que corresponde a los datos de 1983. El valor de k que se encontró para ese año (k= 0.247) fue similar a los correspondientes a 1984 (k= 0.253) y 1985 (k=0.260), obtenidos mediante el retrocálculo. Estos valores, a su vez fueron también semejantes a los estimados a través de los Programas ELEFAN: k = 0.250, k = 0.270 y k = 0.275, para 1983, 1984 y 1985, respectivamente. Con respecto a los valores estimados de la longitud asintótica, se obtuvieron los valores más altos en 1983 con los dos métodos empleados; no sin presentar grandes diferencias entre éstos (405-410). Para las dos temporadas posteriores, los valores resultantes de la aplicación de ambos métodos fueron diferentes, fluctuando entre 324 y 339.8 para 1984, y 330 y 357.3 para 1985 (Tabla l). Una vez obtenidos los parámetros de crecimiento, se calcularon las longitudes medias correspondientes a cada clase de edad (Tabla 2). Para las tres temporadas se obtuvo un mayor número de grupos de edad, resultado de la aplicación del ELEFAN. En 1983 se observan ligeras diferencias entre los valores estimados con ambos métodos en las clases de edad correspondientes (Fig. l), a diferencia de los dos años siguientes, donde los valores medios son similares con los métodos empleados, incluyendo las clases de edad IV a IX en 1984 (Fig. 2) y de la III a la VI en 1985 (Fig. 3). En general, los resultados de los parámetros de crecimiento no mostraron grandes variaciones entre los diversos métodos en los diferentes años, pero dado que en los tres años se aplicó el ELEFAN y la consistencia de los valores de la tasa de crecimiento (k) en los diferentes años y con lo reportado en la literatura (Torres, 1984; Torres y Chávez, 1987; Mexicano y Arreguín, 1987). Se utilizaron los parámetros derivados del ELEFAN para estimar las tasas de mortalidad y patrones de reclutamiento. Mediante el análisis de las curvas de captura del Programa ELEFAN 2A se observó una disminución en los sucesivos valores de la mortalidad total de la primera a la tercera temporada, de tal manera que los correspondientes fueron Z=1.6202, Z=0.5750; y como se puede observar en la Tabla 2, los valores de la mortalidad natural no variaron significativamente en los tres años, por lo que las diferencias se atribuyen a los cambios en la mortalidad por pesca. En relación a los patrones de reclutamiento obtenidos con el ELEFAN, se observaron para 1983 y 1984 dos picos máximos (Figuras 5 y 6) y aún cuando no coinciden con las capturas para dichos años, la tendencia es muy similar. Por último, y habiéndose observado una relación lineal entre la longitud furcal del pez y el radio de las escamas, cuya ordenada no pasa por el origen, se tomó el valor de ésta como la longitud a la cual comienza a formarse el foco de tales estructuras. En este caso sólo se utilizaron las escamas colectadas en 1985, ya que la muestra de ese año incluia organismos de tallas menores a los 190 mm, lo cual dio lugar a que la ordenada indicara un tiempo más temprano de formación de las escamas (a 8-9 mm) que el calculado con individuos mayores provenientes de muestras comercialmente seleccionadas (Carlander, 1982). Esta relación se observa en la Figura 8. 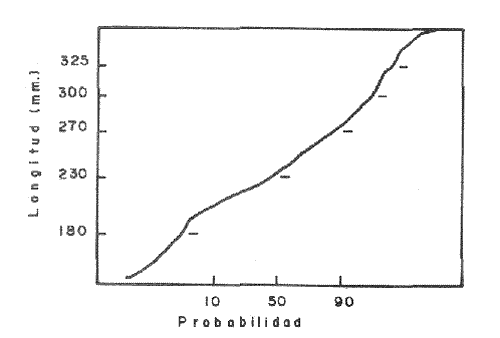 Figura 1. Estimación de clases de edad de Lutjanus synagris mediante el Método de Harding en la temporada 1983. 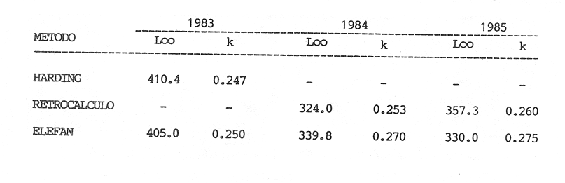 TABLA 1 VALOR DE LOS PARÁMETROS DE CRECIMIENTO POR MÉTODO Y AÑO DE L. synagris 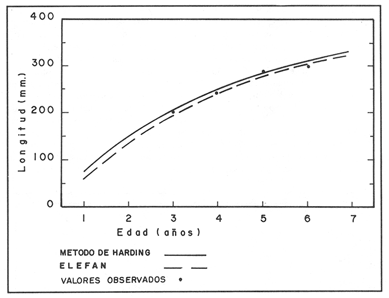 Figura 2. Curvas de crecimiento de Lutjanus synagris en la temporada 1983. 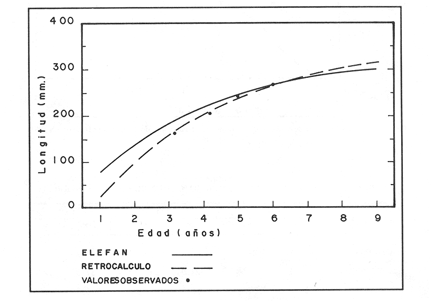 Figura 3. Curvas de crecimiento de Lutjanus synagris en la temporada 1984. 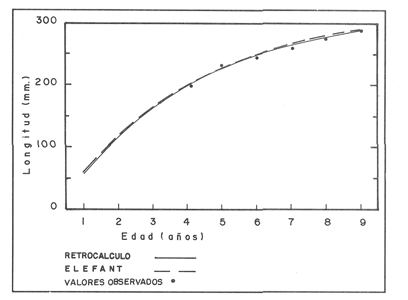 Figura 4. Curvas de crecimiento de Lutjanus synagris en la temporada 1985. 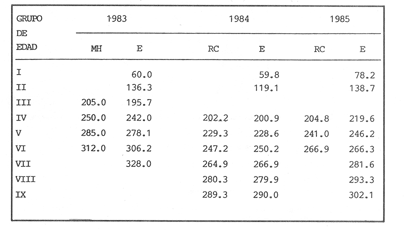 TABLA 2 LONGITUDES PROMEDIO PARA CADA GRUPO DE EDAD, CALCULADAS POR LOS MÉTODOS DE HARDINNG (MH), RETROCÁLCULO (RC) Y ELEFAN (E) DE L. sinagrys 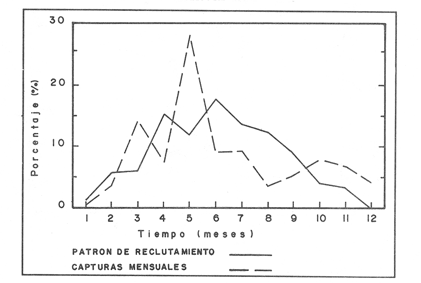 Figura 5. Comparación entre los patrones de reclutamiento y la tendencia de las capturas mensuales de Lutjanus synagris en la temporada 1983. 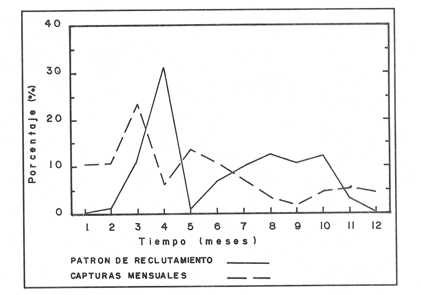 Figura 6. Comparación entre los patrones de reclutamiento y la tendencia de las capturas mensuales de Lutjanus synagris en la temporada 1984. 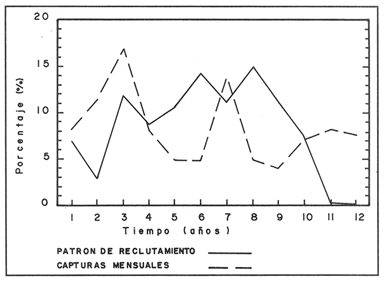 Figura 7. Comparación entre los patrones de reclutamiento y la tendencia de las capturas mensuales de Lutjanus synagris en la temporada 1985. 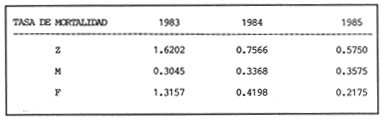 TABLA 3 VALORES DE LA MORTALIDAD TOTAL (Z) NATURAL (M) Y POR PESCA (F) DE L. synagris DE 1983 A 1985 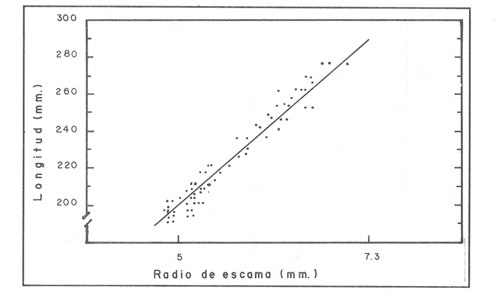 Figura 8. Estimación de la longitud teórica de inicio de formación de las escamas a partir de la relación radio de la escama-longitud furcal. DISCUSIÓN Y CONCLUSIONESLa estimación de los parámetros de crecimiento y de las tasas de mortalidad son de gran importancia en la evaluación de poblaciones ícticas bajo explotación, sobre todo cuando el objetivo final es proponer medidas de regulación para el manejo adecuado de tales recursos; por ello se incluyeron los métodos estadístico y anatómico para tratar de lograr una determinación más sólida de los parámetros de la Ecuación de Crecimiento de Von Bertalanffy. En ese sentido, se encontró que los valores de k obtenidos entre los métodos aplicados y las tres temporadas no variaron significativamente, excepto por una ligera tendencia ascendente del primer al tercer año, y se observa consistencia con lo reportado en la literatura, donde los valores de la tasa de crecimiento fluctúan entre 0.24 y 0.28 (Torres, 1984; Torres y Chávez, 1987; Mexicano y Arreguín, 1987). Estos pequeños aumentos tienen una manifestación más evidente en el valor de la longitud asintótica correspondiente debido a la relación inversa que entre ambos parámetros han establecido Pitcher y Hart (1982), es decir, especies que tienen tasas de crecimiento altas, alcanzan rápidamente su talla asintótica que en términos absolutos, son menores que las correspondientes a especies de lento crecimiento. Los valores de k estimados en las tres temporadas permiten confirmar por otro lado, que la rubia presenta un crecimiento rápido, dado que como lo demostró Torres (1987), en el tercer grupo de edad los individuos alcanzan el 50% de su longitud asintótica, al iniciar la primera madurez sexual comprenden el 62% de la máxima talla observada, y al llevarse a cabo el reclutamiento a la pesquería ya sobrepasaron el 50% de su ciclo de vida. Debido a que no se cuenta con la información de los factores ecológicos que determinan el crecimiento de esta especie, las fuentes de error que se pueden considerar en estas condiciones para la estimación de los valores de k y Loo son: la representativad de la muestra y la subjetividad de algunos de los métodos empleados. En este sentido, la semejanza menor se observó en los valores calculados para 1984 y 1985, entre el Programa ELEFAN y el Retrocálculo, ya que el primero, al igual que el Método de Harding, se basa en el uso de frecuencias de longitud a partir de las cuales se obtienen las longitudes promedio estimadas para los grupos de edad, proporcionando además una evaluación rápida de los parámetros poblacionales que pueden ser utilizados en los modelos predictivos de rendimiento pesquero. El Retrocálculo, por otro lado, amplia el número de datos a analizar, pero tiene la limitante de que la estimación del factor de corrección está fuertemente determinada por el intervalo de tallas de los organismos de los cuales se extrajeron las estructuras óseas para la lectura de marcas de crecimiento. Una adecuada estimación de este factor ayuda en el cálculo correcto de longitudes estimadas para los primeros años de vida (Carlander, 1982). De ahí la importancia de incluir ejemplares (dentro de un amplio intervalo de tallas) pequeños para determinar la relación radio de la escama-longitud furcal del pez, y grandes para no subestimar el valor de la longitud asintótica. Dada la gran dificultad que representa la evaluación de las distintas fuentes de mortalidad natural, en general se ha considerado que ésta presenta un valor constante para todos los grupos de edad posteriores al reclutamiento, y en este caso se observó que no hubo variaciones significativas en los resultados en las tres temporadas de pesca, lo que puede atribuirse al método empleado, ya que como se utilizan parámetros de crecimiento y éstos fueron muy similares en los tres años, esto se refleja en los valores de mortalidad natural que fueron estimados. Lo que es más factible evaluar son los cambios en el modo e intensidad de operación de la flota pesquera entre varias temporadas. A finales de 1983 se verificó una disminución considerable en el esfuerzo pesquero (capacidad neta de bodega) en este caso de la flota arrastrera yucateca, debido a que los barcos con capacidad mayor de 180 toneladas, que operaban en puertos base en Progreso, fueron enviados a otras filiales de la empresa que los administraba. Para los fines de estudios pesqueros, estos cambios se expresan mediante la mortalidad por pesca, y a los cuales se atribuye la disminución en los valores de la mortalidad total de 1983 a 1985. Esta reducción en la autonomía y alcance totales de la flota, trajo como consecuencia una reducción en la tasa de explotación, obteniéndose así los bajos rendimientos que se registraron en 1984 (483 toneladas) y 1985 (66 toneladas) para esta especie, sobre todo considerando las capturas que se alcanzaron en 1981 (1620 toneladas), 1982 (1275 toneladas) y 1983 (936 toneladas). Estos cambios en la mortalidad total no mostraron efectos sobre la tasa de crecimiento individual ni los valores de la mortalidad natural, ya que la estimación de esta última involucra los parámetros de crecimiento que se conservaron más o menos constantes durante las temporadas analizadas. Sin embargo, deben tomarse con reserva estos resultados, dado que al salir la flota arrastrera, la rubia fue capturada con línea. También y por lo tanto, el poder de pesca y consecuentemente el coeficiente de capturabilidad varió no siendo posible definir de manera concreta el efecto de las variaciones en el refuerzo de pesca y por lo tanto en la mortalidad por pesca. Por otro lado, la justificación de los picos estimados de mayor reclutamiento se basa en el hecho de que los nuevos reclutas han alcanzado ya su primera madurez sexual (Torres, 1987), y tanto éstos como los adultos exhiben una conducta de agregación durante su época de reproducción, lo cual aumenta considerablemente las probabilidades de que sean capturados masivamente. Es por ello que los perfiles mensuales de captura y reclutamiento en las tres temporadas analizadas sean tan similares, pudiendo corroborarse que la estación de desove va de marzo a septiembre, con una actividad intensa y masiva entre marzo y julio, lo cual ya ha sido reportado por Claro (1986) y Torres (op. cit.). Para el caso de 1985 se presentó un perfil de capturas distinto a las dos temporadas anteriores, ya que se registraron de marzo a septiembre valores similares en la magnitud de las capturas y del patrón de reclutamiento. Esto concuerda con lo reportado por Claro (1986) en el sentido de que la época de reproducción de esta especie abarca desde marzo hasta septiembre, con picos máximos de marzo-abril y junio-julio. La relación observada entre las capturas y el reclutamiento puede deberse a que la rubia exhibe un hábito gregario durante su periodo de desove, lo cual aumenta las probabilidades de que individuos que recientemente han alcanzado su primera madurez sexual sean capturados (Torres, 1987). Finalmente, a la luz del análisis realizado, se puede concluir que es necesario llevar a cabo una serie continua y sostenida de estudios de la dinámica de las poblaciones bajo explotación, ya que la lógica misma de esta disciplina lleva implícita la noción de fluctuaciones en el tiempo, cuya dirección e intensidad estarán influidas por los niveles del uso pesquero que soporten dichas poblaciones. El conocimiento de esta dinámica proporcionará los elementos necesarios para predecir el régimen de explotación con el cual se alcanzarán los rendimientos óptimos pesqueros en subsecuentes temporadas de pesca. Por otra parte, es importante mencionar que debe probarse más de un método para estimar el crecimiento de las especies, principalmente haciendo la validación de la edad, ya que los parámetros de crecimiento son utilizados para evaluaciones del recurso, y podrían darse sobre subestimaciones, sobre todo en términos de biomasa, repercutiendo por tanto en las medidas propuestas para el aprovechamiento del recurso. LITERATURABEAMISH, R.J. Y G.A. McFARLANE The forgotten requirement for age validation in fishes biology. Trans. Amer. Fish. Soc. 1983.735-743.112 (6): BREY, T. Y D. APULY Electronic lenght frequency analysis. A revised and expanded user's guide to ELEFAN 0.1 and 2. ICLARM Contribution 251Metro ManilaFilipinas. 1986. 49 p. CARLANDER, K.D., Standard intercepts for calculating lengths from scale measurements for some contrarchid and percid fishes. Trans. Amer. Fish. Soc. 1982. 332-336. 111 (5): CASSIE, R.M. Aust. Jour. Mar Pres. Res. Some uses of probability paper in the analysis of size frequency distribution. 1954. 513-522. 5 CLARO, R. Ecología y ciclo de vida del caballerote Lutjanus griseus (L.) en la Plataforma Cubana. II. Edad y crecimiento, estructura de las poblaciones-pesquerías. Acad Cien. Cub., Rep. Inv. Inst. Ocean. 1983. No. 8. CLARO, R. En: Simposio de Ciencias del Mar y VIII Jornada Cieniffica del Instituto de Oceanografía. Ecología de los peces marinos de Cuba. Acad Cien. Cub. 1986.2. JOHNSON, A.G. Age and growth of yellowtail snapper (Ocyurus chrysurus) from South Florida Trans. Amer. Fish. Soc. USA.1983.173-177. 112 (6): MEXICANO-CINTORA, G. Y F. ARREGUIN-SANCHEZ Congreso de Ciencias del Mar.Dinámica de las poblaciones de la rubia (Lutjanus synagris y canané (Ocyrus chrysura) de las costas de Yucatán.La HabanaCuba.1987. PAULY, D. A preliminary compilation of the fish length growth parameters. Berichte Inst. Meereskd Christian Albrechts Univ. Kiel1978.200p. (55) PAULY, D. On the interrelationships between natural mortality, grawoth parameters and mean environmental temperature in 175 fish stocks. J. Cons. Cos. Int. Explor. Mer. 1980.175 -192.19 (2): PAULY, D. Y J. DAVID A basic program for the objective extraction of growth parameters from length frequency data.Meeresfarsh.1981.205-211. (28): PITCHER, T.J. Y PJ. HART Fisheries Ecology.Crom Helm Londres.1982. RICKER, W.E. Bull. Fish. Res. Board Can. Computation and interpretation of biological statistics of fish populations. 1975302 p. (191): TORRES, R. Tesis Profesional. Diagnosis de la pesquería de la rubia (Lutjanus sinagris Linnaeus 1758) en las costas de Yucatán. ENEP-Iztacala, UNAM México.1984. TORRES, R. Tesis de Maestría.Análisis biológico-pesquero de dos temporada de pesca de la rubia Lutjanus synagris L. 1758) de la costas de Yucatán. CINVESTAV-IPN-Unidad Mérida, Yucatán. 1987. TORRES, R. Y E. CHAVEZCiencias Marinas Evaluación y diagnóstico de la pesquería de la rubia (Lutjanus synagris) en el estado de Yucatán. 1987. 7-29.13 (l):
|