|
SHORT- TERM
EVALUATION OF THREE PELLETIZED DIETS FOR THE RED SWAMP CRAYFISH Procambarus
Clarkii (Girad)
Trabajo recibido el 26 de septiembre de 1989 y aceptado
para su publicación el 4 de mayo de 1990.
CIENTIFIC NOTE
Beatriz Cordero
Domenico Voltolina
Centro de Investigación
Científica y de Educación Superior de Ensenada (C.I.C.E.S.E.),
Departamento de Acuicultura, Apdo. Postal 2732, Ensenada, B.C.,
México.
Se evaluaron en el laboratorio, bajo
tres diferentes condiciones de temperatura, tres dietas para
Procambarus clarkii. Los criterios de evaluación
empleados fueron la aceptación y asimilación de las dietas, y el
consumo energético en cada una de las situaciones experimentales, ya que
estos factores determinan la energía potencial para el crecimiento del
organismo. La mayoría de las respuestas de P. clarkii
resultó depender de las dos variables: la aceptación de dos de
las dietas fue mayor a la temperatura más baja, mientras que la tercera
fue consumida preferentemente en condiciones de alta temperatura. La eficiencia
de asimilación fue alta (entre 85.8 y 92.5%) en todas las situaciones
experimentales. La relación entre consumo energético y
temperatura fue diferente para cada una de las dietas, lo cual modificó
el cuadro obtenido de la evaluación basada únicamente sobre
aceptación y asimilación de las mismas
dietas.
Three pelletized diets for Procambarus
clarkii were evaluated in the laboratory under different thermal
conditions, the criteria for evaluation being their acceptance, their
assimilation, and the amount of energy stored (scope for growth). Temperature
and type of diet influenced the responses of p. clarkii: two diets were
accepted better at the lowest temperature, while the acceptance of the third
was significantly higher at the upper limil of the experimental thermal range.
Assimilation efficiencies were high (between 85.8 lo 93.5%) in all situations.
Different trends were observed in the relationships between energy loss
(asmeasured by oxygen consumption) and temperature, for each of the diets. This
variously modified the overall picture of scope for growth, which only
partially paralleled the one described for food acceptance.
The semi-intensive culture of the red swamp crayfish Procambarus clarkii in growout ponds, as well as the production of juveniles in intensive culture units, might become in Europe, where live crayfish commands much higher prices than in the rest of the world and where this species has already been introduced (Huner, 1988), a feasible alternative to the extensive mixed culture practices, currently used (Anonymous, 1980; Huner and Barr, 1984). For this reason, studies related to feeding and to artificial diet formulation and testing (e.g. Tarshis, 1978; Huner and Meyers, 1979; Cange et al., 1982; Garcés and Avault, 1985; see also Huner and Barr, 1984 and biblio-graphy therein), should he viewed with renewed interest, especially with regard lo complete diets for juveniles. in this context, juveniles and sub-adults of one Baja California wild population, as well as offisprings of wild broodstock of the same origin, have been used to test several pelletized diets formulated with natural products available locally and with by-products of the agricultural and fishery industry (Re Araujo and Bückle Ramirez, 1985; Moralez González, 1986). In this work the two high protein diets which these authors considered more effective, are compared to a third one formulated with fishmeal and with the solid residue of aerobic degradation of cow and chicken manure, which Paniagua Michel and Granados Machuca (1987) suggested as a potential source of raw material for the elaboration of animal feeds. MATERIALS AND METHODSThe pelletized diets were prepared as in Re Araujo and Bückle Ramírez (1985), using 5% agar as binding agent. Their composition and the results of the proximate analysis of their components are given in Table 1. The experiments were run at three thermal regimes (18,22 and 26°C), near to the lower, central and upper values of the thermic range of Baja, California Procambarus clarkii (Re Araujo and Buckle Ramírez, 1985). 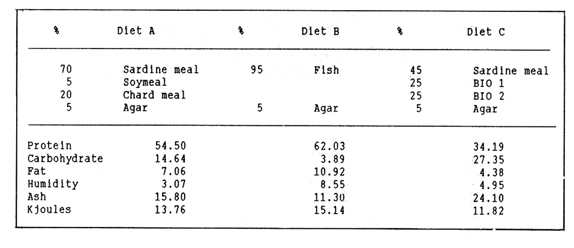 TABLE 1 RELATIVE COMPOSITION, RESULTS OF PROXIMATE ANALYSIS (IN %WEIGTH) AND ENERGY VALUES (KJOULES* 100 g) FOR THE THREE EXPERIMENTAL DIETS. FISH REFERS TO FISH DIS CARDS FROM THE LOCAL MARKET, BIO 1 AND BIO 2 REFER TO SOLID RESIDUE FROM AEROBIC DIGESTION OF COW AND CHICKEN MANURE RESPECTIVELY The organisms, varying in size between 40 and 70 mm, were collected from a small stream close to Ejido Uruapan (31°37'16" N Lat. and 116°26'36" W Long.) some 40 Km SE of Ensenada (Baja California). In the laboratory they were measured, sexed, weighed to the nearest 0.1 g and acclimatized to the experimental feeding and thermal regimes for two weeks in individual acrylic chambers. Feeding was once daily, with one ration of the appropriate diet equivalent to 5% of the initial weight. After acclimatization 17 organisms, previously starved for 48 hours, were transferred to a similar system, consisting of 21 individually acreated acrylic chambers (17 for the experimental organisms, 4 for the control of environmental changes), kept in a water bath for temperature control (Rico Mora, 1987). Each organism was fed 5% of its initial weight of the diet tested. The uneaten food and the faeces were collected separately after 6 hours by pipetting, faeces collection being repeated every two hours for the next 12 hours. Food and faeces were filtered dry ori to Whatman GF/C glass fiber filters, dried at 60°C to constant weight, and weighed to the nearest 0.01 mg. After completion of this phase of the experiment each chamber was cleaned thoroughly by pipetting, the water was completely changed and oxigen con- sumption and ammonia excretion were measured taking care, to prevent any air-water exchange with a layer of mineral oil. Incubation times were 4 to 6 hours, depending on temperature conditions. this experimental procedure was repeated with a new group of organisms, for each diet and temperature setting. At the end of each experiment the dry weight of each animal was determined, to normalize metabolic activities to 1 g of dry weight. Oxygen consumption and ammonia excretion were measurd in indiscrete samples drawn from the experimental closed system (Bayne,1985), using the appropiate electrodes of an Orion 901 specificacion analser. Oxygen samples were analyzed immediately, while ammonia was measured in samples preserved by freezing until the end of all experiments. The criteria of evaluation were the acceptance of each diet as % ingestion (I) by the organisms, the efficiency of its assimilation (AE) and the scope for, growth (P, see Winberg, 1960; Warren and Davis, 1967), which were calculated using equations 1, 2 and 3 respertively: I = [(AP - AS) / AP] × 100 (1) AE= [(F-E)/F]x100 (2) P=A-(R+U) (3) Where: A = Energy absorbed: Energy content of food ingested * AE AP= Dry weight of food ration AS = Dry weight of food uneaten E = Dry weight of faeces F =Dry weight of food ingested AP - AS R = Energy lost by respiration (20.33 J ml-1-¹ 02 consumed, See Bayne, 1985). U = Energy lost by excretion (24.87 J mg-1-¹ NH3-N excreted). Nonparametric statistics was used to compare the results, obtained, since the conditions of homogeneity of variance and goodness of fit to normal distribution were not met. Differences between physiological responses were tested using two way ANOVA and one adaptation for nonparametric statistics of the Student-Newman- Keuls multiple comparison test, elaborated by Dr. Chagoya Guzmán for C.I.C.E.S.E.'s ESIMSL statistical pachage. The results of statistical analysis are summarized throughout the text and are given in detail in Cordero Esquivel (1988). RESULTSThe mean values and standard deviations obtained for each experiment are given in Table 2. Diets A and B were well accepted at all temperatures. The only significant differences were, within diets, the slightly lower consumption, of diet A at 22°C and, between diets, the lower ingestion. of diet B at 26°C, as compared, to A, at 18°C. In the case of diet C, acceptance was as high as that of the others at 26°C, while at the two lower temperatures the percent ingestion was significantly lower than in all other experimental situilions. Two way ANOVA related these differences to the effect of temperature rather than diet (P<0.01 and >0,09 respectively), and to an interactive effect of the two (P<0.01). Assimilation efficiencies were high for all experimental combations, differences being due to temperature and, diet composition (P<0.05, in both cases), but not to the interaction of the two factors. Within diets, B was utilized sigficantly better at 18°C; no temperature-related differences were evident for A an C. Between diets B was assimilated better than A when utilizations at equal temperatures were compared. The diets assimilated with the, greates efficiency were C at 26°C and B at 18°C. Differences in oxygen consumption were shown to depend on both experimental variables and on their interaction (P<0.01 or better), probably reflecting diet-dependent differences in the thermal optimum of P. clarkii. Thus, organisms fed diet A showed oxygen consumption, progressively increasing with temperature, the respiration of those using diet B only increased between 18° and 22°C, while the use of diet C resulted in minimum respiratory activity at the intermediate temperature of 22°C. The data on ammonia excretion are not dealt with in this paper, due to their high, variability resulting from the varying sample preservation time (Cordero Esquivel, 1988 ). Nevertheless, it was decided to use their overall mean (2.45 mg NH3-N.g D.W. day -¹) in order to obtain scope for growth values closer to reality. These resulted positive for, all experimental situations (Fig. 1), with higher values at 18°C for diets A and B, and at, 26°C for diet, C. 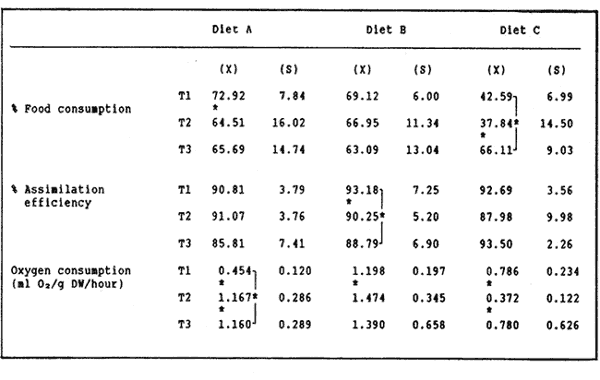 TABLE 2 MEAN (X) AND STANDARD DEVIATION (5) VALUES OF THE RESULTS OBTAINED FOR THE NINE EXPERIMENTAL SITUATIONS. DIFFERENCES (P<0.05) WITHIN DIETS ARE INDICATED BY AN ASTERISK FOR DIFFERENCES BETWEEN DIETS SEE TEXT. TI=l8°C; T2=22°C; T3=26°C DISCUSSIONThe poor acceptance of diet C at the two lower temperatures remains unexplained at this time. A temperature-dependent difference in solubility of repellant and/or attractant substances present in the diet would seem lo be most likely explanation. The data on proximate diet composition are insufficient to explain the few differences noted in assimilation efficiencies. The slightly lower overall assimilation of diet A (89.2% as compared to 90.7% and 91.4 for B and C, respectively) could be due to the presence of a higher content of undigestible plant fiber which had not been subjected, like in the case of diet C, to previous bacterial activity. The data of oxygen consumption seem high, when compared lo those available in the literature (e.g. Ansell, 1973; Chinnayya, 1974; Regnault, 1981). This is probably due to the fair degree of activity displayed by the specimens in all experimental conditions, making the data closer to measurements of active, rather than routine metabolism. Furthermore, the respiratory response to increasing temperatures is different for the three diets. Although contrary to any Q10-based expectation for diets B and C, the low values of the standard deviations calculated for each experimental situation seem to support the validity of these observations, and point to diet-related differences in the thermal optimum for this species. Re Araujo and Bückle Ramírez (1985) reported the preference of Baja CaliforniaP. clarkii for low temperatures (20°C)and its lack of tolerance for higher thermal regimes, and hypothesized a "thermal barrier" between the Lousiana and the Baja California populations. The ingredients used by these authors in the formulation of their diets were the same as in diet A, which resulted in, high respiratory rates and consequent progressively lower values of scope for growth for increasing temperatures. This might explain the high mortalities they obtained at high temperatures. The high oxygen consumptions resulting at all temperatures from the use of diet B are compensated by its higher caloric value. Higher survival was in fact obtained with a purely animal diet by Re Araujo and Bückle Ramírez (1985). Finally, diet C showed the lowest respiration at the intermediate temperature, a fact which might find an explanation in the lower values of food acceptance and assimilation in this thermal condition. 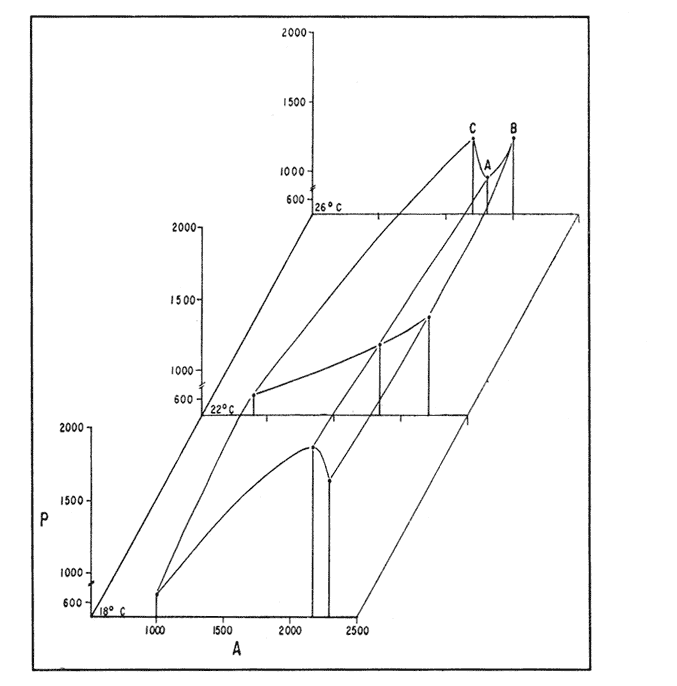 Figure 1. Scope, for growth (P) of Procambarus clarkii al three different experimental temperatures, as a function of energy absorbed through diet (A). The results obtained for scope for growth would seem to indicate a double alternative in the choice of an appropiate diet for P. clarkii: diet B might be used in preference to A and C, since the scopes for growth calculated for animals fed this diet was high in all thermal conditions. On the other hand, the high respiratory activity associated with this, and with diet A at the two higher temperatures, are indicative of an excessive energy expediture. For this reason, the use of diets A and C at the extreme temperature ranges would seem at this time the most indicated, and an improvement in the palatability of diet C might possibly result in a formulation suitable for all thermal situations throughout the season of P. clarkii somatic growth. LITERATURAANONYMOUS A review of crayfish biology, culture, and potential future of the industry in the Pacific Northwest.Univ. of Louisiana, Marine Science Center 1980. 68 p. ANSELL, A.D Neth. J. Sea Res. Changes in oxygen consumption, heart rate and ventilation accompanying starvation in the decapod crustacean Cancer pagurus.1973.455-475. 7. BAYNE, B.L. Praeger Press Biological effects of stress and pollution in marineanimals: ideas and techiniques. Nueva York. 1985. 384p. CANGE, S., M. MILTNER y J.W. AVAULT. Progr. Fish Cult.Range pellets as suplemental crayfish feed.1982.23-24.44 CHINNAYYA, B. Oxygen consuptium of Cardina waberi (Decapoda, Atyidae) in relation to temperature, starvation and size. Broteria1974.150-164.43 CORDERO ESQUIVEL, B.Tesis de Maestría Evaluación de tres dietas artificiales para Procamburus clarkii (Girard): (Decapoda-Cambaridae), en condiciones de laboratorio. C.I.C.E.S.E., Ensenada, México. 1988.83 p. GARCÉS, A.C. y J.W. AVAULT. Aquaculture Evaluation of rice (Oryza sativa), volunteer vegetation and alligatorweed (Alternanthera phyloxeroides) in various combinations as crawfish Procambarus clarkii forages. 1985. 177-186 44 HUNNER, J.V. y S.P. MEYERS.Proc. World Maric. Soc. Dietary protein requirements of the red crawfish, Procambarus clarkii (Girard) (Decapoda-Cambaridae), grown in a closed system.1979.751-760. 10 HUNER, J.V. y J.E. BARR. Sea Grant Publication Number LSU-T-80-001. Red swamp crawfish: biology and exploitation.Louisiana state University, Baton Rouge LouisianaUSA. 1984. 135 p. HUNER, J.V. Aquaculture Magazine Crayfish culture in Europe. 1988. 48-52. March- April MORALES GONZÁLEZ, J.R. Tesis ProfesionalEvaluación la eficiencia de utilización de tres dietas en el crecimiento de Procambarus clarkii (Crustacea-Decapoda) en condiciones de laboratorio Instituto Tecnológico de Veracruz.VeracruzMéxico. 1986.55p. PANIAGUA MICHEL, J. y C. GRANADOS MACHUCA. Ciencias Marinas La fracción sólida de la digestión de desechos órganicos, alimento potencial para peces y crustáceos.México1987. 69-80. (4) 13 RE ARAUJO, D. y L.F. BÜCKILE RAMÍREZ Ciencias Marinas Crecimiento y sobrevivencia de Procambarus clarkii Girard (Crustacea, Decapoda) con diferentes temperaturas y dietas isocalóricas.México1985.39-68.(2) 11 REGNAULT, M. Oceanis Adaptation du metabolisme de la crevette grise a l'environnement acoustique et a des conditions nutritionnelles défavorables.1981.675-686. 7 RICO MORA R Tesis de Maestría Efecto interactivo de la temperatura y de la concentración de microalgas en la fisiología alimenticia y la nergía potencial para el crecimiento de Modiolus capax (Conrad) (Bivalvia: Mytilidae). C.I.C.E.S.E., EnsenadaMéxico.1987. 91 p. TARSHIS, I.B. Proc. World Maric. Soc. Diets, equipment, and techniques for maintaining crawfish in the laboratory.1978. 259-269. 9 WARREN, G.E. and G.E. DAVIS. The biological basis of freshwater fish production. Blackwell Scientific Publications Laboratory studies on the feeding, bioenergetics and growth of fishes. In: S.D. Gerking (Ed.). Oxford1967.175-214. WINBERG, G.G. Fish. Res. Bd. Can. transl. Ser. Rate 7f metabolism and food requirements of fishes. 1960.253 p.194
|