|
ESTUDIO DE LAS
VARIACIONES REGIONALES EN EL CRECIMIENTO, LA LONGEVIDAD Y LA MORTALIDAD NATURAL
DE LA ANCHOVETA Engraulis mordax. Girard (Pisces: Engraulidae) EN EL NORTE DE
BAJA CALIFORNIA, DURANTE LOS AÑOS DE 1986-1987
Trabajo recibido el
28 de julio de 1989 y aceptado para su publicación el 11 de diciembre de
1989.
Manuel Gallardo-Cabello
Xavier Chiappa-Carrara
Instituto de Ciencias del Mar y
Limnología Universidad Nacional Autónoma de México.
Contribución No. 670 del Instituto de Ciencias del Mar y
Limnología. UNAM.
En el presente trabajo, se
determinan las constantes del crecimiento de la ecuación de Brody - Von
Bertalanffy para tres poblaciones de la anchoveta Engraulis mordax en el norte
de Rija California. durante los años de 1986 y 1987. Asimismo, se
determinó el crecimiento ponderal, la longevidad y la tasa de Mortalidad
natural. En todos los casos, la curva de crecimiento que mejor ajustó
los valores observados, fue la calculada aplicando las constantes de la
ecuación de crecimiento obtenidas por el método de Allen (1966).
Los valores del índice K y de la tasa de mortalidad natural, obtenidos
para las poblaciones en estudio, disminuyen conforme se incrementa la latitud,
mientras que los valores de L infinito y de la longevidad aumentan. Las
poblaciones de la anchoveta que se localizan en la zona comprendida entre las
Islas Coronado y Cabo San Quintín, presentan movimientos migratorios
norte -sur y viceversa, en función de las variaciones
atmosféricas e hidrográficas. Con respecto a los
parámetros del crecimiento, es importante señalar que los mayores
rendimientos de la flota anchovetera se obtendrían en las áreas
situadas a mayor latitud.
Brody - Von
Bertalanffy's growth constants were determined for three populations of
Engraulis mordax in west Baja California coasts, during 1986 and 1987. Weight
growth, longevity and natural rates of mortalily were also calculated. In all
cases, the growth curves which gave the better fit to observed values, were
those obtained using the growth parameters calculated by Allen's (1966) method.
the K indexes and the natural rates of mortality of these populations decrease
in northern areas, while those of ∞ and longevity increase. Northern
anchovy populalions located between Coronado Islands and San Quintin Cape show
migrations from north to south and vice versa, related with atmospheric and
hydrographic fluctuations. Greater fishery yields could be obtained in northern
areas.
De acuerdo con los trabajos realizados por Mc Hugh (1951) y Vrooman et al. (1981), se han establecido tres poblaciones de la anchoveta Engraulis mordax en las costas de California, denominadas Norteña cuya distribución abarca desde la Columbia Británica hasta la Bahía Monterrey, la Central, que comparten los Estados Unidos y México y que se localiza frente al sur de California y Norte de Baja California, y finalmente la Sureña, que se ubica frente a la parte central y sur de Baja California. No obstante, estudios más recientes (Gallardo - Cabello, 1985a, 1985b, 1985c; Parrishet al. 1985 y Chiappa-Carrara, 1988) han reportado la presencia de un número mayor de poblaciones, subpoblaciones, o razas geográficas para el área de distribución de E. mordax. tan sólo para la zona que abarca desde las islas Coronado hasta Punta Santo Tomás, se ha detectado la presencia de dos poblaciones (Gallardo-Cabello, 1985a), que quizás puedan corresponder a las denominadas Central y Sureña por Mc Hugh (1951) y Vroomanet al. (1981). Por otra parte, Parrish et al. (1985) señalan la existencia de 11 áreas en las cuales los ejemplares de E. mordax, presentan diferencias con respecto al crecimiento y a la composición por edades. Del total de las zonas mencionadas, 6 se presentan en las costas de los Estados Unidos y 5 en las de México. En el presente trabajo, se realiza un anális is sobre el crecimiento, la longevidad y la mortalidad natural de tres poblaciones de E. mordax localizadas en el área comprendida entre las Islas Coronado y Cabo San Quintín, descritas por Chiappa-Carrara (1988) y por Chiappa-Carrara y Gallardo-Cabello (en prensa). Asimismo, en este estudio se lleva a cabo una comparación tanto entre las poblaciones encontradas, como con las detectadas por Gallardo-Cabello (1985a, 1985b, 1985c) en el año de 1982, estableciéndose así posibles cambios en la distribución deE. mordax en las costas del Pacífico Norte. Por otra parte, en este estudio se han empleado diferentes métodos para la evaluación de los parámetros del crecimiento, los cuales han sido ampliamente desarrollados, con el propósito de establecer las ecuaciones que mejor se ajusten a las poblaciones descritas en el presente análisis. MATERIAL Y MÉTODOSLos datos biológicos fueron obtenidos de un total de 960 ejemplares de la anchoveta Engraulis mordax, provenientes de los muestreos realizados en la región que se localiza frente a las costas de Baja California Norte, desde las Islas Coronado hasta Cabo San Quintín (Fig.1). Las muestras son producto de la captura comercial de los buques "Calafia", "Ciprés", "Concepción", "Kino", "Libertador del Sur", "Loreto", "Sauzal" "Tortugas" y "Vizcaíno", pertenecientes a la Compañia Pesquera Zapata S.A de C.V. El intervalo de tiempo que abarca el análisis de las muestras comprende desde enero hasta octubre de 1986 y desde enero hasta junio de 1987. Durante los meses de noviembre y diciembre de 1986, las poblaciones de la especie en estudio no fueron capturadas por la flota comercial, debido a los períodos de la veda. De cada captura se separó un kilogramo elegido al azar y de los lotes se consignaron en las hojas de muestreo los siguientes datos: nombre del barco, área, fecha y volumen de la captura, número consecutivo de la muestra y peso total de la misma. De los ejemplares obtenidos se registró: la longitud total, el peso entero y el sexo. Por otra parte, se efectuó la extracción de 960 pares de otolitos sagitta. El análisis de los anillos de crecimiento permitió la identificación de ocho grupos de edad. Por otra parte, con base en las tallas promedio para cada edad, se determinaron tres poblaciones en las costas de Baja California Norte, la primera cuya distribución ocupa un área que va desde las Islas Coronado hasta Punta Salsipuedes, la segunda que abarca desde la localidad anterior hasta Punta Santo Tomás y la tercera, que va desde la localidad antes mencionada hasta Cabo San Quintín (Chiappa - Carrara, 1988; Chiappa - Carrara y Gallardo - Cabello, en prensa). En el análisis del crecimiento, se aplicó la ecuación de Brody (1927) y Von Bertalanffy (1938). Para ello, se calcularon las constantes de dicha ecuación por medio de los métodos propuestos por Ford (1933) y Walford (1946), Beverton (1954), Tomlinson y Abramson (1961), Gulland (1964) y Allen (1966). Para la obtención de la edad límite o longevidad, así como las tasas de mortalidad natural (M) se emplearon los métodos de Taylor (1958a; 1958b; 1959; 1960; 1962). RESULTADOS Y DISCUSIÓN
APLICACIÓN DE LA ECUACIÓN DE BRODY - VON BERTALANFFY
Método de Ford - WalfordEmpleando los datos obtenidos a partir de las lecturas de los otolitos, se calcularon los siguientes valores de Lt y de Lt+1 para cada zona de estudio. En 1986: 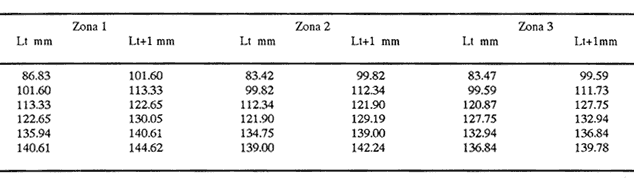 En 1987: 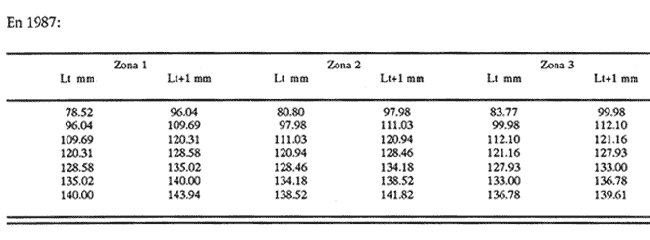 Los valores de los parámetros K y L∞ estimados por este método aparecen a continuación: En 1986:  En 1987: 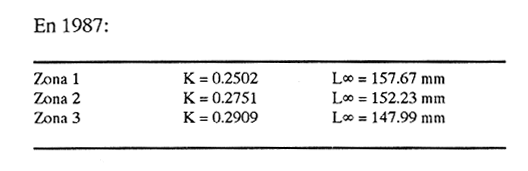 Método de GullandLos valores de to calculados por este método, son los iguientes:  Método de Tomlinson y AbramsonLos resultados obtenidos, para z, son:  Asimismo, los valores calculados para los parámetros de la ecuación de Brody - Von Bertalanffy se presentan a continuación: 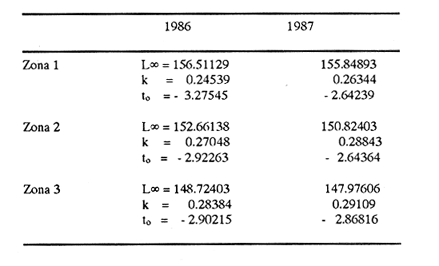 Método de AllenLos parámetros del crecimiento calculados por este método, son:  Método de BevertonPara la aplicación de este modelo, se utilizaron los valores de L infinito obtenidos mediante las técnicas de Ford Walford, Tomlinson - Abramson y Allen. A continuación se presentan los resultados obtenidos: 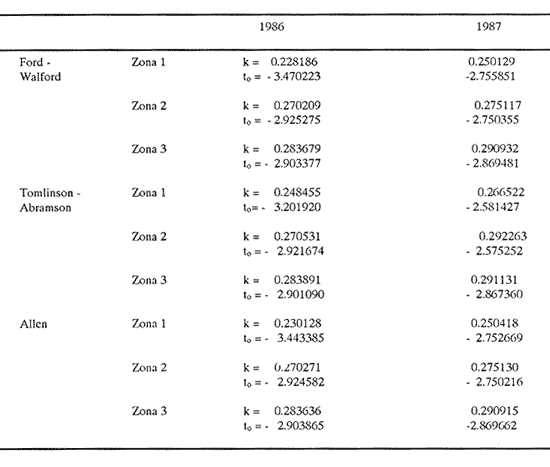 Selección y valoración de los coeficientes de crecimiento. A partir de las estimaciones realizadas para cada coeficiente se han desarrollado las curvas del crecimiento de la anchoveta y obtenido las diferencias de los valores calculados de las tallas para cada edad, con respecto a los observados (Tabla 1). El método de Allen (1966) fue el que mejor ajustó los datos, en la mayoría de los casos, proporcionando las sumas de las diferencias al cuadrado de menor valor, con excepción de la zona 3 en el año de 1986, en la que se obtuvieron valores iguales a los obtenidos mediante la aplicación de las técnicas de Ford - Walford y Tomlinson y Abramson. El método de Ford - Walford, en la mayoría de los casos no presentó un buen ajuste con respecto a la técnica de Allen. Lo anterior puede deberse a que en el desarrollo de la curva de Brody - Von Bertalanffy se empleó el valor de to obtenido por el método de Gulland (1964). Los valores calculados por medio de la técnica de Tomlinson y Abramson (1961), fueron los que presentaron una mayor diferencia con respecto a los observados, con excepción de la zona 3, durante el año de 1986. Lo anterior se debe a que la metodología propuesta por estos autores, requiere que el tamaño de las muestras para cada grupo de edad, sea igual, permitiendo que las varianzas tiendan a ser homoscedásticas, como lo requiere la aproximación de Stevens (1951). Al no cumplirse estos supuestos en la aplicación de dicho método, se obtienen resultados menos precisos, como ocurrió en el presente trabajo. Sin embargo, en otros estudios de poblaciones de E. mordax en que se analizaron muestras de un mayor tamaño que las estudiadas en este trabajo, el método de Tomlinson y Abramson (1961) fue el que permitió un mejor ajuste entre la curva calculada y los datos observados (Gallardo - Cabello, 1985b). Por lo tanto, se puede decir, en términos generales, que en el presente análisis, la técnica de Allen (1966) fue la que presentó un mejor ajuste entre los datos observados con respecto a los calculados debido a que este método no requiere que las muestras estén dispuestas en intervalos regulares de tiempo, ni que sean del mismo tamaño. Así, las varianzas de los valores calculados no tienen que cumplir con las propiedades de homoscedasticidad citadas por Pimentel -Gomes (1953). De acuerdo con lo anterior, puede proponerse la aplicación de la técnica de Tomlinson y Abramson (1961) en el estudio de crecimiento de poblaciones deE. mordax cuando las muestras obtenidas para el análisis de este parámetro sean iguales para cada grupo de edad y de gran tamaño (Gallardo -Cabello, 1985b), mientras que el método de Allen (1966) resultará de mayor utilidad en la mayoría de los casos, para la obtención de las constantes de la ecuación de Brody - Von Bertalanffy, cuando se trabaje con muestras desiguales y de menor tamaño a las estudiadas por Gallardo - Cabello (1985b). La aplicación de la ecuación de Beverton (1954) a los métodos anteriormente citados, no proporcionó, en la mayoría de los casos, un mejor ajuste, debido a que la obtención de constantes más reales puede depender de que tan preciso sea el valor asumido para L infinito. Ecuaciones del crecimiento de Engraulis mordax. Crecimiento en longitudLas curvas calculadas que mejor describen los valores de la longitud observados, son las representadas por las siguientes ecuaciones:  Los valores de las constantes de las ecuaciones, son los calculados por el método de Allen (1966). En la tabla 2, se presentan las longitudes teóricas para cada edad. En las figuras 2 y 3 se muestran las curvas teóricas del crecimiento en longitud de E. mordax para las zonas 1, 2 y 3, correspondientes a las edades de "O" a "VII" años. En las poblaciones analizadas, se observa que E. mordaxpresenta un rápido crecimiento en el transcurso de los primeros meses de vida. Datos similares han sido reportados por Gallardo - Cabello (1985b), para las poblaciones de la anchoveta estudiadas en el año de 1982. Durante el primer año (considerando las tres zonas de estudio), la anchoveta alcanza longitudes comprendidas entre 99.58 y 101.60 mm de longitud estandard (para el año de 1986) y de 96.04 y 99.98 mm de LS (para el año de 1987), lo que equivale al 64% - 67% y al 61% - 68% de L infinito respectivamente. En todos los casos, a partir del primer año de vida, la velocidad del crecimiento disminuye considerablemente (Figuras 4 y 5). Crecimiento en Peso Relación talla - PesoLas ecuaciones que describen las relaciones entre la longitud y el peso, se presentan a continuación. 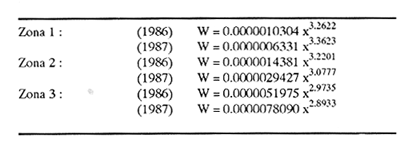 Peso a cada edadDe acuerdo con la relación entre la talla y el peso, se obtienen los pesos a cada edad, a partir de los datos del crecimiento en longitud. En la tabla 3, se muestra el crecimiento entre el peso absoluto y los valores del peso relativo (Expresado como Loge Gi + 1 - Loge Gi) para cada una de las zonas de estudio. De acuerdo con los resultados obtenidos, puede apreciarse que el incremento instantáneo en peso disminuye conforme aumenta la edad. Crecimiento teórico en PesoA continuación se presentan las ecuaciones que describen el crecimiento en peso.  Empleando dichas ecuaciones, se han calculado los pesos teóricos de E. mordax a diferentes edades, que aparecen en la tabla 4. En las figuras 6 y 7 se muestran las curvas teóricas del crecimiento en peso para las tres zonas de estudio, correspondientes a las edades "0" a "VII" años. Edad Límite o LongevidadLas longevidades encontradas para la anchoveta Engraulis mordax para cada una de las zonas de estudio (Taylor 1958a; 1958b; 1959; 1960; 1962), aparecen a continuación. 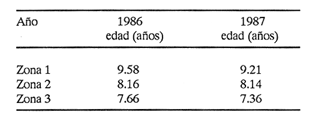 Mortalidad naturalLa tasa instantánea de la mortalidad natural (Taylor 1958a; 1958b; 1959; 1960; 1962) obtenida para E. mordax en cada una de las zonas analizadas, es la siguiente.  Variaciones de los parámetros del crecimiento y de la mortalidad natural en relación a la latitud. Con el objeto de estudiar las características del crecimiento de E. mordax y de analizar algunas de las relaciones con otras poblaciones de esta misma especie, en la tabla 5 se presentan las longitudes promedio para diferentes edades de la anchoveta, en varias localidades del Pacífico oriental. Las zonas que han sido consideradas para el presente análisis se muestran en la figura 8. De acuerdo con lo anterior y considerando los datos proporcionados por Clark y Phillips (1952), Spratt (1975) y Sunada (1975), se observa que la anchoveta del área central de California, alcanza longitudes mayores a una misma edad a partir del tercer año, que la anchoveta del área del sur de California, diferencia que se incrementa conforme aumenta la edad. Por otra parte, Parrish et al. (1985) muestran que la anchoveta E. mordaxpresenta longitudes mayores a una misma edad, en las localidades del norte con respecto a las del sur, disminuyendo el crecimiento conforme decrece la latitud. Así, los ejemplares capturados en las áreas de California Central, San Nicolás y los bancos de tanner y Cortez, alcanzan longitudes comprendidas entre los 116 y los 120 mm al cumplir su primer año de vida, mientras que los individuos pertenecientes a las localidades del área de Catalina y del canal Santa Bárbara, alcanzan longitudes de 119 mm durante su segundo año de edad. De la misma manera, se observa que las anchovetas obtenidas en las zonas del canal de San Pedro, Coronado y Ensenada, alcanzan longitudes comprendidas entre los 118 y 121 mm al cumplir su tercer año de vida y, similarmente, los ejemplares capturados en las áreas cercanas al Cabo San Quintín y la Bahía de Vizcaíno (Baja California Sur), presentan longitudes de 122.5 y 121 mm al alcanzar el cuarto y el quinto año de edad, respectivamente. Para los individuos pertenecientes a áreas situadas más al sur de las antes mencionadas, Parrish et al.(1985) establecen una longitud asintótica de 92 mm, misma que estos organismos alcanzan durante el primer año y medio de vida. Asimismo, los autores mencionados, reportan que la anchoveta E. mordaxalcanza tallas mayores a una misma edad, en áreas alejadas de la costa con respecto a aquellas cercanas a la misma, como es el caso de las localidades de Coronado, Catalina y los bancos de tanner y Cortez, en que los ejemplares alcanzan, durante su primer año de edad, longitudes de 107, 112 y 116 mm, respectivamente. En relación a los organismos de E. mordax estudiados por Gallardo - Cabello (1985b) en Baja California norte (áreas de Coronado y Ensenada), éstos alcanzan longitudes de 120.12 y 116.63 mm durante el cuarto año de vida, respectivamente; valores semejantes a los obtenidos por Parrish et al. (1985) para las zonas de Cabo San Quintín y Bahía Vizcaíno. Las diferencias en las longitudes alcanzadas por la anchoveta E. mordax, para una misma edad (y por lo tanto, en los parámetros del crecimiento y mortalidad natural), en las distintas localidades analizadas, pueden deberse, entre otras, a las diferentes épocas del desove, factores genéticos, diferencias en la alimentación o bien, a las migraciones de esta especie (Collins, 1971; Mais, 1974; Mallicoate y Parrish, 1981; Gallardo-Cabello, 1985b y Chiappa-Carrara, 1988). Ahora bien, las poblaciones de la anchoveta estudiadas por Gallardo - Cabello (1985b) para el año de 1982, no son las mismas que las analizadas en el presente trabajo para los años de 1986 y 1987. De acuerdo con los resultados obtenidos, podría establecerse que las poblaciones de la anchoveta estudiadas en el primer período, se encontraban durante los años de 1986 y 1987, distribuídas en una zona más al sur del área de estudio, es decir, entre la Bahía Vizcaíno y el Cabo San Lucas. Es muy probable que durante 1982 haya existido un desplazamiento hacia el norte en busca de mejores condiciones para su desarrollo. Se trata de anchovetas que alcanzan una longevidad promedio de cuatro años y que de acuerdo con otros parámetros poblacionales representan a las poblaciones que comunmente se localizan en las regiones sureñas. En la tabla 5, puede observarse la gran similitud que existe entre la talla promedio para cada edad de los ejemplares de E. mordax estudiados por Gallardo Cabello (1985b) para el año de 1982 con respecto a los estudiados por Parrish et al. (1985) para el área de Bahía Vizcaíno (Baja California Sur) durante el periodo comprendido entre los años de 1966 a 1983. Las diferencias que se observan entre las tallas promedio a una misma edad entre estas dos series de datos, son debidas a que Parrish et al. (1985) consideran un mayor período para la formación del primer anillo de crecimiento en los otolitos, que el reportado por Gallardo - Cabello (1985a). Por otra parte, las poblaciones estudiadas durante los años de 1986 y 1987, representan anchovetas de una mayor longevidad que alcanzan hasta siete años de edad, casi el doble con respecto a los ejemplares analizados durante el período de 1982. Estas anchovetas estudiadas más recientemente, se encuentran más relacionadas con las poblaciones norteñas de los Estados Unidos, como son las analizadas por Spratt (1975) y Sunada (1975) en California sur y que presentan una longevidad de seis años. O bien, las poblaciones estudiadas por Parrish et al. (1985) para las áreas del sur de California y norte de Baja California, ejemplares de E. mordax que alcanzan los siete años de vida. Asimismo, es importante señalar que las tallas promedio para cada edad de las anchovetas en las áreas de Coronado, Ensenada y Cabo San Quintín analizadas por Parrish et al. (1985) son aproximadamente similares a las encontradas en el presente trabajo, para los grupos de edad de "I" a "V" años. Las mayores diferencias se observan en relación a los grupos de edad "VI" y "VII". Ello es debido a que Parrish et al. (1985) ajustaron los datos de las tallas promedio observados, por medio de una regresión lineal, con lo cual tanto la longitud como la edad tenderán al infinito. Asimismo, los datos presentados por los autores mencionados representan las longitudes para cada edad de la anchoveta resultantes de promediar los datos de 17 años de muestreos, con lo cual no se pueden observar las posibles diferencias en el número de los grupos de edad anualmente con respecto a las áreas de estudio, y comparar tanto con los datos obtenidos por Gallardo - Cabello (1985b) como con los presentados en este trabajo. Todo lo anterior permite suponer que la anchoveta E. mordaxque se distribuye en el área comprendida entre la Columbia Británica en Canadá y Cabo San Lucas en México, comprende un gran número de poblaciones o subpoblaciones, que se encuentran en constante movimiento, ya sea desplazándose a zonas más al norte o bien a áreas más al sur, en función de las variaciones atmosféricas e hidrográficas, y que dichas poblaciones presentan importantes mezclas. Por otra parte, experimentos de marcado han demostrado que las anchovetas emigran, tanto del sur de California al centro de California y viceversa, como del sur de California al norte de Baja California. Los movimientos hacia el norte ocurren durante el verano y los desplazamientos hacia el sur durante el invierno, principalmente (Haugen et al, 1969). De acuerdo con lo anterior, puede establecerse que las regiones del norte de Baja California están constituidas tanto, por ejemplares de la anchoveta del stock central como del sureño (De acuerdo con la denomincación propuesta por Vrooman et al. 1981) como ya ha sido observado por Parrish et al. (1985).- Asimismo, los autores antes mencionados, establecen que las poblaciones del sur de Baja California están básicamente compuestas por el stock sureño. Sin embargo, se han identificado anchovetas del stock central, durante el mes de noviembre de algunos años. De la misma manera, consideran que el área de California Central es una zona de mezcla entre los stocks del norte y el central. De acuerdo con lo anterior, puede considerarse que las anchovetas estudiadas durante el año de 1982, representan a las poblaciones situadas más al sur durante los años más "benignos" y que durante el año mencionado, en que se presentaron importantes variaciones en la temperatura y la salinidad de las aguas, emigraron a zonas más al norte en búsqueda de alimento. Estas variaciones en los parámetros físico-químicos de las masas de agua, asociados a cambios meteorológicos, redujeron notablemente los montos de las capturas de la flota anchovetera. Por el contrario, en septiembre de 1983 cuando bajó la temperatura de las aguas, las poblaciones que se habían desplazado hacía el norte, emigraron hacía el sur (Ornelas, comunicación personal). Este tipo de fenómenos, que se encuentran relacionados con los años "Niño" y las alteraciones hidrográficas tanto en las aguas de California en particular, como en las costas del Pacífico norte oriental en general, determinan la distribución y la abundancia de las poblaciones de la anchoveta Engraulis mordax. Por otra parte, en la tabla 6, se muestran los valores de los parámetros del crecimiento de la ecuación de Brody-Von Bertalanffy, de la longevidad y de la mortalidad natural para las poblaciones de la anchoveta en tres localidades del norte de Baja California. De acuerdo con lo anterior, se observa que cuanto más elevado es el valor del coeficiente k, menor es el valor de ∞ y por tanto, más rápido será alcanzado éste por los individuos de la población en cuestión. Así, las poblaciones presentan -conforme se sitúan a menor latitud-una menor longevidad y una mayor tasa de mortalidad natural. Asimismo, considerando que las poblaciones de Baja California norte habitan en aguas más frías, conforme se encuentran a mayor latitud (Chiappa-Carrara, 1988), se puede añadir que existe una tendencia en la que se observa que la temperatura y los valores de k y M guardan una relación directa entre sí, mientras que la temperatura y la edad límite, muestran una relación inversa. Las diferencias del valor k entre las distintas poblaciones están relacionadas con las condiciones ambientales de un área determinada, como son la temperatura y la disponibilidad del alimento, así como la intensidad de la competencia, entre otros. Por otra parte, es importante señalar que las tasas de crecimiento de una especie, pueden presentar importantes variaciones a causa de la explotación pesquera, misma que puede provocar una disminución en la talla comercial, y por lo tanto, variaciones en el coeficiente k (Gallardo- Cabello, 1985b). De acuerdo con lo anteriormente expuesto, es muy posible que los rendimientos en las capturas de la flota anchovetera sean mayores conforme se incrementa la latitud, en relación a las poblaciones que se sitúan a una latitud menor, debido a un aumento en la biomasa de la población, dado por el valor del índice k, el cual representa longitudes y pesos mayores a una misma edad. Asimismo, es importante señalar que los rendimientos de la flota comercial dependerán de los movimientos migratorios realizados por esta especie, siendo éstos mayores cuando, al descender la temperatura, las poblaciones localizadas a mayor latitud emigren hacia el sur, y menores cuando al aumentar los valores de la temperatura, los cardúmenes se desplazen a una mayor latitud, como ocurrió con la pesca de la anchoveta durante el año de 1982, con respecto a los años de 1986 y 1987. ConclusionesLa curva teórica de crecimiento que se asemeja más a los valores observados por medio de la lectura de los otolitos, es la calculada aplicando las constantes de la ecuación obtenidas por el método de Allen (1966). Las ecuaciones del crecimiento de E. mordax son las siguientes:  Los parámetros de la longividad (A0.95) y de la mortalidad natural (M), son los siguientes: 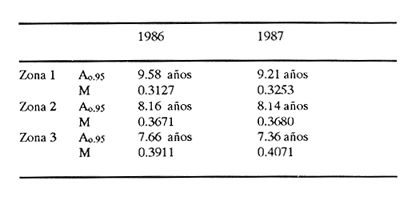 Existen diferencias latitudinales en el crecimiento de la anchoveta. Los valores del índice k y de la tasa de mortalidad natural son mayores en el sur, tendiendo a disminuir conforme se incrementa la latitud. El comportamiento de la longitud máxima teórica (L∞) y de la longevidad, es inverso. Los valores obtenidos para el índice catabólico k y la tasa de mortalidad natural guardan una relación inversa con la latitud, y una relación directa con los valores de L∞ y de la longevidad. Los más altos rendimientos de la flota anchovetera se obtendrían en las áreas situadas a mayor latitud con respecto a las zonas localizadas a menor latitud. Las poblaciones de la anchoveta E. mordax que se distribuyen en el área comprendida entre las Islas Coronado y Cabo San Quintín, en el norte de Baja California, se encuentran en constante movimiento, ya sea desplazándose a zonas más al norte o bien a áreas más al sur, en función de las variaciones atmosféricas e hidrográficas. LITERATURAALLEN, K. R. J. Fish. Res. Bd. A method lo fitting growth curves of the Von Bertalanffy type to observed data. Canada1966.163-277.(3): 23 BEVERTON, R. J. H. Us Fish & Wildl. Serv., Fish. Lab., Beaufort Misc. Contrib. Notes on the use of theoretical models ir. the study of dynamics; of exploited fish populations. 1954. 181 p. 2: BRODY, S. Growth Rates. Univ. Missouri Agric. Exp. Sta. Bull.1927.123p.97: CHIAPPA-CARRARA, X.Tesis Consideraciones bioecológicas sobre la alimentación, crecimiento, hábitos alimenticios y contenido calórico de la dieta de las poblaciones de la anchoveta Engraulis mordax Girard, localizadas en las costas occidentales de Baja California.UACPyP-CCH-UNAM1988. 131 p. CHIAPPA-CARRARA, X. y M. GALLARDO-CABELLO Biól. trop. Análisis de las frecuencias de tallas y de los otolitos sagitta para la determinación de la edad de tres poblaciones de la anchoveta Engraulis mordax Girard, en Baja California Norte (Pisces: Engraulidae),en prensa.. CLARK, F. N., y J. B. PIIILLIPS Calif Fish. Game The northem anchovy (Engraulis mordax) in the California fishery.1952.189-207.38: COLLINS, R. A. Calif Dept. Fish and Game, Fish. Bull. Size and age composition of northern anchovies, Engraulis mordax, in the California reduction and canning fisheries 1968- 1969 season.1971. 283-289. (4):57 FORD, E. J. Mar. Biol. Ass. Uk, Ns An account of the herring investigations conducted al Plymouth during the years from 1924 to 1933.1933.305-384. 19: GALLARDO-CABELLO, M. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoDeterminación de la edad de la anchoveta Engraulis mordax Girard, en aguas de Baja California Norte (Pisces: Engraulidae). 1985a. 221-234. (1):12 GALLARDO-CABELLO, M. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoAnálisis del crecimiento de la anchoveta Engraulis mordax Girard, en aguas de Baja California Norte (Pisces: Engraulidae).1985b. 235 - 252. (1):12 GALLARDO-CABELLO, M.An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México Determinación de la mortalidad total y natural de la anchoveta Engraulis mordax Girard, en aguas de Baja California Norte (Pisces: Engraulidae). 1985c. 253-260.(1): 12 GULLAND, J. A. Fish. Tech. PaperManual of methods for fish stock assessment. Part I: Fish poputation analysis. FAO1964. rev. 1. 40 HAUGEN, C. W., J. D. MESSERSMITH, y R. H. WICKWIRE Calif Dept. Fish. Game Fish. Bull.Progress report on anchovy tagging off California and Baja California, March 1966 through May 1969. 1969.75-89.147: MAIS, K. F. Pelagic fish surveys in the California Current. Calif. Dept. Fish and Game, Fish. Bull.1974.1-79.162: MALLICOATE, D. L., y R. H. PARRISH Calif. Coop. Oceanic Fish. Invest. Rep. Seasonal growth patterns of California stocks of northern anchovy, Engraulis mordax, Pacific mackerel, Scomber japonicus, and jack mackerel, Trachurus symmetricus.1981.69-81.22: Mc HUGH, J. L. Bull. Scripps Inst. Oceanogr. Meristic variations and populations of northern Anchovy (Engraulis mordax mordax).1951.123-160.6: PARRISH, R. H., D. L. MALLICOTE, y K. F. MAIS Engraulis mordax. Fish. Bull.Regional variations in the growth and age composition of northern anchovy 1985.483-496. 83: PIMENTEL-GÓMEZ, F.,Biometrics The use of Mitscherlich's regression law in the analysis of experiments with fertilizers. 1953.498-516.(4): 9 SPRATT, J. D., Calif. Fish. Game Growth rate of the northern anchovy, Engraulis mordax, in southern California waters, calculated from otoliths.1975.116-125.61: STEVENS, W. L.Biometrics Asymptotic regression. 1951.247-267.(3): 7 SUNADA, J. S. Calif. Dept. Fish and Game, Fish. Bull.Age and length composition of northern anchovies, Engraulis mordax, in the 1972-1973 season California anchovy reduction fishery. 1975133-143.(3): 61 TAYLOR, C. C. Spe. Publ., ICNAF A note on Lee's phenomenon in Georges Handdock Bank.1958a.243-251. 1: TAYLOR, C. C. J. Conseil Cod growth and temperature.1958b.366-370.(3): 23 TAYLOR, C. C. J. Conseil1959.93-101.(1): 25 TAYLOR, C C. J. Conseil Temperature, growth and mortality - the Pacific cockle1960.177-224.(1): 26 TAYLOR, C- C. J. Conseil Growth equations with metabolic parameters. 1962. 270.286.(3): 27 TOMLINSON, P. K. y N. J. ABRAMSON Fish. Bull. Fitting a Von Bertalanffy growth curve by least squares. 1961.69-101.(1): 116 Von BERTALANFFY, L. Human Biology A quantitative theory of organic growth (Inquiries on growth laws. II). 1938. 181-213.(2): 10 VROOMAN, A. M., P. A. PALOMA, y J. R. ZWEIFEL Calif. Fish. Game Electrophoretic, morphometric, and meristic studies of subpopulations of northern anchovy, Engraulis mordax 1981. 39-51. 67: WALFORD, L A. Biol. Bull. A new graphic method of describing the growth of animals.1946. 141 147. (2): 90 LAMINAS
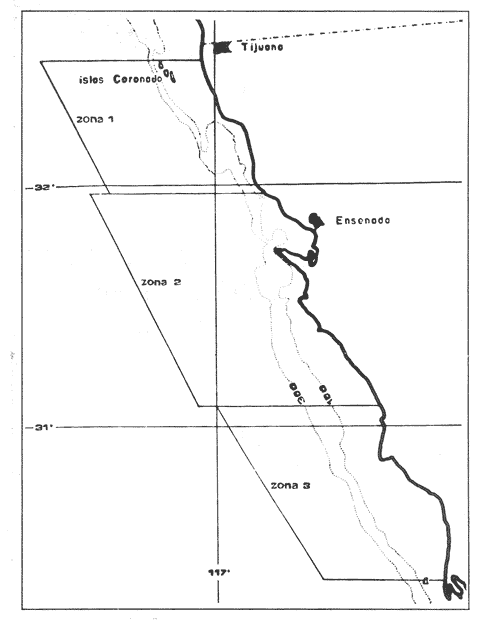 Figura 1. Localización del área de estudio. 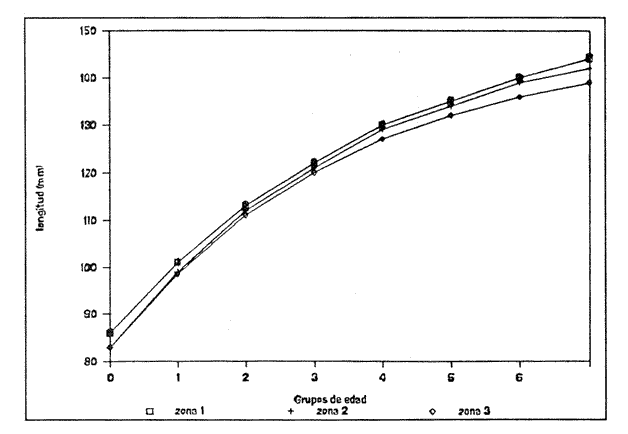 Figura 2. Crecimiento teórico en longitud, zonas 1, 2, y 3, 1986. 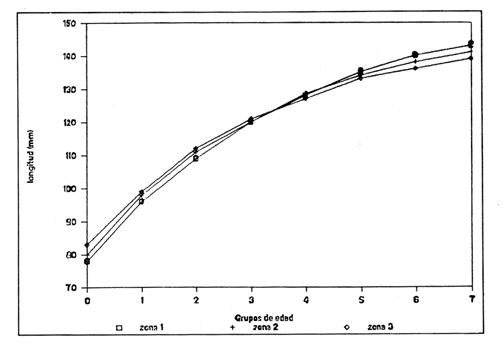 Figura 3. Crecimiento teórico en longitud, zonas 1, 2, y 3, 1987. 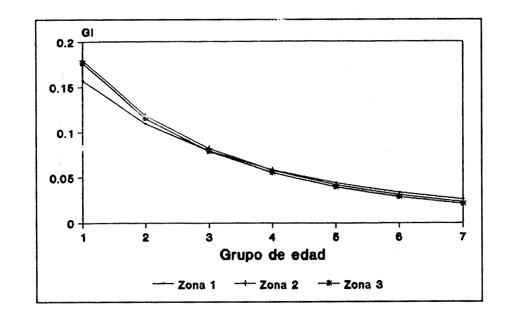 Figura 4. tasa instantánea del incremento en longitud, zonas 1, 2, y 3, año de 1986. 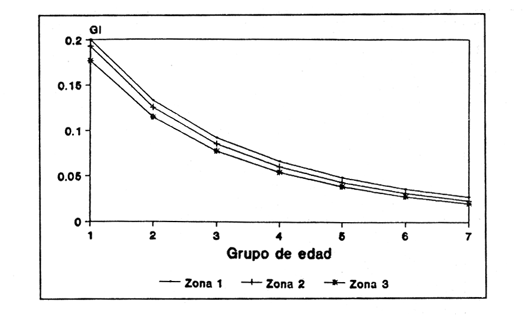 Figura 5.tasa instantánea del incremento en longitud, zonas 1, 2, y 3, año de 1987. 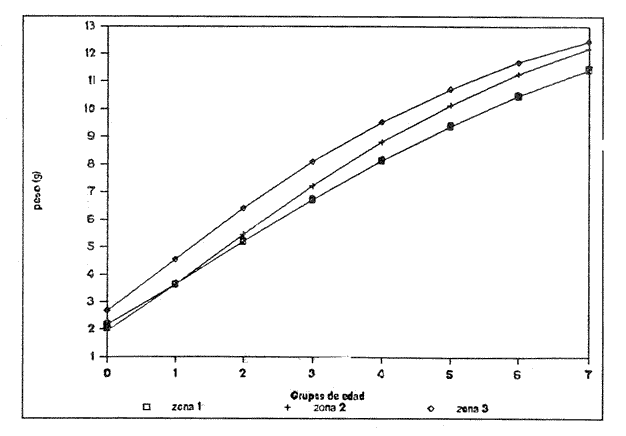 Figura 6. Curva teórica del crecimiento en peso- zonas 1, 2. y3. año de 1986. 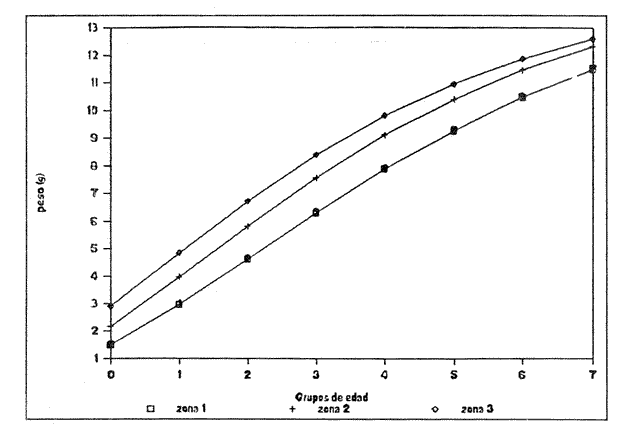 Figura 7. Curva teórica M crecimiento en peso. zonas 1, 2, y3. año de 1987. 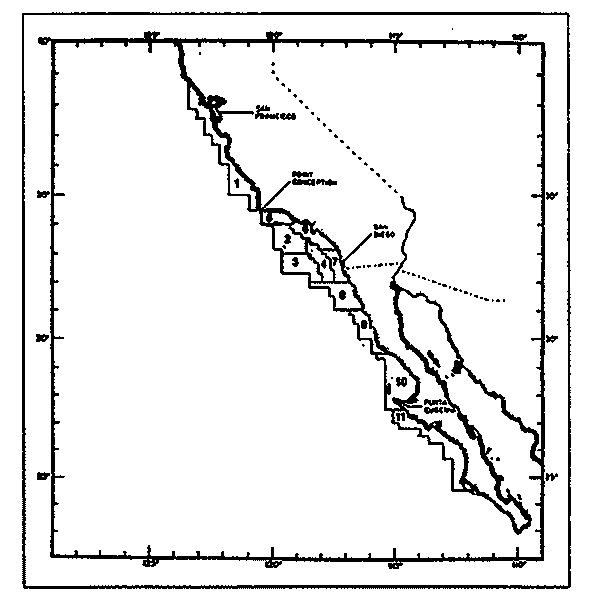 Figura 8. Localización de diversas áreas de distribución de la anchoveta Engraulis mordax, en relación a las características de su crecimiento (tomado de Parrish, et al., 1985). 1= California Central. 2= San Nicolás, 3= Bancos de tanner, Cortéz, 4= Área de Catalina, 5= Canal de Santa Bárbara 6= Canal de San Pedro, 7= Zona de Coronado, 8= Ensenada, 9= Cabo San Quintín, 10= Baja California Sur (Bahía Vizcaíno), 11 = California Sur. 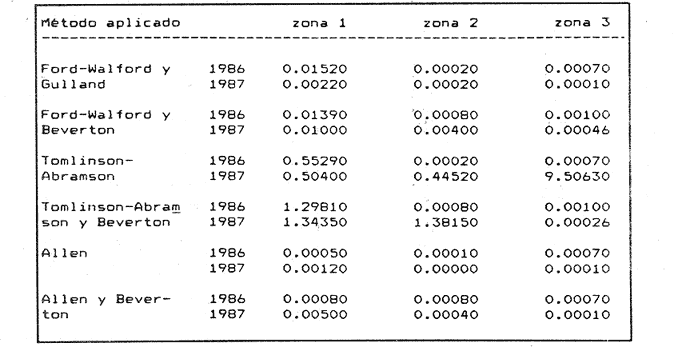 TABLA 1 SUMAS DE LAS DIFERENCIAS AL CUADRADO ENTRE LOS VALORES DE LA LONGITUD OBSERVADOS Y LOS CALCULADOS CON LOS PARÁMETROS OBTENIDOS CON DIFERENTES MÉTODOS 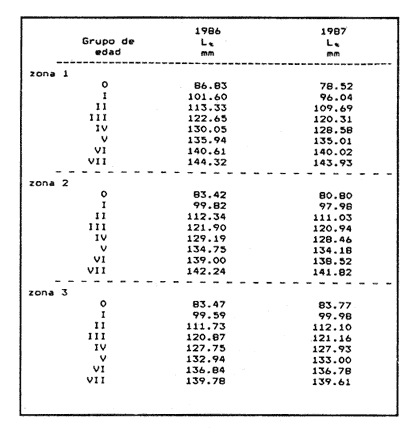 TABLA 2 LONGITUDES TEÓRICAS DE LA ANCHOVETA 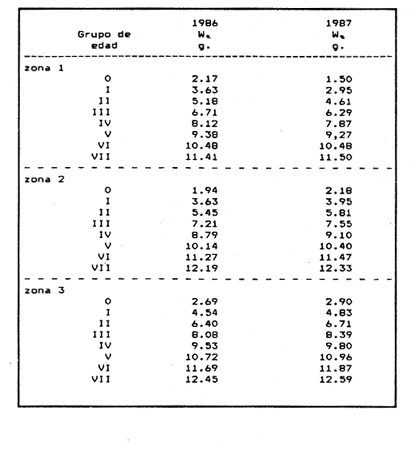 TABLA 3 CRECIMIENTO EN PESO ABSOLUTO Y RELATIVO DE LA ANCHOVETA F.. morda.v. ZONAS 1, 2 Y 3. AÑOS 1986 Y 1987 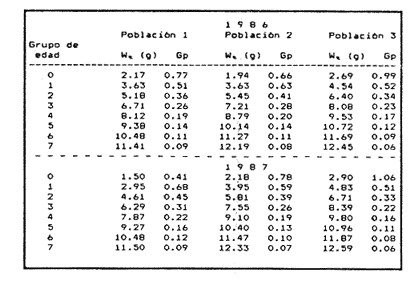 TABLA 4 PESOS TEÓRICOS A CADA EDAD DE LA ANCHOVETA E. mordax. ZONAS 1, 2 Y 3. AÑOS 1986 y 1987  TABLA 5 LONGITUDES PROMEDIO PARA CADA EDAD DE LA ANCHOVETA E. morda.r. EN DISTINTAS LOCALIDADES DEL PACIFICO ORIENTAL 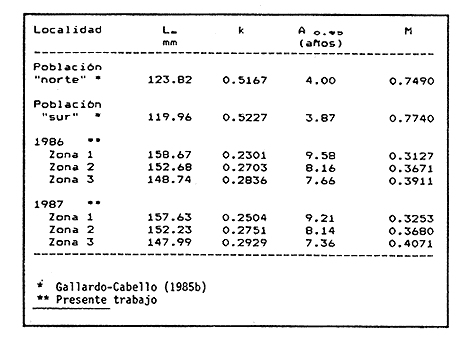 TABLA 6 PARÁMETROS DEL CRECIMIENTO, LONGEVIDAD Y MORTALIDAD NATURAL, PARA LAS POBLACIONES DE LA ANCHOVFTA F. Otorda y EN DISTINTAS ÁREAS DE BAJA CALIFORNIA NORTE
|