|
ALIMENTACIÓN DE LAS LANGOSTAS
Panulirus guttatus y P. argus (Latreille 1804) EN EL CARIBE MEXICANO
Trabajo recibido el 8 de enero 1990 y aceptado para su publicación
el 4 de mayo 1990.
Félix Colinas-Sánchez
Fuente de Marcela 28, Lomas de
Tecamachalco, 53950 México, D. F.
Patricia Briones-Fourzán
Estación "Puerto Morelos", Instituto de Ciencias del Mar y
Limnología, UNAM. Ap. Postal 1152, Cancún, Q. R., 77500
México. Contribución No. 675 del Instituto de Ciencias del Mar y
Limnología, UNAM.
En este trabajo se estudian los patrones de alimentación de las
langostasPanulirus guttatus y P. argus
(Latreille 1804) en el área de Puerto Morelos, Q.R., en el litoral del
Caribe mexicano, así como las diferencias en el tipo de
alimentación entre los sexos de ambas especies y, en el caso de
P. guttatus, su variación dietética
estacional. Para esto, se llevó a cabo el análisis del contenido
estomacal de 226 ejemplares de P. guttatus y 90 de
P. argus, capturados entre noviembre de 1986 y noviembre de
1987, con un porcentaje de vacuidad de 53.1 y 72.2 respectivamente. No hubo
diferencias significativas en el patrón alimenticio de ambos sexos en
ninguna de las dos especies. Se identificaron diez grupos tróficos para
P. guttatus y nueve para P. argus. P.
guttatusmostró una marcada preferencia por los
crustáceos. Los moluscos y las algas coralinas ocuparon un lugar
secundario. Otros grupos, tales como materia vegetal, ascidias, equinodermos y
esponjas tienen un carácter de alimento ocasional. En esta especie se
encontraron indicios de canibalismo circunstancial. Los patrones
dietéticos en cada estación del año no muestran grandes
diferencias, excepto en el invierno, que alcanzó el índice de
similitud más bajo en comparación con las demás
estaciones. P. argusmuestra preferencia por los
crustáceos y moluscos siendo los demás grupos alimentos
ocasionales. Se hace evidente el carácter generalista, oportunista,
plástico y selectivo en el comportamiento alimenticio de estas
especies.
The feeding patterns of the spiny lobsters
Panulirus gutatus and P. argus(Latreille
1804) in the area of Puerto Morelos, Q.R., in the Mexican Caribbean, were
studied, as well as the differences in food between sexes of both species and,
for P. guttatus, the seasonal variation in diet. This was
carried out through the stomach content analysis of 226 P.
guttatus and 90 P. argus, caught between November
1986 and November 1987. The vacuity percentage was 53.1 and 72.2 respectively.
No significant differences were found in the food of both sexes in either
species. Ten trophic groups were identified for P.
guttatusand nine for P. argus. P.
guttatus showed a marked preference for crustaceans. Mollusks and
coralline algae occupied a secondary place. Other groups, such as plant matter,
ascidians, equinoderms and sponges were occasional food items. This species
showed signs of circumstancial cannibalism. The seasonal diet patterns of
P. gunattus did not differ greatly, except in winter, when
the similarity index reached the lowest value compared to the rest of the
seasons. P. argusprefers crustaceans and mollusks, whereas
the rest of the trophic groups were accasional food items. Both species showed
a generalistic, opportunistic, plastic and selective feeding
behavior.
Las relaciones tróficas son un aspecto importante de la biología de una especie, ya que constituyen un elemento del nicho ecológico, y determinan en gran medida algunos parámetros poblacionales, como la abundancia, densidad y distribución (Berry, 1977); ciertas relaciones de simbiosis, depredación y parasitismo, asi como aspectos de migración y selección de habitat (Kanciruk, 1980; Olsen et al., 1975). Panulirus argus y P. guttatus son especies de langosta cuya distribución geográfica es muy similar, encontrándose principalmente en el Caribe (Phillips et al. 1980a). Pertenecen a la familia Palinuridae del orden Decapoda y presentan una larga serie de fases larvarias planctónicas, que difieren considerablemente de las de los demás decápodos y de cuyos alimentos naturales y hábitos alimenticios apenas se tiene información (Provenzano, 1968). Robertson (1968) indica que los nauplios de Artemia spp.,son satisfactorios para alimentar larvas filosoma de P. argus y P. guttatus en sus primeros estadios. Las langostas adultas poseen un sistema quimiorreceptor bien desarrollado (Heydorn, 1968; Conklin, 1980) que les permite detectar la fuente de alimento. Una vez capturado, éste es triturado por las mandíbulas y pasado al estómago, donde el molino gástrico ejerce una segunda trituración (Heydorn, 1968; Herrnkind, et al., 1975; Phillips et al., 1980a.). Esto dificulta en gran medida la identificación detallada de los organismos ingeridos. La familia Palinuridae contiene especies con un amplio espectro de patrones alimenticios que van desde un régimen de herbivoría parcial descrito por Beurois (1971) paraJasus paulensis, hasta la preferencia por la carroña fresca en Panulirus cygnus (Phillips et al. 1980b). Sin embargo, parece haber un predominio del omnivorismo tal como lo demuestran los trabajos de Lindberg (1955) en P. interruptus, Heydom (1968) en Jasus lalandii, Berry (1971) en P. homarus, Joll (1984) en P. cygnus, y Aramoni (1982) en P. inflatus y P. gracilis. Aunque no demostrado plenamente, se ha reportado cierto grado de canibalismo en algunas especies. Tal es el caso deP. gracilis y P. inflatus en el Pacífico mexicano (Aramoni, 1982) y de P. argusen las Islas Virgenes (Olsenet al., 1975). Los patrones alimenticios de P. argus han sido estudiados por Herrnkind et al. (1975) y Olsen et al. (1975) en las Islas Vírgenes, describiendo sus movimientos de forrajeo y la composición de su dieta. Estos autores identificaron ocho diferentes tipos de alimento, y concluyeron que esta especie tiene una alimentación tipicamente omnívora. También demostraron que los organismos adultos y los juveniles de P. argus exhiben distintos patrones de alimentación. No se llegó a demostrar si esto se debe a las preferencias alimenticias de los organismos, o bien a la disponibilidad de los alimentos en los distintos ambientes en los que se encuentran, ya que los ejemplares adultos de esta especie ocupan diversos tipos de ambientes, mientras que los juveniles se distribuyen casi exclusivamente en áreas someras con vegetación marina (Kanciruk, 1980; Marx y Herrnkind, 1985). Se ha observado también que las hembras ovígeras tienen alteraciones con respecto al comportamiento alimenticio en los períodos de actividad reproductiva (Phillips et al., 1980a), aunque Chittleborough (1975) encontró una excepción en P. cygnus. No existen en la literatura citas sobre la alimentación de P. guttatus. Se plantea como objetivo general de este trabajo, conocer y comparar los patrones de alimentación de las especies Panulirus guttatus y P. argus en la zona de Puerto Morelos, Q.R., asi como determinar las diferencias alimenticias que pudieran presentarse entre los sexos de ambas especies y detectar las posibles variaciones estacionales en la estructura de estas dietas. ÁREA DE ESTUDIOEl área de estudio abarcó la zona costera del Estado de Quintana Roo comprendida entre Punta Nizuc: y Punta Brava (Fig. l), que corresponde al área de pesca de la Cooperativa "Pescadores de Puerto Morelos". A lo largo de la linea de costa de Quintana Roo y a una distancia variable, corre una barrera arrecifal. Esta constituye el elemento mas importante que caracteriza geomorfológica y ecológicamente esta área. Jordán (1979) divide el ambiente arrecifal en cuatro zonas principales. (1) La zona lagunar, que se extiende desde la orilla hasta el borde de la plataforma arrecifal en su parte posterior, y se caracteriza por sus fondos arenosos cubiertos porThalassia testudinum, con escasas formaciones coralinas y con profundidades generalmente no mayores a los 10 m. (2) La zona posterior, limitada hacia tierra por la zona lagunar y hacia el mar por densos bancos deAcropora palmata se distingue por una gran diversidad en especies de corales. (3) La zona de rompiente, que es una zona somera y dominada por pequeñas y aisladas colonias de coral. (4) Por último, la zona frontal, que se extiende desde la zona de rompiente, hasta la parte inferior de la formación arrecifal y que queda expuesta a mar abierto. Su extensión está determinada tanto por el grado, generalmente suave, de la pendiente del sustrato rocoso calcáreo, como por la profundidad a la que se encuentre la plataforma arenosa, en el orden de los 20-30 m. 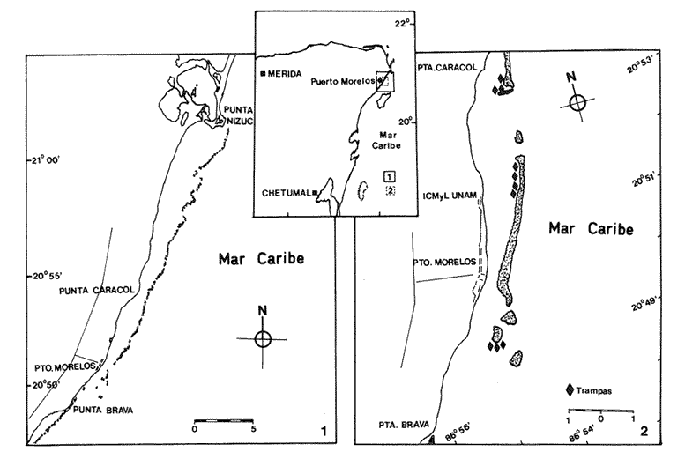 Figura 1. Área de estudio. 1)Área de colecta de especímenes de Panuliru argus; 2) Área de colecta de especímenes de P. guttatus. MATERIAL Y MÉTODOSLa información que contiene este trabajo se obtuvo durante el lapso comprendido entre noviembre de 1986 y noviembre de 1987, con la finalidad de abarcar por lo menos un período anual de muestreo. El método de muestreo utilizado en la obtención de los ejemplares fué distinto para cada especie, ya que sus estadios adultos tienen diferentes hábitats. La captura de langosta en el Estado de Quintana Roo, como en casi toda la zona del Caribe, está marcadamente sesgada hacia la especieP. argus, que se encuentra fundamentalmente en la zona frontal del arrecife y sobre la plataforma continental, constituyendo cerca del 100% del total de la captura de estos crustáceos. Por el contrario, la especie P. guttatus se encuentra en las formaciones del coral escleractinio Acropora palmata (Sutcliffe, 1953; Munro, 1974; Aiken, 1975; Briones et al., 1988), y por tanto se restringe casi exclusivamente al área que ocupa la barrera arrecifal. Prácticamente no existe una pesca comercial de P. guttatus. Los ejemplares de P. argus fueron colectados por algunos pescadores, quienes después de capturar al animal empleando el gancho langostero por medio de buceo autónomo, seccionan el abdomen del cefalotórax, cediendo éste último para el presente estudio. Los ejemplares de P. guttatus y algunos de P. argus se capturaron mediante trampas langosteras colocadas a lo largo de la parte posterior de la linea arrecifal. La carnada consistió en orejas de res. Se intentó que las langostas que penetraran en las trampas no pudieran ingerir la carnada, pero esto no siempre se logró. Las trampas se colocaron durante quince días cada mes, abarcando los períodos lunares de cuarto menguante hasta cuarto creciente, períodos en los que se ha registrado la mayor actividad de estos organismos (Sutcliffe, 1953; Phillips et al., 1980b). Las trampas se revisaban diariamente durante los periodos mencionados, entre las 06:00 y las 07:00 horas, mediante buceo libre. Los ejemplares capturados fueron sexados, medidos y transportados inmediatamente al laboratorio. Cuando no fué factible realizar la disección inmediatamente, los ejemplares se congelaron para detener el proceso digestivo. La extracción del estómago se realizó según el método descrito por Aramoni (1982), que consiste en separar la porción dorsal del cefalotórax para dejar expuestos los órganos internos, facilitando la extracción del tracto digestivo anterior y el estómago. Una vez retirados, los estómagos se fijaron en formol al 10% por lo menos durante dos horas, después de lo cual se conservaban en alcohol al 70%. Posteriormente, se extraía el contenido estomacal de cada tracto, registrando el número de estómagos vacíos (EVT) para poder obtener un porcentaje de vacuidad por especie y por estación del año. Este porcentaje incluyó tanto los estómagos totalmente vacios (EV) como aquéllos cuyo contenido consistió exclusivamente de carnada (ECAR), que para fines prácticas también se consideraron como vacíos. Este valor (EVT), se relacionaba con el número total de estómagos obtenidos (IND) de cada sexo. El contenido estomacal se separó en distintos grupos tróficos, procurando llevar la identificación de cada fracción separada hasta el menor taxón posible, teniendo en cuenta la digestión y trituración sufrida en el estómago de la langosta. La posición taxonómica de las especies de gasterépodos, es la adoptada por Thielle (García-Cubas, 1981), y la de los crustáceos por Bowman y Abele (1982). Una vez separadas, las muestras se secaban a temperatura ambiente con objeto de obtener el peso seco de cada fracción, mediante una balanza analítica (Sauter 414) con 0.1 mg de precisión. Los grupos tróficos considerados fueron los siguientes: algas coralinas, materia vegetal, esponjas, equinodermos, crustáceos, género Panulirus, moluscos y ascidias. Los grupos con muy escasa representación, como los anélidos, foraminíferos, fragmentos óseos y escamas de peces, briozoarios y sipuncúlidos, se agruparon como "otros". También se tomó en cuenta la materia orgánica no identificada (MONI). Se consideró al género Panulirus como un grupo aparte, con objeto de poder analizar el posible papel que juega el canibalismo en la dieta de las langostas estudiadas. Se cuantificó el número de grupos tróficos presentes en cada estómago, de tal forma que al agrupar la muestra por sexos, se pudiera encontrar la distribución de estos estómagos en función de la diversidad que presentaron, obteniéndose un patrón de diversidad para los machos y otro para las hembras de cada especie. PROCESAMIENTO DE LA INFORMACIÓNHyslop (1980) menciona que ningún método de análisis del contenido estomacal, por sí mismo, ofrece una imagen completa del patrón dietético, por lo que debe utilizarse por lo menos un método que permita la estimación de las cantidades y otro que mida el volumen o el peso del contenido estomacal. Con base en esto, se obtuvieron dos valores para cada uno de los grupos tróficos presentes en cada estómago: por un lado, su porcentaje de peso (gravimétrico) con respecto al peso total de la muestra y por otro, su frecuencia de aparición o presencia con respecto al número total de estómagos analizados en cada muestra. Ambos porcentajes pueden calcularse después de agrupar por especie, por sexo y por fecha, con objeto de obtener los patrones de alimentación para ambos sexos, así como su comportamiento estacional. Para comparar los resultados del porcentaje de peso en el análisis por sexo y por estaciones, se utilizó el índice de similitud (IS) de Jaccard modificado por Spatz (Müller-Dumbois y Ehlemberg, 1974), que es uno de los que demuestran una mayor sensibilidad a las diferencias tanto cuantitativas como cualitativas entre losbloques a comparar. La notación de este índice (ISsp) es la siguiente:
donde R = la relación entre el porcentaje de peso más pequeño y el más grande de cada uno de los grupos tróficos comunes a los dos bloques a comparar; Mc = suma de los valores de porcentaje de peso de los grupos tróficos comunes a ambos bloques; Ma = suma de los valores de porcentaje de peso de los grupos tróficos del bloque A, y Mb = lo mismo que Ma, pero para el bloque B. Los valores umbrales del índice de similitud son los siguientes: por debajo del 25%, los bloques pueden ser considerados como entidades separadas; entre 25 y 50%, los bloques comparados pueden considerarse como subconjuntos de una misma muestra, y con valores superiores al 50%, la similitud es tan grande que deben ser tomados como bloques iguales. Los grupos tróficos identificados pudieron clasificarse de acuerdo a la importancia que tienen dentro de la dieta del organismo, según el índice de importancia relativa (IIR), descrito por Yañez-Arancibia et al. (1976) y Hyslop (1980), dado por la relación IIR = F x W / 100, donde IIR es el indice de importancia relativa que tiene cada grupo trófico en particular, F representa la frecuencia o porcentaje de presencia y W representa el porcentaje gravimétrico del alimento ingerido (grupo tráfico) en cuestión. Este índice ha resultado de gran utilidad para caracterizar principalmente las dietas de peces (Gracia y Lozano, 1980; Yáñez-Arancibia et al., 1976). La combinación de la frecuencia y el peso, evaluados por el IIR, permite graficar el espectro trófico, que sirve como auxiliar para hacer una clasificación de los grupos tróficos, dependiendo del cuadrante en el cual incidan estos valores, en: I) Grupos Tróficos, Ocasionales, Accidentales o Circunstanciales, cuyos porcentajes de peso o frecuencia no sobrepasan el valor de 20% y el de IIR no es mayor a 10%; II) Grupos Tróficos Secundarios, donde los valores de presencia y peso van de 20 a 40% y su IIR varia entre 10 y 40%; III) Grupos Preferenciales, definido por los valores de peso y presencia entre 40 y 100%, y con el mismo intervalo de valores para el, IIR. TRATAMIENTO ESTADÍSTICOSe utilizó una prueba de diferencia entre proporciones detectar posibles- variaciones entre los valores del índice de vacuidad por sexo, para cada una de las e species. A los valores de los índices de vacuidad por estación, se les aplicó una prueba de X² en un nivel de significancia (p) de 0.05 (Hoel, 1976). La misma prueba se llevó a cabo con los resultados de diversidad de alimentos en los estómagos de hembras y machos de ambas especies. RESULTADOS Y DISCUSIÓN
Panulirus gattatus
Indices de vacuidadSe colectaron 226 ejemplares adultos durante todo el período de estudio (44.5 a 87.5 mm longitud cefalotorácica, X = 63.3 mm), de los cuales 68 fueron hembras y 158 fueron machos. Del total de hembras capturadas, solo 34 presentaron ontenido estomacal, lo que representa un 50.0% de vacuidad. Para los machos el porcentaje de vacuidad fué de 52.5% (83 individuos). La diferencia entre estos valores no resultó significativa. El porcentaje de vacuidad para ambos sexos en conjunto fue de 51.8%. Esto significa que no existe una diferencia en el tipo de comportamiento alimenticio entre los sexos. Aramoni (1982) encuentra también una similitud en el porcentaje de vacuidad para ambos sexos en P. inflatus y P. gracilis. El hecho de que más de la mitad de los ejemplares hayan tenido el estómago vacío puede deberse a varios factores, entre los cuales se encuentra el muestreo con trampas, que tiende a favorecer la captura de organismos hambrientos por ser atraídos más fuertemente por la carnada, o la hora de revisión de las trampas. No se ha determinado la hora de máxima actividad de alimentación de esta especie, aunque es probable que tengan cuando menos cierta actividad en las primeras horas de la madrugada. En la Tabla 1, se observan las variaciones estacionales del porcentaje de vacuidad para esta especie, así como el número de individuos capturados por estación y el número, de estómagos que contuvieron algun tipo de contenido estomacal. Destacan los valores de primavera 1987, en el cual se obtuvo el mayor número de organismos (61) y el porcentaje de vacuidad más bajo (44.3%). Los mayores porcentajes de vacuidad corresponden al invierno y verano de 1987. El periodo con el menor número de individuos capturados (29), correspondió al otoño de 1986, No hubo diferencias significativas entre los valores estacionales de vacuidad. Esta homogeneidad en el porcentaje de vacuidad estacional podría indicar que esta especie mantiene sin cambios su comportamiento alimenticio a lo largo del año, es decir, que no hay una estación en la que marcadamente dejen de alimentarse. 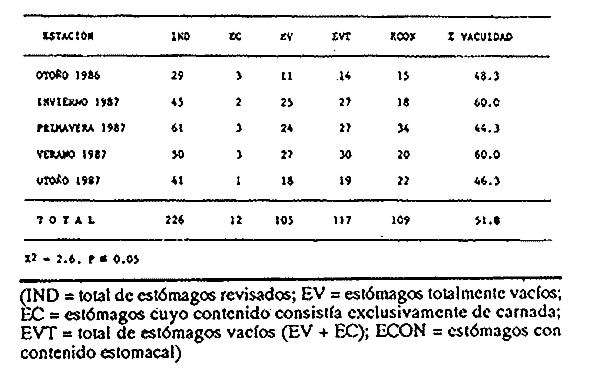 TABLA 1 PORCENTAJE DE VACUIDAD EN ESTOMAGOS DE EJEMPLARES DE Panulirus guttatus POR ESTACIÓN DEL AÑO DiversidadLa diversidad de grupos tróficos encontrados en los estómagos de P. guttatus (Tabla 2), evidencia la gran cantidad de estómagos vacíos en ambos sexos, así como el que fueran más frecuentes los ejemplares con solamente uno o dos grupos tróficos en sus estómagos, siendo sumamente raros los que contenían mas de cuatro. Tan solo una hembra en toda la muestra presentó siete de los diez grupos tróficos identificados. Las diferencias de estos valores entre machos y hembras, no resultaron significativas (p=0.05). Esto indicaría que el carácter generalista y oportunista de la especie en términos alimenticios, no está simplemente dado por la conjunción de las preferencias individuales de los organismos que constituyen la población sino que, además, cada individuo refleja en su dieta estas características. Por tanto, es posible que todos los elementos constituyentes de la dieta jueguen un papel importante en el desarrollo del organismo, aunque quizá en diferentes grados, y que los requerimientos individuales se vean reflejados en los requerimientos de la especie en forma global. 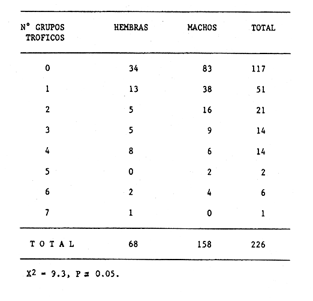 TABLA 2 NÚMERO DE HEMBRAS Y MACHOS DE Panulirus guttatus QUE CONTENÍAN ENTRE 0 Y 7 GRUPOS TRÓFICOS EN SU CONTENIDO ESTOMACAL Porcentajes de peso y presencia de los grupos tróficos.Los porcentajes globales de peso y de presencia de los distintos grupos tróficos encontrados en los estómagos de hembras y machos de esta especie para todo el año, se muestran en la figura 2. Destacan, en ambos sexos, los grupos de crustáceos, moluscos y algas coralinas. El grupo tróficoPanulirus adquiere importancia sólo en los machos. 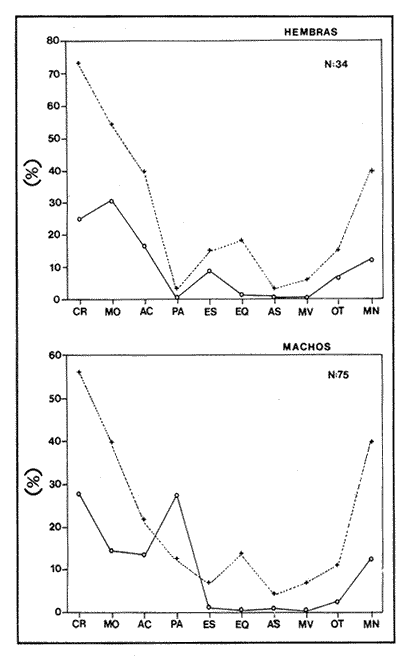 Figura 2. Espectro trófico para, hembras y machos de Panulirus guttatus. (CR = crustáceos; MO = moluscos; AC = algas coralinas; PA = Panulirus; ES = esponjas; EQ = equinodermos; AS = ascidias; MV = materia vegtal; OT = "otros"; MN=materia orgánica no identificada. Línea continua=porcentaje de presencia; línea quebrada=porcentaje de peso. El porcentaje de presencia de grupos tróficos es adecuado en el caso del análisis del contenido estomacal de las langostas, ya que la extrema trituración que sufren los alimentos dificulta la cuantificación del número de individuos ingeridos, aunque tiene la desventaja de sobrestimar los alimentos constitutídos por organismos pequeños o en escaso número. Por otro lado, el porcentaje de peso (gravimétrico) tiende a subestimar los alimentos con escaso p m., aún cuando se hubiesen presentado frecuentemente, y a sobrestimar alimentos de gran peso y poca frecuencia. Ambos métodos, tanto el de presencia como el gravimétrico, se ven afectados por el hecho de que los crustáceos, moluscos, equinodermos y algas coralinas, están constituídos en gran medida por partes duras que, por un lado, tienen un peso específico mayor que las partes blandas, y por otro, pueden tener una tasa de evacuación diferente. Esto confiere cierto sesgo a los resultados en el sentido de aumentar el valor de participación gravimétrica, así como el porcentaje de presencia de estos grupos. Joll (1984), encontró una rápida tasa de evacuación de alimentos en Panulirus cygnus, alcanzando un 50% de ejemplares con el estómago vacío cuatro horas después de ser alimentados con moluscos gasterópodos, y seis horas después de alimentarse con algas coralinas. Los resultados de Joll contrastan con una más lenta evacuación de las partes duras en la dieta del portúnido Scylla serrata, en la que se han encontrado remanentes de éstas seis dias después de su ingestión (Hill, 1976), y con las de Homarus americanus, en la que los restos de equinodermos, moluscos, crustáceos y poliquetos permanecen en el tracto digestivo hasta, 180 dias después de su ingestión (Carter y Steele, 1982). Aunque la tasa de evacuación de los alimentos puede variar de especie a especie, se ha considerado que, en general, los palinúridos la poseen elevada y la tendencia es que las partes duras sean evacuadas más lentamente (Berry, 1971; Herrnkind et al., 1975). De aquí que la comparación entre ambos coeficientes sea necesaria para obtener una idea más certera del patrón de alimentación. Es así como pueden explicarse las supuestas discrepancias entre los valores del porcentaje de peso y aquéllos del porcentaje de presencia para un determinado grupo trófico. Tal es el caso de los equinodermos (Fig. 2), que constituyen un grupo menor en términos de participación gravimétrica, aunque los datos de presencia indican que son un componente dietético común. Solamente en los machos se presentaron cantidades importantes de restos de organismos de la especiePanulirus, aunque no con un gran porcentaje de frecuencia. Estos resultados son compatibles con los registrados por Joll (1984), quien encontró porcentajes variables de este componente en estómagos deP. cygnus. Heydorn (1968) explica que la presencia de estos restos se debe a la ingestión de fragmentos de exuvias de congéneres como fuente de oligoelementos para las langostas en proceso de muda, lo que implicaría un fuerte ahorro energético. Sin embargo, en este trabajo también podría jugar un papel importante el estado de "stress" al que pueden verse sometidos los organismos en las trampas, lo que alteraría su comportamiento, además de que podría favorecer la autotomía de los apéndices, que son las partes encontradas con mayor frecuencia en los contenidos estomacales. En algunas ocasiones en que dos o más langostas eran capturadas en la misma trampa, alguna de ellas había perdido un apéndice, mientras que otra se encontraba ingiriéndolo al momento de la colecta, sin que se pudiera establecer si el apéndice en cuestión había sido o no autotomizado. Todos estos elementos, aunados al hecho de que en ocasiones las partes de exoesqueletos encontrados en el tracto digestivo del animal aún presentasen tejidos blandos, ponen en duda, en el caso de P. guttatus, que la ingestión de estos fragmentos se deba exclusivamente a la necesidad de oligoelementos para el endurecimiento del caparazón, y viene a reforzar la idea de una alimentacion oportunista en esta especie, sin implicar necesariamente un comportamiento agresivo. Estacionalmente, se observan fluctuaciones en ambos porcentajes de los grupos tróficos que conforman la dieta de la especie P. guttatus (Fig. 3), aunque pueden identificarse grupos que se mantienen con valores relativamente elevados a lo largo del año, tales como los crustáceos, los moluscos, y las algas coralinas. Otros grupos, por el contrario, conservan valores bajos durante el período de estudio (por ejemplo, ascidias y materia vegetal). Los grupos restantes:Panulirus, esponjas, equinodermos y "otros", se caracterizan por tener grandes fluctuaciones en el tiempo. Esta variación estacional, así como la diversidad de los componentes alimenticios, podrían parcialmente reflejar las condiciones del hábitat de esta especie (Miller et al., 1971). Conell (1978) sugiere que las comunidades arrecifales, de una gran diversidad, mantienen un equilibrio dinámico basado en oscilaciones constantes tanto en el número de especies como en el número de individuos de las poblaciones que conforman a cada especie. Por otro lado, se ha encontrado que las langostas Homarus americanus parecen tener cierta selectividad en sus alimentos, es decir que la proporción de los grupos tróficos en los estómagos difiere de su abundancia relativa en el hábitat (Phillips et al., 1980a). Ennis (1973) encontró cambios importantes en la proporción de crustáceos y equinodermos en la dieta de H. americanus relacionados con los períodos de muda. Los crustáceos proveen a H. americanus de alrededor de quince veces más energía y cuatro veces más proteína que los equinodermos (Evans y Mann, 1977), pero éstos últimos les proveen de más calcio cuando necesitan endurecer su exoesqueleto (Ennis, 1973). De esta manera, podrían interpretarse las fluctuaciones en la proporción de algunos de los grupos mencionados, como el resultado de una cierta plasticidad en la dieta de estos crustáceos ante circunstancias ambientales, así como a preferencias y requerimientos intrínsecos de la especie. 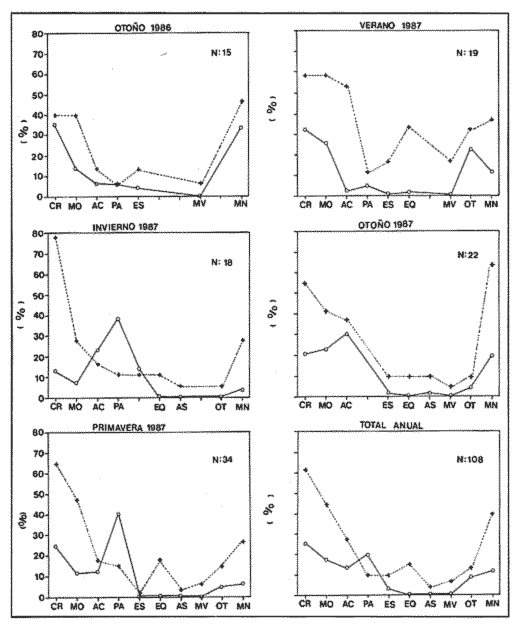 Figura 3. Espectro trófico estaciona¡ y anual de Panulirus guttatus. (CR = crustáceos; MO = moluscos; AC = algas coralinas; PA = Panulirus; Es = esponjas; EQ = equinodermos; AS = ascidias; MV = materia vegetal; OT = "otros"; MN=materia orgánica no identificada. Línea continua=porcentaje de presencia; línea quebrada=porcentaje de peso). Indices de similitud (IS)El índice de similitud entre los porcentajes gravimétricos del contenido estomacal de hembras y machos de P. guttatus, fue de 53.6, es decir que los sexos no presentan diferencias significativas en su alimentación. A partir de los valores del porcentaje de peso se computaron los índices de similitud entre las distintas combinaciones de pares de estaciones del año, los cuales se exponen en la Tabla 3. Dados los bajos valores del índice de similitud estacional entre grupos tróficos que se presentan en casi todas las combinaciones en las que se incluye el invierno, podría pensarse que es en esta estación en donde se presentan los cambios más severos en todo el año, pues al ser comparados con el otoño de 1986, el otoño del 1987 y el verano de 1987, ninguno de estos valores sobrepasa el valor umbral del 25% para poder considerarse como similares.  TABLA 3 INDICES DE SIMILITUD ENTRE LOS CONTENIDOS ESTOMACALES DE Panulirus guttatus EN LAS ESTACIONES DEL AÑO, CON BASE EN LOS PORCENTAJES DE PESO DE LOS GRUPOS TRÓFICOS Los demás binomios estacionales comparados mantienen sus valores dentro del rango de aceptabilidad de similitud (25-50%), lo que podría deberse a una graduación en el cambio de condiciones alimentarias entre estación y estación, a excepción del invierno. De nueva cuenta esto es reflejo de la plasticidad dietética ante circunstancias ambientales. Indice de importancia relativa (IIR)La figura 4 representa el diagrama trófico combinado de P. guttatus, y muestra los valores anuales del porcentaje de peso y de presencia, así como el índice de importancia relativa (IIR), de los distintos grupos tróficos. Este diagrama permite caracterizar al grupo de los crustáceos como un alimento preferencial, seguido por los moluscos y las algas coralinas, que ocupan un relevante lugar como alimento secundario. Joll y Phillips (1984) subrayan la importancia que tienen para P. cygnus las algas. coralinas como fuente de calcio para la formación del exoesqueleto de los individuos que se encuentran en las primeras fases de intermuda. Los demás grupos tróficos caen en el rango de alimentos ocasionales o circunstanciales, haciendo patente la mínima contribución que tiene la materia vegetal en la conformación del patrón alimenticio de P. guttatus. Panulirus argus
Indices de vacuidadDe esta especie se obtuvo un total de 90 ejemplares (56.0 a 152.9 mm de longitud cefalotorácica, X= 91.2 mm). En las 45 hembras obtenidas, se alcanzó un 75.6% de vacuidad. En el caso de los 45 machos, la vacuidad fué de 66.7%. No existen diferencias significativas entre los porcentajes de vacuidad de los dos sexos de esta especie. Este mismo porcentaje para machos y hembras en conjunto fue de 71.1%. Se observan porcentajes de vacuidad elevados en todas las estaciones (Tabla 4), siendo el mayor el correspondiente al verano 1987, y el menor el de otoño de 1986. Estos altos porcentajes de vacuidad podrían deberse a un artificio del muestreo, ya que la pesca comercial de este crustáceo se lleva a cabo entre las 08:00 y las 14:00 horas. Herrnkind et al. (1975) determinaron que el periodo de mayor actividad alimentaria de esta especie está comprendida entre las 17:00 y las 06:00 horas, por lo que transcurrió un lapso considerable entre la alimentación y la hora del procesamiento de las muestras. DiversidadLos valores de diversidad de grupos tróficos en los estómagos deP. argus, no muestran diferencias significativas entre sexos (p=0.05). Al igual que en P. guttatus, abundan los estómagos con pocos grupos tróficos en su interior, siendo un estómago perteneciente a una hembra el que mayor diversidad presentó, con ocho diferentes tipos de alimentos (Tabla 5). Porcentajes de peso y presenciaLa figura 5 muestra los porcentajes de peso y presencia de los componentes tróficos en ambos sexos. Estos componentes se presentan en la misma secuencia que para la especie P. guttatus, con fines comparativos. Del total de la muestra, incluidos ambos sexos, dos grupos destacan sobre todos los demás como los poseedores de los valores más altos, tanto del porcentaje de presencia como del de peso: crustáceos y moluscos. La materia orgánica no identificada (MONI) tiene una representación elevada, sobre todo en términos del porcentaje de presencia, pues se trata en gran parte de material ya digerido, debido al tiempo transcurrido desde el momento de la ingestión hasta el de la fijación. El grupo de los moluscos es el que alcanza un mayor porcentaje de aparición en los tractos de los ejemplares de esta especie, causado tal vez por la tasa de evacuación diferencial de acuerdo a la dureza del tejido. Es posible también que algunos de los fragmentos de conchas presentes en la muestra y que se contabilizan como moluscos, fueran en realidad conchas que albergaban anomuros (cangrejos ermitaños), que debieran formar parte del grupo de los crustáceos. Las ascidias no se presentaron en los contenidos estomacales de las hembras; sin embargo, se encontró un mayor porcentaje de peso de los equinodermos y las algas coralinas en las hembras que en los machos. 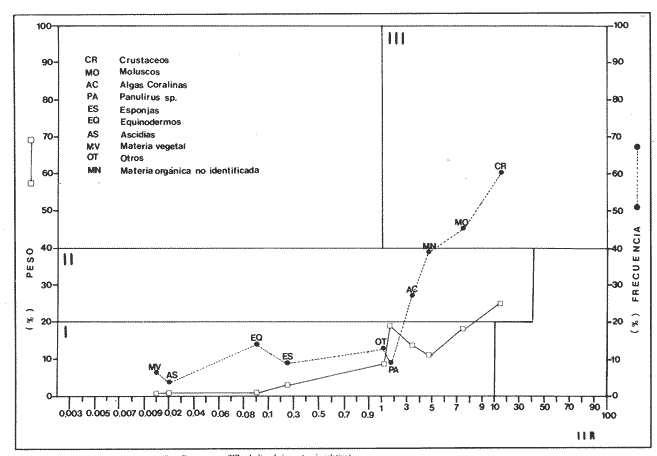 Figura 4. Diagrama trófico combinado de Panulirus guttatus (HR = Indice de importancia relativa). 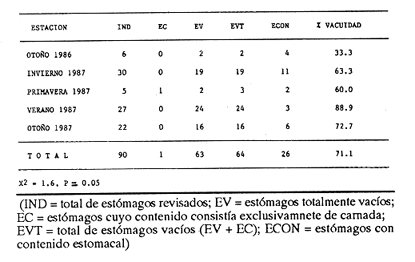 TABLA 4 PORCENTAJE DE VACUIDAD EN ESTÓMAGOS DE EJEMPLARES DE Panulirus argus POR ESTACIÓN DEL AÑO (IND = total de estómagos revisados; EV = estómagos totalmente vacíos; EC = estómagos cuyo contenido consistía exclusivamente de camada, EVT = total de estómagos vacíos (EV + EC); ECON = estómagos con contenido estomacal) 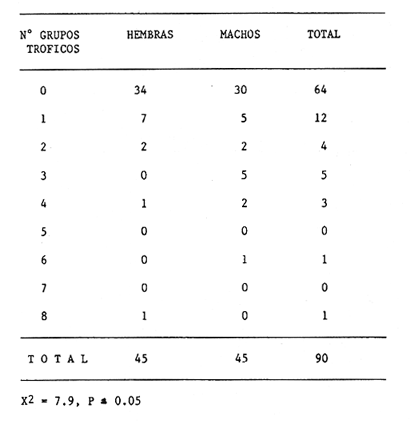 TABLA 5 NÚMERO DE HEMBRAS Y MACHOS DE Panulirus argus QUE CONTENÍAN ENTRE 0 Y 8 GRUPOS TRÓFICOS EN SU CONTENIDO ESTOMACAL Resalta en esta especie la ausencia del componente Panulirus, a diferencia de lo encontrado en P. guttatus, cuyos ejemplares fueron capturados por trampas, lo cual apoya la hipótesis del "stress" para explicar el cariz oportunista del canibalismo. También es evidente la ausencia de materia vegetal, aunque en ambos casos existe la posibilidad de la influencia del tamaño pequeño de la muestra. El análisis estacional de esta especie no se pudo efectuar debido al escaso número de ejemplares colectados. Indice de similitud (IS)Se obtuvo un índice de similitud entre los contenidos estomacales de hembras y machos de esta especie de 35.3, es decir que, a pesar de las diferencias particulares en el porcentaje gravimétrico de los grupos tróficos constitutivos de la dieta de ambos sexos, sus patrones dietéticos son en gran medida similares. Indice de importancia relativa (IIR)La figura 6 representa el diagrama trófico combinado de P. argus, y muestra los índices de importancia relativa de los grupos tróficos encontrados en los tractos digestivos de los ejemplares de esta especie. Este diagrama indica claramente la importancia que tienen los crustáceos y los moluscos como alimentos preferenciales de P. argus, lo que delinea un patrón similar al deP. guttatus. Existe, sin embargo, una diferencia en lo concerniente a las algas coralinas, ya que si bien éstas son un alimento secundario para P. guttatus, devienen en alimento ocasional paraP. argus. Identificación taxonómica de los alimentosEl grado de trituración y digestión a que son sometidos los organismos ingeridos impide realizar una determinación taxonómica detallada. En algunos casos, sin embargo, fueron identificados algunos elementos a nivel de especie, aunque la mayoría de ellos solo pudieron determinarse a nivel de clase o familia. Los resultados de este análisis se presentan en las tablas 6 y 7, que corresponden a los contenidos estomacales de P. guttatusy P. argus respectivamente. Es posible que, en el caso de P. argus, la lista fuera mayor si se hubiera obtenido un número de ejemplares más abundante o en horas más cercanas a su horario de alimentación. Inicialmente se había planteado la posibilidad de una superposición de nichos ecológicos en lo referente a alimentación entre ambas especies, con base en su cercanía taxonómica, su distribución geográfica similar y la parcial, aunque temporal, compartición del hábitat arrecifal. Los resultados de este estudio señalan una gran similitud en la configuración de los patrones dietéticos de ambas especies, a pesar del menor número de ejemplares de P. argusanalizados. 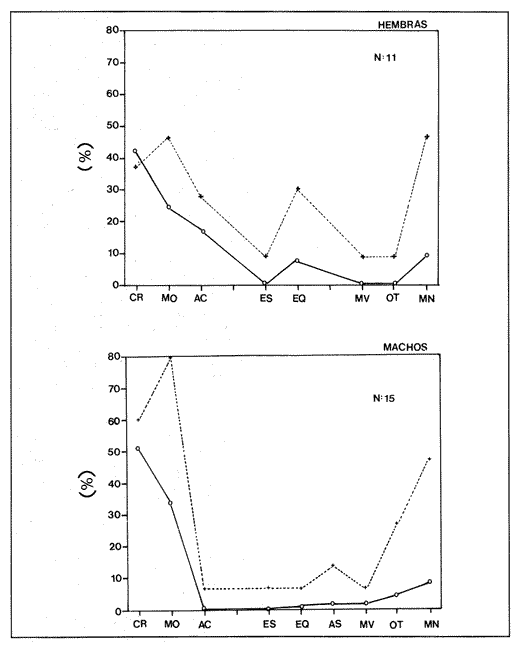 Figura 5. Espectro trófico para hembras y machos de Panulirus argus. (CR = crustáceos; MO = moluscos, AC = algas coralinas; ES = esponjas; EQ = equinodermos; AS = ascidias; MV = materia vegetal; OT = "otros"; MN=materia orgánica no identificada Línea continua=porcentaje de presencia; línea quebrada=porcentaje de peso) 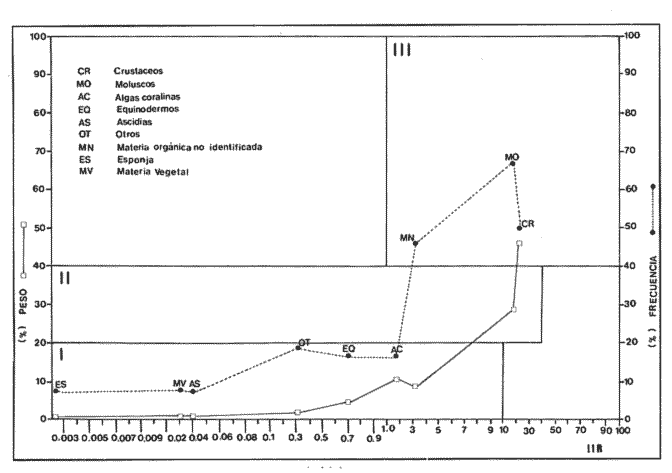 Figura 6. Diagrama trófico combinado de Panulirus argus. (IIR = Indice de importancia relativa).  TABLA 6 IDENTIFICACIÓN TAXONÓMICA DE LOS ORGANISMOS ENCONTRADOS EN LOS CONTENIDOS ESTOMACALES DE Panulirus gattatus Sin embargo, Gause (1934) establece que la intensidad de la competencia no está dada por la afinidad sistemática, sino por la similitud de los requerimientos de los competidores potenciales en el medio, y Andrewartha y Birch (1954) señalan que, aún cuando dos o más especies consuman los mismos tipos de alimento, esto podría denotar una superabundancia de ese alimento en particular, o bien que estas especies se alimenten de distinto material a nivel específico. Los resultados del presente estudio permiten determinar que P. argus y P. guttatus tienen patrones alimenticios similares, pero no que ambas especies compitan por las mismos alimentos.  TABLA 7 IDENTIFICACIÓN TAXONÓMICA DE LOS ORGANISMOS ENCONTRADOS EN LOS CONTENIDOS ESTOMACALES DE Panulirus argus Conclusiones- Panulirus guttatus posee un espectro alimenticio ligeramente más amplio que P argus, aunque esta comparación es tentativa debido al menor número de ejemplares de P argus analizados. - En ambos casos el grupo de los crustáceos debe ser considerado como un alimento preferencial. - El grupo de los moluscos es un alimento preferencial para P. argus, y secundario, pero relevante, para P. guttatus. - Las algas coralinas tienen un carácter de alimento secundario para P. guttatus, y de ocasional para P. argus. - Los equinodermos son un grupo encontrado frecuentemente en los estómagos de ambas especies, aunque su contribución gravimétrica los relega a un papel de alimento ocasional. - Se detectó la existencia de canibalismo en P. guttatus capturadas con trampas, infiriéndose el cariz oportunista en su alimentación. No se presentaron restos de congéneres en los especímenes de P. argusanalizados. - La materia vegetal, ascidias, esponjas y el resto de los grupos considerados como "otros", quedan relegados a alimentos ocasionales, tanto en P. argus como en P. guttatus. - No existe una notable diferencia entre los patrones dietéticos de los dos sexos de ninguna de las dos especies, a excepción hecha del grupo Panulirus, que se encuentra con mayor frecuencia en los machos de P. guttatus. - Estacionalmente, hay ligeras variaciones en la composición dietética de P. guttatus, siendo el invierno la estación que presenta una mayor diferencia debido, tal vez, a que en esta estación imperan condiciones más severas, lo que significa un cierto grado de plasticidad en la alimentación de esta especie. - Puede considerarse la dieta de estos crustáceos como omnívora - aunque fundamentalmente carnívora -, generalista, plástica y selectiva. AgradecimientosLos autores desean agradecer la valiosa ayuda de Fernando Negrete, Gabriela Contreras, Martha Fonseca y Jorge Simonín en el trabajo de campo; de MariCarmen Martínez en la identificación de restos de crustáceos; de David Gutiérrez en el tratamiento estadístico, y de Carmen Becerril en la elaboración de las figuras. Lourdes Segura y Enrique Lozano revisaron acertadamente el manuscrito. La realización del trabajo fué posible gracias a una beca del CONACYT para el primer autor, y a un permiso de pesca científica extendido por la Secretaría de Pesca. Este trabajo forma parte del proyecto "Biología y Dinámica Poblacional de las Langostas del Caribe Mexicano" realizado por convenio entre el ICMyL- UNAM y el CONACYT (Clave PCECBNA-021927). LITERATURAAIKEN, K.A.Min. Agric. Fish. Div., Mar. Fish. Res. Rep. Spiny lobster investigations.(Jamaica)1975.1-24. ANDREWARTHA, H.G. y L.C. BIRCH The Distribution and Abundance of Animals.The University of Chicago PressChicago.1954.782p. ARAMONI, G.P. Tesis profesional Alimentación de las langostas Panulirus inflatus (Bouvier) y P. gracilis Streets en Zihuatanejo, Guerrero, y su relación con el bentos. Fac. de Ciencias, Univ. Nal. Autón. México.1982. 66 p. BERRY, P.F. Oceanogr. Res. Inst. (Durban) Invest. Rep. The biology of the spiny lobster Panulirus homarus (Linnaeus) off the east coast of southem Africa. 1971.1-75.28 BERRY, P.F. Circ. CSIRO Div. Fish. Oceanogr. A preliminary account of a study of biomass and energy flow in a shallow subtidal reef community on the east coast of South Africa, involving the rock lobster Panulirus homarus. (Australia)1977.1-24.7: BEUROIS, J.Tethis Régime alimentaire de la langouste Jasus paulensis (Heller1862) des îles Saint-Paul et Amsterdam (Océan Indien). Résultats préliminaires. 1971.943-948.(4): 3 BOWMAN, T.E. y L.G. ABELE The Biology of Crustacea, Vol. 1: Systematics, the Fossil Record and Biogeography. Classification of the Recent Crustacea. In: Abele, L.G. (Ed.) Academic Press Nueva York. 1982. 319 p. BRIONES, P., E. LOZANO, F. COLINAS y F. NEGRETE Informe final del Proyecto Biología de las Langostas del Caribe Mexicano. Convenio Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México / Consejo Nal. Cienc. tecnol., Clave PCECBNA-021927.1988. 231 p. CARTER, J.A. y D.H. STEELECan. JI. Zool. Attraction to and selection of prey by immature lobsters (Homarus americanus).1982 326-336. 60: CONELL, J.H. Science Diversity in tropical rain forest and coral reefs.1978.1302-1310.199 CONKLIN, D.E. The Biology and Management of Lobsters. Vol. 1: Physiology and BehaviorNutrition. In: Cobb, J.S. y B.F. Phillips (Eds.) Academic Press Nueva York. 1980. 462 p. CHITTLEBOROUGH, R.G. Aust. J. Mar. Freshwater Res.Environmental factors affecting growth and survival of juvenile Western rock lobster Panulirus longipes (Milne-Edwards).1975.177-196. 26: ENNIS, G.P. J. Fish. Res. Board Can. Food, feeding and condition of lobsters, Homarus americanus, throughout the seasonal cycle in Bonavista Bay, New foundland. 1973.1905-1909.30: EVANS, P.D. y K.H. MANN J. Fish. Res. Board Can. Selection of prey by American lobster, Homarus americanus, when offered a choice between sea urchins and crabs. 1977. 2203-2207.34: GARCIA-CUBAS, A. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Publ. Esp. Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de términos, Campeche). 1981.1-182. 5: GAUSE, G.F. Niche: theory and Application. Benchmark Papers in Ecology The struggle for existence. Hutchinson & Ross, Inc. DowdenPennsylvania1938.46-55. Vol. 3.1975. GRACIA, A. y E. LOZANOAn. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México Alimentación del bagre marino Netuma platypogon y su importancia como indicador de reclutamiento de postlarvas de langosta (Decapoda: Palinuridae) en Guerrero, México.1980.199-206. (2): 7 HERRNKIND, W.F., J. VANDERWALKER y L. BARR Nat. Hist. Mus. Los Angeles Cty. Sci. Bull. Population dynamics, ecology and behavior of spiny lobster Panulirus argus, of St. John, U.S.V.I.: Habitation and patterns of movements.1975.31-45.20: HEYDORN, A.E.F. S. Afr. Div. Sea Fish., Invest. Rep. The rock lobster of the South African west coast, Jasus lalandii (H. Milne-Edwards). II. Population studies, behaviour, reproduction, moulting, growth and migration.1968.1-52.71: HILL, BJ. Mar. Biol. Natural food, foregut clearance rate and activity of the crab Scylla serrata.1976.109-116.34: HOEL, P.G. Elementary Statistics. John Wiley & Sons, Inc. Nueva York.1976. 361 p. HYSLOP, E.J. J. Fish. Biol. Stomach content analysis -a review of methods and their application. 1980. 411-429.17: JOLL, L.M. Tesis Doctoral. Natural diet and growth of juvenile western rock lobsters, Panulirus cygnus George. Department of Zoology, University of WesternAustralia.1984. 194 p. JOLL, L.M. y B.F. PHILLIPS J. Exp. Mar. Biol. Ecol. Natural diet and growth of juvenile western rock lobsters Panulirus cygnus George. 1984.145-169.75: JORDAN, E. An. Centro Ciencias del Mar y Limnol. Univ. Nal. Autón. México Estructura y composición de arrecifes coralinos en la Región noreste de la península de Yucatán, México. 1979.87-96.(1): 6 KANCIRUK, P. The Biology and Management of Lobsters. Vol. 2. Ecology and Management. Ecology of juvenile and adult Palinuridae. In: Cobb, J.S. y B.F. Phillips (Eds.) Academic Press Nueva York.1980. 390 p. LINDBERG, R.G Growth, population dynamics and field behavior in the spiny lobster, Panulirus interruptus (Randall). Univ. Calif. Publ. Zool. 1955.157-248.59: MARX, J.M. y W.F. HERRNKINDBull. Mar. Sci. Macroalgae (Rhodophyta: Laurencia spp.) as habitat for young juvenile spiny lobsters, Panulirus argus. 1985.423-431.36: MILLER, R.J., K.H. MANN y DJ. SCARRATT J. Fish. Res. Board Can.Production potential of a seaweed-lobster community in eastern Canada. 1971.1733-1738. 28: MUNRO, J.L. Res. Rep. Zool. Dep., Univ. West Indies The biology, ecology, exploitation and management of Caribbean reef fishes. Part V.1. the biology, ecology and bionomics of Caribbean reef fishes: Crustaceans (spiny lobsters and crabs). 1974. 1-57.3: MÜLLER-DUMBOIS, D. y H. EHLENBERG Aims and Methods in Vegetation Ecology John Wiley & Sons Nueva York.1974. OLSEN, D.A., W.F. HERRNKIND y R.A. COOPER Nat. Hist. Mus. Los Angeles Cty.. Sci. Bull Population dynamics, ecology and behavior of spiny lobsters Panulirus argus, of St John, U.S.V.I.,II. Introduction and general population characteristics.1975. 11-16. 20: PHILLIPS, B.F., J.S. COBB y R.W. GEORGE General Biology. The Biology and Management of Lobsters. VoL 1: Physiology and Behavior. In: Cobb, J.S. y B.F. Phillips (Eds.) Academic Press, Nueva York.1980a. 462 p. PHILLIPS, B.F., G.R. MORGAN y C.M. AUSTIN Synop. Synopsis of biological data on the Westem rock lobster Panulirus cygnus George FAO fihs 1980b.1-64.128:1962. PROVENZANO, A.J. Proc. Gulf Caribb. Fish. Inst. Recent experiments on the laboratory rearing of the tropical lobster larvae.1968. 152-157. 21: ROBERTSON, P.B. Bull. Mar. Sci. The complete larval development of the sand lobster, Scyllarus americanus (Smith), (Decapoda, Scyllaridae) in the laboratory, with notes on larvae from the plankton. 1968.294-342. (2): 18 SUTCLIFFE, W.H. Ecology Notes on the biology of the spiny lobster Panulirus guttatus in Bermuda. 1953.794-796.(4): 34 YAÑEZ-ARANCIBIA, A., J. CURIEL-GOMEZ Y V.L. DE YAÑEZ An. Centro. Cienc. del Mar y Limnol. Univ. Nal. Autón. México Prospección biológica y ecológica del bagre marino Galeichthys caerulescens (Gunther) en el istema lagunar costero de Guerrero, México (Pisces: Ariidae) 1976.125-180(1): 3
|