|
CYTOGENETIC STUDY OF
Diplectrum eumelum Rosenblatt and Johnson, 1974 (PISCES: SERRANIDAE)
Trabajo recibido el 11 de enero de 1990 y aceptado Para su
publicación el 6 de marzo de 1990.
Alicia Luz Durán-González
Alfredo Laguarda-Figueras
Instituto de Ciencias del Mar y Limnología, Universidad Nacional
Autónoma de México. Contribución No. 674 del Instituto de
Ciencias del Mar y Limnología, UNAM. Trabajo presentado en el X Congreso
Nacional de Zoología, efectuado del 25-28 de Octubre de 1989 en Fac. de
Ciencias, UNAM, México D.F.
Tomando en cuenta la alta
complejidad taxonómica que presentan los Osteichthyes, en particular los
agrupados en la Familia Serranidae cuya diferenciación y
clasificación es difícil, el presente trabajo contribuye con el
análisis citotaxonómico de la especie Diplectrum
eumelum, para facilitar su caracterización. Los ejemplares
fueron colectados en tres cruceros oceanográficos (CAPECAL II, III y IV)
efectuados en las costas del Pacífico Mexicano durante
septiembre-octubre de 1984, 1985, 1986. Por medio de técnicas
citogenéticas convencionales, fueron procesados quince organismos, los
cuales presentaron un número diploide de 48 cromosomas, cuya
clasificación permitió caracterizar a un par metacéntrico,
dos pares submetacéntricos y 21 pares acrocéntricos, sin
presencia de cromosomas sexuales. El número haploide (n) de esta especie
es 24 y el número fundamental es n. f. = 54, en virtud de que presenta
21 elementos monorrámeos y tres birrámeos. El cariotipo difiere
del patrón de 48 cromosomas acrocéntricos que se considera
ancestral, por la presencia de los tres pares birrámeos. De acuerdo a
los antecedentes citogenéticos de la familia Serranidae, lo anterior
representa una valiosa ayuda para diferenciar a las especies que la
integran.
Taking in lo account the high taxonomic
complexity of the Osteichthyes, in particular those of Serranidae family whose
classification are difficult, the present work contributes with the
cytotaxonomic analysis ofDiplectrum eumelum in order to make
easier the characterization and philogenetic location of this biologic entity.
The studied specimens were colected during the oceanographic campaigns (CAPECAL
II, III and IV) that took part in the coast of the Mexican Pacific Ocean during
september - october of 1984,1985 and 1986. Fifteen organisms were processed by
conventional cytogenetic technique, which shows a diploid number of 2n = 48.
The chromosomes are classified as one metacentric pair, two submetacentric
pairs and 21 acrocentric pairs. No sexual chromosomes were identified. The
haploid number is n = 24 and the major chromosome arm number is n. f. = 54,
because it shows 21 monoarmed and 3 biarmed elements. The karyotype differs of
the 48 acrocentric chromosomes pattern which is considerated ancestral, by the
presence of 3 biarmed pairs. According with the cytogenetics antecedent of the
Serranidae family, this is an important criterion that presents a valuable help
in order to differentiate the species which form this taxonomic
entity.
The classification into the Serranidae Family is clearly insatisfactory, owing this to the similar characteristics presented by the organisms which are considered properly Serranidae and their close relatives, in such a manner that the location in the different genera is based more frequently in the absence of common specific characteristics than to the presence of basic similarities (Smith, 1965). The Serranidae is one of the bigger and diversified families of the teleost fishes. lt is located among other 64 families in the Percoid Superfamily (Nelson, 1984); nevertheless, these families show general characteristics that make difficult lo stablish the limits among them. In general it is located in the base of the evolution of Percoidei Suborder and it has even been used as a "lay aside" taxonomical entity which includes near 35 genera and about 370 species: all the lower percoidei poorly envolved fishes (Gosline, 1966 and Nelson, 1984). For this reason the organisms which integrate the Percoidei Suborder have been subjet of various systematic reviewings in order to get a solution to this complex problem (Gosline, 1966, Smith, 1971, Kendall, 1979, Randall 1980). The Diplectrum genus (Holbrook: 1855 - 32) which has been very well studied, reviewed and characterized by Bortone (1977) and previously by Rosenblatt and Johnson (1974) is located in the Serranidae Family. There has been a great confusion in the literature in order to located the organisms and the different species, taking in to account the evident morphoanatomic and meristic homogenity, and the consequent need to use characters submited to wide variation as coloring (of fin nose, operculum, and caudal pedunculus), number of scales (on cheeks and lateral lines), gillspines and pseudogill filaments; a mistake in the "weight" given to this characteristic may result, in consequence, in an erroneous determination of the species. However, it is considered that the Diplectrum genus is formed by nine endemic species in the Pacific Ocean East Coast and three in the Atlantic Ocean West Coast, (tropical or subtropical). Its species are hermaphrodites and in some cases the authors propose the recognizing of subspecies as in the case of D. formosum(Bortone, 1977 and Darcy, 1985). Due to the differences between the species are very subtle, and in some cases the distinctive characters are overlapping, the objective of the present study it is to stablish the karyotype ofDiplectrum eumelum contributing to its characterization, in order to help to define the systematic location of the group and the Serranidae Family into the evolutionary context. COLLECTING AREAThe studied organisms were collected and processed on board of the B/O "El Puma" vessel during the oceanographic campaigns CAPECAL II, III and IV, which took place in september - october of 1984, 85 and 86, respectively. The specimens were caught in the Pacific Ocean area located between the 28°19.7'-21°18.4' North Latitude and the 105°32.5'-111°34.8' West Longitude (Fig. 1). MATERIAL AND METHODSFifteen hermaphrodites organisms were processed by the cytogenetical techniques for gill epithelium which were established by McPhail and Jones (1966), Denton (1973) and Kligerman and Bloom (1977). The metaphases which showed a complete chromosomic complement, high definition of chromosomes and adequate concentration of chromatine (not very high) were chosen in order to stablish the karyotype. Those fields with overlapping chromosomes were omited. The chromosomes were classificated by the methods proposed by Levan et al. (1964) and Al -Aish (1969). RESULTSA number of 98 metaphases with different levels of chromosome spiralization were obtained. In all cases they showed a diploid number (2n) of 48 chromosomes. Chromosomic heteromorphism or negative heteropygnosis which makes evident the presence of sexual chromosomes, was not found (Fig. 2). Likewise, 18 mitotic fields obtained from organisms presented the desired characteristics for their analysis, showed two pairs of submetacentric, one pair of metacentric and 21 pairs of telocentric (acrocentric) chromosomes (Tabla 1). The karyotype and Idiogram of Diplectrum eumelum are characterized by: a diploid number (2n) of 48 chromosomes, a haploid number (n) of 24, a Karyotype formula of 2sm + 1 m + 21a and major chromosome arm number of 54 because three of its elements are biarmed an 21 monoarmed (Figs. 3 and 4). 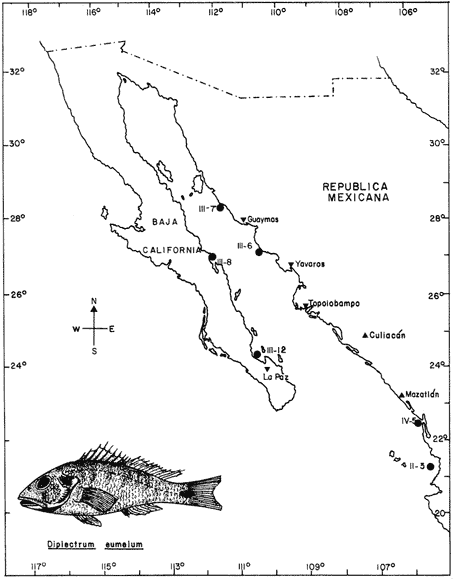 Figure 1. Collecting sites of Diplectrum eumelum corresponding to Oceanographic campains CAPECAL II, III and IV. 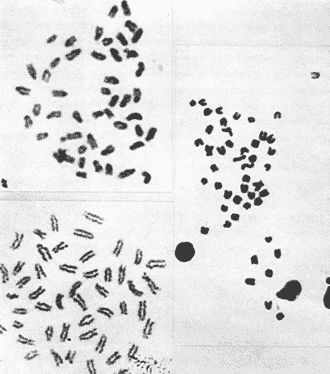 Figure 2. Mitotic Fields of Diplectrum eumelum (2n=48). 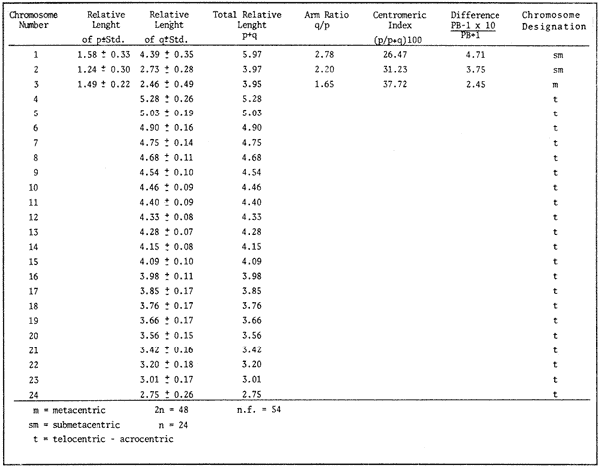 TABLE 1 RESULTS OF THE STATISTICAL ANALYSIS OF THE KARYOTYPES CHROMOSOMES OF Diplectrum eumelum 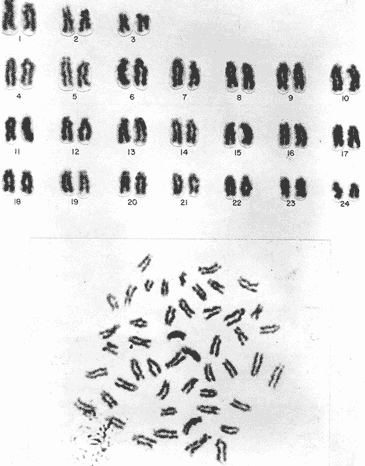 Figure 3. Karyotype of Diplectrum eumelum Rosenblatt and Johnson, 1974. 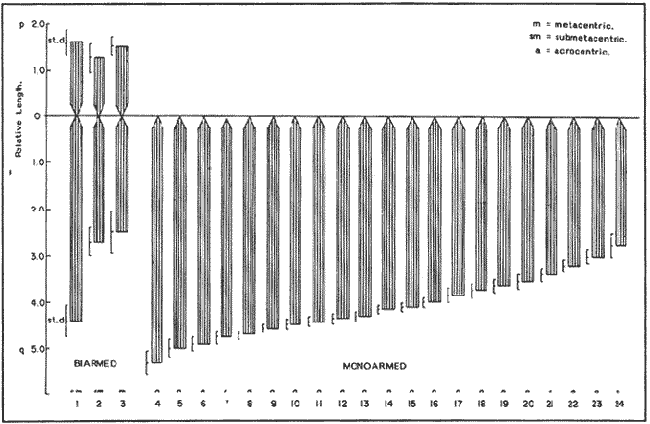 Figure 4. Idiogram of the Karyotype of Diplectrum eumelum. DISCUSSIONTaking in to account that the biarmed elements (metacentric and submetacentric) come from the monoarmed chromosomes (acrocentric: telocentric), several authors (Ohno et al., 1968, Denton, 1973, Thompson, 1979 and Uyeno et al., 1983) consider that the karyotype of the ancestral teleost fishes was of 48 achrocentric chromosomes. The antecedents about cytogenetic: works of the perciform fishes are scarce. The most studied families from this point of view are the Centrarchidae (23 species) and Cichlidae (41 species) but only one species has been studied from the Serranidae Family (Denton, 1973, Thompson 1979). The diversity of the Serranidae Family has made difficult its study, nevertheless it is very interesting from the taxonomic and evolutionary points of view. As an example, the genusHypoplectrus forms a group of 12 hermaphroditic: morphospecies which are sinchronic in the new world and differ among them in the patterrn of coloring but a little in morphometry, meristic, ecology or reproductive behavior. For this reason Graves and Rosenblatt (1980) made the electrophoretic analysis of seven different kinds of colored fishes of this genus located in the same reef: the conclusion was that there are not enough arguments in order to recognize more than one species, because it is a clear example of a polimorphic population; notwithstanding that at the same time Fischer (1980), by means of this "SCUBA" observations, conclude that it was a case of an multispecific complex (Nelson, 1984). From this perspective, the morphoanatomic homogenity of the genusDiplectrum is not a "hig weight" to locate the organisms in different species. For this reason more analysis is needed, taking to account that there are few precise characters in order to achieve a correct taxonomic determination, besides the interesting reviews of Bortone (1977) and Rosenblatt and Johnson (1974) related to Diplectrum eumelum as a recent taxonomic species. Some research in the Gulf of California and the Pacific Ocean done by Castro - Aguirre et al. (1970), Heiden (1985) and Pérez -Mellado and Findley (1985), show the variations which can be present in the species composition of Diplectrum genus, and thus the evident need for defining these taxonomical entities. According to the classification of the Serranidae family and its sexual patterns, which were analized by Smith (1965) and Smith and Young (1966), the Serranus genus is considered one of the most primitive of the sinchronic hermaphroditic Serranidae as well as theHypoplectrus and Diplectrumgenera. There are only two cytogenetic antecedents of the Serranidae Family which are the Centropristes ocyurus study with a diploid number of 48 chromosomes: 28 metacentric and 20 submetacentric (Durán-González and Laguarda-Figueras, 1987) and theCoreoperca kawamebari studys with 48 acrocentric chromosomes (Nogusa, 1960 and Denton, 1973). In the present time some genera of this Family are in an uncertain taxonomical location. The Coreopercagenus, was located in the Perichthydae Family by Gosline (1966), which is still subjects to change in its composition (Nelson, 1984). Apart from the results obtained in the Diplectrum eumelum study which is characterized by a diploid number of 48 chromosomes, 4 submetacentric, 2 metacentric and 42 acrocentric, there is information of the karyotype ofSerranus phoebe that shows 48 acrocentric chromosomes (study in progress). The results are similar to those obtained by Smith and Young (1966) through their analysis of the Serranidae patterns of sexuality. The Centropristes genus has its own morphologic characteristics, such as the shape of the caudal fin, the pelvic fins and an accentuated sexual dimorphism. Notwithstanding, it is a protogenic hermaphrodite whose skull morphology presents certain similarities that suggest affinity with Paralabrax, a secondary gonocoric. According to this, it is considered that both come from a common ancestral and that difference between their sexual mechanisms makes evident that they must be recognized as separated genera (Smith and Young, 1966). Also, the cytogenetic antecedents of Centropristes ocyurus show a karyotypic evolved genus by the presence of its biarmed chromosomes. According to their cytogenetic and karyotypic antecedents, these genera can be located from the most to the less evolved, being ordered as follows: Serranus, Diplectrum andCentropristes. ConclusionesThe karyotype of Diplectrum eumelum is characterized by a diploid number of 2n = 48 (n = 24). The analysis of the complement shows the existence of one metacentric, 2 submetacentric and 21 acrocentric pairs, with a mayor chromosome arm number of 54 (n. f. = 54). In accordance with the cytogenetic studies, it is suggested that in the Serranidae family the Diplectrum genus is less evolved than Centropristes and more evolved thanSerranus. Taking in to account the significant specialization of the reproductive systems presented by the Serranidae family, the cytogenetic and electrophoretic studies are very important to know which differences are significant in order to get a better taxonomic ordaining. AgradecimientosThe authors are grateful to Dr. Antonio García-Cubas and to M. in C. Felipe Flores, from the "Instituto de Ciencias del Mar y Limnología (ICMyL)" of the "Universidad Nacional Autónoma de México (UNAM)" for the help they gave us in order to participate in the oceanographic campaigns CAPECAL. In like manner to the M. in C. Margarito Alvarez, from the ICMyL, and to the M. in C. Patricia Fuentes and the Biól. Héctor Espinosa from the "Instituto de Biología" of UNAM for the valuable help in the classification of the biologic material. LITERATURAAL-AISH, M. Can. J. Genet. Cytol.Human chromosome morphology. I. Studies on normal chromosomes characterization and karyotyping. 1969. 370-381.11: BORTONE, S.A. Revision of the sea basses of the genus Diplectrum (PISCES: Serranidae). U.S. Dep. Commer., NOAA Tech. Rep. NMFS Circ 1977.49 p.404: CASTRO-AGUIRRE, J.L., J. ARVIZU M. y J. PAEZ B. Rev. Soc. Mex. Hist. Nat. Contribución al conocimiento de los peces del Golfo de California.1970.107-181.31: DARCY, G.H. Synopsis of Biological Data on the Sand Perch, Diplectrum formosum (Pisces: Serranidae). U.S. Dep. Commer., NOAA Tech. Rep. NMFS 26. FAO Fisheries Synopsis No. 143:1985. 21 p. DENTON, T.E. Fish Chromosome. Methodology. Charles C. Thomas Publisher U.S.A.1973. 466 p. DURÁN-GONZÁLEZ, A.L. y A. LAGUARDA-FIGUERAS Res. IX. Cong. Nal. de Zool. Estudio citogenético de Centropristes ocyurus (Jordan y Evermann) (Pisces: Serranidae). VillahermosaTabasco. 1987. No. 18.Octubre. Res. FISCHER, J.E.Copeia Speciation in the hamlets (Hypoplectrus: Serranidae) - a continuing enigma. 1980.649-659.(4): 1980 GOSLINE, W.A. Proc. Cal. Acad. Sci. The limits of the fish family Serranidae, with notes On other lower percoids. 4 th series. 1966. 91-112.33: GRAVES, J.E. and R.H. ROSENBLATT. Evolution Genetic relationships of the Color morphs of the serranid fish Hypoplectrus unicolor1980. 240 - 245. 34: HEIDEN, VAN DER, A.M. Recursos pesqueros potenciales de México: La pesca acompañante del camarón. Taxonomía, biología y evaluación de la ictiofauna demersal del Golfo de California In Yáñez-Arancibia, A. (Ed.) Prog. Univ. de Alimentos, Inst. Cienc. del Mar y Limnol., Inst. Nal. de Pesca. UNAMD.F.México1985. Cap. 4, 149-200. 748 p. KENDALL, A.W. Jr.Morphological Comparisons of North American Sea Bass Larvae (Pisces: Seranidae). U.S. Dep. Comer NOAA Tech. Rep. NMFS Circ.1979. 50 p. 428: KLIGERMAN, A.D. and S.E. BLOOM. J. Fish. Res. Board Can. Rapid chromosome preparations from solid tissues of fishes. 1977. 266-269.34: LEVAN, A., K. FREDGA and A.A. SANDBERG. Hereditas Nomenclature for centromeric position on chromosomes.1964. 201 - 220. 52: Mc PHAIL, J.D. and JONES, R.L. J. Fish. Res. Bd. Can.A simple technique for obtaining chromosomes from teleost fishes. 1966.23: 5 NELSON, J.S. Fishes of the world. Wiley Interscience. 2a Ed.U.S.A. 1984. 416 p. NOGUSA, S. Memoirs of the Hyogo Univ. of AgricultureA comparative study of the chromosomes in fishes with particular considerations on taxonomy and evolution.1960.1.(1): 3 OHNO, S., U. WOLF and N.B. ATKIN. HereditasEvolution from fish to mammals by gene duplication. 1968.169-187.(6): 59 PÉREZ-MELLADO, J. y L.T. FINDLEY. Recursos Pesqueros potenciales de México: La pesca acompañante del camarón Evaluación de la ictiofauna acompañante del camarón capturado en las costas de Sonora y Norte de Sinaloa, México In; Yáñez-Arancibia, A. (Ed.). Prog. Univ. de Alimentos, Inst. Ciene. del Mar y Limnol., Inst. Nal. de Pesca. UNAMD.F.México 1985. Cap. 5: 201 - 254, 748 p. RANDALL, J.E.Micrones Revision of the genus Plectranthias (Serranidae: Anthiinae) with descriptions of 13 now species. 1980.101-187.16: ROSENBLATT, R.H. and G.D. JOHNSON. Calif Fish and Game Two new species of sea basses of the genus Diplectrum with a key to the Pacific species. 1974.178-191.(4): 60 SMITH, C.L. Amer. Museum The patterns of sexuality and the classification of serranid fishes.NOVITATES 1965. 1-20.2207: SMITH, C.L. Bull. Am. Mus. Nat. Hist. A revision of the American grouper: Epinephelus and Allied genera.1971.69-241.146: SMITH. C.L and P.H. YOUNG. Calif Fish and Game Gonad structure and lile reproductive Cicle of the kelp bass, Paralabrax clathratus (Girard), with comments on the relationships of the serranid genus Paralabrax. 1966. 283- 292. (4): 52 THOMPSON, K.W. Copeia Cytotaxonomy of 41 species of Neotropical Cichlidae.1979. 679 - 691. (4): UYENO, T., R.R. MILLER and J.M. FITZSIMONS CopeiaKaryology of Cyprinodontoid fishes of the mexican family Goodeidae. 1983.497-510(2)
|