|
ANÁLISIS DEL
RÉGIMEN ALIMENTARIO DE TRES POBLACIONES DE LA ANCHOVETA Engraulis mordax
Girard (Pisces: Engraulidae), EN EL NORTE DE BAJA CALIFORNIA
Trabajo
recibido el 25 de agosto de 1989 y aceptado para su publicación el 16 de
noviembre de 1989.
Xavier Chiappa-Carrara
Manuel Gallardo-Cabello
Mercedes Jacob-Cervantes
Instituto de Ciencias del Mar y
Limnología, Universidad Nacional Autónoma de México.
Contribución No. 665 del Instituto de Ciencias del Mar y
Limnología, UNAM.
En el presente
trabajo se determina la dieta alimentaria de los ejemplares de tres poblaciones
de la anchoveta E. mordax en el norte de Baja California. Asimismo, se
analizaron las variaciones del régimen alimentario en relación a
la latitud, la edad, la periodicidad estacional y las épocas de las
surgencias, con especial referencia a la nutrición, el crecimiento y la
reproducción. La proporción de sexos encontrada es muy
próxima a un macho por cada hembra y la época de desove ocurre
durante la primavera y el otoño. Las diatomeas, los dinoflagelados, los
protozoarios, los crustáceos y los huevecillos son los principales
grupos alimentarios de la anchoveta. En las poblaciones analizadas, a medida
que decrece la latitud, aumenta el número de especies en el contenido
estomacal. E. mordax es un consumidor primario durante las primeras etapas de
su desarrollo (edades "0" a "I" años ). Posteriormente, se comporta como
un consumidor secundario (edades "II" a "VII" años). El requerimiento
calórico específico para una anchoveta con un cielo de vida de
siete años es de 1670 Kcal. Los periodos de más activa
alimentación ocurren en el verano, épocas en que aumenta tanto la
intensidad de las surgencias, como los índices de diversidad de la
dieta. Existe una marcada relación entre el desgaste corporal y la
freza, conforme tiene lugar el desove el peso de la anchoveta disminuye
considerablemente.
The diet of three
populations of E. mordax off west Baja California coasts was determined.
Dietary variations referred to latitude, age, seasonal periodicity and
upwelling periods, specially relate to nutrition, growth and reproduction, were
also analyzed. Sex ratio was nearly one-to-one, spawning occurring throughout
the year, with peaks during spring and autumn. Diatoms, dinaflagellates,
protozoarians, crustaceans and E. mordaxeggs are the most
important dietary components of northern anchovy. Diversity indexes of the
preys in the guts are greater in southem than in northern areas. In terms of
numbers of items per prey category, phytoplankton is the grealer contributor to
the diet of the smallest E. mordax examined (age groups "0" and "I"), but as
fish growth ("II" to "VII" years) zooplankton preys replace the former as the
major component of the diet. Along a seven years life history, northern anchovy
requires 1670 Kcal for maintenance, growth and reproduction. Most active
feeding activity was identified to be during summer, due to the increase of
upwelling intensity as well as diversity indexes of the diet. There is a strong
relationship between body weight and reproduction. While spawning, anchovy
weight decreases.
La interacción entre factores positivos - como son el crecimiento y el reclutamiento - y negativos - como la mortalidad natural y por pesca, mantiene un equilibrio y produce el rendimiento de una población sometida a explotación pesquera (Gallardo- Cabello, 1985). Así, en el estudio de las poblaciones marinas el factor de su evolución en el tiempo es el que más interesa conocer para administrar adecuadamente los recursos. Dicho factor se da por medio de las interacciones de los distintos elementos que conforman el sistema dinámico, tanto biológicamente - la depredación y la competencia - como en el medio exterior - la disponibilidad del alimento y las variaciones de los parámetros físicos-químicos. Dichos elementos, constituyen la estructura del sistema y determinan su comportamiento dinámico a través del tiempo. De esta manera, se puede decir que la interacción entre los componentes de un sistema tiene más importancia en su persistencia, que cada una de las partes que lo conforman. Por lo tanto, para el manejo adecuado de los recursos pesqueros, es necesario obtener información sobre la estructura de la transferencia energética y los cambias tróficos que sufre un ecosistema. De esta manera, es posible concluir que el análisis del régimen alimentario de una especie es un factor fundamental en el estudio de la dinámica de las poblaciones. Las características de este elemento estarán representadas por una serie de parámetros matemáticos a los cuales se deberá recurrir para analizar su efecto sobre la nutrición, el crecimiento y la reproducción en relación con las demás fuerzas, que intervienen en la dinámica trófica. De acuerdo con lo anterior, la recolección de los datos para esta clase de estudios, conduce a diferentes fases fundamentales, una de ellas consiste en el análisis de la alimentación. A pesar de la importancia ecológica y económica que tiene la anchovetaEngraulis mordax en el sistema de la corriente de California (Chiappa-Carrara, 1988), se han realizado pocos trabajos sobre la ecología trófica de los juveniles y de los adultos de esta especie (Leong y O'Connell, 1969; Loukashkin, 1970; O'Connell, 1972).
Por lo tanto, en el
presente trabajo se emplearon los contenidos estomacales de los ejemplares de
tres poblaciones de la anchovetaEugraulis mordax
Los
límites de las poblaciones consideradas en el presente trabajo, son las
establecidas por Chiappa - Carrara (.1988) y Chiappa - Carrara y Gallardo -
Cabello (en prensa; poblaciones "1", "2" y "3", figura 1).
MATERIAL Y METODOSLos datos biológicos fueron obtenidos de un total de 960 ejemplares de la anchoveta Engraulis mordax, provenientes de los muestreos realizados en la región que se localiza frente a las costas de Baja California, desde las Islas Coronado hasta Cabo San Quintín. Las muestras de E. mordax son producto de la captura comercial de los B/P. "Calafia", "Ciprés", "Concepción", "Kino" "Libertador del Sur", "Loreto", "Sauzal", "Tortugas" y "Vizcaíno", pertenecientes a la Compaña Pesquera Zapata, S.A. de C.V. El intervalo de tiempo que abarcan las muestras analizadas comprende desde enero hasta octubre de 1986, y desde enero hasta junio de 1987. Durante los meses de noviembre y diciembre de 1986 las poblaciones de la especie en estudio no fueron capturadas por la flota comercial, debido a los periodos de la veda. Los ejemplares fueron separados de acuerdo con las localidades en que fueron capturados. Se obtuvieron los tractos digestivos, cortando desde el inicio del esófago hasta el final del intestino. Las vísceras fueron guardadas individualmente, fijándolas en formol neutralizado al 4% (Steedman, 1976) y conservándolas, posteriormente, en alcohol al 70%, señalando además, la longitud estándar y el peso de cada organismo. El sexo y los estadios de madurez gonádica se determinaron in visu siguiendo los criterios de Nikolsky (1963), modificados por Chiappa-Carrara (1988). Se examinó el contenido del estómago cardíaco y pilórico, así como el del esófago. Se descartaron los intestinos de los organismos para reducir el error debido a las diferentes tasas de digestión del alimento (Gannon, 1976; Berg, 1979; Hyslop, 1980). Asimismo, no se consideró el peso del estómago, debido a la desviación que se presenta en la obtención de este parámetro, dada la cantidad de jugos gastricos existentes en el tracto digestivo (Thomas y Davidson, 1962). Por otra parte, se examinó el contenido de cada estómago empleándose una lupa binocular estercoscópica y un microscopio óptico. Todos los items del contenido fueron identificados hasta el menor nivel taxonómico posible, utilizando las claves de Cupp (1943; 1950), Tregouboff y Rose (1957) y Rose (1975).  Figura 1. Localización del área de estudio, mostrando los límites geográficos de las poblaciones analizadas. A partir del conteo de los diferentes componentes dietéticos, se calcularon los índices de frecuencia de ocurrencia de cada grupo (Hynes, 1950; Hyslop, 1980), expresados como el porcentaje de estómagos que contienen dicho componente con respecto del total. Asimismo, se calcularon los porcentajes numéricos de las presas (Windell y Bowen, 1978). Los análisis de la alimentación con respecto a los distintos grupos de edad, se llevaron a cabo empleando las relaciones sobre la edad-longitud encontradas por Chiappa-Carrara y Gallardo-Cabello (en prensa). Por otra parte, se relacionó el régimen alimenticio de esta especie con las épocas y las intensidades de las sugerencias, estimadas a partir de los índices de Bakun (1973), para las tres zonas de estudio, completando el ciclo anual considerado en este trabajo. Los cálculos, se realizaron por estación climática para las latitudes y longitudes de 33ºN, 119ºW; 30ºN, 119ºW y 27ºN, 116ºW. De la misma manera, se calcularon los índices de diversidad de la dieta tanto mensual como estacionalmente, utilizando la expresión de Levins (1968). Con los datos biométricos obtenidos se determinó la relación entre la alimentación, el crecimiento y la nutrición, utilizando el índice del factor de condición de Fulton (1902). Finalmente, para estimar el requerimiento calórico teórico, se aplicaron los índices de las eficiencias de conversión calórica (Pauly, 1986) a los modelos del crecimiento desarrollados por Gallardo-Cabello y Chiappa-Carrara (1990). RESULTADOS Y DISCUSIÓN
REPRODUCCIÓNLa reproducción de una especie es uno de los principales factores aditivos a la misma población, proporcionando información sobre el reclutamiento. El estudio del ciclo de madurez gonádica y su relación con los períodos del crecimiento y las épocas de mayor alimentación, aportan conocimientos valiosos acerca de las adaptaciones de la especie en relación a su entorno. PROPORCIÓN DE SEXOSDe 960 ejemplares pertenecientes a tallas de 85 a 140 mm de longitud estándar, 454 fueron identificados como machos, 504 como hembras y 2 cuyo sexo no pudo ser determinado, y que correspondían a individuos jóvenes en estadio virgen. En la Tabla 1, se muestra la proporción de sexos encontrada. En todos los casos, se observa que dicha proporción es muy próxima a 1:1.  PROPORCIÓN DE SEXOS PARA TRES POBLACIONES DE LA ANCHOVETA Engrandis mordax En la anchoveta E. mordax, las gónadas pueden ser diferenciadas sexualmente "a simple vista" cuando los ejemplares han alcanzado los 85 mm de longitud estándar. La madurez sexual de esta especie se observa antes de cumplir el primer año de edad. CICLO DE MADUREZ GONÁDICALa determinación del estado de madurez gonádica, se realizó en base a la escala propuesta por Nikolsky (1963), modificándola de acuerdo al análisis realizado sobre las gónadas de la anchoveta, resultando: Estadio II : Unión de los estadios I y II de la escala de Nikolsky (1963). Reposo (utilizado también para designar a los individuos vírgenes). Apariencia de las gónadas: Estas se presentan pequeñas y filamentosas, adheridas entre la vejiga natatoria y el eje vertebral en la zona media de la cavidad visceral. Se encuentran protegidas por una envoltura adiposa. Son translúcidas y la irrigación sanguínea no es visible. En el caso de los ovarios, éstos son finos y redondeados, con una leve coloración rosácea; ovocitos no visibles a simple vista. Testículos finos, aplanados y blanquecinos. Estadio III: Inicio de la maduración virginal (en individuos en los que se activan por primera vez las gónadas) o recuperación (postdesove). Gónadas de mayor tamaño en relación al estadio anterior. Ocupan aproximadamente la mitad de la cavidad visceral. Translúcidas en las hembras, opacas en los machos. Los ovarios en maduración virginal, presentan una coloración rosácea, tejido granuloso e irrigación sanguínea poco manifiesta. Los ovarios en recuperación, presentan, además de las características antes mencionadas, una irrigación sanguínea más evidente y un volumen mayor. Los testículos son blanquecinos, con escasa vascularización. Estadio IV: Maduración (estadios IV y V de Nikolsky). Gónadas voluminosas que ocupan hasta las 2/3 partes de la cavidad visceral. La irrigación sanguínea es muy visible, abundante y ramificada. Los ovarios son grandes y de color amarillento-rojizo, muy irrigados. Los ovocitos son opacos, visibles a simple vista y presentan tejido granuloso. Los testículos son blanco-cremoso, ligeramente vascularizados. Semen denso. Estadio V DesoveGónadas que ocupan un volumen mayor al correspondiente a las 2/3 partes de la cavidad visceral, cubriendo parcialmente al intestino en la parte ventral. Irrigación abundante y capilares muy ramificados. Ovarios frágiles y muy vascularizados; ovocitos translúcidos y visibles a simple vista. Los testículos son blandos y presentan una abundante irrigación sanguínea en la región caudal. El semen es fluido. Resumiendo, los estadios de madurez gonádica considerados en este trabajo son los siguientes:  En la Tabla 2, se presentan los resultados obtenidos con respecto a las frecuencias mensuales de los distintos estadios de madurez gonádica y en la Tabla 3, se muestran las frecuencias porcentuales de dichos estadios, en relación a la estación climática. En términos generales, se observa que durante el invierno aumentan considerablemente los individuos en estadio II (reposo), seguidos en menor grado por aquellos en el inicio de la maduración. Durante los meses correspondientes a la primavera, aumenta la frecuencia de organismos desovantes. Así, en el verano (véase principalmente el caso de 1987, zonas 2 y 3), predominan los individuos en recuperación o postdesove (estadio III). Este estadio, se prolonga hasta el mes de septiembre, en el que comienza la maduración, continuándose hasta finales del otoño, en el que se presenta otro desove. Por lo tanto, el desove masivo de la anchoveta, se realiza en dos temporadas. Estas son la primavera y el otoño. Lo anterior coincide con las observaciones realizadas por Tapia-Vázquezet al. (1988) y por Parrish et al. (en prensa). RÉGIMEN ALIMENTARIO
Variaciones de la dieta con respecto a la latitud.Los principales grupos de presas que constituyen la alimentación de la anchoveta, así como los porcentajes numéricos obtenidos, se presentan en la figura 2. Es importante notar, que la importancia numérica relativa de las diatomeas, en las tres zonas, es considerable. En la zona 2, se observan las mayores proporciones, seguida por la 3 y la 1, en orden decreciente. Siguen en importancia los dinoflagelados, que junto con las diatomeas, ocupan cerca del 50% del total de individuos en la dieta. Los crustáceos y otros organismos del zooplancton (larvas de peces, moluscos y poliquetos), ocupan el 13.1% del alimento en la zona 1, el 11 % en la 2 y el 17.4% en la 3. Asimismo, los radiolarios representan el 12.3%, 5.4% y 8.3% en el sentido norte-sur de las zonas. Similarmente, los huevecillos de E. mordax, mostraron valores de 11.2%, 16.4% y 7.7%, respectivamente. El material orgánico presente no identificable, fluctúa entre valores volumétricos de 6.2% (zona 2), 13.3% (zona 1) y 16.4% (zona 3). En la Tabla 4, puede observarse que entre las zonas de estudio no se encuentran diferencias sustanciales entre los porcentajes numéricos de los principales grupos alimenticios de la anchoveta. Sin embargo, existe dentro de cada grupo taxonómico, una variación significativa en el número de especies que los conforman (Chiappa-Carrara, 1988). En las Tablas 5, 6 y 7, se muestran, tanto el número como el porcentaje de las presas para las tres zonas consideradas. Es importante notar que conforme decrece la latitud, aumenta el número de especies en relación a las zonas, de la manera siguiente: existe un aumento en el número de especies de diatomeas (5 especies en la zona 1, 6 en la zona 2 y 8 en la zona 3), aunque Coscinodiscus sp. domina en las tres zonas. Le siguen en importancia, en términos generales, Nitzschiasp,Rhizosolenia sp y Asterionella sp, respectivamente.  TABLA 2 FRECUENCIAS OBSERVADAS DE LOS ESTADOS DE MADUREZ GONÁDICA PARA TRES POBLACIONES DE LA ANCHOVETA Engraulis mordax, POR MES Y AÑO 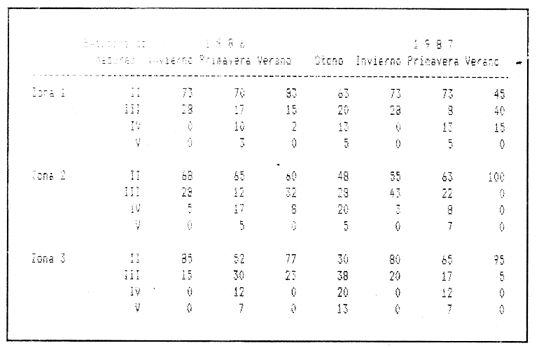 TABLA 3 PORCENTAJES ESTACIONALES DE LOS ESTADIOS DE MADUREZ GONÁDICA, EN TRES POBLACIONES DE LA ANCHOVETA Engraulis mordax (Años de 1986 y 1987)  Figura 2. Porcentajes de los grupos de presas de la anchoveta Engraulis mordax, para cada una de las áreas analizadas.  TABLA 4 VALORES NUMÉRICOS Y PORCENTUALES DE LOS PRINCIPALES GRUPOS DE PRESAS DE LA ANCHOVETA E. mordax, EN RELACIÓN A LAS ZONAS DE ESTUDIO  TABLA 5 VALORES NUMÉRICOS Y PORCENTUALES DE LAS PRESAS DE LA ANCHOVETA E. mordax, PARA LA ZONA 1  TABLA 6 VALORES NUMÉRICOS Y PORCENTUALES DE LAS PRESAS DE LA ANCHOVETA e. mordax PARA LA ZONA 2 En relación a los dinoflagelados, Ceratiumsp tiende a hacerse menos importante conforme decrece la latitud. Contrariamente,Dinophysis sp adquiere relevancia. Similarmente, el número de especies de radiolarios aumenta en relación a la disminución de la latitud, y se incrementan los grupos y familias que constituyen a los crustáceos. Asimismo, es importante señalar que en la zona 3 aparecen los moluscos (larvas Velliger). Régimen Alimentario y su Variación con Respecto a los Grupos de EdadEn las Tablas 8, 9 y 10, se presentan los resultados de los índices de frecuencia de ocurrencia, en relación a las clases de talla y en la figura 3, se muestran los porcentajes de los componentes alimenticios. La dieta presenta variaciones importantes a través de las tallas y las edades. Del análisis de las Tablas 8,9 y 10, se infiere que existe un cambio gradual de la alimentación, conforme el organismo alcanza tallas mayores. Así, la dieta de individuos de 81 a 85 mm de longitud total (edades de "0" y "I" años), está constituida principalmente por diatomeas, dinoflagelados, radiolarios y huevecillos. Por lo contrario, en organismos de tallas de 96 mm de longitud total y superiores a ésta (edades de "II" a "VII" años) aumenta el índice de frecuencia de ocurrencia de crustáceos y otros organismos del zooplancton, en relación a los demás componentes de la dieta. Analizando la figura 3, se demarcan diferencias notables en relación a la alimentación de los peces a diferentes edades. De esta manera, las diatomeas están representadas en todos los grupos de edad, en las tres zonas de estudio. Sin embargo, los porcentajes tienden a disminuir, conforme el pez crece. Contrariamente, los organismos pertenecientes al zooplancton (crustáceos y larvas de peces) aumentan su frecuencia en los peces de mayor longitud. Para el análisis de las relaciones existentes entre la alimentación y el crecimiento deEngraulis mordax durante las diversas etapas de su desarrollo, a continuación se muestran los valores promedio del factor de condición (Fulton, 1902), en relación a los grupos de edad. 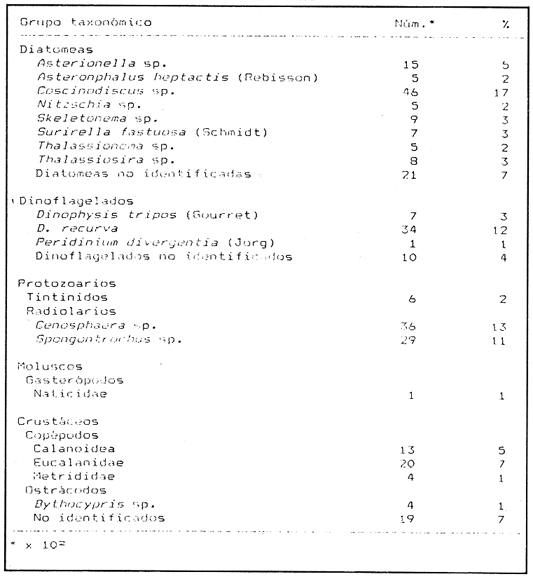 TABLA 7 VALORES NUMÉRICOS Y PORCENTUALES DE LAS PRESAS DE LA ANCHOVETA E. mordax PARA LA ZONA 3  TABLA 8 ÍNDICE DE FRECUENCIA DE OCURRENCIA DE LAS PRESAS, EN RELACIÓN A LAS CLASES DE TALLAS DE LA ANCHOVETA E. mordax, ZONA 1  TABLA 9 ÍNDICE DE FRECUENCIA DE OCURRENCIA DE LAS PRESAS, EN RELACIÓN A LAS CLASES DE TALLAS DE ANCHOVETA E. mordax, ZONA 2 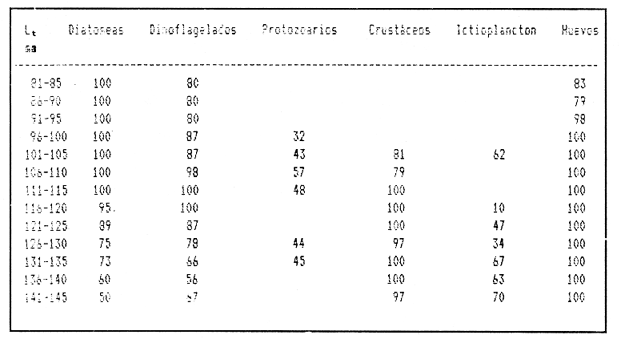 TABLA 10 ÍNDICE DE FRECUENCIA DE OCURRENCIA DE LAS PRESAS, EN RELACIÓN A LAS CLASES DE TALLAS DE LA ANCHOVETA E. mordax, ZONA 3  Figura 3. Porcentajes de los grupos alimenticios de la anchoveta E.mordax, en relación a la edad, en cada una de las áreas analizadas.  Se observa que los incrementos del factor de condición (^ ), disminuyen a través de la edad. Esto se debe a que el crecimiento en longitud de Engraulis mordax es muy rápido durante las primeras fases de su desarrollo (Chiappa-Carrara y Gallardo-Cabello, en prensa), con lo cual evita una depredación mayor por parte de otras especies, y obtiene una mayor capacidad de competencia interespecífica (Chiappa-Carrara, 1988). El fitoplancton ha sido considerado como la fuente principal de alimento de los engraúlidos (Ryther, 1969; Longhurst, 1971; Rojas de Mendiola, 1971; King y Macleod, 1976; Durbin, 1979; Walsh, 1981). Sin embargo, considerandolo anteriormente expuesto, puede concluirse que la anchoveta no es un consumidor de primer orden durante todas las etapas de su ontogenia. Los datos presentados en este trabajos así como los encontrados por James y Chiappa-Carrara (1989), indican que la anchoveta Engraulis mordax, durante la mayor parte de su ciclo de vida, es un consumidor secundario en las áreas de surgencia de la corriente de California. Lo anterior coincide con las observaciones de Angelescu (1982a; 1982b) y Angelescu y Anganuzzi (1986) para E. anchoita, de Koslow (1981) paraE. mordax y de James (1987) para E. capensis. Variaciones Estacionales en la Composición de la DietaEn la figura 4, se muestran las frecuencias porcentuales de ocurrencia de los principales grupos de alimento de la anchoveta Engraulis mordax, a lo largo del tiempo. De la misma, manera, en la figura 5, se presentan los porcentajes numéricos, en el mismo orden. Con respecto a las frecúencias porcentuales, se observa cierta constancia en la ocurrencia de las diatomeas y de los huevecillos durante todos los meses del año. Otros grupos, como el de los protozoarios, ocurre principalmente en los meses de primavera y principios del verano. Este mismo comportamiento que se registra para los crustáceos. Los dinoflagelados, siguen una tendencia parecida a la de las diatomeas, aunque con fluctuaciones más acentuadas. En lo referente a los porcentajes numéricos (Figura 5), se observa que los picos en las fluctuaciones de la diatomeas y de los dinoflagelados muestran una tendencia en la cual, durante los meses en que aumentan los valores de un grupo, disminuyen los del otro y viceversa. Los protozoarios, aunque presentan generalmente valores bajos, tienden a adquirir cierta importancia en junio. Los crustáceos son más numerosos durante los meses de primavera y de verano, después de los períodos del florecimiento de las diatomeas. En el caso de los huevos, su abundancia es mayor en febrero, abril y octubre. Estos últimos, son los meses del desove masivo de la anchoveta en esta área (Chiappa-Carrara, 1988).  Figura 4. Valores mensuales de las frecuencias de ocurrencia de los principales grupos alimenticios de la anchoveta E. mordax. A. Diatomeas, B. Dinoflagelados, C. Protozoarios, D. Crustáceos, D. Huevecillos de E. mordax.  Figura 5. Porcentajes numéricos mensuales de los principales grupos alimenticios de la anchoveta E. mordax. A. Diatomeas, B. Dinoflagelados, C. Protozoarios, D. Crustáceos, D. Huevecillos de E. mordax. Variaciónes en el Régimen Alimentario en Relación a las SurgenciasEn la Tabla 11, se muestra la relación entre los índices de surgencias de Bakun (1973), de la diversidad de la dieta (B) de Levins (1968) y de la relación peso-longitud (b) con respecto al ciclo estacional. De acuerdo con lo anterior, se observa que tanto los índices de las surgencias y de la diversidad de la dieta (B), como los del coeficiente (b), alcanzan sus más altos valores durante los meses del verano especialmente en junio. En cambio, los valores más bajos, se presentan durante el Invierno principalmente en enero. De acuerdo con lo anterior, se puede establecer que durante los meses de verano, la alimentación deE. mordax es más abundante y diversa, coincidiendo con los períodos de mayor afloramiento de nutrientes y de producción primaria. Mientras que durante los meses del invierno, al disminuir la intensidad de las surgencias, decrece la producción primaria y la alimentación de la anchoveta es menos intensa. ALIMENTACIÓN Y DESARROLLOLa eficiencia de conversión calórica del alimento obtenida fue del 12.5% lo que equivale a una tasa de consumo de alimento en el mar, de 348 cal/día. Asimismo, este valor corresponde al 3% del peso húmedo de la anchoveta, por día (Mikhman y Tomanovich, 1977). Los valores del requerimiento calórico con respecto a los grupos de edad de la anchoveta, se muestran en la Tabla 12. De acuerdo con lo anterior, se observa que los resultados obtenidos corresponden a una función exponencial, que tiende a ser asintótica alrededor de las 400 Kcal. Por lo tanto, el requerimiento calórico específico, expresado como el número total de calorías es de 1670.6 Kcal. para una anchoveta cuyo ciclo de vida es de siete años. En relación al alimento, esta energía corresponde, aproximadamente a 1500 g en peso húmedo, lo que equivale a que el alimento consumido anualmente por una anchoveta, es de 10 veces su peso, aproximadamente. Por otra parte, Hunter y Leong (1981), establecen que del requerimiento calórico total, la anchovetaE. mordax emplea el 5% para su crecimiento y el 8% para el desove. De acuerdo con lo anterior, en la figura 6, se muestran los valores del exponente (b) de la relación peso-longitud, de los índices de las surgencias de Bakun (1973) y de la diversidad de la dieta de Levins (1968), así como los porcentajes de los estados de madurez gonádica, en relación al ciclo estacional. Al respecto se observa, que durante el verano, los índices de Bakum y el coeficiente b alcanzan sus más altos valores, asociados con una mayor diversidad de la dieta. Es durante este período cuando la alimentación de los organismos de la población en general, y el crecimiento de los juveniles en particular, son mas intensos (Chiappa-Carrara y Gallardo-Cabello, en prensa). Durante el otoño, descienden los valores de los índices de surgencia, relacionados con una menor variedad y disponibilidad del alimento, por lo que el valor del coeficiente b, decrece. Este período está asociado, en los individuos adultos, con la época del desove, en que el desgaste corporal es muy grande y su peso disminuye considerablemente. En el invierno, los valores descienden en relación a los meses anteriores, debido a un decrecimiento de la productividad asociado a cambios en el fotoperíodo y en la intensidad de las surgencias.  TABLA 11 RELACIÓN ENTRE LOS ÍNDICES DE SURGENCIAS DE Bakun (1973), DE LA DIVERSIDAD DE LA DIETA (b) DE Levins (1968) Y DE LA RELACIÓN PESO-LONGITUD (b) DE LA ANCHOVETA E. mordax, EN FUNCIÓN DEL PERÍODO ESTACIONAL, AÑOS DE 1986 Y 1987 Durante la primavera, aumentan los valores de los índices de Bakun y de la diversidad de la dicta con respecto al invierno. Sin embargo, los valores del coeficiente b son los más bajos en relación a los demás períodos. Lo anterior es debido, fundamentalmente, a que en esta época aumenta la proporción de los individuos maduros, y se presenta el desove masivo en las poblaciones de la anchoveta, con lo cual la energía es canalizada, principalmente, hacía la actividad reproductora, disminuyendo notablemente el peso de los organismos.  TABLA 12 VALORES DE REQUERIMIENTO CALÓRICO TEÓRICO (Kcal) EN RELACIÓN A LOS GRUPOS DE EDAD Y AL PESO DE LA ANCHOVETA E. mordax  Figura 6. Variaciones estacionales del valor del coeficiente b de la relación peso-longitud, del índice de diversidad de la dieta (B) y de los porcentajes de las frecuencias de los estadios de madurez gonádica de la anchoveta Conclusiones- La proporción de sexos encontrada es muy próxima a 1: 1. - La madurez sexual es alcanzada por esta especie antes de cumplir el primer año de vida. - El desove masivo de la anchoveta se realiza durante la primavera y el otoño. - Las diatomeas, los dinoflagelados, los protozoarios, los crustáceos y los huevecillos son los principales grupos alimenticios de la anchoveta. - En relación a las tres zonas de estudio no se encontraron diferencias sustanciales, con respecto a los porcentajes numéricos de los grupos alimenticios de la anchoveta. Sin embargo, dentro de cada grupo taxonómico existe una variación significativa en el número de las especies que les conforman. A medida que decrece la latitud, aumenta el número de especies presentes en el contenido estomacal. - La dieta de E. mordax presenta variaciones importantes a través de la edad, existiendo así un cambio gradual en la alimentación. De esta manera, en individuos jóvenes está constituida principalmente por diatomeas, dinoflagelados, radiolarios y huevecillos. En organismos más grandes, aumentan las frecuencias de los crustáceos y otros organismos del zooplancton en relación a los demás componentes de la dieta. - Los incrementos en los valores del índice del factor de condición disminuyen a través de la edad. El crecimiento en longitud de esta especie es rápido durante las primeras fases del desarrollo. - La anchoveta E. mordax es un consumidor primario durante las primeras etapas de su desarrollo (edades "0" a "I" años). Posteriormente (edades de "II" "VII" años) se comporta como un consumidor secundario. - El requerimiento calórico específico, calculado para una anchoveta con un ciclo de vida de siete años, equivale a 1670 Kcal. - Los períodos de más activa alimentación ocurren durante los meses del verano, épocas en que son mayores los índices de las surgencias de Bakun y de la diversidad de la dieta de Levins. - Durante los meses de la primavera y del otoño, épocas en que la anchoveta lleva a cabo los desoves masivos, existe una marcada relación entre el desgaste corporal y la freza. Conforme tiene lugar el desove, el peso de los organismos disminuye considerablemente. LITERATURAANGELESCU, V. Zoología Neotropical.Ecología trófica de la anchoita del mar Argentino (Engraulidae: Engraulis anchoita). Parte 1: Morfología del sistema digestivo en relación con la alimentación. P.J. Salinas Actas VIII Congr. Latinoamer. Zool.MéridaVenezuela1982a1317-13502 ANGELESCU, V.Ecología trófica de la anchoita del mar Argentino (Engraulidae: Engraulis anchoita). Parte II: Alimentación, comportamiento y relaciones tróficas en el ecosistema. Contrib. Inst. Nac. Invest. Des. Pesq. Mar del Plata1982b83 p.409 ANGELESCU, V. y A. ANGANUZZIEcología trófica de la anchoita (Engraulidae: Engraulis anchoita) del mar Argentino. Parte III: Requerimiento trófico individual en relación con el crecimiento, ciclo sexual y las migraciones estacionales. Rev. Invest. Des. Pesq.1986194-2235 BAKUN, A. Coastal upwelling índices, west coast of North America, 1946-71.NOAA Tech. Rep. NMFS, SSRF-6711973103 p. BERG, J. Discussion of methods of invesfigating the food of fishes with reference to a preliminary study of the prey of Gobiasculus flavescens (Gobiidae).Mar. Biol.1979263-27350(3) CHIAPPA-CARRARA, X.Tesis Consideraciones bioecológicas sobre la alimentación, crecimiento, hábitos alimenticios y contenido calórico de la dieta de las poblaciones de la anchoyeta ENGRAULIS MORDAX Girard, localizadas en las costas occidentales de Baja California.UACPy-CCH-UNAM1988131p. CHIAPPA-CARRARA, X. y M. GALLARDO-CABELLO, Análisis de las frecuencias de tallas y de los otolitos sagitta para la determinación de la edad de tres poblaciones de la anchoveta EngrauKs mordax Girard (Pisces: Engraulidae), en Baja California Norte. en prensa. Biol. Trop. CUPP, E.E. Bull. Scpipps Marine plankton diatoms of the west coast of North America. Inst. Oceanogr.19431-2385(1) CUPP, E. E. Bull. Scripps Inst. Oceanogr. Marine planklon of the westem coast of North America.19501-2375 DURBIN, A. G.Predator-Prey Systems in Fisheries Management.Food selection by plankton feeding fishes. H. Clepper Sport Fishing Institute Washington D.C.1979203-218 FULTON, T. Rates of groyith of sea-fishes.Sci. Invest Fish. Div. Scot. Rept.19021-3720 GALLARDO-CABELLO, M. An. Inst Cienc. del Mar y Limnol. Determinación de la edad de la anchoveta Engraulis mordax Girard en aguas de Baja California Norte (Pisces: Engraulidae). Univ. Nal. Autón. México1985221-23412(1) GALLARDO-CABELLO, M. y X. CHIAPPA-CARRARA An. Inst. Cienc. del Mar y Limnol.Estudio de las variaciones regionales en el crecimiento, la longevidad y la mortalidad natural de la anchoveta Engraulis mordax Girard (Pisces: Engraulidae) en el norte de Baja California, durante los años de 1986-1987.Univ. Nal. Autón. México199017(1) GANNON, J. E. The effects of differential digestion rates of zooplankton by alewife, Alosa pseudoharengus, on determinations of selective feeding. Trans. Am. Fish. Soc.197689-95105(1) HUNTER. J.R. y R. LEONGFish. Bull. The spawning energetics of female northem anchovy, Engraudis mordax.1981215-23079(2) HYNES, H. B. N. The food of freshwater stickleebacks (Gasterosreus aculeatus and Pygosteus pungitius), with a review of melhods used in atudies of the food of fishes. J. Anim. Ecol.195035-3819 HYSLOP, E. J. Stomach contenta analysis. A review of methods and their application.J. Fish. Biol.1980411-42917 JAMES, A. G. Feeding ecology, diel and field-based studies on feeding selectivity of the Cape anchovy Engraulis capensis Gilchrist. S. Afr. J. Mar. Sci.1987673-6925 JAMES, A.G. y X. CHIAPPA-CARRARAA comparision of field based studies of the trophic ecology of Engraulis capensis Gilchrist and E. mordar Girard.Proc. 24 th. EMBS, , en prensa.ObaputU.K1989 KING, D.P.F. y P.R. MACLEODComparison of the food and the fillering of pitchard Sardinops ocellata and anchovy Engrauhs capensis off South West Africa, 1971-1972.Invest. Rep. Sea Fosh. Brch. S. Afr.197629 p.111 KOSLOW, J. A. Fish. Bull. Feeding selectivity of schools of northem anchovy, Engraulis mordair, in the southem Califomia Bight.1981131-14279(1) LEONG, R.C. y C.P. O'CONNELLA laboratory study of particule and filter feeding of the northem anchovy (Engraulis utorda). J. Fish. Res. Bd Canada1969557-58226 LEVINS, R. Evolution in changing environments. The theory of niche. R. H. Mac Arthur Princeton Univ. Press.1968 LONGHURST, A. R.Oceanography and Marine Biology. An annual Review 9The clupeoid resources of tropical seas. H. Bames George Alien and UnwinLondon1971349-385 LOUKASHKIN, A. S. On the diet and feeding behavior of the northern anchovy Engraulis mordax (Girard).Proc. of the Calif. Acad of Sciences1970419-45837(13) MIKHMAN, A.S. y L.V. TOMANOVICHThe feeding of the Azov anchovy, Engraulis encrasicholus moeoticus. J. Ichthyol, (Scripta Publish. Wash.)1977240-24417 NIKOLSKY, G. V. The Ecology of Fishes.Academie PressLondon-New York1963352 p. O´CONNELL, C. P.The interrelation of biting and filtering in the feeding activity of the northem anchovy (Engraulis mordax) J. Fish. Res. Bd. Canada1972285-29329(3) PARRISH, R.H., D.L. MALLICOATE, K.F. MAIS y R.A. KLINGBEIL, en prensa. Fish. Bull.Age dependent maduration stages, sex ratio, spawning frequency and fecundity in the northen anchovy, Engraulis mordax. PAULI, D. Fish. Bull. A simple method for estimating the food consumption of fish populations from growth data and food conversion experiments.1986827-84084(4) ROJAS DE MENDIOLA, B.Fertilily of the sea. Some observations on the feeding of the peruvian anchoveta Engraulis ringens J. in two regions of the peruvian coast. Gordon y Breach Science Publish.1971417-4402 ROSE, M. Manuel de plancionologie Mediterraneenne.Centre National de la Recherche ScientifiqueParis1975Tomos I y II. RYTHER, J. H.Science Photosynthesis and fish production in the sea.196972-76166 STEEDMAN, H. F.Monogmphs on Oceanographic Metbodology Zooplankton Fixation and Preservation. UNESCO1976350 p.4 TAPIA-VAZQUEZ, O., C.E. COTERO-ALTAMIRANO y C.M. GARCIA-CUELLARDeterminación de madurez gonadal y fecundidad en anchoveta Engraulis mordax de la subpoblación central.Ciencia Pesquera, Inst. Nal. de la Pesca, Sría. de Pesca.México198869-1011(6) THOMAS, H. J. y C. DÁVIDSON The food of the Norway lobster.Mar. Res.19625-153 TREGOUBOFF, G. y M. ROSEManuel de plactonologie Mediterraneénne. Centre National de la Recherche ScientifiqueParis1957Tomos I y II. WALSH, J. J. A carbon budget for overfishing off Peru. Nature, Long.1981300-304290 WINDELL, J.T. y S.H. BOWEN Methods for the Assesmment of Fish Production in Fresh Walers. Methods for study the fish diets based on analysis of stomach contents. T. BagnedOxford Blackwell Scientific Public.3era ed.1978219-226
|