|
PROTEÍNAS Y
CARBOHIDRATOS DE LA MATERIA ORGÁNICA PARTICULADA FOTOSINTÉTICA Y
NO FOTOSINTÉTICA EN LA COSTA OCCIDENTAL DE BAJA CALIFORNIA SUR,
MÉXICO
Trabajo recibido el 23 de agosto de 1989 y aceptado para su
publicaci6n el 11 de diciembre de 1989.
Carlos H. Lechuga-Devèze
José Bustillos-Guzmán
David Lopez-Cortés
Centro de Investigaciones
Biológicas de Baja California Sur Apdo. Postal 128. 23000 La Paz, Baja
California Sur. México.
Muestras de Materia Orgánica Particulada (MOP) fueron
fraccionadas en dos rangos de tallas:2-38 um (fracción "nano") y 38-200
um (fracción "micro"),para aguas de la costa occidental de Baja
California Sur, México. Las muestras fraccionadas fueron analizadas para
estimar su biomasa en términos de proteínas totales,
carbohidralos totales y clorofila a. La fracción de MOP correspondiente
al nanofitoplancton mostró una distribución homogénea en
la capa 0-35 m en toda el área de estudio. El microfitoplacton estuvo
ausente. La fracción de la MOP correspondiente a nanoparticulas no
fotosintéticas presentó una elevada biomasa en la capa 0-20 m
siguiendo estrechamente la incursión de una masa de agua superficial de
origen tropical. La biomasa expresada en proteínas y carbohidratos de
las microparticulas, se distribuyó homogéneamente en la columna
de agua. Se discute y analiza la posible importancia que la fracción no
fotosintética de la MOP (principalmente nanoparticulas detriticas +
bacterias asociadas),representa como fuente alterna de alimento en la cadena
trófica marina.
Proteins and
carbohydrates, of photosynthetic and nonphotosynthetic particulate organic
matter of the Baja California west coast. México.- Samples of
Particulate Organic Matter (POM) were obtained and fractionated in two, range
sizes: 2-38 um (size"nano") and 38-200 um (size "micro"),for the photic zone of
Baja California west coast waters, México.The fractionated samples were
analized to estimate the biomass expressed as total protein, total carbohydrate
and chlorophyll a. The nanophytoplankton fraction was distributed homogeneously
in the layer 0-35 m of the studied area. The microphytoplankton was absent.The
nonphotosynthetic nanoparticles shows a high biomass in terms of proteins and
carbohydrates,in the layer 0-20 m following closely the intrusion of tropical
warm surface water.The biomass in terms of proteins and carbohydrates of
microparticles was distributed homogeneously in the water column. The
importance, as a potentially available food, of the photosynthetic fraction of
POM (maily nanoparticles as detritus + associated bacteria), is discussed and
analised.
El estudio de la Materia Orgánica Particulada (MOP) en el océano es uno de los problemas fundamentales en la oceanografía biológica debido a que básicamente, ésta se constituye de una combinación de partículas detríticas, bacterias, fitoplancton y zooplancton, difíciles de disociar unas de otras en términos de biomasa. Sin embargo, cada uno de este tipo de partículas representa una fuente potencial de alimento para los organismos heterótrofos (Seki et al., 1968; Paffenhofer y Strickland, 1970; Wiebe y Hendricks, 1977; Barlow, 1982; Biddanda, 1988; Biddanda y Pomeroy, 1988; Mann, 1988), por lo que el conocimiento de su composición bioquimica en términos de proteínas y carbohidratos es importante desde el punto de vista de transferencia energética en la pirámide alimenticia. La costa occidental de Baja California Sur aloja, en parte, una de las mayores pesquerías mexicanas como es la de la sardina y anchovta (c.f. Hewitt , 1988); sus fluctuaciones en captura han sido objeto de estudio y han sido el origen de varias hipótesis (Levastu, 1984; Baumgartner y Chiristensen, 1985; Mysak, 1986; Kawasaki y Omori, 1986; Lluch-beldaet al., 1986). Entre ellas, se considera que la fuente de alimento, su composición bioquímica y por consiguiente su digestibilidad (Lasker y Zweifel, 1978; Leiby, 1984), pueden contribuir en el desarrollo de estas fluctuaciones. Como una contribución al conocimiento que la fuente de alimento representa para los organismos heterótrofos, el presente trabajo se orienta a conocer las variaciones espaciales de las diferentes partículas que componen la MOP, así como su composición bioquímica indicadora del valor nutricional de la partícula. MATERIALES Y METODOSLos datos que aquí se presentan fueron obtenidos de un crucero oceanográfico a bordo del B/O "EL PUMA" en julio de 1987, llevado a cabo en la costa occidental de Baja California Sur, México (Fig.1). Se tomaron muestras discretas a tres profundidades estándar (5,20 y 35 m) con la ayuda de botellas NISKIN de 9 litros de capacidad. Este volumen de agua fue filtrado a través de mallas de nylon de 200 um y 38 um de luz de malla. La MOP de la malla de 38 um y aquella menor a 38 um, fue recuperada sobre filtros de fibra de vidrio tipo Whatman GF/C, obteniendo las fracciones "nano"(2-38 um) y "micro"(38-200 um). Estas fracciones fueron obtenidas por triplicado para el análisis de proteínas totales (Lowry et al., 1951), carbohidratos totales (Dubois et al., 1956) Y Pigmentos Fotosintéticos (jeffrey y Humphrey, 1975). Los filtros fueron almacenados a -20°C hasta su análisis en laboratorio. La densidad óptica de los extractos fue medida en un espectrofotómetro Bausch & Lomb Spectronic 2000. Temperatura, salinidad y oxígeno disuelto, fueron obtenidos con un CSTD en cada estación de muestreo, por nuestros colegas del Centro Interdisciplinario de Ciencias Marinas (CICIMAR). Las tablas de datos son reportadas por Cervantes-Duarte et al. (en prensa). RESULTADOS
CONDICIONES FÍSICASLa figura 2, muestra la distribución vertical de temperatura, oxígeno disuelto y los valores de sigma-t, del transecto más oceánico del área de estudio (transecto.60). Los restantes dos transectos (.40 y .20), más cercanos a la costa, básicamente presentan la misma estructura. Es posible observar un avance superficial (0-20 m) de aguas relativamente cálidas. Por abajo de los 20 m se encuentra una masa de agua homogénea con temperaturas oscilando entre 13 y 15°C. ESTRUCTURA BIOQUÍMICAEl análisis de varianza factorial efectuado sobre el conjunto de datos, muestra que las concentraciones de proteínas y carbohidratos de las micropartículas se distribuyen homogéneamente en la columna de agua de 35 m (Tabla 1); valores significativos de clorofila a no fueron obtenidos para las micropartículas. Las concentraciones de proteínas y carbohidratos de las nanopartículas señalan la presencia de un estrato superior, de 0-20 m, con una concentración media similar para proteínas y carbohidratos de 200 mg/m³ , mientras que a la profundidad de 35 m se presentan valores de 70 mg de proteína/m³ y 52 mg de carbohidrato/m³ (Tabla 1). Las concentraciones de clorofila a para las nanopartículas no señalan una diferencia significativa entre las tres profundidades consideradas, con una concentración media de 0.92 mg/m³. Efectuando diagramas de dispersión entre clorofila-proteínas clorofila- carbohidratos y proteínas-carbohidratos, para las nanopartículas, se observa una ausencia de correlación en el estrato de 0-20 m para proteínas y carbohidratos con la clorofila a (Figs. 3a y 3b); situación inversa en el estrato de 35 m donde se obtuvieron correlaciones significativas (Figs. 3c y 3d). Los análisis de correlación entre proteína y carbohidrato para ambos estratos, presentaron altos coeficientes significativos (Fig. 4a, b). Para las micropartículas, los análisis de correlación entre proteínas y carbohidratos presentaron altos coeficientes significativos en todo el estrato de 0-35 m (Fig. 4c). 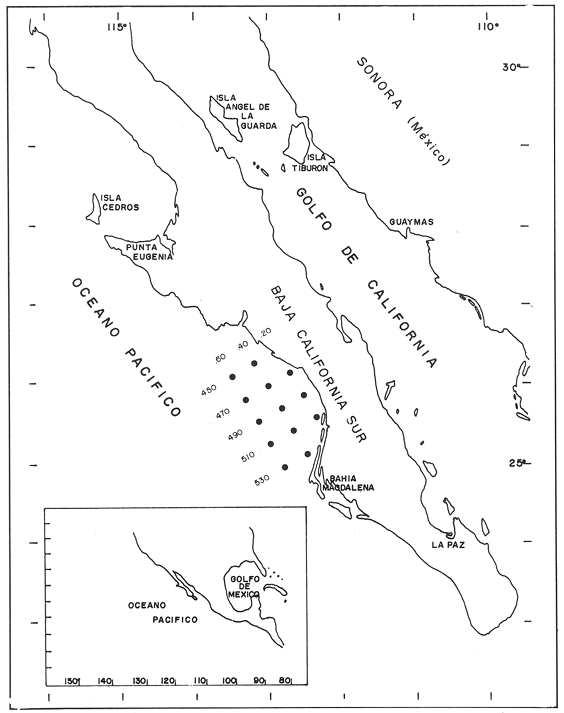 Figura 1. Localización del área de estudio y ubicación de las estaciones de muestreo. Cada estación de muestreo se ubica según las coordenadas X, Y, señaladas (inicio: 450.60; fin: 530.40). 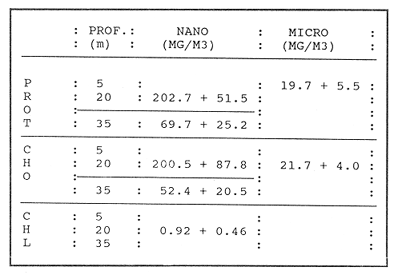 TABLA 1 CONCENTRACIONES MEDIAS (mg/m3) Y DESVIACIÓN ESTÁNDAR (95% LÍMITE DE CONFIANZA) DE PROTEÍNAS, CARBOHIDRATOS Y CLOROFILA a DE LAS FRACCIONES "nano" Y "micro" EN LAS DIFERENTES PROFUNDIDADES MUESTREADAS EN SU TOTALIDAD DE ÁREA DE ESTUDIO 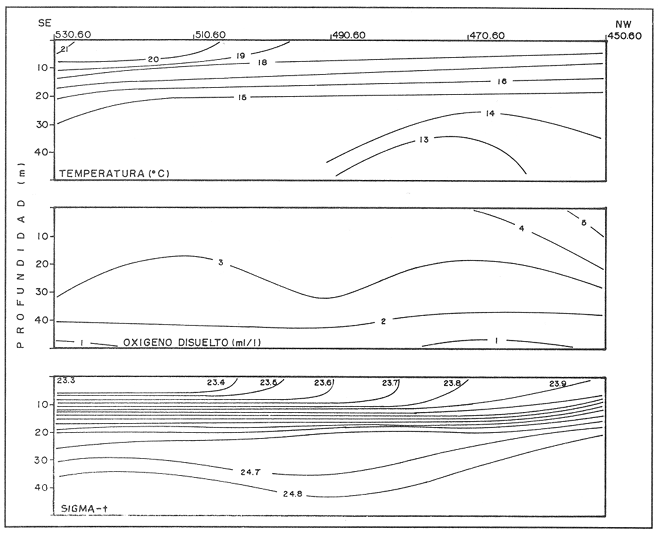 Figura 2. Secciones SE-NW del transecto más oceánico (60), mostrando la estructura física en términos de temperatura, oxígeno diselto y sigma - t. 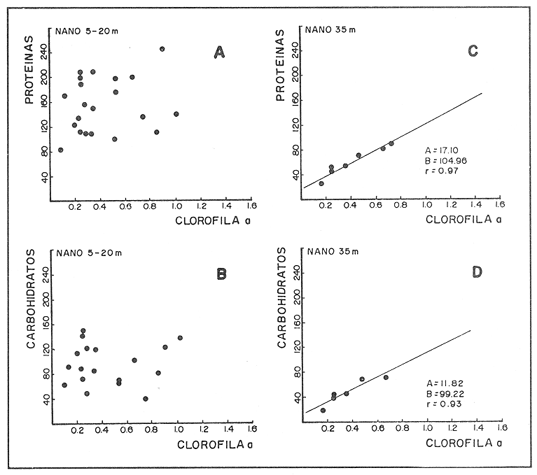 Figura 3. Diagramas de dispersión y análisis de correlación de proteínas, carbohidratos y clorofila a, para la fracción "nano", en los estratos 5-20 ni (AyB) y 35 ni (CyD). DISCUSIÓNEl análisis factorial de varianza y los análisis de regresión sobre el conjunto de datos obtenidos en este crucero, señala principalmente: a) el nanofitoplancton fue el único contribuyente a la biomasa expresada en clorofila a, y ésta, se distribuye homogéneamente en el estrato 0-35 m de toda el área de muestreo. b) Las concentraciones de proteínas y carbohidratos de la fracción "nano" en el estrato de 0-20 m están fuertemente influenciadas por material no fotosintético, presumiblemente nanodetritus. c) El microfitoplancton estuvo ausente en este estrato de 0-35 m por lo que las concentraciones de proteínas y carbohidratos, presumiblemente están asociadas al microzooplancton más micropartículas detríticas. La estrecha correlación que guardan los parámetros de proteínas y carbohidratos con la clorofila a la profundidad de 35 m, nos indica la dominancia del nanofitoplancton sobre el nanodetritus. Los cocientes promedio entre proteína/clorofila y carbohidrato/clorofila obtenidos en este estudio (75.8 y 57.0 respectivamente), son comparables con los obtenidos en cultivos uniespecíficos (c.f. Darley, 1977). La homogeneidad en la distribución vertical de la clorofila a, el incremento en las biomasas de proteínas y carbohidratos a los 5 y 20 m, y la ausencia total de correlación entre estos compuestos bioquimicos con la clorofila a, nos hacen suponer que la biomasa nanodetrítica aumentó considerablemente en esta zona la cual tiene como frontera el límite inferior de la termoclina. Los análisis de correlación entre la biomasa expresada en términos de carbón o nitrógeno, con la clorofilaa, han sido utilizados con frecuencia para discriminar el carbón perteneciente al fitoplancton del que pertenece a otras partículas no fotosintéticas (Mcallister et al., 1960; Banse, 1977; Sakshaug, 1978; Lancelot-van Beveren, 1980; Herbland y Le Bouteiller, 1983; Verlencar y Qasim, 1985). Lechuga-devezeet al. (en prensa), menciona la influencia que la proteína y el carbohidrato de origen detrítico puede representar para la MOP total, en ocasiones alcanzando el 95% de su biomasa expresada en carbohidrato. 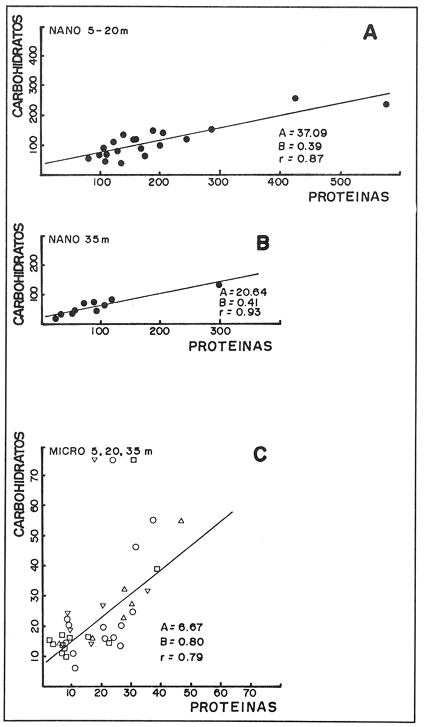 Figura 4. Correlaciones entre proteínas y carbohidratos para las fracciones "nano" (5-20 m A; 35 m, B) y "micro" (5-20-35 m, C). La división por rangos de tallas de esta MOP, señala que en la fracción "nano" se encuentra la mayor biomasa detrítica y fotosintética. En medios ambientes oceánicos, gran parte de la MOP deriva del fitoplancton vivo o muerto (Biddanda, 1988). Este último es un sustrato preferencial para la instalación de bacterias cuya actividad provoca la disgregación de la partícula detrítica, en un proceso continuo (BIddanda y Pomeroy, 1988). La importancia del material detrítico en la cadena alimenticia como sostén de invertebrados y de la producción pesquera, aún no está bien definida (Mann, 1988). Biddanda (1988) y Biddanda y Pomeroy (1988) mencionan que los agregados detritusbacterias pueden constituir una fuente significativa de carbón para metazoarios capaces de consumirlos. Paffenhofer y Strickland (1970), Seki et al. (1968), Wiebe y Hendricks (1974), Heinle et al. (1977), Barlow (1982), Lechuga-deveza et al.(en prensa), consideran que estos complejos detritus-bacterias poseen una composición bioquímica competitiva con la que presenta el fitoplancton, y por lo tanto, su aprovechamiento potencial por organismos en niveles tróficos superiores en la cadena alimenticia, puede ser de magnitudes aún no consideradas. En el desarrollo de este trabajo es notorio el avance de aguas cálidas superficiales creando dos zonas bien definidas: una superficial (0-20 m) bien estratificada, con temperaturas entre 15 y 21°C; y otra (20.50 m), bien homogénea, con temperaturas entre 13 y 15°C. A pesar de esta estructura, nuestros resultados sugieren que la biomasa del nanofitoplancton expresada en clorofila a, se mantiene significativamente homogénea en la capa 0-35 m, mientras que, aparentemente, la biomasa "nano" de MOP, expresada en proteínas y carbohidratos, en la capa 0-20 m, tiene una distribución significativa y dominante, por material detrítico y las bacterias asociadas a éste. Dicha distribución de los contribuyentes principales de la MOP no ha sido reportada aún. Dos teorías pueden ser posibles: el debilitamiento de la corriente de California durante el verano y el avance del frente tropical (Wyrtki, 1965) que acarrea consigo material particulado justo en la capa que corresponde a este avance tropical; por otro lado, podría considerarse una formación de MOP a partir de la materia orgánica disuelta (MOD) sujeta a la acción de efectos físicos (viento, oleaje) los cuales estuvieron presentes en el desarrollo de este crucero (Biddanda, 1988; y Pomeroy, 1988; Mann, 1988). Cualquiera que haya sido el mecanismo (transporte o generación física y biológica) originario de este material particulado, es evidente que su biomasa expresada como proteína y carbohidratos, representa una fuente energética importante. Investigaciones sobre la alimentación deEngraulis mordax señalan su preferencia hacia dinoflagelados (Lasker, 1974) sobre cadenas de diatomeas (Scura y Jerde, 1977). Sin embargo, Hitchcock (1982), señala que estas observaciones no demuestran que los dinoflagelados son preferidos como alimento, sino que otros factores, como el "sabor" de la partícula, debe ser tomado en consideración. La captura no selectiva de E. modaxpuede incluir en su dieta a partículas detríticas del tamaño y valor nutricional adecuados. Por tal motivo, si este material detrítico domina bajo ciertas circunstancias en el medio acuático, su captura puede llegar a ser de un valor nutricional importante. De los resultados expresados en este trabajo, se refuerza la hipótesis de considerar al detritus como una fuente importante de alimento en el cadena trófica marina. Se ha hecho mención que mas energía y material particulado es transferido al zooplancton por la vía del detritus que por fitofagia directa Heile, 1977; Mann, 1988). Falkowski et al. (1988), menciona que el consumo por herbívoros es del orden del 34%; el 51% de este fitoplancton se hunde quedando a merced de la acción bacteriana, ya sea en la columna de agua o en el fondo de la plataforma continental, y del 10% al 20% se exporta al océano profundo. Está generalmente establecido que aproximadamente el 18.5% de la producción primaria total marina, la cual se genera en la zona de la plataforma continental y por eventos de surgencias, soporta más del 95% de las pesquerías comerciales; sin embargo, la inclusión del material detersivo como fuente potencial de alimento puede modificar sustancialmente este concepto y puede otorgar una alternativa para la explicación parcial de las grandes fluctuaciones en la pesquería, principalmente de sardina y anchoveta, ocurridas a lo largo de toda la costa. Este del Océano Pacífico, y en otras regiones oceánicas. LITERATURABANSE, K. A review. J. Phycol. Rates of growth, respiration and photosynthesis of unicellular algae as related to cell size.1977135-14012 BARLOW, R. G. J. Exp. Mar. Biol. Ecol.Phytopiankton ecology in the southem Benguela current. 1. Biochemical composition.1982209-22763 BAUNIGARTNER, T. R. y N. Christensen Jr. Coupling of the Gulf of California to large-scale interannual climatic variability.J. Mar. Res.1985825-84843 BIDDANDA, B. A.Microbial metabolismMicrobial aggregation and degradation of phytoplankton-derived detritus in seawaler. II.Mar. Ecol. Progr. Ser.198889-9542 BIDDANDA, B. A. y L. R. Pomeroy.l. Microbial succession Microbial aggregation and degradation of phytoplankton-derived detritus in seawater.Mar. Ecol. Progr. Ser.198879-8842 CERVANTES-DUARTE, R. J. M. Cendejas, y R. Guerrero-Godínez, (en prensa). Parámetros físico quínuicos de la costa Occidental de Baja California Sur. Atlas CICIMARCruceros 8707, 8710.La Paz, B.C.S.México DARLEY, W. M. The Biology of Diatoms. Biochemical coniposition Werner , D. University of Cilifornia Press.1977498 p. DUBOIS, M. K., A. Gilles, J. K. Hamilton, P. A. Rebers, y F. SmithAnal. Chem.Colorimetric method for determination of sugars and related substances.1956350-35628 FALKOWSKI, P. G., C. N. Flagg, G. T. Rowe, S. L. Smith, T. E. Whitledge, y C. D. Wirick, The fate of a spring phyloplinkton bloom: export or oxidalion?.Cont. Shelf. Res.1988457-4848 HEINLE, D. R., R. P. Harris, J. F. Ustach, y D. A. Flemer Detritus as food for estuarine copepods. Mar. Biol.1977341-35340 HERBlAND, A., y A. Le BoutellierDynamique du phytoplancton et matiere organique particulares dans la zone cuphotique de l'Atllntique equatorial. Mar. Biol.1983265-27872 HEWITT, R. P. Historical review of the oceanographic approach to fishery research.CalCOFI Rep.198827-4129 HITCHCOCK, G. L. A compirative study of the size-dependent organic composition of marine diatoms and dinoflagellates. J. PL. Res.1982363-3774 JEFFREY, S. W. y G. F. HumphreyNew spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in algal, phytoplankton and Brigadier plants.Biochem. Physiol. Pfatiz. B. P. P.1975191-194167 KAWASAKI, T., y M. OmoriFluctualions in the three major sardine stocks in the pacific, and global trends in temperature.Internat. Symp. on Long Term Changes in Marine Fish PopulationsVigoEspaña198618-21 Nov. LANCELOT-VAN BEVEREN, C.A statistical method to estimate the biochemical composition of phytoplankton in The southern bight of the North Sea.Est. Coast. Mar. Sci.1980467-47810 LASKER, R. Fish. Bull. Field criteria for survival of anchovy Larvae: the relation between inshore chlorophyll maximum layers and successful first feeding. 1974453-46273 LASKER, R. y J. R. SwifelSpatial pattern in plankton communities, Growth and survival of first-feeding northern anchovy larvae (Engraulis mordax) in patches containing differents proportions of large and small prey. J. H. Steele Plenum Press.1978470 p. LECHUGA-DEVEZE, C. H., B. Ayala-Rocha, y I. Gárate-Lizárraga An. Inst. Cienc. del Mar y Linmol., Proteínas, carbohidratos y clorofila de la Materia Orgánica Particulada en tres diferentes ambientes oceánicos: Mar de Portugal, Golfo de Califórnia y NW de Baja California Sur.UNAM1989 (en prensa).16(1) LEIBY, M. M. Marine Plankton Life Cycle StrategiesLife history and ecology of pelagic fish eggs and larvae K. A. Steindinger y L. M. Walker CRC Press.1984158 p. LEVAST, T. The effects of temperature anomalies on the fluctuation of fish stock. Rapp. P. v. Reun. Cons. Int. Explor. Mer.1984214-225185 LOWRY, O. H., N. J. Rosebrough, A. L. Farr, y R. J. Randal Protein measurement with folin phenol reagent. J. Biol. Chem.1951265-275193 LLUCH-BELDA, D., F. J. Magallon, y R. A. SchwartzloseLarge fluctuations in the sardine fishery in the Gulf of California. Possible causes. CalCOFI REP1986136-14327 MANN, K. H. Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems. Limnol. Oceanogr.1988910-93033 McALLISTER, C. D., T. R. Parsons, y J. D. H. StricklandPrimary productivity and fertility station "P" in the north-east Pacific Ocean. J. Cons. Internat. Explor. Mer.1960240-25925 MYSAK, L. A. El Niño, interannual variability and fisheries in the northeast Pacific Ocean. Can. J. Fish Aquat. Sci.1986464-49743 PFAFFENHOFER, G.A., y J. D. H. Strickland, . A note on the feeding of Calanus helgolapodicus on detritus.Mar. Biol, .197097-995 SAKSHAUG, E. The influence of environmental factors on the chemical composition of cultivaled and natural populations of marine phytoplankton.University of Trondheim. Trondjem Biological Station, Norway.197882 p. SCURA, E. D., y C. W. JerdeFish. Bull. Various species of phytoplankton as food for larva, northern anchovy, Engraulis Mordar, and relative nutritional value of the dinoflagellates Gymnodinium splendes and Gonyaulax polyedra.1977579-58375 SEKI, H., J. Skelding, y T. R. Parsons Observations on the decomposition of a marine sediment. Linmol. Oceanogr.1968440-44713 VERLENCAR, X. N., y S. Z. QasimParticulate organic matter in the coastal and estuarine waters of Goa and its relationships with phytoplankton. Est. Coast. Mar. Sci.1985235-24321 WIEBE, W. J. y C. W. Hendricks, Effect of the Ocean Environment on Microbial Activities.Distribution of heterotrophic bacteria in a transect of the Antartic Ocean. R. R. Colwell y R. Y. Morita Univ. Park. Press.1977535 p. WYRTKI, K. Corrientes superficiales del Océano Pacífico Oriental tropical. Com. Infer-Am Atun Trop.1965195-30419
|