|
MOLUSCOS DE LA LAGUNA
DE ALVARADO, VERACRUZ: SISTEMÁTICA Y ECOLOGÍA
Trabajo
recibido el 27 de julio de 1989 y aceptado para su publicación el 21 de
septiembre de 1989.
Martha Reguero
Antonio García-Cubas
Instituto de Ciencias del Mar y
Limnología, Universidad Nacional Autónoma de México.
Contribución No. 664 del Instituto de Ciencias del Mar y
Limnología, UNAM.
En este trabajo se hace un
análisis de la estructura de la comunidad malacológica de la
laguna de Alvarado, la cual se encontró compuesta por 23 especies de las
Clases Gastropoda y Bivalvia, destacando por su valor de importancia
relativaRangia flexuosa, Neritina reclivata, Mulinia lateralis y
Littoridina sphinctostoma. Entre los gasterópodos predominaron
las especies epifaunales y herbívoro-raspadoras. La mayoría de
los bivalvos presentaron nutrición suspensívora y hubo un
número igual de formas infaunales que de epifaunales. Los valores medios
de similaridad fueron de 71.8% para la Clase Gastropoda, 65.5% para la Clase
Bivalvia y 69.2% para el phylum Mollusca. El cálculo de la diversidad y
sus componentes se hizo tanto para la comunidad total (vivos + muertos) como
para la viva. En el primer caso, los valores más altos se registraron en
la Clase Gastropoda y, en el segundo, correspondieron al Phylum. El
análisis de variación de la diversidad indica que hay diferencias
estadísticamente significativas entre los resultados de la comunidad
total y los obtenidos para la comunidad viva. Con base en las condiciones
hidrológicas y la fauna presente se diferenciaron tres ambientes
halinos, con mayor registro de diversidad en el
oligohalino.
In this paper, an analysis of the Alvarado's lagoon
mollusks community structure was made. 23 species of the Classes Gastropoda and
Bivalvia were founded, and because of their relative importance values detached
Rangia flexuosa, Neritina reclivata, Mulinia lateralis and
Littoridina sphinctostoma. Epifaunal and herbivorous-scraper
species dominated among the gastropods. Suspensivorous nutrition was present in
the majority of bivalves, having equal number of infaunal and epifaunal forms.
Mean similarity values were 71.8% for Class Gastropoda, 65.5% for Class
Bivalvia, and 69.2% for Phylum Mollusca. Diversity indexes as weil as its
elements computations were made both for the total community (alive + shells)
and the alive one. In the former, highest values were recorded for the Class
Gastropoda, and in the latter for the Phylum. Diversity variance analysis
pointed out the existence of statistically significant differences between the
results of total community and those obtained for the alive fauna. Three haline
environments were differentiated on the basis of hydrological conditions and
the presence of fauna, with the greater diversity record in the oligohaline
one.
Los complejos sistemas lagunares aledaños a la costa son fuentes importantes de recursos naturales accesibles a las comunidades humanas. De ahí la necesidad de llevar a cabo investigaciones científicas sobre estas zonas, que permitan incrementar su productividad y sustentar estudios posteriores. En el presente trabajo se abordan aspectos relacionados con la sistemática, abundancia y distribución de los moluscos de la Laguna de Alvarado, Ver., como parte de un proyecto más amplio que desarrolla el Laboratorio de Malacología del Instituto de Ciencias del Mar y Limnología de la UNAM, en el que se incluye el estudio de la malacofauna de 22 lagunas costeras del litoral mexicano del Golfo de México. La fauna malacológica es uno de los componentes bióticos más relevantes de estos ecosistemas, debido a su posición de consumidores dentro de la red trófica y a que algunas especies nativas, como ostiones y almejas, están sujetas a explotación comercial y son objeto de cultivos extensivos. Así, el análisis de la estructura de la comunidad de moluscos hará más comprensible su relación con el ambiente y posibilitará su mejor aprovechamiento. ANTECEDENTESLa Laguna de Alvarado ha sido objeto de numerosos estudios, entre los que figuran los de Villaloboset al. (1966), sobre hidrografía y productividad; Villalobos et al. (1969), en el que se establecen las relaciones entre postlarvas planctónicas de camarón y el ambiente; Villalobos et al. (1975), acerca de la hidrobiología del área; Signoret (1969), sobre medusas; Reséndez (1973), relacionado con la ictiofauna; Gómez-Aguirre (1977), el cual es un estudio comparativo del plancton en las lagunas de Alvarado, Términos y Tamiahua; Flores-Coto y Méndez Vargas (1982), quienes trabajaron el ictioplancton; Vargas Maldonado (1986), que trata sobre la estructura de las comunidades de peces; Fernández (1989), acerca de variaciones poblacionales de crustáceos; García Montes (1989), sobre estructura de comunidades de macroinvertebrados epibénticos y un trabajo en preparación de Raz-Guzmánet al. en el que se hace una caracterización hidrológica y sedimentológica del área. Los trabajos realizados en el país sobre malacofauna del Golfo de México son escasos, sobre todo los que se refieren a lagunas costeras. Entre ellos se encuentran los de García-Cubas (1963, 1968, 1969, 1981, 1988) y Antolí y García-Cubas (1985), los cuales versan sobre sistemática y ecología de moluscos. DESCRIPCIÓN DEL ÁREA DE ESTUDIOEl complejo lagunar de Alvarado está constituído, de Norte a Sur, por las lagunas Camaronera, Buen País, Alvarado y Tlalixcoyan. Se localiza al Sureste del Estado de Veracruz, entre las coordenadas geográficas 18º 44' 00" y 18° 52' 15" de latitud Norte y 95º 44' 00" y 95º 57' 00" de longitud Oeste (Flores-Coto y Méndez Vargas, 1982). El presente trabajo no abarca todo el sistema, sino que se restringe a las Lagunas de Alvarado y Buen País, incluyendo una pequeña porción de influencia de Tlalixcoyan (Fig. l). La Laguna Camaronera, por sus características peculiares, será objeto de un estudio posterior. CLIMATOLOGÍADe acuerdo con García (1971), la Laguna de Alvarado presenta un clima de tipo Aw"2 (i), que corresponde a cálido subhúmedo -el más húmedo de los subhúmedos-, con lluvias en verano. La temperatura media anual es de 26°C y la media del mes más frío por arriba de 18°C, con una oscilación entre 5 y 7°C. La temporada de sequía se presenta entre los meses de enero a mayo, la de lluvias inicia en junio y los Nortes tienen lugar en noviembre, generalizándose en enero (Reséndez, 1973). HIDROGRAFÍAEn la región Sureste del complejo descarga sus aguas el río Papaloapan; los ríos Camarón y Blanco desembocan en Tlalixcoyan y el Acula se conecta directamente con el cuerpo de agua central (Flores-Coto y Méndez Vargas, 1982). HIDROLOGÍALos aportes de agua continental que recibe permanentemente la Laguna de Alvarado a través de los ríos, se ven incrementados en la época de lluvias y "Nortes". Por otra parte, el flujo de aguas marinas se lleva a cabo por la desembocadura del río Papaloapan y por un canal artificial que comunica a la laguna Camaronera con el mar. Esta apertura fue posterior a los estudios de Villaloboset al. (1975), quienes propusieron la división del complejo lagunar en subsistemas, que han sido modificados en función de los cambios salinos e hidrológicos del área. Así, Vargas-Maldonado (1986) identifica tres zonas: A: Area de Gradiente Hidrológico. Zona de gran mezcla de agua dulce y marina, presenta gradientes horizontales y estratificación la mayor parte del año. Incluye lo que es propiamente la Laguna de Alvarádo, Buen País y la boca del río Papaloapan. 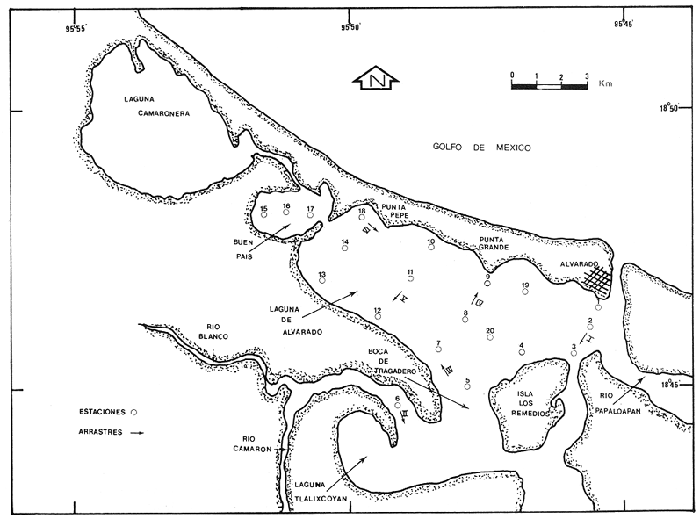 Figura 1. Localización del área de estudio y estaciones de muestreo. B. Area de Influencia Dulceacuícola. Zona con gradientes horizontales de menor magnitud e influencia preponderante de agua dulce. Sin estratificación. Laguna de Tlalixcoyan. C. Area de Influencia Nerítica. Con gradientes horizontales, influencia dulce y marina, pero en mayor grado esta última. Sin estratificación. Laguna Camaronera. BatimetríaReséndez (1973) señala que, en términos generales, Alvarado es una laguna somera, con profundidades máximas de 2.2 m en su porción central, a excepción del lecho del río Papaloapan y canales en donde se han registrado profundidades hasta de 13 m. TransparenciaLos porcentajes promedio para las tres épocas son: nortes 36.0% secas 38.8% lluvias 16.5% Alvarado es un sistema de aguas muy turbias, con transparencias aún menores para la época de lluvias (Vargas-Maldonado, 1986). Temperatura del aguaFlores-Coto y Méndez Vargas (1982) registran para el verano temperaturas superficiales entre 25 y 30ºC, muy similares a las de fondo que van de 25.1 a 31ºC, mientras que Fernández (1989) y García-Montes (1989) señalan variaciones entre 29 y 34ºC para el mes de julio. SalinidadLos rangos de salinidad reportados por Vargas-Maldonado (1986) para cada una de las épocas son: nortes 5.00 a 22.00 % secas 1.65 a 35.58 % lluvias 0.12 a 5.92 % García-Montes (1989) registra, para el mes de julio, salinidades que van desde 3.5 hasta 12 %. SedimentosContreras (1985) señala que en el área pueden encontrarse sedimentos de tipo arenoso y limo-arcilloso, coincidiendo con Raz-Guzmán et al. (en preparación) quienes indican que el sistema lagunar se caracteriza por sedimentos limo-arcillosos con un alto contenido de materia orgánica en el extremo noroeste de la laguna y por sedimentos arenosos, pobres en materia orgánica, en el resto del sistema. VegetaciónEl contorno de la laguna se encuentra poblado por manglares, predominando el "mangle rojo"Rhizophora mangle, aunque se observan tambiénAvicenia germinans y Laguncularia racemosa (Reséndez, 1973). En la época de lluvias abunda el lirio acuático Eichornia crassipes(Contreras, 1985). La vegetación sumergida del área litoral inmediata a la zona de manglar está integrada casi totalmente porRuppia maritima. A dicha flora se asocia estrechamente una fauna muy particular, entre la que figuran varios moluscos. En aguas más profundas se encuentran diversas especies de algas, destacando la rodofitaGracilaria verrucosa (Reséndez, 1973). MATERIALES Y MÉTODOSLa recolecta del material y el registro de datos ambientales se llevaron a cabo durante una campaña realizada en el mes de julio de 1980, coincidiendo con la temporada de lluvias. Se hizo un reconocimiento del área para excluir zonas con presencia de bancos de ostión y almeja y se estableció una red de 20 estaciones distribuídas en el sistema lagunar (Fig. 1). En cada una de ellas se midió la temperatura y salinidad con un salinómetro de inducción Beckman, la transparencia con un disco de Secchi y la profundidad con una sondaleza de mano. Se extrajo un volumen de cinco litros de sedimento superficial húmedo, por medio de una draga tipo Van Veen, con capacidad de dos litros, efectuándose los lances necesarios para extraer la cantidad deseada. Los sedimentos obtenidos se tamizaronin situ, con una malla de plástico, con abertura de 1.5 mm. Los materiales resultantes fueron preservados en bolsas de polietileno con formol al 10%, neutralizado con borato de sodio y debidamente etiquetados para su traslado al Laboratorio de Malacología del Instituto de Ciencias del Mar y Limnología de la UNAM, en donde se lavaron con agua corriente, se secaron a temperaturas inferiores a 70°C, separando las formas recolectadas vivas de los exoesqueletos para proceder a su identificación y cuantificación a nivel específico. En el caso de los bivalvos con conchas desarticuladas se adoptó el criterio de García-Cubas (1963), quien considera cada valva como un organismo. Para fines cualitativos, se efectuaron 6 arrastres mediante una red biológica con boca de 300 cm² y malla de 2 cm, durante 5 min, a una velocidad promedio de 2 nudos. Las especies obtenidas fueron incluídas en la lista sistemática, aunque no se consideraron para el análisis cuantitativo de la comunidad, ya que la técnica de captura no es adecuada para ese propósito. Los ejemplares representativos se incorporaron a la colección malacológica del Instituto de Ciencias del Mar y Limnología. La comunidad de moluscos se caracterizó mediante los componentes de abundancia, frecuencia y densidad de las especies (individuos/m²), de donde se derivó el valor de importancia relativo a cada una de ellas y el índice de similaridad entre las estaciones de muestreo (Krebs, 1972). Lo anterior se complementó con indicaciones de distribución geográfica y local, hábitos alimenticios y hábitat. Asimismo, se calcularon los índices de diversidad (H'n), diversidad máxima (Hmax), equitatividad (J) y riqueza de especies (D) para cada una de las Clases de moluscos y por ambiente halino, mediante las expresiones matemáticas propuestas por Shannon-Wiener (1963), Pielou (1975) y Margalef (1958) respectivamente, efectuando un análisis de variación de la diversidad (Basharin, 1959; Lloyd, Zar y Karr, 1968) para estimar las diferencias entre los resultados obtenidos del cálculo de este índice aplicado a la comunidad total y a las poblaciones vivas (Hutcheson, 1970; Zar, 1974). El análisis incluyó también la estimación de los coeficientes de determinación entre los valores de H'n vs. J y H'nvs. D, para establecer el factor dominante sobre la diversidad en cada taxón, trabajando por separado la comunidad viva y la total. RESULTADOS
HIDROLOGÍAPor lo que se refiere a los rangos de temperatura y salinidad de fondo en el área, éstos variaron de 28.8 a 30.7ºC y de 0.2 a 5.6 %, respectivamente. La transparencia del agua fluctuó entre 0.20 y 0.60 m, dependiendo de la profundidad, para la cual se registraron valores desde 0.80 a 6.60 m. Los datos detallados de estos parámetros para cada una de las estaciones de muestreo se concentran en la Tabla 1. 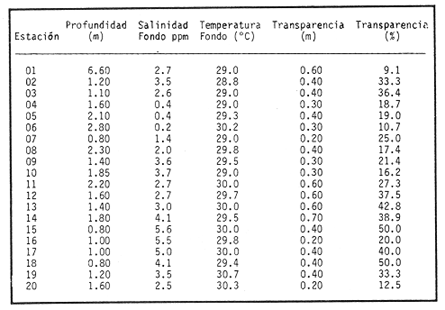 TABLA 1 PARÁMETROS HIDROLÓGICOS EN LAS ESTACIONES DE MUESTREO Con base en los valores de salinidad y siguiendo el criterio de García-Cubas (1981), modificado de Välikangas (1933), fue posible caracterizar el sistema lagunar para la temporada de colecta en 3 ambientes distintos: 1) Limnético (0 - 0.5 %), ubicado en la zona de comunicación entre las Lagunas de Alvarado y Tlalixcoyan, donde es notable la influencia de aguas dulces aportadas por los ríos Blanco, Camarón y Papaloapan; 2) Oligohalino (0.5 - 5 %), que corresponde al cuerpo lagunar principal de la Laguna de Alvarado; y 3) Mesohalino de tipo "A" (5 - 10 %), que comprende el área de Buen País, donde se hace evidente la influencia de aguas procedentes de la Laguna Camaronera (Fig.2). CONSIDERACIONES A LA SISTEMÁTICAEl arreglo que se presenta a continuación se basó en el criterio morfológico, usando únicamente las conchas. Para el caso de Bivalvia se hizo énfasis sobre escultura externa, charnela, impresiones musculares, presencia o ausencia de seno palial y sus dimensiones. En la Clase Gastropoda se tomó en cuenta la longitud y amplitud de los canales anal y sifonal, pliegues columelares, número de vueltas constituyendo la espira, ornamentaciones y, en algunos casos, caracteres del opérculo. La identificación de la malacofauna se efectuó de acuerdo con García-Cubas (1981) y Antolí y García-Cubas (1985), quienes adoptan para la Clase Bivalvia el sistema de clasificación propuesto por Newell (1965) y para la Clase Gastropoda, el criterio de Thiele (1934-1935). Asimismo, las indicaciones sobre distribución geográfica, habitat y tipo de nutrición de las especies fueron extraídas de las dos primeras obras citadas anteriormente. Los organismos quedaron ubicados sistemáticamente en 2 Clases, 15 Familias, 19 Géneros y 23 Especies. SISTEMÁTICA Y ECOLOGíA
 SISTEMÁTICA Y ECOLOGÍA  Continuación SISTEMÁTICA Y ECOLOGÍA  Continuación SISTEMÁTICA Y ECOLOGÍA  Continuación SISTEMÁTICA Y ECOLOGÍA ANÁLISIS CUANTITATIVO Y PORCENTUALEn un total de 20 estaciones de muestreo se recolectaron 63,283 individuos, de los cuales 2,040 (3.2%) fueron capturados vivos y 61,243 (96.8%) correspondieron a conchas. En el ecosistema estudiado se identificaron 13 especies para la Clase Gastropoda y 10 se ubicaron en la Clase Bivalvia. De un total de 23 especies, sólo 4 (Pomacea flagellata, Cingula floridanus, Nassarius acutus y Rangía cuneata) fueron excluídas del análisis cuantitativo por haber sido capturadas con red de arrastre biológica y no mediante draga Van Veen. Por lo que se refiere a abundancia (Tabla 2), la Clase Bivalvia representó el 83.5% (52,839 ejemplares) del total de organismos recolectados y en la Clase Gastropoda el valor porcentual resultó de 16.5% (10,444 individuos). Las estaciones con mayor número de ejemplares fueron la 5 y 17, con 20,088 y 10,856 individuos, respectivamente; mientras que el menor número de organismos se registró en las estaciones 1 (181 ejemplares) y 15 (70 ejemplares). En lo que respecta a estaciones con el número más elevado de especies, éstas fueron la 1, 6, 9 y 17 (con 13 en cada una) y las que presentaron menor número fueron la 20, 16 y 8, con 7, 6 y 4 especies, respectivamente. 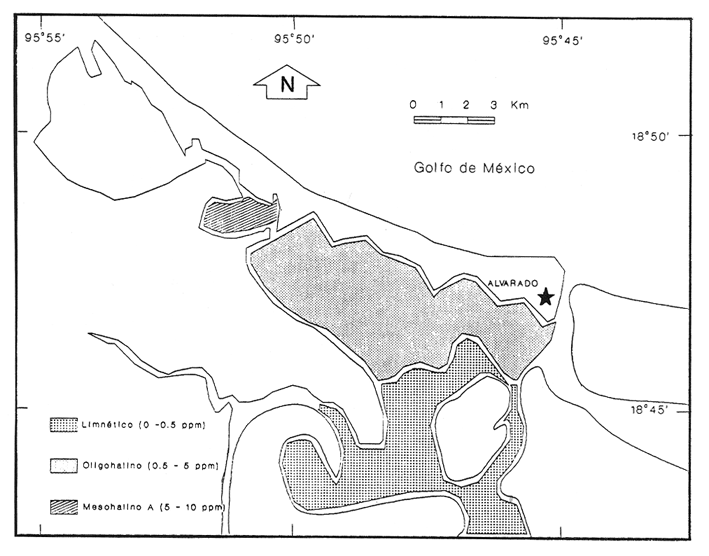 Figura 2. Perfil halino del área de estudio. De las 19 especies a las que se les aplicó análisis cuantitativo, 11 (57.89%) tuvieron representantes vivos: 7 pertenecientes a la Clase Bivalvia y 4 a la Gastropoda, predominando, en la época de colecta,Polymesoda caroliniana, Brachidontes exustus, Ischadium recurvum y Neritina reclivata (Tabla 3). El porcentaje de bivalvos capturados vivos se eleva a un 87.6% (1,788 individuos) sobre 12.4% (252 individuos) de gasterópodos, registrándose una mayor abundancia en las estaciones 17, 9 y 19. En el sistema lagunar se reconocieron áreas en donde comienzan a recuperarse poblaciones que en épocas anteriores fueron objeto de una explotación comercial intensiva. Así, en la zona de la Laguna de Alvarado próxima a Buen País se detectaron bancos de Rangia cuneata y dispersos en el cuerpo lagunar principal se observaron otros deCrassostrea virginica. Sin embargo, de ninguno de ellos se extrajeron muestras para el presente trabajo. De manera que al excluir estas formas que se encuentran constituyendo bancos, el análisis cuantitativo vierte como resultado que el área de estudio está caracterizada por 10 especies: 6 de la Clase Bivalvia (Rangia flexuosa, Mulinia lateralis, Mytilopsis leucophaeata, Polymesoda caroliniana, Brachidontes exustus, Ischadium recurvum) y 4 de la Clase Gastropoda (Neritina reclivata, Littoridina sphinctostoma, Neritina virginea, Cerithidea pliculosa), que en conjunto representan el 98.5% respecto al total de individuos recolectados. No obstante, cuatro de las especies citadas son las que destacan por su gran abundancia y distribución generalizada: Rangia flexuosa, Neritina reclivata, Mulinia lateralis y Littoridina sphinctostoma, las cuales constituyen el 86% de la población total, siendo R. flexuosa la forma más importante en la comunidad malacológica, alcanzando por sí sola un valor porcentual más elevado (65.6%) que el resto de las especies en conjunto. 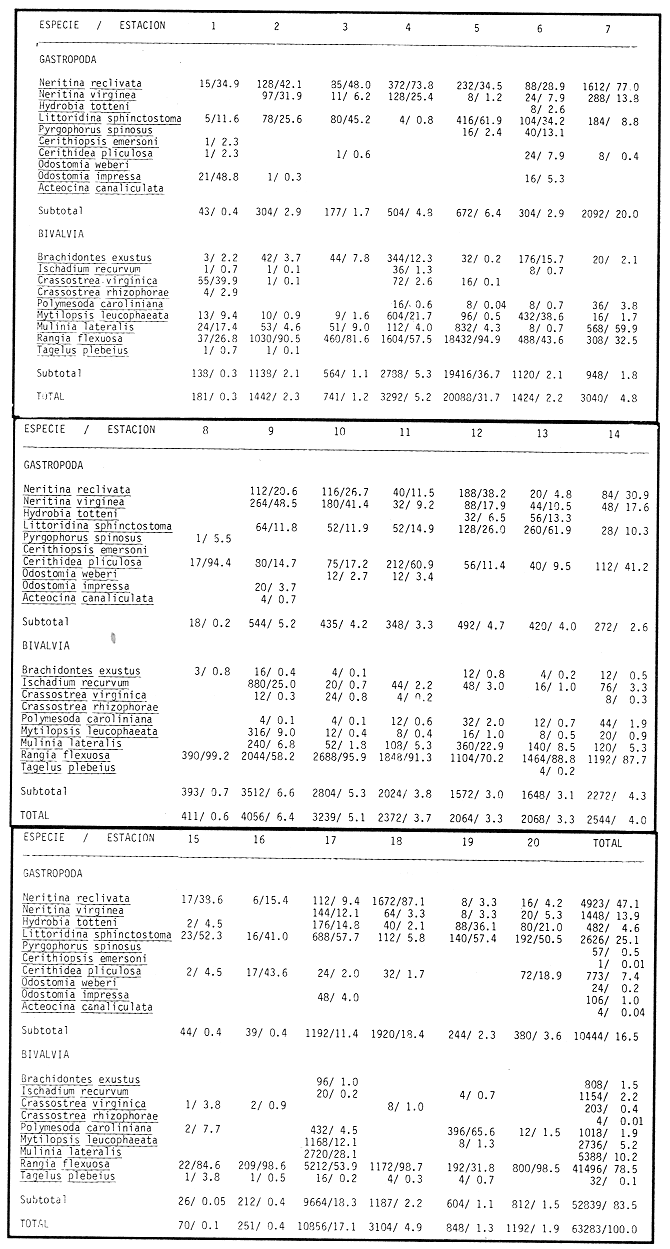 TABLA 2 ABUNDANCIA NUMÉRICA Y PORCENTUAL DE LAS ESPECIES DE MOLUSCOS 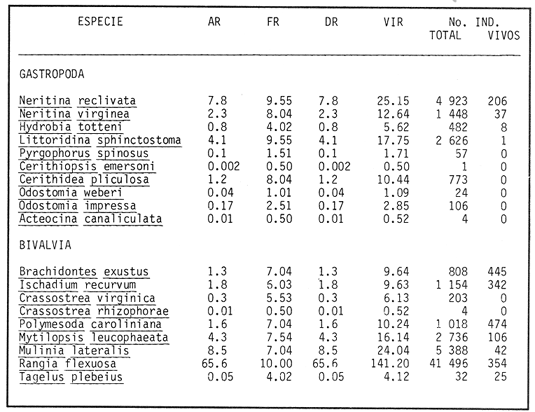 TABLA 3 ABUNDANCIA RELATIVA (AR), FRECUENCIA RELATIVA (FR), DENSIDAD RELATIVA (DR), VALOR DE LA IMPORTANCIA RELATIVA (VIR) Y NÚMERO DE INDIVIDUOS (No. IND.) DE LAS ESPECIES DE MOLUSCOS Los valores de abundancia, frecuencia, densidad e importancia relativas de las especies, que sustentan esta caracterización del área de estudio, se encuentran detallados en la Tabla 3, conforme a su arreglo sistemático. Con el fin de establecer una relación de la distribución y asociaciones de las especies que integran la comunidad de moluscos, se aplicó porcentualmente el índice de similaridad de Krebs (1972), el cual permite evaluar la semejanza entre los componentes de las muestras o estaciones del sistema. Con los valores obtenidos de comparar la ocurrencia de especies comunes en las 20 estaciones se elaboraron diagramas de enrejado (Figs. 3- 5), en donde se observa que las estaciones más afines en componentes específicos de la Clase Gastropoda fueron, en orden decreciente, la 3, 7, 14, 12 y 13; para la Clase Bivalvia, la 4, 9, 10 y 14, y para el Phylum Mollusca, la 14,13,12,17 y 10, registrándose una similitud media por taxón en todo el sistema lagunar de 71.8%, 65.5% y 69.2%, respectivamente. Nutrición y habitatDe acuerdo con la información bibliográfica (Purchon, 1968; Andrews, 1971; Abbott, 1974; García-Cubas, 1981; Antolí y García-Cubas, 1985), las especies de moluscos ocupan diversos habitat y presentan distintos tipos de nutrición, como se indica en la lista sistemática, observándose que la forma de nutrición más frecuente en los bivalvos correspondió a la filtración de partículas orgánicas en suspensión, presente en 9 de las 10 especies identificadas (90%), con una sola especie (10%) recolectora de materia orgánica depositada. Respecto a su relación con el sustrato, 5 especies (50%) son epifaunales y un número igual infaunales. En los gasterópodos el tipo de nutrición dominante fue el herbívoro raspador, registrado en 7 de las 13 especies identificadas (53.8%), le siguen en orden de importancia las formas carnívoras con 3 especies (23%), los ectoparásitos con 2 especies (15.4%) y una especie que se alimenta de materia orgánica depositada (7.7%). Por lo que se refiere a su posición respecto al sustrato, 12 especies (92.3%) resultaron epifaunales y sólo una (7.7%) seminfaunal. Sin embargo, tanto los gasterópodos como los bivalvos pueden presentar diversas alternativas alimenticias y de habitat. Estructura de la comunidadEl análisis de la estructura comunitaria a partir de los índices de diversidad (H'n), diversidad máxima (Hmax), equitatividad (J) y riqueza de especies (D), se llevó a cabo tanto de manera general, considerando la suma de organismos vivos y muertos, como en forma particular, tomando en cuenta sólo los ejemplares recolectados vivos, debido a que las diferencias entre los valores del índice de diversidad de Shannon-Wiener (1963), calculado para las Clases Gastropoda y Bivalvia así como para el Phylum Mollusca fueron estadísticamente significativas (t = 15.05; 57.38; 33.01, respectivamente; P > 0.001), al comparar los resultados de la comunidad total (vivos + muertos) con los obtenidos de la comunidad viva. 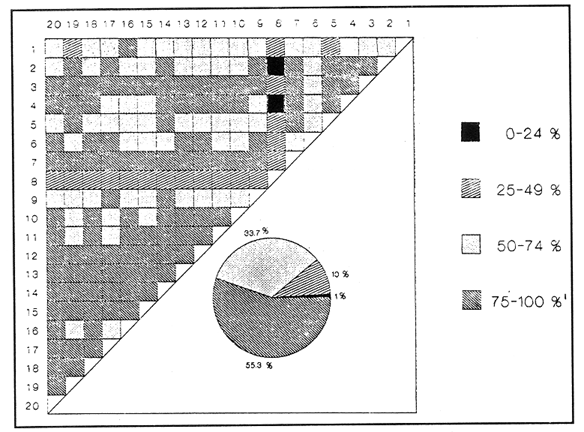 Figura 3. Diagrama de enrejado de la afinidad de la Clase Gastropoda en las estaciones de muestreo. 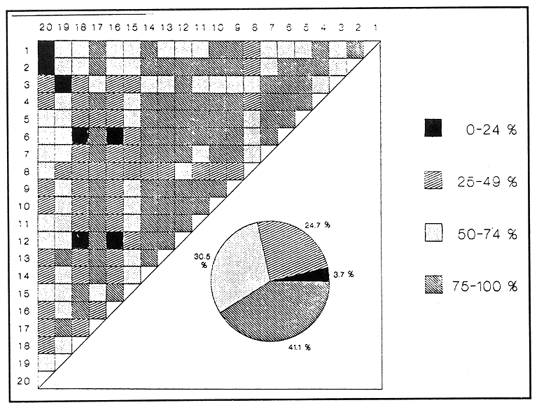 Figura 4. Diagrama de enrejado de la afinidad de la Clase Bivalvia en las estaciones de muestreo. 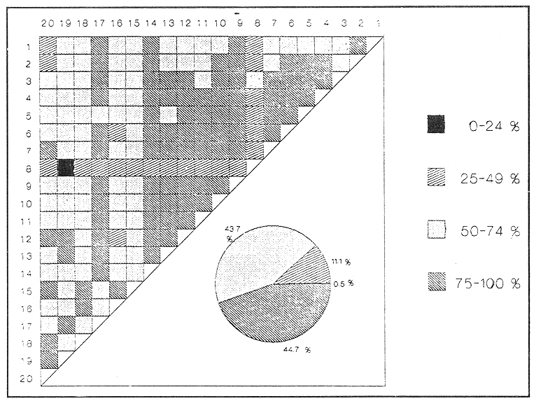 Figura 5. Diagrama de enrejado de la afinidad del Phylum Mollusca en las estaciones de muestreo. En el primer caso, los valores más altos se observan en la Clase Gastropoda, con excepción de la riqueza específica, la cual es mayor a nivel de Phylum; mientras que en la comunidad viva, los índices más elevados corresponden al Phylum, salvo el de equitatividad, el cual es superior para la Clase Bivalvia (Tabla 4; Figs. 6 y 7). Asimismo, estos parametros presentaron fluctuaciones notables en las distintas estaciones de muestreo, encontrándose que en el análisis general (vivos + muertos), la diversidad (bits/individuo) se determinó en un intervalo de 0.31 a 2.37 para la Clase Gastropoda, de 0.06 a 2.17 para la Clase Bivalvia y de 0.33 a 2.84 para el Phylum Mollusca, con medias de 1.51, 0.90 y 1.71, respectivamente. La equitatividad varió de 0.31 a 0.92, de 0.06 a 0.72 y de 0.17 a 0.77; mientras que la riqueza específica registró valores en intervalos de 0.22 a 0.74, de 0.10 a 0.98 y de 0.34 a 1.60, según el conjunto analizado (Tabla 5; Figs. 8- 10). En cambio, para la comunidad viva, la diversidad mostró resultados en un intervalo de 0 a 0.92 para la Clase Gastropoda, de 0 a 1.92 para Bivalvia y de 0 a 2.31 para el Phylum, con medias de 0.14, 0.90 y 1.16, respectivamente; la equitatividad varió de 0 a 0.92 para gasterópodos y de 0 a 1, tanto para bivalvos como para el total de moluscos vivos, y la riqueza específica registró valores de 0 a 0.41, de 0 a 0.56 y de 0 a 0.96, de acuerdo con cada taxón (Tabla 6; Figs. 11- 13). 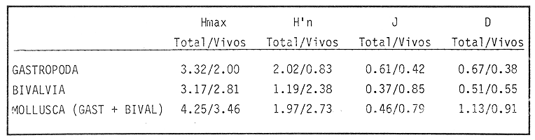 TABLA 4 ÍNDICES DE DIVERSIDAD MÁXIMA (Hmáx), DIVERSIDAD (H'n), EQUITATIVIDAD (J) Y RIQUEZA DE ESPECIES (D) DE LAS COMUNIDADES MALACOLÓGICAS TOTAL Y VIVA 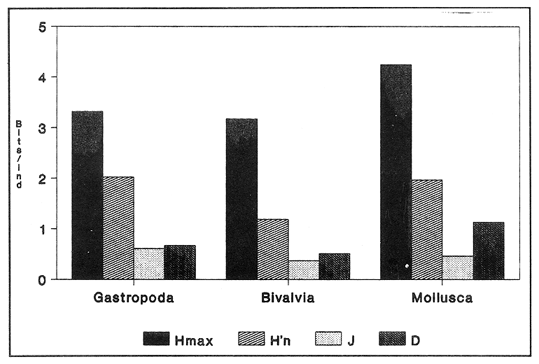 Figura 6. Índices de divenidad máxima (Hmáx), diversidad (H'n), equitatividad (1) y riqueza de especies (D) para la comunidad malacológica total (vivos+muertos). 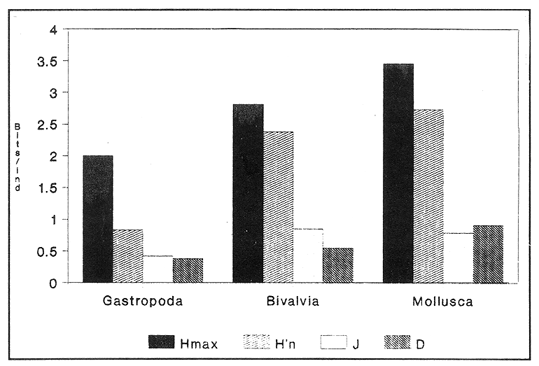 Figura 7. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (1) y riqueza de especies (D) para la comunidad malacológica viva. 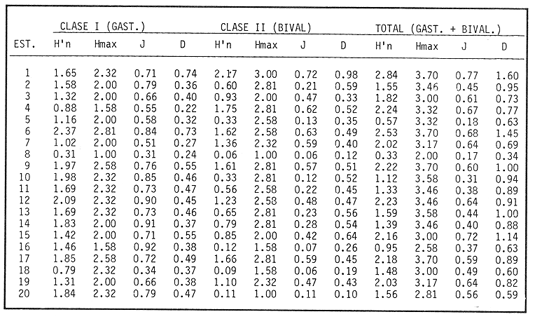 TABLA 5 ÍNDICES DE DIVERSIDAD (H'n), DIVERSIDAD MÁXIMA (Hmáx), EQUITATIVIDAD (J) Y RIQUEZAS DE ESPECIES (D) DE LA COMUNIDAD MALACOLÓGICA TOTAL EN CADA ESTACIÓN DE MUESTREO 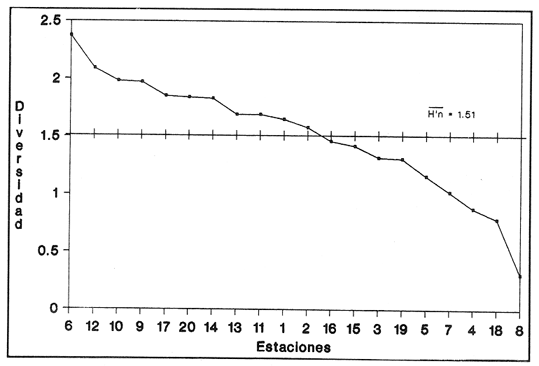 Figura 8. Índices de diversidad de Clase Gastropoda (vivos + muertos) en cada estación de muestreo 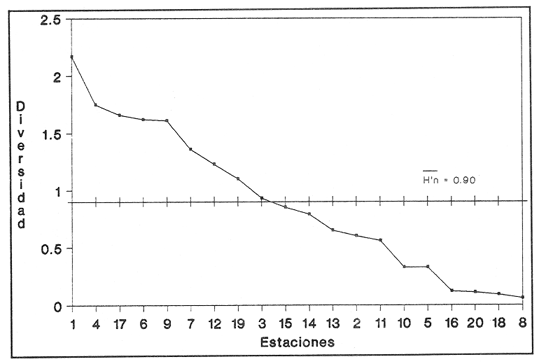 Figura 9. Índices. de diversidad de la Clase Bivalvia (vivos + muertos) en cada estación de muestreo. 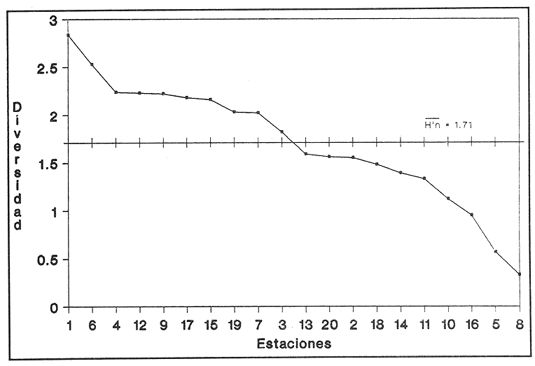 Figura 10. Índices de diversidad del Phylum Mollusca (vivos + muertos) en cada estación de muestreo. Por otra parte, los coeficientes de determinación (R²) entre H'nvs. J y H'n vs. D señalan que la equitatividad tuvo mayor influencia sobre la diversidad calculada para la comunidad total (Fig. 14 A-F). Sin embargo, en el análisis de vivos, los coeficientes muestran variaciones importantes en los taxaestudiados e indican que la equitatividad determina notablemente la diversidad de gasterópodos; que la equitatividad y la riqueza de especies ejercen una influencia similar en la diversidad de bivalvos, y que la riqueza específica es el factor que influye en mayor grado sobre la diversidad del Phylum (Fig. 15 A-F). El comportamiento espacial de la diversidad se determinó, también, con base en su variación por ambiente halino (Tabla 7; Figs. 16 y 17), observándose que en el análisis general (vivos + muertos), los valores más bajos corresponden a la Clase Bivalvia y a la zona limnética; mientras que en los ambientes oligohalino y mesohalino "A" los índices obtenidos fueron semejantes, con variaciones por taxón. No obstante, la comunidad viva es más diversa en ambiente oligohalino, registrándose valores de cero en los índices de la Clase Gastropoda para las zonas limnética y mesohalina "A"; mientras que los resultados entre la Clase Bivalvia y el Phylum Mollusca son muy cercanos y las diferencias se presentan entre cada uno de los ambientes. 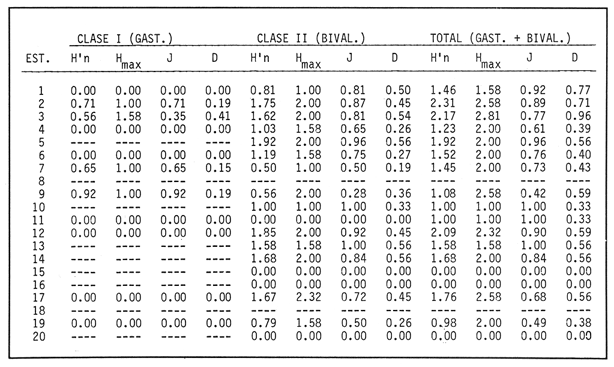 TABLA 6 ÍNDICES DE DIVERSIDAD (H'n), DIVERSIDAD MÁXIMA (Hmáx), EQUITATIVIDAD (J) Y RIQUEZAS DE ESPECIES (D) DE LA COMUNIDAD MALACOLÓGICA VIVA EN CADA ESTACIÓN DE MUESTREO 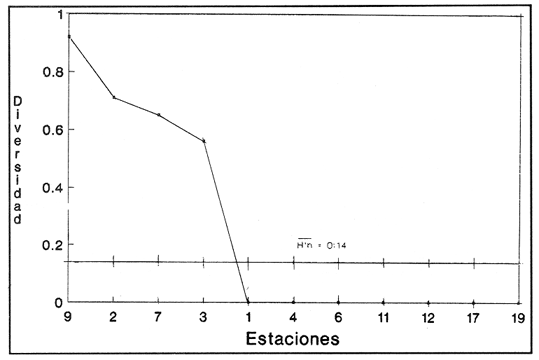 Figura 11. Índices de diversidad por estación para gasterópodos vivos. 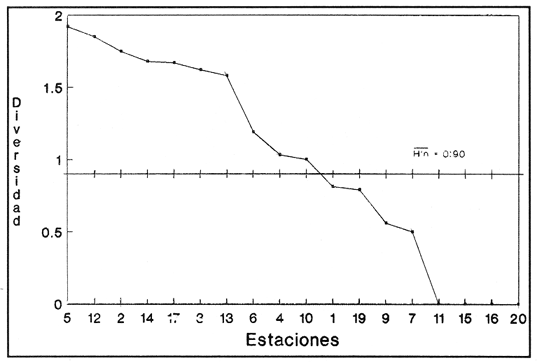 Figura 12. Índices de diversidad por estación para bivalvos vivos. 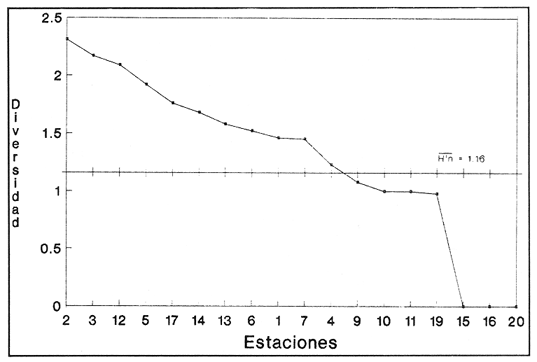 Figura 13. Índices de diversidad por estación para moluscos vivos. 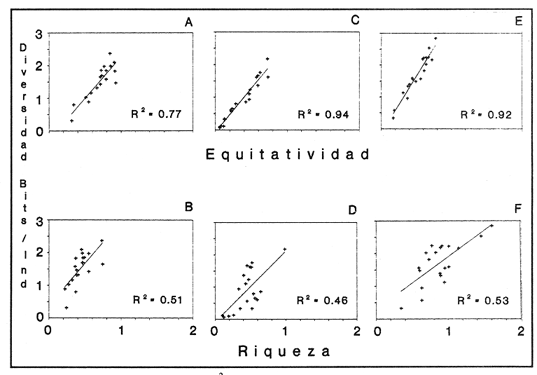 Figura 14. Coeficientes de determinación (R 2) entre la diversidad de la comunidad total (vivos + muertos) y los componentes de equitatividad y riqueza de especies. Clase Gastropoda: A = H'n vs. J; B = H'n vS. D. Clase Bivalvia: C = H'n vs. J; D = H'n vs. D. Phylum Mollusca: E = H'n vs. J; F = H'n vs. D. 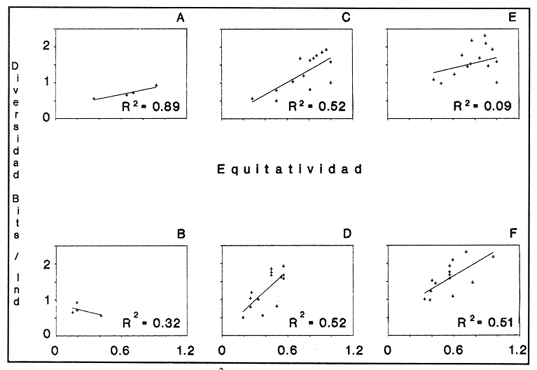 Figura 15. Coeficientes de determinación (R) entre la diversidad de la comunidad viva y los componentes de equitatividad y riqueza de especies. Clase Gastropoda: A = 1 H'n vs. J; B = H'n vs. D. Clase Bivalvia: C H'n vs. J; D = H'n vs. D. Phylum Mollusca: E = H'n vs. J; F = H'n vs. D. 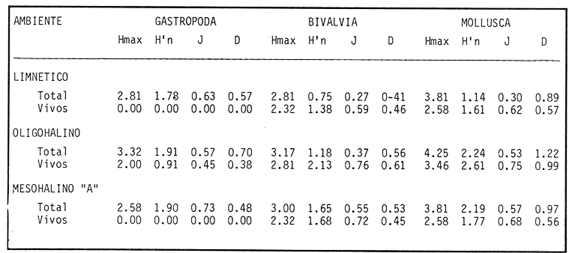 TABLA 7 ÍNDICES DE DIVERSIDAD (H'n), DIVERSIDAD MÁXIMA (Hmáx), EQUITATIVIDAD (J) Y RIQUEZAS DE ESPECIES (D) PARA LAS COMUNIDADES MALACOLÓGICAS TOTAL Y VIVA, POR AMBIENTE HALINO 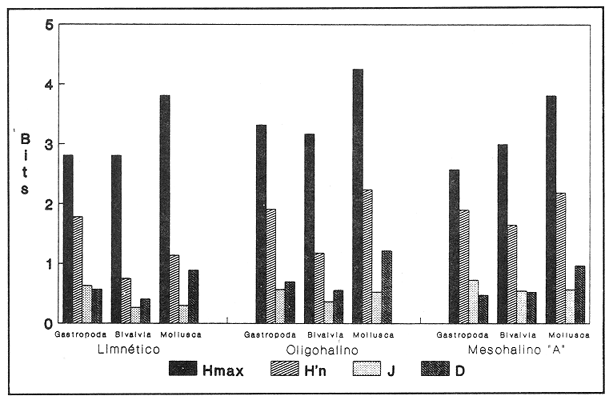 Figura 16. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D) de la comunidad malacológica total (vivos + muertos), por ambiente halino. 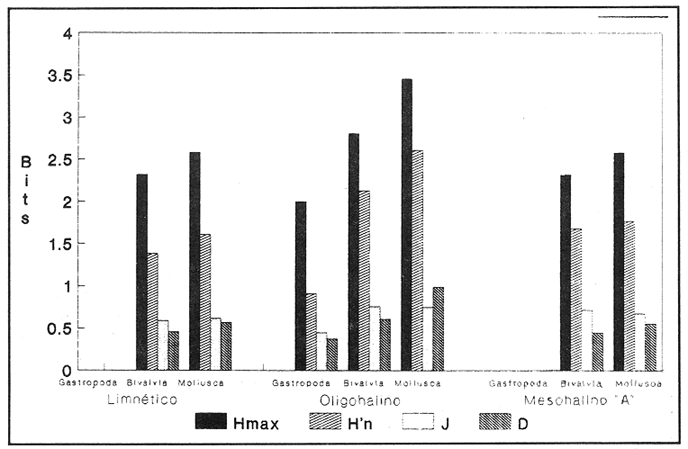 Figura 17. Índices de diversidad máxima (Hmáx), diversidad (H'n), equitatividad (J) y riqueza de especies (D) de la comunidad malacológica viva, por ambiente halino. ConclusionesDISCUSIÓN Y CONCLUSIONESLos resultados de temperatura y profundidad obtenidos concuerdan con los reportados por Reséndez (1973) en verano, para el área de estudio. En lo que respecta a la distribución horizontal de la salinidad, se observa un comportamiento similar al que señala Vargas Maldonado (1986) para época de lluvias, predominando el ambiente oligohalino (0.5-5° %), debido al efecto de mezcla en la columna de agua, producido por la precipitación y el creciente aporte de los ríos en esa temporada. Cabe señalar, sin embargo, que el perfil halino del área en el que se diferenciaron 3 ambientes, no pudo ser complementado con la caracterización faunística para cada uno de ellos, pues con excepción de las formas marcadamente dulceacuícolas (Pyrgophorus spinosus y Pomacea flagellata), las especies de moluscos que conforman la comunidad malacológica total (vivos + muertos) son en su mayoría de ambientes salobres y muestran tolerancia a variaciones de salinidad, encontrándose distribuidas indistintamente desde ambientes mesohalinos hasta francamente limnéticos. No obstante, si se considera sólo la comunidad viva, se observan diferencias en la abundancia y distribución local de las 11 especies que la integran. Así, aunque todas tienen representantes en el ambiente oligohalino, sóloNeritina virginea, Hydrobia totteni y Littoridina sphinctostoma se encontraron restringidas a él, sin ser las más abundantes. En la zona limnética destacó la presencia de Brachidontes exustus y Mytilopsis leucophaeata; mientras que en la mesohalina "A" predominaron Rangia flexuosa y Tagelus plebeius, aún cuando no son formas exclusivas de esos ambientes. Por tanto, puede afirmarse que la mayoría de las especies con representantes vivos en el sistema lagunar de Alvarado son eutópicas y eurihalinas para salinidades de 0 a 10 %. Por lo que se refiere a transparencia del agua, se apreciaron grandes variaciones en el sistema lagunar. Así, los porcentajes más bajos de transparencia (16% en promedio) se registraron en las estaciones 4, 5 y 6, situadas en la porción que une a las Lagunas de Alvarado y Tlalixcoyan, en donde hay gran cantidad de materiales acarreados por los ríos circundantes, cuya influencia es notable hasta la zona comprendida entre Punta Grande y Boca de Tragadero (22%). Las aguas más claras son las de Buen País (36.6%), debido a que la abertura de un canal artificial, que comunica a la Laguna Camaronera con el mar, permite el flujo de agua cuyo patrón de circulación evidente es hacia Buen País y la región noroccidental de la Laguna de Alvarado. No obstante, el porcentaje promedio de transparencia en el área de estudio fue de 28%, difiriendo del que indica Vargas Maldonado (1986) para la misma época del año (16.5%), cuestión que puede ser atribuida a que su trabajo se refiere a todo el complejo lagunar y es bien sabido que la Laguna de Tlalixcoyan presenta una elevada turbiedad, que seguramente influyó en el valor promedio. Por otra parte, la mayor abundancia de los bivalvos no se relaciona, en este caso, con el criterio adoptado para su cuantificación; pues aún cuando cada valva desarticulada se consideró como un ejemplar, si el total de bivalvos recolectados se divide entre dos, éste duplica al de los gasterópodos. Así, el predominio de la Clase Bivalvia quizá se deba a que los sustratos blandos característicos del área favorecen su presencia. Es importante mencionar, también, que las estaciones 17, 9 y 19, en las que se registró el número más elevado de organismos capturados vivos, han sido caracterizadas por Raz-Guzmán et al. (en preparación) como localidades en las que predominan sedimentos limo-arcillosos con alto contenido de materia orgánica y carbonatos, en donde hay aportes de carbono orgánico que derivan de vegetación sumergida (Ruppia maritima), como ocurre en las estaciones 9 y 19, o bien de la vegetación circundante (Rhizophora mangle), como sucede en la estación 17. Las diferencias estadísticas encontradas entre los índices de diversidad calculados para la comunidad viva y el total de moluscos, incluyendo exoesqueletos, podrían inducir a pensar en una falta de representatividad de los remanentes esqueléticos en estudios ecológicos. Sin embargo, este aspecto ha sido abordado por diversos autores (Valentine, 1961; Ager, 1963; Johnson, 1964; Johnson, 1965; Craig, 1966; Wilson, 1967; Warme, 1969, 1971; Garcia-Cubas, 1973), quienes coinciden en afirmar que hay suficientes líneas de evidencia que indican que las conchas depositadas en los fondos blandos no consolidados, predominantes en las lagunas costeras, reflejan adecuadamente a lostaxarecolectados vivos, por ser insignificante su transportepost-mortem. Asimismo, las diferencias en los índices de diversidad podrían deberse al tipo de muestreo efectuado pues, como ya se señaló, no se obtuvieron muestras de los organismos que se encuentran constituyendo bancos, debido a que el objetivo del proyecto general, en el que se incluye este trabajo, contempla el reconocimiento de toda la fauna malacológica que habita en las lagunas litorales y no sólo de la comunidad estable, la cual ya está bien identificada por soportar pesquerías en esas áreas y que está integrada por especies de ostión (Crassostrea virginica) y almejas (Rangia cuneata, R. flexuosa y Polymesoda caroliniana). De manera que con la metodología empleada se asegura la extracción de las especies de moluscos presentes en el sistema lagunar, aunque se subestiman la abundancia y frecuencia de la comunidad viva característica de estos cuerpos de agua; la cual, dado su carácter gregario, habría sido capturada en grandes cantidades con un sólo lance de draga. Por otra parte, la diversidad de gasterópodos vivos se sustenta en una comunidad integrada por sólo 4 especies, que difieren notablemente en abundancia y distribución, reflejándose en valores bajos de equitatividad para esa Clase. En cambio, la comunidad de bivalvos se compone de 7 especies con representantes vivos, ampliamente distribuidas y con abundancias más cercanas entre sí, por lo que la mayor equitatividad de esta Clase influyó también en los valores de diversidad obtenidos a nivel de Phylum. Las localidades en las que se observaron los mayores registros de diversidad específica para los conjuntos analizados se encuentran influídas por descargas de los ríos (estaciones 2, 3, 5 y 6), o bien se trata de áreas asociadas a presencia de vegetación sumergida y/o circundante (estaciones 9 y 17). Así, el incremento en el flujo de agua de origen fluvial durante la temporada de lluvias provoca a su vez un aumento de la productividad en el área (Villalobos et al., 1975) y de la cantidad de materia orgánica suspendida; lo cual, aunado a la presencia de pastos parvozostéridos y algunas especies de algas (García-Montes, 1989), provee de alimento, sustrato y refugio a moluscos filtradores y herbívoros raspadores. El cálculo de los coeficientes de determinación entre H'nvs. J y H'n vs. D permitió identificar a la equitatividad como el factor que controla la diversidad de gasterópodos vivos y de la comunidad malacológica total (vivos + muertos), indicando que los parámetros de dominancia son más importantes que la riqueza específica; mientras que en las poblaciones vivas de la Clase Bivalvia y el Phylum Mollusca, la adición de especies a la comunidad y la dominancia ejercen una influencia similar sobre la diversidad. El análisis espacial revela una estabilidad relativa en la composición de la comunidad malacológica para los ambientes oligohalino y mesohalino "A", característicos de sistemas estuarinos, lo cual sugiere que, en esa época del año, la heterogeneidad estructural del habitat podría estar regulada por la variación de las praderas deRuppia maritima y la presencia de ficoflora estacional, más que por el gradiente de salinidad. Es importante mencionar, finalmente, que dentro de la fauna característica del área se encuentran especies de moluscos que podrían ser utilizadas más ampliamente como un recurso alimenticio. Tal es el caso de las almejas Rangia flexuosa y Polymesoda caroliniana y de los mejillones Ischadium recurvum y Brachidontes exustus. AgradecimientosLos autores agradecen a Xavier Chiappa Carrara su valiosa ayuda en el manejo estadístico de los datos, elaboración de material gráfico y revisión del manuscrito; a Iván Proaño Muciño y Gerardo Barrientos Mac Gregor por el desarrollo y aplicación de un programa de cómputo para el diseño de diagramas de enrejado; a Felipe Flores Andolais, Vicente Antolí Figuerola y Francisco Escobar de la Llata por su participación en la recolecta del material biológico. LITERATURAABBOTT, R. T. American seashells. The marine Mollusca of the Atlantic and Pacific coast of North America. Van Nostrand Reinhold Co.2a. ed.New York1974666 p. AGER, D. V. Principles of Paleoecology. Ed. Mac Graw Hill New York1963371 p. ANDREWS, J. Sea shells of the Texas coast.University of Texas PressAustin1971 298 p. ANTOLÍ, V. y A. GARCÍA-CUBAS An. Inst. Cienc. del Mar y Limnol.Sistemática y ecología de los moluscos en las lagunas costeras Carmen y Machona, Tabasco, México.Univ. Nal. Autón. México1985145-19812(1) BASHARIN, G. P. Theory Probability Appl.On a statistical estimate for the entropy of a sequence of independent variables.1959333-3364 CRAIG, G. W. Concepts in Paleoecology.Earth Sciences Rev.1966127-1552 CONTRERAS, F. Las lagunas costeras mexicanas.Centro de Ecodesarrollo, Secretaría de Pesca, D. F.1985253 p. FERNÁNDEZ BUCES, N. Tesis Profesional.Variación poblacional de Discapseudes holthuisi (TANAIDACEA) (Bacescu y Gutu, 1975) en la laguna de Alvarado, Veracruz.Facultad de Ciencias, UNAM198997 p. FLORES-COTO, C. y L. MÉNDEZ VARGAS An. Inst. Cienc. del Mar y Limnol.Contribución al conocimiento del ictioplancton de la Laguna de Alvarado, Veracruz.Univ. Nal. Autón. México1982141-1609(1) GARCÍA-CUBAS, A.Sistemática y distribución de los micromoluscos recientes de la Laguna de Términos, Campeche, México.Bol. Inst. Geol. Univ. Nal. Autón. México19631-55, 24 figs., 4 láms.67(4) GARCÍA-CUBAS, A.Ecología y distribución de los micromoluscos recientes de la Laguna Madre, Tamaulipas, México. Bol. Inst. Geol. Univ. Nal. Autón. México19681-44, 15 figs., 8 láms.86 GARCÍA-CUBAS, A.Ecología y distribución de los micromoluscos recientes de la Laguna de Tamiahua, Veracruz, México. Bol. Inst. Geol. Univ. Nal. Autón. México19691-53, 32 figs., 11 láms.93 GARCÍA-CUBAS, A.Tesis Doctoral.Ecología y distribución de los micromoluscos de tres lagunass costeras del Golfo de México.Facultad de Ciencias, UNAM, México1973257 p., 10 Láms. GARCÍA-CUBAS, A.An. Inst. Cienc. del Mar y Limnol. Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de Términos, Campeche).Univ. Nal. Autón. México19811-182Publ. Esp. 5 GARCÍA-CUBAS, A.Ecología de los Ecosistemas Costeros en el Sur del Golfo de México: La región de la Laguna de Términos.Características ecológicas de los moluscos de la Laguna de Términos, Cap. 16:277-304. Yánez-Arancibia, A. y J. W. Day Jr. Inst. Cienc.del Mar y Limnol. UNAM. Coast. Ecol. Inst. LSU. Editorial UniversitariaMéxico, D. F.1988518 p. GARCÍA, E. An. Inst. Biol. Univ. Nal. Autón. México. Los climas del Estado de Veracruz (según el sistema de clasificación climática de Köppen modificado por la autora).Ser. Botánica19713-4241 (1) GARCÍA-MONTES, J. F.Tesis de Maestría Composición, distribución y estructura de las comunidades de macroinvertebrados epibénticos del sistema lagunar Alvarado, Veracruz.UACPyP-CCHUNAM1989124 p. GÓMEZ-AGUIRRE, S.Observaciones comparativas de resultados de estudios del placton de lagunas costeras del Golfo de México. Mem. II Simp. Lat-Amer, Oceanogr. Biol. Univ. de OrienteCumanáVenezuela197719-33124-28 nov. 1975 HUTCHESON, K. A test for comparing diversities based on the Shannon formula. J. Theoret. Biol.1970151-15429 JOHNSON, R. G. The community approach to PaleoecologyApproaches lo Paleoecology. Imbrie, J. y N. D. Newell. Wiley New York1964432 p. JOHNSON, R. G. Pelecypod death assemblages of Tomales Bay, California.Jour. Paleont.196580-8539 KREBS, J. C. Ecology The experimental analysis of distribution and abundance. Harper International EditionNew York1972694 p. LLOYD, M., J. H. ZAR y J. R. KARR On the calculation of information-theoretical measures of diversity. Amer. Midland Natur.1968257-27279 MARGALEF, D. R.Information theory in ecology. General systems. Transl. from Mem. Real Acad. Cienc. Arts.Barcelona1958373-44932 NEWELL, N. D.Classification of the Bivalvia.Amer. Mus. Nov.19651-24, figs. 1-3.(2206) PIELOU, E. C. Ecological diversity. Wiley New York1975165 P. PURCHON, R. D. The Biology of the Mollusca. New YorkPergamon Press1968560 p. RAZ-GUZMÁN, A., G. DE LA LANZA y L. A. SOTO Fuentes, distribución y 13C de la materia orgánica sedimentaria y detrito, y 13C de la vegetación de la Laguna de Alvarado, Veracruz. (En preparación)120 p. RESÉNDEZ, M. A.Estudio de los peces de la Laguna de Alvarado, Veracruz, México. Rev. Soc. Mex. Hist. Nat.1973183-28134 SHANNON, E. C. y N. WIENER The mathematical theory, of comunication.University of Illinois Press, Urbana1963117 p. SIGNORET, P. M. Tesis Profesional.Contribución al conocimiento de las medusas de las lagunas de Tamiahua y Alvarado, Ver., México. Fac. de Ciencias, Univ. Nal. Autón. México196991 p. THIELE, J. Handbuch der Systematischen Weichtierkunde.Jena1934-1935779-1154, figs. 784-7892 VALENTINE, J. W.Paleoecological molluscan geography of the Californian Pleistocene. Univ. Calif. Publ. Geol. Sci.1961309-34234 VÄLIKANGAS, I.Über die Biologie der Ostsee als Brackwassergebiet. Verh. int. Ver. theor. u. angew. Limnologie193362-1126 VARGAS MALDONADO, I.Estudios ecológicos básicos de las Lagunas costeras del Golfo de México. Estudio ecológico de la ictiofauna de la Laguna de Alvarado, Veracruz. Estructura de las comunidades de peces para las épocas de nortes y secas. Informe Técnico.PCECBNA/UAM IZTAPALAPA/CONACYT.1986 VILLALOBOS, A., J. A. SUÁREZ, S. GÓMEZ, G. DE LA LANZA, M. ACEVES, F. MANRIQUE y J. CABRERAConsiderations on the hidrography and productivity of Alvarado Lagoon, Veracruz, México. Proc. Gulf. Carib. Fish. Inst. Nineteenth Anual Sess.196675-85 VILLALOBOS, A., J. CABRERA, F. MANRIQUE, S. GÓMEZ, V. ARENAS y G. DE LA LANZA Lagunas Costeras un Simposio.Relaciones entre postlarvas planctónicas de Penaeus sp. y caracteres ambientales en la Laguna de Alvarado, Veracruz, México. Ayala-Castañares, A. y F.G. Phleger Mem. Simp. Intern. Lagunas Costeras, UNAM-UNESCOMéxico1969601-620Nov. 28-30,1967 VILLALOBOS, A., S. GÓMEZ, V. ARENAS, A. RESÉNDEZ y G. DE LA LANZA An. Inst. Biol. Univ. Nal. Autón. México Estudios hidrobiológicos en la Laguna de Alvarado (Febrero-Agosto, 1966). Ser. Zool.19751-341 WARME, J. E. Live and dead molluscs in a coastal lagoon.Jour. Paleont.1969141-15043 WARME, J. E.Paleoecological aspects of modem coastal lagoon.Univ. Calif. Publ. Geol. Sci.19711-13187 WILSON, J. B.Paleoecological studies on shells beds and associated sediments in the Solway. Firth Scottish Jour. Geology1967329-3713 ZAR, J. H.Biostatistical analysis.Prentice-HallLondon1974 620 P. LÁMINAS
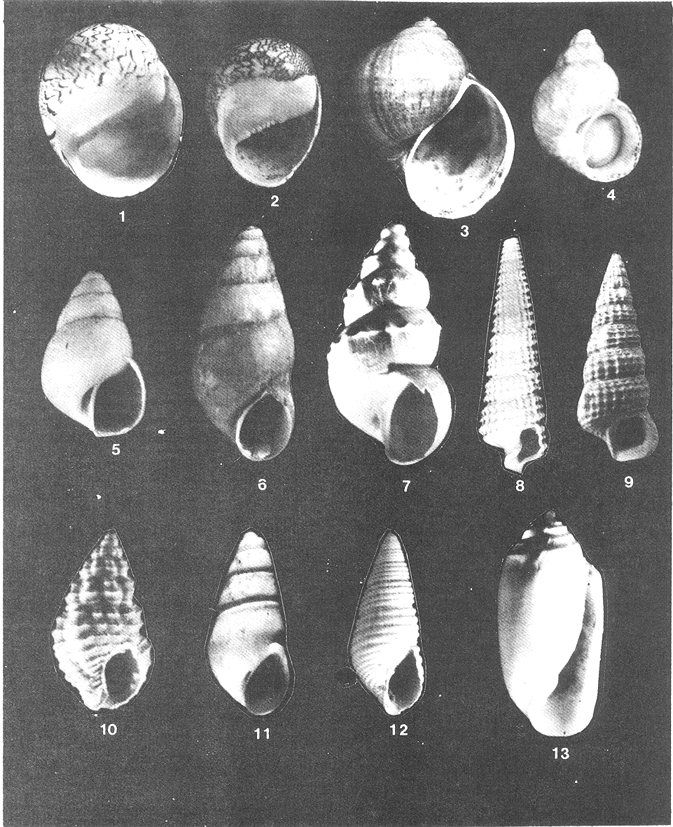 Lámina 1. Fig. 1: Neritina reclivata. Fig. 2: Neritina (Vitia) virginea. Fig. 3: Pomacea flagellata. Fig. 4: Cingula (Microdochus) floridanus. Fig. 5: Hydrobia totteni. Fig. 6: Littoridina(Texadina) sphinctostoma. Fig. 7: Pyrgophorus spinosus. Fig. 8: Cerithiopsis (Laskeya) emersoni. Fig. 9: Cerithidea (Cerithideopsis) pliculosa. Fig. 10: Nassarius (Nassarius) acutus. Fig. 11: Odostomia weberi. Fig. 12: Odostomia (Menestho) impressa. Fig. 13: Acteocina canaliculata. 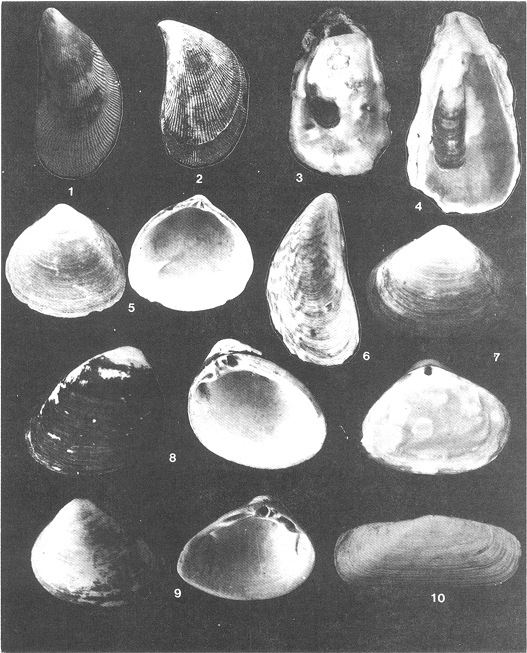 Lá¡mina 2. Fig. 1: Brachidontes (Hormomya) exustus. Fig. 2: Ischadium recurvum. Fig. 3: Crassostrea virginica. Fig. 4: Crassostrea rhizophorae. Fig. 5: Polymesoda caroliniana. Fig. 6: Mytilopsis leucophaeata. Fig. 7: Mulinia lateralis. Fig. 8: Rangia cuneata. Fig. 9: Rangia (Rangianella) flexuosa. Fig. 10: Tagelus (Mesopleura) plebeius.
|