|
ESTRUCTURA DE
TALLAS, RECLUTAMIENTO Y CRECIMIENTO DEL MEJILLÓN Módiolus capax
(CONRAD) (BIVALVIA-MYTILIDAE) EN LA BAHÍA DE LOS ANGELES, BAJA
CALIFORNIA, MÉXICO
Trabajo recibido el 26 de enero de 1989 y
aceptado para su publicación el 3 de noviembre de 1989.
María del Carmen Garza-Aguirre
Luis Fernando Bückle-Ramírez
Centro de
Investigación Científica y de Educación Superior de
Ensenada (C.I.C.E.S.E.), Av. Espinoza 843, Ensenada, Baja California,
México.
En la bahía de Los
Angeles Baja California, se estudió la estructura de tallas y el
crecimiento deModiolus capax (CONRAD) (BIVALVIA-MYTILIDAE)
desde febrero de 1985 hasta febrero de 1986. La estructura de tallas de la
población de dos estaciones de muestreo denotó que los organismos
de longitudes anteroposteriores mayores de 50 mm en la estación 1 y 75
mm en la estación 2 constituyeron generalmente más del 75%. El
asentamiento de los juveniles en la población no fue abundante. En
agosto de 1985 se observó por primera vez un número significativo
de organismos recién fijados y durante los meses siguientes, hasta
febrero de 1986, se captó el efecto acumulativo de los nuevos reclutas
que fue menos del 19% de la población estudiada. El estudio del
crecimiento de M. capax, durante un año, determinado
a partir de organismos marcados (1.29 mm/mes) y del movimiento de la moda de
los animales recién fijados (1.22 mm/mes) revelan que crece lentamente.
La velocidad de crecimiento medida durante nueve meses en los reclutas con una
longitud anteroposterior promedio inicial de 5 mm, disminuye de 1.83 a 0.75
mm/mes. Lo señalado debe considerarse antes de iniciar cualquier forma
de explotación de las poblaciones de esta especie en el área
estudiada. El crecimiento relativo de los organismos indicó que los
más grandes tienden a adquirir formas más cilíndricas
debido a un incremento relativamente más rápido del ancho de la
concha con respecto al alto.
Size structure,
recruitment and somatic growth rate of the musselModiolus
capax (CONRAD) (BIVALVIA-MYTILIDAE) in bahía de Los Angeles,
Baja California, México. Studies carried out at two sites of
bahía de los Angeles, from February 1985 to February 1986, showed that
in one of these approximately 75% of the Modiolus capax
population was made up by individuales with a total shell length of more than
50 mm, and that the other one had a similar precentage of even larger organisms
( 75 mm). Recruitment was generally low: significant new settlement was first
noticed in August 1985, and continued throughout the period of Observations,
its commulative effect resulting in less than 19% of the total final
population. Somatic growth rate was measured directly in marked specimens (1.29
mm/month), and determined from the progressive shift of the modal shell
measurement values of the new recruits (1.22 mm/month), The data show that this
la a slow-growing organism. These biological Characteristics should be taken
into considerafion before planning any lype of explotation of this species in
the study area. Other measurements indicate that the intervalvar distance tends
to increase with age more rapidly than valve length, conferring a more
cylindrical shape to aged specimens.
En México hay muchas especies de moluscos bivalvos de importancia económica de los cuales algunos como Pinna rugosa, Atrina tuberculosa, Atrina maura, Argopecten circularis, Liropecten subnodosus, Tivela stultorum son intensamente explotados (Baqueiro, 1984). La insuficiencia del manejo de las poblaciones ha sido el resultado del desconocimiento de aspectos biológicos y ecológicos que ha generado el agotamiento de algunas poblaciones como Pteria sterna truncando en consecuencia el desarrollo de biotecnologías modernas. Con ésta y otras contribuciones realizadas anteriormente, se espera evitar que en las poblaciones de Modiolus capax que recientemente han comenzado a explotarse en el Golfo de California, suceda lo mismo, ya que los abundantes bancos, de fácil acceso y captura, están expuestos a la sobrepesca si la explotación se intensifica sin la debida regulación. Las investigaciones referentes a la biología de M. capax son recientes y escasas. Se ha estudiado la actividad reproductiva de las poblaciones de la bahía de la Paz, Baja California Sur (Ochoa-Báez, 1985), y de la bahía de los Angeles, B.C. (Garza y Bückle, 1989); el desarrollo embriológico y larval en condiciones de laboratorio (Orduña-Rojas, 1986) y las diferentes respuestas fisiológicas a los efectos interactivos de la temperatura y el alimento (Rico-Mora, 1987); en el laboratorio se investigó la extensión de la actividad reproductiva durante hasta 60 días adicionales al ciclo natural de la especie (Espinoza-Peralta, 1989) y en condiciones naturales Aguirre-Hinojosa (1987) analizó las preferencias en la fijación de las larvas sobre diferentes sustratos artificiales siguiendo el crecimiento y la sobrevivencia de los reclutas; y Serrano-Guzmán, (en preparación) estudió la abundancia y la distribución de las larvas en un periodo de primavera-verano. El propósito de este estudio realizado en la bahía de Los Angeles, es de contribuir al conocimiento de la biología de M. capax a través del análisis de la estructura de tallas y el crecimiento de la población durante un año. MATERIALES Y MÉTODOSLos mejillones se colectaron en la bahía de los Angeles, localizada en la región Oeste del Golfo de California, en la intersección de los 29º de Lat. Norte y los 113º 30' de Long. Oeste. En esta área se ubicaron dos estaciones de muestreo en la zona intermareal; la estación 1 quedó situada en la parte noroeste, frente al campamento del Programa de Estudio de la Tortuga Marina de la Secretaría de Pesca, y la estación 2 en la zona conocida como El Rincón, al sureste de la Bahía (Fig.1). Las dimensiones de las dos estaciones se establecieron arbitrariamente y ambas se fijaron a una longitud de 200 m, paralelos a la línea de costa, con un ancho aproximado de 30 m en la estación 1 y de 15 m en la estación 2, lo cual corresponde al ancho de la franja inferior del intermareal que está la mayor parte del tiempo inmersa en el agua y queModiolus capax ha colonizado densamente. Las colectas de los organismos se hicieron en febrero y marzo de 1985 en la estación 1 y a partir de abril de 1985 en las dos estaciones, alternando una cada mes (a excepción del mes de julio) hasta febrero de 1986. Las muestras se tomaron al azar con 10 cuadrantes de 50 X 50 cm colectando todos los mejillones incluidos en ellos. A cada individuo se le midió la longitud anteroposterior, el alto, el ancho y el peso húmedo total al 0.1 mm o gr más cercano. La distribución de frecuencia de tallas se hizo con los datos de la longitud anteroposterior. 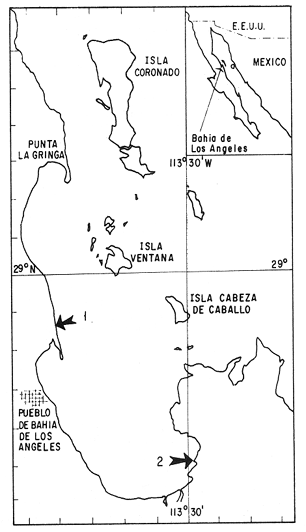 Figura 1. Ubicación geográfica de la Bahía de Los Ángeles y de las estaciones de muestreo. 1: estación 1; 2: estación 2. (Tomado de Barnard y Grady, 1968). Para el estudio del crecimiento absoluto se utilizó el método de análisis de la distribución de tallas (Cassie, 1954). También se trabajó con organismos marcados para lo cual desde marzo de 1985 hasta febrero de 1986 se retuvieron cinco grupos de mejillones de diferentes tallas iniciales en ocho cajas experimentales de madera (50 X 50 X 7.5 cm) con bases de malla anchovetera y tapas de malla de plástico de 1 cm de luz. Los grupos confinados se formaron en base a las clases de tamaño encontradas en febrero de 1985, y se distribuyeron de la siguiente forma: 5 organismos entre 0-43 mm, y 10 de cada uno de los siguiente intervalos, 43-61 mm, 61-80 mm 80-95 mm y mayores de 95 mm. El primer intervalo es el reagrupamiento de las primeras dos clases de tamaño (0-23 y 23-43 mm), debido a que se encontraron muy pocos mejillones de estas tallas en el banco natural. Las cajas se dispusieron en dos niveles de zona intermareal de la estación 1, cuatro en el nivel superior, aproximadamente a 70 m de la línea de costa durante la marea más alta y cuatro más en la parte inferior del intermareal a 30 m de las primeras. Los organismos de dos cajas de cada nivel se enumeraron con tinta imborrable y se midieron cada dos meses. Los individuos de las cajas restantes se enumeraron con una excavación ligera en el periostraco y se midieron sólo a mediados y al final del experimento. Al final del experimento se aplicó la prueba de U Mann Witthney (Siegel, 1980) para comparar los incrementos en longitud entre los mejillones de cada par de intervalos de tamaño de las cajas dispuestas en el mismo nivel y con iguales períodos de medición. También se utilizaron análisis de varianza no paramétricos de tres vías (Wilson, 1956) para detectar diferencias en el crecimiento de los organismos entre los intervalos de tamaños, entre los distintos niveles y entre los diferentes periodos de medición. El crecimiento relativo de Modiolus capax se estimó utilizando los datos de la longitud anteroposterior, ancho, alto y peso húmedo total de los organismos colectados. Estos parámetros se relacionaron de acuerdo a una regresión lineal simple (ecuación alómetrica) Y = aXb o en su forma logarítmica: Log Y = Log a + bLog X. Las constantes a y b se estimaron por regresiones de mínimos cuadrados (Zar 1974). Para comparar las pendientes de regresión con las pendientes teóricas de isometría y para comparar los pares de las pendientes de regresión entre las estaciones 1 y 2, se aplicaron pruebas t de Student (Zar, 1974). RESULTADOS
ESTRUCTURA DE TALLASLa población deModiolus capax estuvo formada principalmente por organismos de tallas grandes. En la estación 1 (Fig. 2) los individuos mayores de 50 mm constituyeron durante la mayor parte del tiempo más del 75 % del conjunto, donde los organismos de tallas entre 80 y 90 mm fueron los más abundantes. Los primeros reclutas (organismos menores de 15 mm) se observaron en el mes de mayo y aunque a partir de agosto su presencia fue más notoria, nunca excedieron el 18% de la población. El animal más grande recolectado en esta estación fue de 126 mm. En las muestras de la estación 2 (Fig. 3), a excepción de enero, más del 75 % de los individuos fueron mayores de 75 mm, dominando numéricamente los organismos con longitudes anteroposteriores entre 80 y 100 mm. Los organismos menores de 15 mm solo fueron relativamente abundantes en noviembre de 1985 y en enero de 1986 (10 y 19.5 % respectivamente). El organismo más grande recolectado en esta estación fue de 130 mm y en el banco se hallaron individuos muertos de 200 mm. CRECIMIENTODebido al alto grado de traslape y el poco desplazamiento de la mayoría de las clases de tamaño determinadas por el método de Cassie (1954), el estudio del crecimientoMódiolus capax por este método sólo se pudo hacer con los organismos recién asentados en la estación 1 (Fig. 2) en el mes de mayo de 1985, siguiendo el desplazamiento de su moda hasta febrero de 1986. Las tallas promedio alcanzadas en los diferentes meses y sus desviaciones estándar se muestran en la Tabla 1. La tasa de incremento mensual promedio de la longitud anteroposterior fue de 1.83 mm de mayo a agosto de 1985, disminuyó a 1.0 mm entre agosto y diciembre y a 0.75 mm/mes entre diciembre de 1985 y febrero de 1986. El distanciamiento total de la moda de estos organismos, a partir de una longitud promedio de 5 mm, fue de 11 mm en nueve meses (1.22 mm/mes). El crecimiento de los organismos marcados fue significativamente similar entre los mejillones del mismo intervalo de longitud pertenecientes a cada par de cajas bajo las mismas condiciones ambientales. Las pruebas U' de Mann Wittney (Siegel, 1980), indicaron que no hubo diferencias significativas en los incrementos de la longitud anteroposterior entre los mejillones de cada par de intervalos de tamaños pertenecientes a cada par de cajas que se midieron en los mismos intervalos de tiempo en cada nivel de la estación 1. Por esta razón las observaciones fueron reagrupadas. El análisis de varianza no paramétrico de tres vías (Wilson, 1956), demostró que hubo diferencias significativas entre los incrementos de la longitud de los mejillones de cada intervalo de tamaño reagrupados y entre los mejillones medidos en diferentes intervalos de tiempo (Tabla 2.). Las Tasas. de crecimiento de los organismos medidos aproximadamente cada dos meses (Tabla 3a) fueron menores que las de los mejillones medidos sólo dos veces durante el experimento (Tabla 3 b) donde los incrementos mayores sucedieron en los mejillones 3 con tallas entre 15 y, 43 mm (1.29 mm/mes). En general, en la medida que aumentó la talla disminuyó la velocidad de crecimiento. Los mejillones de 15 mm murieron probablemente a causa de la ligera incisión que se les hizo en el periostraco. En la Tabla 4 se iudican los valores de los coeficientes de las regresiones de las variables morfométricas deM. capax en las dos estaciones de muestreo. Todas las regresiones fueron significativas (p<0.05), y tuvieron diferencias significativas entre las pendientes teóricas de isometría y las observadas. Los resultados indicaron que en ambas estaciones la longitud anteropoterior aumenta relativamente más rápido que el alto y el ancho (alometría positiva) y que el ancho crece relativamente un poco más rápido que el alto (alometría negativa). El peso húmedo total tuvo una relación alométrica negativa con respecto a la longitud anteroposterior, al ancho y al alto. (Fig. 4). La comparación de los pares correspondientes de las pendientes de regresión entre las dos estaciones de muestreo denotó diferencias significativas entre ellas. 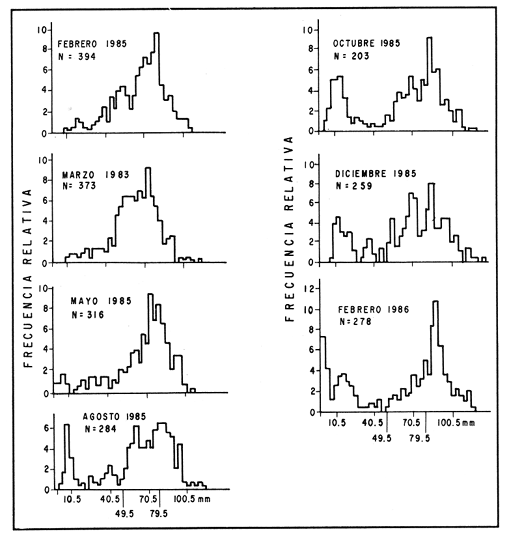 Figura 2. Histogramas de la longitud anteroposterior de los mejillones Modiolus capax en la estación de muestreo 1. 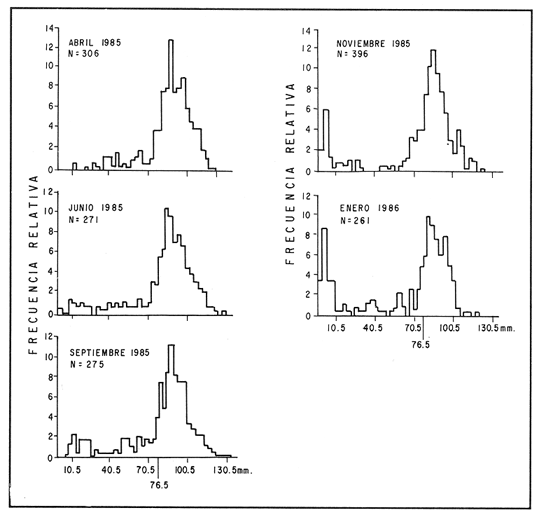 Figura 3. Histogramas de la longitud anteroposterior de los mejillones Modiolus capax de la estaciÓn 2 en los diferentes meses de muestreo. 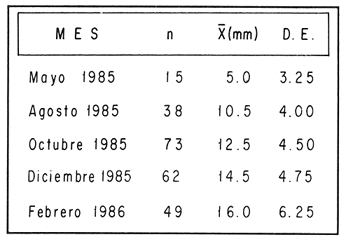 TABLA 1 VALOR PROMEDIO DE LA LONGITUD ANTEROPOSTERIOR ALCANZADA POR LOS MEJILLONES Modiolus capax RECIEN ASENTADOS EN EL BANCO NATURAL DE LA ESTACIÓN 1. n: NÚMERO DE INDIVIDUOS; D.E.: DESVIACIÓN ESTÁNDAR 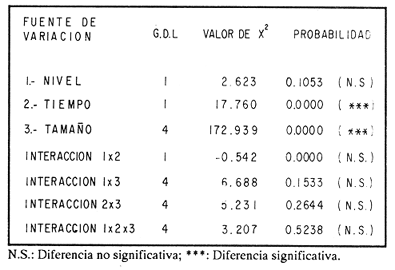 TABLA 2 ANÁLISIS DE VARIANZA NO PARAMÉTRICO DE TRES VÍAS (Wilson; 1956) DEL CRECIMIENTO DE LOS MEJILLONES Modiolus capax MANTENIDOS EN LAS CAJAS EXPERIMENTALES EN LA ESTACIÓN 1 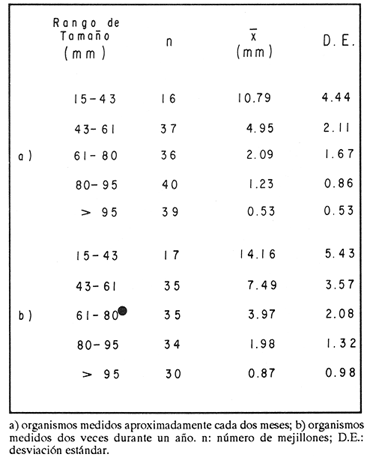 TABLA 3 INCREMENTO MEDIO ANUAL POR LA TALLA DE LA LONGITUD ANTEROPOSTERIOR DE LOS MEJILLONES Modiolus capax DE LAS CAJAS EXPERIMENTALES 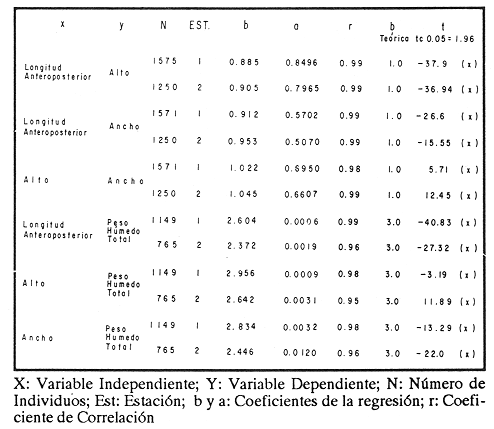 TABLA 4 RELACIONES ALOMÉTRICAS ENTRE LAS VARIABLES MORFOMÉTRICAS DE LOS MEJILLONES Modiolus capax DE LAS DOS ESTACIONES DE MUESTREO Y PRUEBA t DE STUDENT (Zar, 1974) SOBRE LA SIGNIFICANCIA DE LA PENDIENTE TEÓRICA 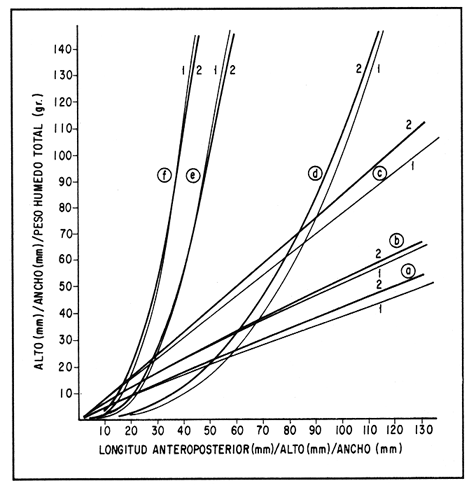 Figura 4. Relación entre las diferentes variables morfométricas de Modiolus capax en cada una de las estaciones de muestreo. Estación 1, 1; Estación 2,2; a) relación longitud anteroposterior-ancho; b) relació longitud anteroposterior-alto; c)relación ancho-alto; d) relación longitud anteroposterior-peso húmedo total; e) relación alto-peso húmedo total; f) relación ancho-peso húmedo total. DISCUSIÓNDel análisis de la estructura de tallas deMódiolus capax se determinó que la población está formada principalmente por organismos grandes y que los individuos más pequeños forman un grupo reducido y variable en el tiempo. En general, aunque el asentamiento de los reclutas, que se observó durante gran parte del año, fue más numeroso a partir de agosto, estos mejillones nunca excedieron el 20 % del total de la población. Este valor es muy bajo en comparación con otras especies de mejillones comomytilus californianus del ejido de Eréndira, B. C. Méx. (García et al., 1982), donde el 55 al 85% de la población corresponde a organismos menores de 6 mm. Sin embargo, la estructura de tallas y el patrón de asentamiento de M. capax es muy similar a los que caracterizan las poblaciones deModiolus modiolus en Irlanda del Norte (Seed y Brown, 1977) y en la costa de Escocia (Comely, 1978). La variabilidad en el reclutamiento entre las dos poblaciones se observa en la estación 2 donde al parecer el reclutamiento es muy bajo y se refleja en que los organismos menores de 70 mm son muy pocos. En las costas de Escocia, Comely (1978) encontró también diferencias similares en el reclutamiento entre poblaciones vecinas y sugirió que las condiciones hidrográficas pueden, en gran parte, ser las causantes de ello. Este estudio carece de datos suficientes para esclarecer esas diferencias. Sin embargo, considerando la desigualdad de reclutas que se asentaron entre febrero de 1985 y febrero de 1986 en la estación 1 (Fig. 2), parece indicar que no sólo existen variaciones en el asentamiento larval de un año a otro, sino que probablemente también en el ciclo reproductivo de la especie entre diferentes años, ya que Garza y Bückle (1989) demostraron que el ciclo reproductivo de M. capax se caracterizó por una liberación continua de gametos de intensidad variable desde febrero de 1985 a enero de 1986 y Espinoza-Peralta (1989) reporta que de octubre a diciembre de 1986 los organismos de las mismas poblaciones naturales se encontraron en etapas posteriores a la liberación de gametos o en reposo. En este sentido es posible considerar que el notable aumento de las fijaciones de reclutas detectado en agosto de 1985, pudo corresponder a una liberación de gametos más intensa acaecida antes de agosto del mismo año' según los resultados del estudio del ciclo reproductivo realizado paralelamente por Garza y Bückle, 1989, investigación que también puede explicar el efecto acumulativo de los nuevos asentamientos durante los siguientes meses al relacionarlo con las liberaciones continuas de gametos, pero de menor intensidad, detectadas desde septiembre de 1985 hasta enero de 1986. Junto a estas observaciones se hizo otra investigación para conocer la distribución temporal y espacial de las larvas de M. capax en la bahía de Los Angeles con la cual se determinó que las formas plantónicas de esta especie son dominantes, pero cuantitativamente variables en la composición del zooplancton. Comparativamente las larvas Veliger son mucho más numerosas que las Pediveliger prontas a metamorfosearse (Serrano-Guzmán, en preparación), Al parecer en la bahía de Los Angeles existe una correspondencia entre la cantidad de larvas Pediveliger que hay en el Zooplancton (sin considerar que en esta etapa normalmente existe una mortalidad natural adicional antes de la metamorfosis) y el exiguo reclutamiento que se constata en las poblaciones naturales donde a su vez pudieran existir limitaciones en el sustrato de asentamiento ocupado por poblaciones adultas como es el caso en esta investigación. Por otra parte los resultados de una investigación realizada en 1989 por Fárfán (com. personal) (CICESE) para hacer un estudio comparativo de otras poblaciones de este organismo, que se extendió por toda la zona del Pacífico a lo largo de Baja California y Baja California Sur hasta San Felipe, por el Golfo de California, indican que el reclutamiento en esta especie es muy limitado. En los mejillones marcados se observó el efecto sobre el crecimiento de las mediciones repetidas frecuentemente causando que los organismos crecieran significativamente menos. Probablemente influyó en ello la tensión a que fueron sometidos y al gasto de energía que significó la formación de biso para volver a fijarse. El crecimiento promedio mensual de los organismos marcados de longitudes entre 15 y 43 mm (1.29 mm/mes) fue muy similar al incremento promedio de los organismos recién asentados en la población natural (1.22 mm/mes). Esto sugiere que el crecimiento de los reclutas pudo haber sido subestimado debido a la falta de información sobre el asentamiento durante los meses de junio y julio (meses sin colecta), teniendo en cuenta que la tasa de crecimiento es más alta en los organismos de tallas menores como se constató en las cajas experimentales. Los resultados que encontró Aguirre-Hinojosa (1987) con los mejillones de la misma especie que se fijaron en sartas hechas de red anchovetera suspendidas de una estructura flotante en la bahía de Los Angeles, indican que el crecimiento de esos mejillones con tallas promedio iniciales de 0.7 y 0.9 mm fue de 17.6 mm (2.9 mm/mes) a 20.8 mm (3.4 mm/mes) en 6 meses. Estos valores son bajos con respecto a otras especies de mejillones como Perna perna, que en cinco meses creció aproximadamente 21.6 mm (4.32 mm/mes) (Cayré, 1978);Mytilus californianus cuya velocidad de crecimiento en el primer año fue de 4,6 mm/mes hasta hacerse casi constante (0.2 mm/mes) después del tercer año (García et al., 1982). Sin embargo, en especies del mismo género también se han registrado bajas tasas de crecimiento como por ejemploModiolus modiolus de Irlanda del Norte (Brown y Seed, 1977; Seed y Brown, 1977, 1978) y Modiolus demissuss granossissimus en Louisiana, E.E.UU. (Rodríguez-Ortega y Day, 1978). El patrón del crecimiento relativo deM. capax determina que los organismos mayores sean relativamente más anchos y adquieran formas más cilíndricas. Este cambio gradual en las proporciones corporales con el aumento en la longitud anteroposterior explica las diferencias en la morfología de los individuos en la estación 2, donde la mayoría tuvieron formas un poco más cilíndricas. Las diferentes estructuras de tamaños de M. capax en las dos poblaciones estudiadas se reflejan en los distintos valores de las pendientes de regresión entre las variables morfométricas. Con esta investigación es posible reconocer que Modiolus capaxes un organismo que vive en el intermareal, "tolerante al estrés" en el sentido de Vermeij (1946), que se caracteriza por crecer lentamente y porque el reclutamiento es muy bajo. En consecuencia una población puede ser severamente afectada si no se delinea una forma racional de explotación. AgradecimientosEste trabajo fue financiado parcialmente por el Consejo Nacional de Ciencia y Tecnología (Proyecto, Clave: PCECCNA-050018). LITERATURAAGUIRRE-HINOJOSA, E.Tesis de Maestría en Ciencias. Análisis de la fijación, crecimiento y sobrevivencia del mejillón Modiolus capax (Conrad) (BIVALVIAMYTILIDAE), durante el desarrollo de una comunidad sobre sustratos artificiales en la bahía de los Angeles, B.C.Centro de Investigación Científica y de Educación Superior de Ensenada.Ensenada, Baja CaliforniaMéxico1987106 P. BAQUEIRO, C.E.Aquaculture Status of molluscan aquaculture on the Pacific coast of México.198483-9339 BARNARD, J.L.; y J.R. GradyA biological survey of Bahía de Los Angeles. Gulf of California. México. I: General account.San Diego Soc. Nat. Hist. Trans.196851-6615(6) BROWN, R.A., y R. SEEDModiolus Modiolus (L.). An autoecological study.Proc. 11th Europ. Symp. mar. biol. Pergamon Press. 197793-100 CASSIE, R.M. Some uses of probability paper in the analysis of size frecuency distribution.Austjour. of Mar. and Fresh. Res1954513-5225 CAYRE, P. Etude de la moule Perna perna L. et des possibilites de myticulture en Republique Populaire au Congo.Can. ORSTOM, Sér. Océanogros.19789-1716(1) COMELY, C.A. Biology. Ophelia Modiolus Modiolus (L.) from the Scottish west coast. I.1978167-19317(2) ESPINOZA PERALTA, A.M.Tesis de Licenciatura. Dilación del desove de Modiolus capax Conrad (Bivalvia:Mytilidae) en condiciones controladas de temperatura y alimentación. Escuela de Biología. Universidad Autónoma de Baja California. Ensenada, Baja CaliforniaMéxico1989106 p. GARCÍA, P.F., A. SALAS G. y A. OLIVA DE LA P.Ecología de Mytilus californianus. En: estudios para el desarrollo del cultivo comercial de los mejillones Mytilus californianus y Mytilus edulis en las costas de Baja California. Informe anual 1982.UABCIIO-SEP. Universidad Autónoma de Baja California Ensenada, B.CMéxico1982 Tomo II GARZA-AGUIRRE, M.C., y L. F. BÜCKLE R. An. Inst. Cien. del Mar y Limnol.Ciclo reproductivo del mejillón Modiolus capax (Conrad, 1837) (BIVALVIA, MYTILIDAE, ANISOMYARIA) en la bahía de los Angeles, Baja California, México. Univ. Nal. Autón. México198916(1) KAESTNER, A. Lehrbuch der Speziellen Zoologie. Teil I: Wirbellose, 1 Halbband.VEB Gustav Fischer Verlag. Jena1955298-299 OCHOA-BAEZ, R.L.Antecedentes sobre el ciclo de reproducción de Modiolus capax (Conrad, 1837) (BIVALVIA:MYTILIDAE) en la bahía de La Paz Baja California Sur, México. Inv. Mar. CICIMAR198586-1032(2) ORDUÑA-ROJAS, J.Tesis de Licenciatura.Desove y desarrollo embriológico y larval del mejillón Modiolus capax CONRAD (BIVALVIA: MYTILIDAE) en condiciones controladas.Escuela Superior de Ciencias Marinas. Universidad Autónoma de Baja California.Ensenada, Baja CaliforniaMéxico198652 p. RICO-MORA, R. Tesis de Maestría en Ciencias. Efecto interactivo de la temperatura y de la concentración de microalgas en la fisiología alimenticia y la energía potencial para el crecimiento de Modiolus capax Conrad. Centro de Investigación Científica y de Educación Superior de Ensenada. Ensenada, Baja CaliforniaMéxico1987 91 P. RODRÍGUEZ-ORTEGA. D. y J.W. DAY. An. Centro Cienc. Mar y Limnol. An ecological study of Modiolus demissus granossissimus Sowerby in Louisiana (USA).Univ. Nal. Auton. México1978215-2245(1) SEED, R. AND R.A. BROWN.A comparation of the reproductive cycles of Modiolus modiolus (L.), Cerastoderma (Cardium) edule (L.), and Mytilus edulis L. in Strong Ford Lough, Northern Ireland.Oceologia1977173-17830 SEED, R. AND R.A. BROWNGrowth as strategy for survival in two marine bivalves, Cerastoderma edule and Modiolus ModiolusJour. Mar. Ecol.1978283-29247 SERRANO-GUZMÁN, SJ.Tesis de Maestría en Ciencias. En preparación. Distribución espacial y temporal de las larvas de Bivalvos durante la Primavera y el Verano de 1986 en la bahía de Los Angeles, B.C., México. Centro de Investigación, Científica y de Educación Superior de Ensenada.Ensenada, Baja CaliforniaMéxico SIEGEL, S. Non parametric statistics for the behavioral sciences. Mc.Graw-Hill, Kogakusha, LTD.International student edition.1980 346 p. VERMEIJ J. GEERATBiogeography and Adaptation. Patterns of maríne lífe. Harvard University Press. Cambridge, Massachusetts.1946322p. WILSON, R.V.Psychological Bulletin A distribution-free test of analysis of varianze hypotesis.195690-10153(1) ZAR, J. H.Biostatistical analysis. Prentice Hall, biological sciencies series.1974620 p.
|