|
LOS CROMOSOMAS DE LOS PECES Oreochromis
urolepis hornorum y Oreochromis mossambicus (PISCES: CICHLIDAE)
Trabajo
recibido el 2 de marzo de 1988 y aceptado para su publicación el 14 de
marzo de 1988.
Julio Arreguín-Espinosa
Manuel Uribe-Alcocer
Instituto de Ciencias del Mar y Limnología, Universidad
Nacional Autónoma de México. Contribución No. 657 del
Instituto de Ciencias del Mar y Limnología, UNAM.
Con el fin de incrementar la producción alimentaria, varias
especies ícticas han sido introducidas a México. Entre ellas, las
tilapias han sido utilizadas en pruebas para obtener progenie monosexual
masculina por medio de la cruza entre diferentes especies o líneas. En
este trabajo se informa de los estudios citogéneticos llevados a cabo a
fin de establecer un cariotipo patrón, auxiliado por técnicas de
bandas tipo G para las especiesOreochromis urolepis hornorum y O.
mossambicus,así como en su híbrido, provenientes de las
estaciones piscícolas "El Rodeo" y "Zacatepec" del Estado de Morelos. En
los estudios realizados no se detectaron marcadores cromosómicos
útiles en la acuicultura, pero se enfatiza el potencial de las tilapias
para producir líneas híbridas mejoradas de valor alimentario
incrementado.
Some exotic species have been
introduced into Mexico in order to increase food production. The african
tilapias are particularly relevant in this respect, as they have been
repeatedly used in trials to obtain monosexual progeny by crossing different
species or strains. This paper reports the cytogenetic studies carried out to
establish a conventional and G band karyotype of the species
oreochromis urolepis hornorum and O. mossambicus, as well as
their hybrid, used in the fish factories of "El Rodeo" and "Zacatepec" of the
Mexican state of Morelos. Our results could not bring out chromosomal markers
useful in acuicultural practices, although they pointed out the potential of
the tilapia stocks to produce hybrid selected strains of high alimentary
value.
El abastecimiento de proteína de origen animal para la población nacional constituye un problema prioritario en nuestro país. A fin de resolverlo se han introducido a México diecinueve especies de peces, aproximadamente la mitad de las cuales se explotan en pesquerías en las aguas interiores del país, entre las que destacan diversas especies de carpas, lobinas, truchas, bagres y mojarras (Rosas Moreno, 1976). Entre las mojarras, destaca la importancia de la familia Cichlidae, de la cual se introdujeron a México en 1964, tres especies de tilapias procedentes de Auburn, Alabama, EUA, y que son Oreochromis mossambicus, O. nilotica y T. melanopleura (T. rendalli), originarias de Africa. Hacia 1981, fueron introducidas la variedad albina de O. mossambicus y O. urolepis hornorum procedentes de Palmetto, Florida, EUA. Rosas Moreno en 1976 informó que en ese año se obtuvo una captura de 7 a 8000 toneladas en la presa de Temazcal, Oax. Este volumen ha ido incrementándose gradualmente, y en el primer semestre de 1986 se ha informado de un volumen de producción nacional de tilapias, procedentes de prácticas acuiculturales, de 32000 toneladas (Sepesca, 1986). Una de las limitantes del cultivo de la tilapia consiste en que, bajo condiciones de sobrepoblación, se da una disminución de tamaño y reproducción precoz, situaciones a evitar por la baja de producción que provocan. A fin de controlar la sobre población se han establecido diferentes metodologías, como la introducción de depredadores de crías o huevos, y la separación de los machos. Una de las metodologías utilizadas más ampliamente para lograr un mayor rendimiento alimentario de estos peces, ha sido la producción de progenie monosexual masculina por medio de cruzamientos interespecíficos (Jalabert et al., 1971). Si en el transcurso de un programa de cría para producción de progenie híbrida masculina se incorporaran accidentalmente individuos híbridos a las cepas progenitoras, es muy probable que se abatieran los rendimientos de producción de progenie masculina, en contra de los objetivos del programa de cría. En esta situación es imperativo contar con características diagnósticas que permitan reconocer la especie a la que pertenezcan los individuos de los lotes de reproductores. Una de las características puede ser el cariotipo, ya que el número de cromosomas, así como su tamaño y estructura son caracteres particularmente estables dentro de las especies, y normalmente los cariotipos de especies diferentes, son asimismo distintos. En la literatura se informa de estudios cromosómicos realizados en Oreochromis mossambicus en los que se ha encontrado que tienen un número diploide de 44 cromosomas (Tabla 1). Hasta donde es de nuestro conocimiento no se han realizado estudios cromosómicos en O. urolepis hornorum, y dado que en algunas especies de tilapias se ha encontrado un número cromosómico distinto de 44, tal como 40 en O. niloticus (T. nilotica) y 38 enT. zilii (Badr y El-Dib, 1977), etc., se investigó el número cromosómico de O. u. hornorum, y se verificó el número cromosómico de O. mossambicus presente en el país, a fin de estudiar su posible uso como marcador genético de las líneas en cultivo en las Piscifactorías de el Rodeo y Zacatepec, Mor. Es de suponerse que de haber hibridación entre especies con número cromosómico diferente, el híbrido mostrará un número de cromosomas intermedio, así como asociaciones cromosómicas anormales, en la meiosis. Híbridos resultantes de una cruza de Oreochromis mossambicus (2n=44) x T. nilotica (2n=40) tendrían 42 cromosomas. Una cruza de Oreochromis mossambicus (2n=44) x T. melanopleura = T. zilii(2n=38) originarían individuos híbridos de 41 cromosomas. Resulta conveniente investigar si las poblaciones presentes en el país pueden ser diferenciadas en base a su cariotipo, puesto que si las poblaciones son portadoras de complementos cromosómicos distintos, se podrían llevar a cabo cruzas interespecíficas con descendencia estéril en gran proporción o totalmente, lo que resultaría de relevancia en el cultivo de estos peces. En el país se han iniciado los estudios citogenéticos de poblaciones híbridas de tilapias de tres localidades, en las que se ha asumido existen diferencias ecológicas notorias: Piscifactoría de los Amates, Ver., Piscifactoría del Rodeo, Mor. y Yuriria, Gto. (Castorena-Sánchez y colaboradores, 1983). Los objetivos de este estudio fueron: a) Llevar a cabo el estudio convencional de los cariotipos, a fin de encontrar características diagnósticas diferenciales. b) Establecer el cariotipo patrón, por medio de la utilización de parámetros estadísticos, a fin de establecer elementos diferenciales entre las poblaciones en estudio y realizar la comparación estadística de los pares cromosómicos individuales de ambas poblaciones. c) Establecer los cariotipos patrón, auxiliados por técnicas de bandas cromosómicas tipo "G", de las especies en estudio, como elementos de administración de este recurso. MATERIAL Y MÉTODOSa) Para los estudios citogenéticos convencionales, se estudiaron un total de 18 ejemplares (10 machos y 8 hembras) de Oreochromis urolepis hornorum, y 6 de O. mossambicus (3 machos y 3 hembras), y 7 individuos híbridos F1O. mossambicus x O. u. hornorum (4 machos y 3 hembras). Se trabajó con tejido branquial y con las gónadas de los híbridos provenientes de la estación Piscícola El Rodeo, Mor. de acuerdo a la técnica de McPhail y Jones (1966) con adaptaciones realizadas en el Laboratorio de Genética de Organismos Acuáticos (Maldonado-Monroy,. 1984 y Uribe-Alcocer, 1985). b) Se seleccionaron 12 campos mitóticos deO. mossambicus, 6 de O. u. hornorum y 17 de híbridos, provenientes de diferentes organismos, de la calidad requerida para llevar a cabo la medición de sus elementos, y para la elaboración de los cariotipos con bandas G. En los organismos híbridos machos se seleccionaron adicionalmente 16 figuras meióticas.  TABLA 1 NÚMEROS CROMOSÓMICOS DE DIFERENTES ESPECIES DE LA TRIBU Tilapiine c) El análisis estadístico realizado fue la comparación de medias de las longitudes de cada brazo cromosómico obtenidas de las mediciones provenientes de cada una de las dos poblaciones estudiadas. d) La obtención de las bandas "G" cromosómicas se hizo mediante un tratamiento con una solución 5M de urea y cloruro de sodio, a un pH de 7.8, por pulsos que variaron de 2 a 10 segundos, a una temperatura de 37 °C durante todo el proceso (Lara, 1985). Posteriormente se enjuagaron las laminillas en agua destilada, con el objeto de detener el proceso al que fue sometido el DNA y a su vez retirar los residuos de urea para evitar la formación de cristales que impidan la observación microscópica. La tinción de las laminillas se llevó a cabo con colorante de Giemsa, elaborado a partir de una solución stock que se diluye en Buffer Sörensen de fosfatos (KHPO Y NaHPO) en una proporción de 1:50 Giemsa-Sörense respectivamente, y a un pH de 6.8. La tinción se llevó a cabo durante 30 min. Las laminillas se enjuagaron con agua destilada y se secaron al aire. e) A fin de elaborar los idiogramas con los patrones de bandas G de los distintos pares cromosómicos, se llevaron a cabo los siguientes pasos: 1) se preparó un idiograma basado en las mediciones de longitud relativa de los brazos cortos, brazos largos y longitud total de cada par cromosómico. 2) se identificaron las bandas más constantes y notorias de cada par cromosómico y se subdividió el cromosoma tomando como límite de cada región alguna de las bandas presentes. 3) se integraron estas subdivisiones en el idiograma anteriormente establecido, obteniéndose finalmente una representación gráfica de las bandas encontradas. RESULTADOS Y DISCUSIÓNEn la figura 1 se muestra un cariotipo y un campo mitótico representativo de Oreochromis mossambicus, con un número diploide de 44 cromosomas. Los cinco pares de homólogos del primer renglón son birrámeos y los diecisiete pares restantes son monorrámeos. Esta estructura cariotípica fue encontrada en la totalidad de la muestra estudiada de esta especie. En la figura 2 se muestran el cariotipo y un campo mitótico representativo de O. u. hornorum, mostrando igualmente un número diploide de 44 elementos cromosómicos. Al igual que en la muestra de O. mossambicus, los cinco pares de cromosomas homólogos que han sido colocados en el primer renglón son birrámeos. Los diecisiete pares restantes son monorrámeos. Esta estructura cariotípica es común de las muestras estudiadas.  Figura 1. Cariotipo y campo mitótico representativo de Oreochromis mossambicus, mostrando un 2N=44.  Figura 2. Cariotipo y campo mitótico representativo de Oreochromis urolepis hornorum, mostrando igualmente un 2N = 44. En la figura 3 se muestra un esquema de una metafase de epitelio branquial de un ejemplar híbrido O. mossambicus xO. u. hornorum, procedente de la Piscifactoría del Rodeo, Mor., mostrando un número diploide de 44 cromosomas (10 birrámeos y 34 monorrámeos). Puede verse también una profase meiótica I con 22 bivalentes y el cariotipo de la metafase ilustrada. No es posible distinguir los elementos provenientes de las líneas parentales debido a la gran semejanza, o a la identidad cromosómica de las líneas parentales. El estudio de campos meióticos llevado a cabo para detectar rearreglos internos del cromosoma, difícilmente distinguibles mediante la utilización del cariotipo teñido convencionalmente, no mostró la presencia de dichos reacomodos cromosómicos, ya que se observaron apareamientos normales, y en la metafase I se observaron claramente en todos los casos veintidós bivalentes. El número fundamental (número total de brazos cromosómicos) resultó ser igual en ambas especies, con un valor de 54. La fórmula cromosómica de las especies fue asimismo igual, 4sm + 1st + 17t, correspondiendo a la presencia de 5 pares de cromosomas birrámeos y 17 pares unirrámeos. El número diploide de los especímenes estudiados corresponde a aquéllos de que se informa en todos los estudios en el género Oreochromis a que se hace mención en la Tabla 1, excepto los que presentan Badr y El-Dib (1977) sobre O. niloticus y Post (1965) sobre O. alcalicus (Tabla 1). Los valores obtenidos de las mediciones de los 12 cariotipos deOreochromis urolepis hornorum se observan en la Tabla 2, donde se presentan los pares de cromosomas en orden de longitud decreciente y se muestran las clasificaciones obtenidas según los métodos de Levan et al., 1964, y Al-Aish, 1969. Los valores obtenidos de la medición de los cariotipos de O. mossambicus, se muestran en la Tabla 3, así como su respectiva clasificación. En la figura 4 se grafican comparativamente las longitudes relativas de los elementos cromosómicos de los cariotipos estudiados, y se enfatiza el alto grado de semejanza entre ellos. El análisis estadístico de los cromosomas que, con excepción de los brazos del par cromosómico 3, y de los cromosomas 6, 7 y 19, donde se encontraron valores significamente diferentes (P = 0.95), indica que se puede considerar que el resto de los brazos cromosómicos son iguales, y que las diferencias encontradas pueden ser atribuidas a factores fortuitos.  Figura 3. Esquema elaborado con una cámara lúcida de una metafase de epitelio branquial de un ejemplar híbrido de Oreochromis mossambicus x O. u. hornorum, procedente de la Piscifactoría del Rodeo, Mor., mostrando un 2N = 44 (10 birrámeos y 34 monorrámeos). En la figura 5 se muestra un idiograma con las bandas G encontradas en los cromosomas deOreochromis mossambicus, y en la 6 con las de O. u. mossambicus. En los pares cromosómicos mayores (pares 1 a 5), se encontraron las siguientes diferencias en el patrón de bandas "G": a) Par cromosómico 4: En O. u. hornorum no se aprecian bandas en la porción correspondiente al brazo "p", mientras que en O. mossambicus se observa una banda en la región telemétrica del mismo brazo. c) Par cromosómico 5: Se observa una banda en el brazo "p" de O. u. hornorum y una banda telemétrica en su brazo "q", mientras que en O. mossambicus la banda del brazo "p" abarca la totalidad del brazo y en el brazo "q" se identificaron dos bandas. Las diferencias encontradas en los cromosomas monorrámeos no son fáciles de analizar, ya que la longitud de estos cromosomas es muy pequeña en comparación con los cromosomas birrámeos y la constricción producida por la colchicina no permite evaluar con consistencia el patrón de bandas. La estructura de los cromosomas de las especies del géneroOreochromisestudiadas aparentemente es idéntica ya que, aunque el arreglo del cariotipo difiere del presentado por Kornfieldet. al. (1979) o por Majumdar y McAndrew (1986), existen los mismos elementos cromosómicos y la posición del centrómero es muy parecida. La longitud de los brazos cortos del segundo par birrámeo subtelocéntrico del cariotipo del primer trabajo mencionado es muy semejante a la del par mayor de los cromosomas telocéntricos del cariotipo que se presenta en este trabajo. El índice centromérico encontrado en este par cromosómico, corresponde a un elemento telocéntrico. Sin embargo, brazos cromosómicos de tal longitud son susceptibles de modificaciones mayores debidas básicamente al tiempo de exposición al mitostático, que brazos de mayor tamaño. Estas modificaciones se ven reflejados en las mediciones e interpretación correspondientes. Nuestra Interpretación concuerda con la hecha por Prassad y Manna (1976) en O. mossambicus. Por lo anterior, se puede considerar que los cariotipos convencionales de los organismos estudiados, no presentaron diferencias entre los individuos de una población ni entre ellas, por lo que se pueden considerar iguales, de no ser por cambios en las longitudes de los cromosomas atribuibles a la concentración o tiempo de administración del mitostático o a la dispersión de los cromosomas. Se juzga conveniente que los brazos cromosómicos sean estudiados en una muestra más amplia en estudios posteriores, ya que, pudiendo llegar a tener importancia evolutiva no es claro qué papel biológico puedan llegar a tener en la diferenciación de ambas poblaciones. Por otra parte no parece factible la posibilidad de utilizarlos como marcadores genéticos, ya que la diferencia que se ha encontrado es poblacional, y carecería de valor diagnóstico a nivel individual, siendo muy importante el poder determinar la pertenencia a un grupo particular de cada uno de los individuos que se quieran evaluar como reproductores. Es posible que estas poblaciones en estado de simpatría hayan fortalecido su aislamiento reproductivo, de tal modo que no exista hibridación entre ellas, pero que, al momento de entrar en contacto con otras poblaciones filogenéticamente próximas, carentes de este mecanismo de aislamiento fortalecido, puedan establecer con ellas flujo genético, de una manera semejante a lo que se presenta en las poblaciones del roedorPeromyscus melanotis (Bowerset al., 1973). Probablemente la especiación de las tilapias siga patrones de aislamiento reproductivo distintos de la diversificación cariotípica, y se pudiera caracterizar por la especialización conductual en el momento del apareamiento o cuidado de la cría, o bien por la ocupación de distintos nichos ecológicos. 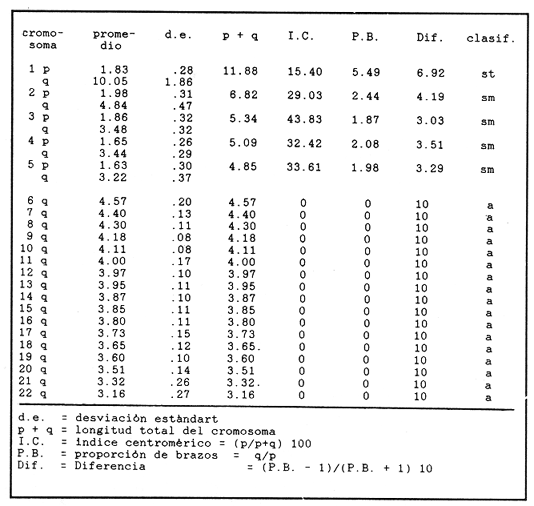 TABLA 2 MEDICIONES DE CAREOTIPO DE Oreochromis urolepis hornorum Y CLASIFICACIÓN 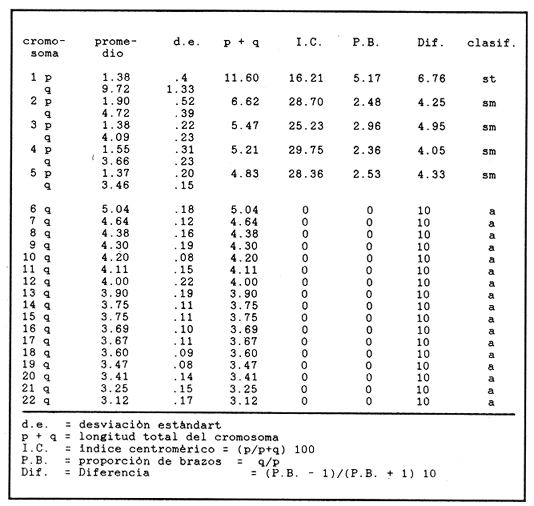 TABLA 3 MEDICIONES DE CAREOTIPO DE Oreochromis mossambicus Y CLASIFICACIÓN 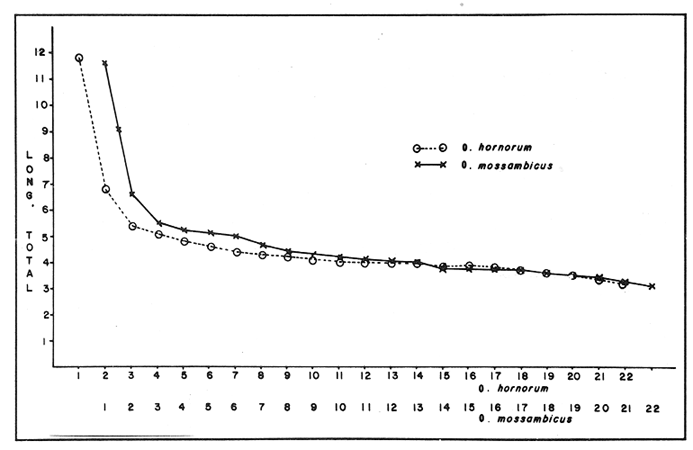 Figura 4. Comparación de la longitud relativa de O. mossambicus y O. u. hornorum. 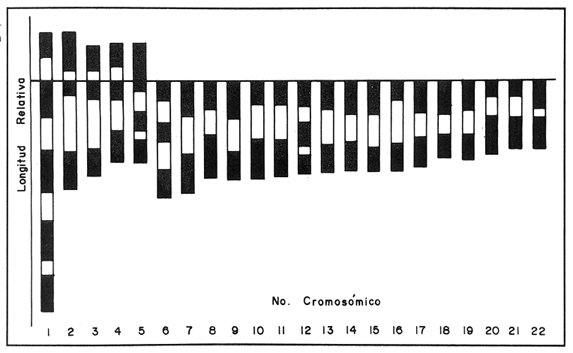 Figura 5. Idiograma de O. mossambicus mostrando el patrón de bandas G. 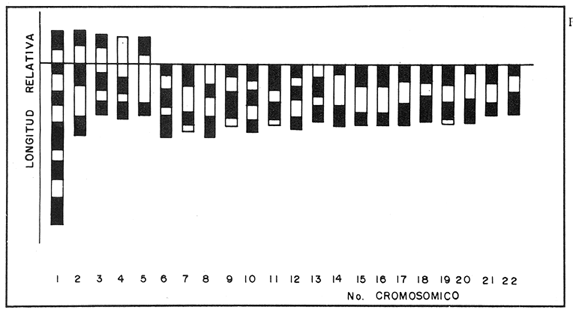 Figura 6. Idiograma de O. u. hornorum. mostrando el patrón de bandas G detectadas en su complemento cromosómico Conclusiones1. En el estudio de las tres muestras, se observó indistintamente un número diploide de 44 cromosomas, de los cuales, cuatro pares son cromosomas submetacéntricos, un par es subtelocéntrico y diecisiete pares tienen centrómero terminal. 2. El análisis cariotípico convencional no resulta útil en la identificación a nivel específico de las poblaciones de tilapias utilizadas en la Piscifactoría de Zacatepec, Mor., para la producción de progenie híbrida. 3. El flujo genético que se presenta en algunas tilapias, ofrece promisorias perspectivas de investigación aplicada, al permitir buscar mediante hibridaciones, cepas mejoradas de alto rendimiento. AgradecimientosEste trabajo fue realizado con fondos del presupuesto interno otorgado por la UNAM al Instituto de Ciencias del Mar y Limnología, y con fondos canalizados por el CONACYT al Proyecto clave PCCBBNA - 021431 "Estudios genéticos en peces de importancia alimentaria y económica. a) Género tilapia. b) GéneroDormitator" Se agradece la colaboración técnica de Juan Carlos Lara y de Roberto Arreguín E. en el desarrollo del presente trabajo. LITERATURAAL-AISH, M. Human chromosome morphology. I. studies on human chromosome characterization, classification and karyotyping.Can. Journ. Gen. and Cytol.196937O-38111 ARAI, R. y Koike, A.Bull. Nat. Sci. Mus. A karyotype study on two species of freshwater fishes transplanted into Japan(Tokyo)1980275-2786 BADR, E. A. y El-Dib, S. I.Cytological studies on three species of ciichlid fish. Egypt. J. Genet. Cytol.197744-516 BOWERS, J. H., Baker, R. J. y Smith, M. Chromosomal, electrophoretic and breeding studies of selected populations of deer mice (Peromyscus maniculatus) and black-eared mice (P. melanotis) Evolution1973378-38627 CASTORENA SÁNCHEZ, I., URIBE ALCOCER, M. Y ARREGUIN ESPINOSA, J. Estudio cromosómico de poblaciones del género Tilapia Smith. (Pisces, Cichlidae) provenientes de tres regiones de México. Veterinaria, Méx.1983137-14114 JAKOWSKA, S.Spermatogenesis in the cichlid fish Tilapia macrocephal (Beeker). Trans. Am. Microsc. Soc.1950403-41369 JALABERT, B., KAMMACHER, P. Y LESSENT, P. Determinisme du sexe chez les hybrides du gendre Tilapia. Ann. Biol. Anim. Bioch. Bioph.1971155-16511 KORNFIELD, I.L., RITTE, U., RICHLER, F. Y WAHRMAN, J.Evolution Biochemical and cytological differentiation among cichlid fishes on the Galilee.19791-1433 KRISHNAJA, A.P. Y REGE, M.S.Some observations on the chromosomes of certain teleosts usign a simple method. Indian J. Exp. Biol.1980268-27018 LARA PEREA, J.C. Tesis Profesional Estudios de bandeo cromosómico en trucha arco iris (Salmo gardnieri)". Facultad de Ciencias, UNAM.1984 LEVAN, A., FREDGA, K. Y SANDBERG, A. HereditasNomenclature for centromeric position of chromosomes.1964201-22052 MAJUMDAR, K.C. Y MCANDREW, B.J.GeneticaRelative DNA content of somatic nuclei and chromosomal studies in three genera, Tilapia, Sarotherodon, and Oreochromis of the tribe Tilapiini (Pisces, Cichlidae).1986175-18868 MALDONADO MONROY, M.C., URIBE - ALCOCER, M., ARREGUIN ESPINOSA, J. Y CASTRO PÉREZ. A.CytologiaKaryotypical studies on Dormitator maculatus Bloch and Gobiomorus dormitor Lacepede (Gobiidae: Perciformes).(Tokyo)198515-2150 MAJUMDAR, K.C. Y MCANDREW, B.J.GeneticaRelative DNA content of somatic nuclei and chromosomal studies in three genera, Tilapia, Sarotherodon and Oreochromis, of the tribe Tilapiini (Pisces, cichlidae)1986175-18868 MCPHAIL, J.D. Y JONES, R. L.A simple technique for obtaining chromosomes from teleost fishes. J. Fish. Res. B. Canada1966767-76812 MICHELE, J.L. Y TAKAHASHI, C. S.Cytolgia Comparative Cytology of Tilapia rendallii and Geophagus brasiliensis (Cichlidae, Pisces).(Tokyo)1977535-53742 NATAJARAN R. Y SUBRAHMANYAM, J.A preliminiry study on the chromosomes of Tilapia mossambica (Peters). curr. Sci.1968262-26337 POST, A. Zool Syst. Evolutionsforsch.Vergleichende untersuchungen der Chromosomen zahlen bei Susswasser-Teleosteern. Z.196547-933 PRASSAD R. Y MANNA, G.K.Chromosomes of the fishes Tilapia mossambica and Notopterus notopterus. Chrom. Inf. Ser. Univ. Kalyani.197611-1312 ROSAS-MORENO, M. Peces dulceacuícolas que se explotan en México y datos sobre su cultivo. Inst. Nac. de Pesca. Centro de Estudios Económicos y Sociales del Tercer Mundo., A.C.197675-85 SECRETARÍA DE PESCAAnálisis de la Actividad Pesquera. Sistema Nacional Integrado de Información del Sector Pesca.1986Septiembre 1986. THOMPSON, KW.Thesis Some aspect of chromosomal evolution of the Cichlidae (Teleostei, Perciformes) with emphasis of neotropical forms. Ph. D. The University of Texas al Austin.1976 THOMPSON, K.W. Karyotypes of six species of African Cichlidae (Pisces: Perceiformes).Experientia1981351-35237 URIBE-ALCOCER, M., ARREGUÍN-ESPINOSA. J., TORRES-PADILLA, A. Y CASTRO PÉREZ, A. An. Inst. Cienc. del Mar y Limnol. Los cromosomas de Dormitator latifrons (Pisces: Gobiidae).Univ. Nal Auton. México198323-3010 VERVOORT, A.Cytologia The karyotypes of seven species of Tilapia (Teleostei: Cichlidae).(Tokyo)1980651-65645
|