|
EFECTO DE LA
MORFOLOGÍA DEL SUSTRATO EN EL DESARROLLO DE LA COMUNIDAD CORALINA
Trabajo recibido el 9 de junio 1988 y aceptado para su publicación
el 6 de febrero de 1989.
Eric Jordán Dahlgren
Instituto de Ciencias del Mar y
Limnología, Estación "Puerto Morelos", Apdo. Postal 1152, Puerto
Morelos 77500 Q. R. México. Contribución No. 649 del Instituto de
Ciencias del Mar y Limnología,
UNAM.
Los arrecifes coralinos del la región Centro-Norte
del litoral de Quintana Roo, frente al Mar Caribe, se caracterizan por un pobre
desarrollo coralino en la zona frontal; mientras que en las zonas protegidas,
el desarrollo es elevado. Esta atípica conformación predomina a
lo largo de los arrecifes coralinos del Caribe Mexicano, por más de 300
km lineales, tanto en el litoral continental como en el insular. Un
análisis comparativo entre áreas de bajo y alto desarrollo
coralino en la zona frontal de arrecifes de esta región, permiten
formular un sencillo modelo conceptual que pretende explicar el porqué
de esta atípica pero extensa conformación arrecifal. Los
elementos de este modelo se basan en: 1) La presencia de un sustrato arrecifal
de bajo relieve y escasa pendiente, que resulta de la morfología de la
plataforma continental. 2) Que esa disposición del sustrato favorece la
existencia de condiciones de alta severidad ambiental para corales
escleractíneos por deposición y resuspensión
contínua de sedimento no estabilizado y, 3) Que durante el
período formativo de la comunidad, ha existido una alta frecuencia
relativa de efectos catastróficos producidos por ciclones, que destruyen
las incipientes formaciones
arrecifales.
An atypical reef community structure extends for more than
half of the coral reef tract on the Eastern Yucatán coastline (over 300
km).,One of the main resultants of this configuration is a poorly developed
scleractinian community on the fore reef, that in turn determines the absence
of a depositional matrix. Based on a comparison of community structure in
excepcionally well developed sites on the for reef, a simple model that
explains this type of coral community development is discussed. Basic elements
of the model are: 1) Low relief sustrate in a low angled slope, as a result of
the processes that shaped the continental platform; 2) Conditions that
encouraged the continuos accumulation and resuspension of reefal sediments,
therefore imposing severe environmental conditions on scleractinian corals and,
3) relative high frecuency of hurricane and severe storms that disrupt the
rising reef structure on the fore reef
La mayor parte del litoral de México, frente al Mar Caribe, se encuentra bordeado por una extensa formación arrecifal. En la parte Centro-Norte de ese litoral, los arrecifes coralinos se caracterizan por un bajo desarrollo de la zona frontal a la vez que presentan un elevado desarrollo coralino en la zona posterior (Jordán, 1979; 1989). Esta situación es atípica ya que en condiciones normales, un arrecife coralino muestra un crecimiento considerablemente mayor en la zona frontal que en la posterior; situación que resulta entre otras cosas, de que el propio desarrollo del arrecife genera condiciones subóptimas para los corales escleractíneos en las zonas protegidas (Darwin, 1842; Schlanger, 1981). A pesar del bajo grado de desarrollo coralino en la zona frontal, la riqueza y diversidad específicas son elevadas (Jordán et al, 1981), así como la abundancia y diversidad de otros componentes de la comunidad coralina (particularmente los gorgonáceos (Jordán (a), En prensa), que al igual que los corales son organismos sésiles, coloniales y simbióticos), lo que indica claramente que las condiciones ambientales son adecuadas para el crecimiento de la biota arrecifal coralina. Por lo tanto, deben existir algunos factores que específicamente, inhiben el desarrollo de los corales escleractíneos en la zona frontal. Jordán (1980) y Jordán et al., (1981) basándose en un estudio de la estructura comunitaria de escleractíneos, propusieron la hipótesis de que esta situación obedece a la existencia de un ambiente severo, generado por acumulación de sedimento libre sobre el sustrato arrecifal. Estudios posteriores mostratron la existencia de lugares excepcionales dentro de la región Centro-Norte en los que el desarrollo de corales escleractíneos, así como el de la propia estructura arrecifal de la zona frontal, es elevado (Jordán, 1988). Un análisis de las diferencias en la morfología arrecifal y en la estructura de la comunidad coralina de los ambientes de alto y bajo desarrollo coralino, permiten formular un sencillo modelo conceptual que pretende explicar el escaso desarrollo coralino en la zona frontal de los arrecifes de esta región del Caribe Mexicano. ÁREA DE ESTUDIOEl área con bajo desarrollo coralino en la zona frontal, se extiende desde la Punta Pulticub (19º05' LN) hasta la Isla Contoy (21º30' LN), a lo largo del litoral continental de Quintana Roo (Figs. 1 y 2). Esta situación predomina también a sotavento de la Isla de Cozumel (Jordán, 1988) y en la parte noroeste del Banco Chinchorro (Jordán y Martín, 1987). Dentro de esta extensa región se encontratron cinco localidades en las que se observó un excepcionalmente alto grado de desarrollo coralino en la zona frontal: Punta Allen, Akumal y Punta Petempich en el litoral continental; Punta Chiqueros y Microatolones (Cocos), en la Isla de Cozumel (Fig. 1). Lo más sobresaliente de estos lugares es que el área de alto desarrollo coralino no se encuentra a lo largo de todo el perfil arrecifal, sino que existe únicamente en áreas aisladas de la zona frontal, las que se caracterizan por la existencia de un fuerte relieve del sustrato y en cuyas crestas se desarrolla la comunidad coralina. Este tipo de sustrato de alto relieve (del orden de varios metros) es de extensión limitada (cientos de metros) y se encuentra en un rango de profundidades entre 5 y 35 m (Fig. 3). En contraste, el sustrato dominante en la región tiene un relieve considerablemente menor a un metro (medido entre las cimas y las depresiones), y se extiende sin grandes variaciones a todo lo largo del perfil arrecifal. Con el objeto de probar la hipótesis de que la diferencia en el grado de relieve del sustrato basal es determinante en el desarrollo de la comunidad coralina, se seleccionaron tres localidades con sustrato de diferente relieve: Puerto Morelos (Bajo Relieve), Punta Maroma (relieve intermedio) y Punta Petempich (Alto Relieve). En estas localidades se realizaron censos y observaciones tendientes a cuantificar las diferencias en el grado de desarrollo de la fauna escleractínea. MÉTODOSEn cada una de las localidades indicadas, a una profundidad de 10m, se estimó la abundancia de las diferentes especies de corales escleractíneos que cubren el sustrato arrecifal. El censo se realizó mediante el uso de transectos unidimensionales, donde la frecuencia y magnitud de intercepción de las colonias de coral con el transecto, determina la importancia relativa de cada especie (Loya, 1972; 1978). La longitud de los transectos fué definida en 20m, con base en la curva que resulta de graficar la longitud del transecto contra el número acumulativo de especies (Fig. 4), considerando que el punto donde la curva tiende a la asíntota señala el largo adecuado (Gleaon, 1922; Loya, 1972; Jordán, 1980). Cinco de estos transectos dispuestos al azar, sobre una zona definida constituyen un tamaño de muestra adecuado de acuerdo con curvas de diversidad acumulativa (Pielou, 1975), obtenidas en un estudio anterior (Jordán, 1980). El transecto es delimitado por cadenas plásticas (que siguen el variable contorno del sustrato) y las mediciones de cobertura son estimadas contando el número de eslabones que cubren a cada colonia de coral que queda bajo el transecto (un eslabón mide 2.3 cm); a la vez, cada colonia es identificada a nivel de especie. Con los datos de los censos de corales escleractíneos se obtuvieron los siguientes indicadores de estructura: Diversidad específica (H', Shanon-Weiner); Equitabilidad (J', Pielou) y Porcentaje de cobertura. 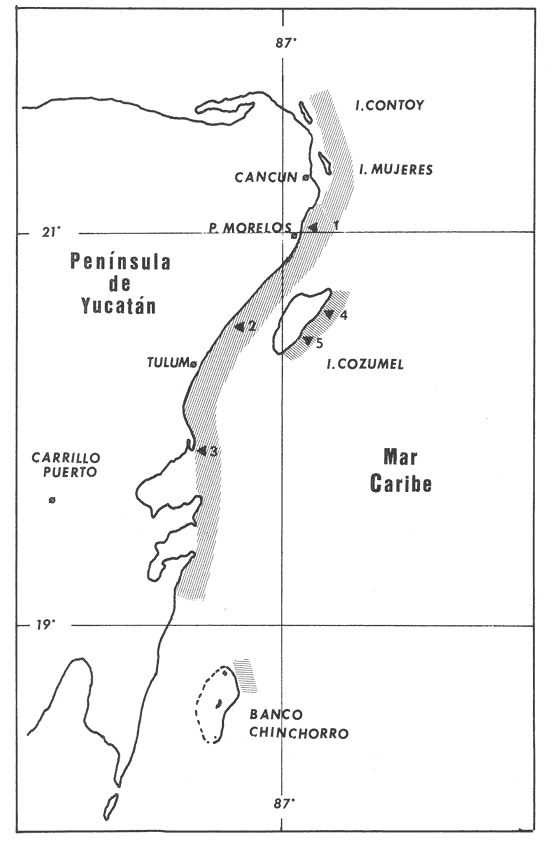 Figura 1. Mapa de Localización. El área sombreada sobre el litoral continental e insular, muestra la región en que predominan los arrecifes con bajo desarrollo coralino en la zona frontal. Las flechas señalan la localización de sitios donde el desarrollo coralino es excepcionalmente elevado en la zona frontal: 1= Punta Petempich; 2= Akumal; 3= Punta allen; 4= Cocos (Microatolones); 5= Punta Chiqueros. 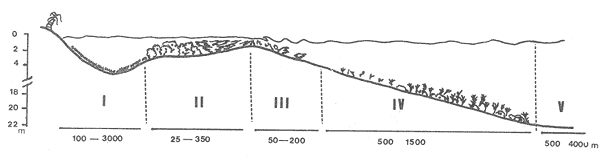 Figura 2. Patrón de Zonación. Perfil perpendicular a la costa mostrando la zonación y morfología arrecifal en los arrecifes coralinos de la zona de estudio. I= Zona Lagunar, II= Zona Posterior. III = Zona de Rompiente; IV= Zona Frontal; V= Plataforma Arenosa. 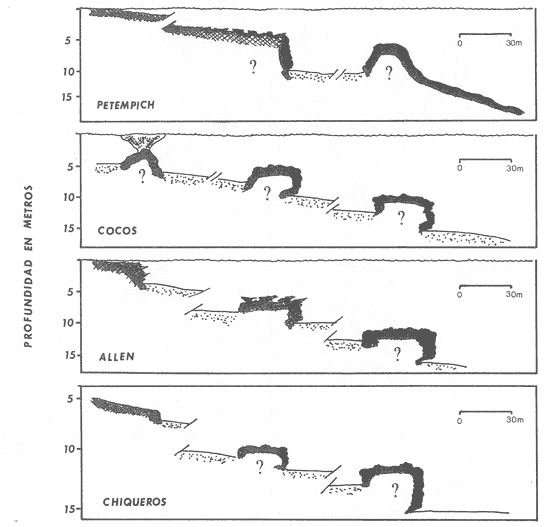 Figura 3. Perfil de la Zona Frontal en Arcas con alto relieve (Comparar con el perfil que aparece en la fig. 2).. La escala vertical se aplica a todo pefil, mientras que la horizontal solo es válida entre las marcas que dividen el perfil de cada localidad. las interrogaciones señalan los lugares en que se desconoce la composición interna de los parapetos, aunque el exterior indica calizas consolidadas. Los parapetos someros de Punta Allen están cubiertos por Acropora palmata El tamaño de la colonia se estimó midiendo el diámetro mayor y el diámetro menor de todas las colonias, que quedaron bajo transectos similares. Diámetros que se promediaron para cada colonia, asumiendo que la forma de la colonia en un plano, tiende a la forma circular por su forma de crecimiento (Weinberg, 1981; Marsh et al., 1984). El tamaño de la muestra, se definió arbitrariamente a partir de que la varianza acumulativa de tamaño fuera igual o menor al 5% del valor máximo. Se estimó la cantidad de sedimento que puede depositarse en un punto dado del arrecife en áreas con bajo y alto relieve. Los colectores (2 por cada sitio de muestreo) se colocaron a 10 m de profundidad, en dos posiciones: Una sobre el fondo, entre colonias de coral, y otro a 1.0 m de altura sobre el fondo (fijos a barretas de hierro afianzadas entre los resquicios del sustrato), cuidando que la posición de los colectores fuera horizontal. Las muestras se obtuvieron con colectores redondos de 20 cm de diámetro. La tapa superior del colector tiene la forma de un embudo invertido que termina en un orificio de 2.5 cm de diámetro en el centro, esta forma evita que el sedimento atrapado sea resuspendido y transportado fuera del colector por la acción del oleaje. En todos los casos se evitó poner los colectores cerca de lugares donde se acumulara el sedimento naturalmente. Los colectores se retiraron una vez a la semana, cubriéndolos bajo el agua con una tapadera ajustable y transportándolos así, hasta la superficie. El sedimento fué trasvasado en la lancha a bolsas de plástico, previamente marcadas, mediante suaves chorros de agua producidos con jeringas plásticas de 25cc de capacidad. Los colectores eran vueltos a colocar en su posición correspondiente inmediatamente después. Posteriormente en el laboratorio el sedimento fué secado y pesado en una balanza analítica. El muestreo tuvo una duración de 9 semanas entre enero y marzo. 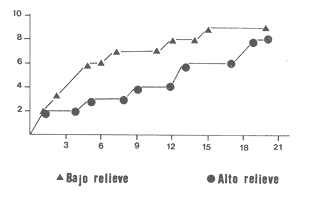 Figura 4. Tamaño del Transecto. La gráfica indica el número acumulativo de especies de corales escleractíneos en función de la longitud del transecto, en áreas con diferente relieve. RESULTADOSSedimentación.-La cantidad de sedimento retenido en las parejas de colectores no fué significativamente diferente, por lo que se promediaron los valores en cada sitio de muestreo. Las diferencias entre los sitios de muestreo fueron significativas, tanto entre las muestras del fondo y las muestras alejadas del fondo, como para los dos ambientes muestreados: bajo relieve y el de alto relieve (Tabla 1). Se observa también que la cantidad de sedimento retenido entre los dos ambientes es claramente diferente, lo que reafirma que un grado de desarrollo coralino elevado contribuye en forma significativa a estabilizar el sedimento no consolidado (Stoddart, 1969), mientras que en ausencia de este desarrollo no existe un control del mismo. El diseño del muestreo, que solo pretendió definir la existencia de diferencias en la dinámica del sedimento entre los dos ambientes, no permite dilucidar si la cantidad de sedimento colectada corresponde a niveles de "stress" o incluso letales, para los corales escleractíneos del área de estudio. Sin embargo, en un estudio de la capacidad de rechazo del sedimento por corales escleractíneos, en condiciones naturales, Bak y Elgershvizen (1976) encuentran que constructores arrecifales importantes comoAcropora palmata y Montastrea annularis, tardan más de 24 horas en rechazar de 1 a 3g de arena sobre el corallum, dependiendo de las condiciones locales. Por lo que en principio, se puede considerar que la cantidad de sedimento retenido en los colectores es elevada en términos de la capacidad de rechazo de sedimento que tienen los corales. Sin embargo, parece ser que la eficiencia del rechazo de sedimento es seriamente influenciada por las condiciones del habitat y de otros factores como la forma, tamaño y disposición de la colonia (Bak y Elgershvizen, 1976). 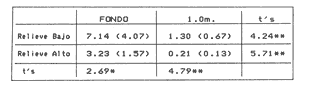 TABLA 1 CONTENIDO DE SEDIMENTOS EN COLECTORES. VALOR PROMEDIO DE DOS COLECTORES POR SEMANA(7 DIAS). FONDO= DIRECTAMENTE SOBRE EL SUSTRATO ARRECIFAL; 1.0m= A UN MEMO DE ALTURA SOBRE EL SUSTRATO; t's= VALOR DE LA PRUEBA DE t, AJUSTADA PARA VARIANZAS DESIGUALES(SOKAL y ROI ILF, 1981); *= SIGNIFICATIVOAL 0.05 DE PROBABILIDAD; **= SIGNIFICATIVO AL 0.001 DE PROBALIDAD La capacidad de rechazar el sedimento varía fuertemente entre especies, por lo que la identidad de las especies dominantes puede ser utilizado como un indicador biológico de ambientes más o menos afectados por sedimento libre. Hubbard y Pockock, (1972) y Kohlemainen, (1973), coinciden en señalar que las especies más resistentes a altas tasas de sedimentación sonSiderastrea siderea, Montastrea cavemosa y Diploria strigosa. Loya (1976) comparando la comunidad coralina de ambientes arrecifales con alta y baja tasa de sedimentación relativa en arrecifes de Puerto Rico, encuentra una estructura radicalmente diferente y reporta que son estas tres especies las que dominan en el ambiente con alta dinámica de sedimento. Posteriormente Jordán (1980) y Jordán et al., (1981) determinan que en el NE de la Península de Yucatán, esas mismas tres especies son las que dominan en los sustratos de bajo relieve; mientras que este estudio señala que en los ambientes de alto relieve dominan Acropora palmata y Montastrea annularis. Desarrollo y Estructura. Para determinar lo favorable que pueda ser un ambiente para el desarrollo coralino en las áreas de alto y bajo relieve, se utilizan en esta estudio dos indicadores relativos: El porcentaje de cobertura del sustrato por corales escleractíneos y el tamaño medio de colonia de coral bajo el transecto. El grado de cobertura indica lo "adecuado" del ambiente para estos organismos, que en condiciones óptimas tienden a cubrir totalmente el sustrato arrecifal (Bradbury y Young, 1983; Hubbard, 1986). El tamaño medio de la colonia es utilizado como un indicador relativo de la estabilidad temporal de esas condiciones ambientales; ya que tiende a haber una relación positiva entre el tamaño de una colonia y el tiempo transcurrido desde su implantación (Buddemeir y Kinzie, 1976; Dustan, 1975). Se encontró que en términos de estos parámetros existe una clara diferencia entre las zonas con sustrato de alto relieve y las de bajo relieve (Fig. 5); indicando que las condiciones son mejores y más estables para el crecimiento de los corales escleractíneos en las áreas de alto relieve que en las de bajo relieve. Los datos de las áreas con bajo y medio relieve, no mostraron una diferencia significativa en términos del porcentaje de cobertura (Man-Whitney U = 20; p > 0.20), lo que se debe a la amplia desviación que muestra el grupo de medio relieve. En términos del tamaño medio de colonias la diferencia si es significativa (Man-Whitney U = 24: 0.05 > p > 0.01). Por lo que aparentemente en las áreas de medio relieve las condiciones son sólo marginalmente mejores que en las de bajo relieve, lo que sugiere que es necesario un grado de relieve mínimo, relativamente fuerte, para que exista un alto grado de desarrollo del componente escleractíneo en la zona de estudio. Hecho que podría estar relacionado con la dinámica del sedimento libre, entre otros factores. 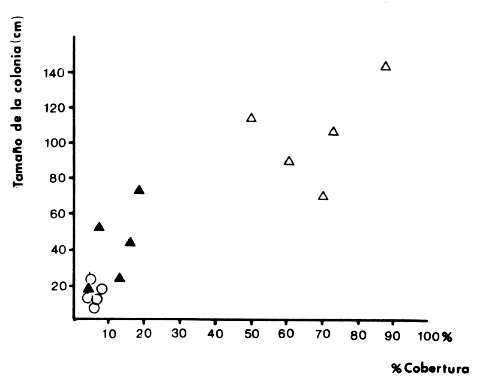 Figura 5. Porcentaje de Cobertura y Tamaño Medio de Colonia en Arcas con sustrato de Diferente Relieve. 0= Bajo relieve (Puerto Morelos); s = Relieve Intermedio (Punta Maroma); s= Alto Relieve (Punta Petempich). En términos de otros indicadores de estructura la separación entre los diferentes ambientes es similar a la que resulta de considerar el desarrollo relativo (Fig. 6). Si bien, el número de especies (S) por transecto tiende a ser mayor en las áreas de alto relieve, las amplias variaciones a escala local determinan que estas diferencias no sean significativas (Kruskal-Wallis, 0.10 < p < 0.20 ). De hecho, el número total de especies de cada uno de los ambientes de relieve (Bajo = 16; Medio = 19; Alto = 17), no son realmente diferentes. En cambio, en términos de la diversidad específica (H'), si existen diferencias significativas entre los tres ambientes de relieve (Kruskal-Wallis; 0.01 > p > 0.05). Un análisis posterior con una prueba de rango múltiple, similar a la de Newman-Keuls (Zar, 1974), indicó que las diferencias solo son significativas entre el ambiente de alto relieve y los otros dos. Esto se debe a que en el ambiente de alto relieve existe una marcada bidominancia de Acropora palmata y Montastrea annularis. No obstante lo anterior, la riqueza y la composición específica de la fauna escleractínea del área de estudio (Jordán et al., 1981), son similares a la que se encuentra a la misma profundidad en otros arrecifes del Caribe (Goldberg 1973; Loya, 1976; Liddell y Olhorst, 1987); lo que lleva a considerar que las diferencias observadas en el desarrollo y en la estructura comunitaria de los tres ambientes, corresponden a diferencias de crecimiento y expansión colonial, y no a condiciones limitantes en el reclutamiento de las diferentes especies que constituyen la comunidad. DISCUSIÓNRelieve del Sustrato Arrecifal.- Los resultados sugieren que efectivamente hay una relación entre los sitios de la zona frontal del arrecife, que presentan un alto relieve topográfico y un alto desarrollo coralino. 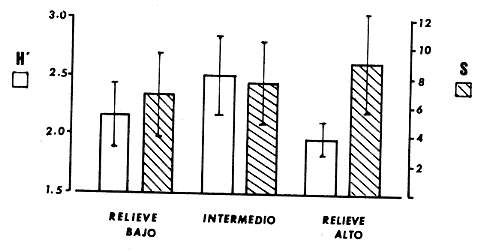 Figura 6. Indicásemos de Estructura de la Comunidad de Escleractineos en Arcas con Sustrato de Diferente Relieve. Bajo relieve (Puerto Morelos) ; Relieve Intermedio (Punta Maroma); Alto Relieve (Punta Petembich). H'= Indice de Diversidad de Shanon-Wiener; S= Número de especies. Las barras verticales dentro de las columnas señalan el valor de una deesviación estandard. Esta situación concuerda con la tésis de que no bastan condiciones hidrológicas apropiadas, sino que se requiere además de una adecuada morfología del sustrato basal para que un arrecife mantenga un proceso de acreción sostenido. Este planteamiento nace de que debido al rápido incremento del nivel del mar durante la transgresión Holocénica (Ginsburg y Choi, 1983; Digerfeldt y Hendry, 1987), un larga etapa formativa implicaba el riesgo de que la biota coralina quedara a demasiada profundidad, donde las tasas de crecimiento de los corales disminuyen drásticamente y por ende, de la propia estructura arrecifal. (Dustan, 1975; Davies et al., 1985; Jordán (b), en prensa). En la zona de estudio, los sustratos de alto relieve no parecen corresponder a una formación coralina reciente, sino a viejos escarpes costeros que forman parapetos parcialmente conservados en unas cuantas localidades aisladas. Esta opinión se basa en que no se observó que estas estructuras estuvieran constituídas por una trama arrecifal, sino por calizas sedimentarias bien consolidadas (Jordán, 1988). A la vez, los parapetos tienden a una disposición paralela a la línea de costa y están confinados a un mismo rango de profundidad (Fig 3). En la mayor parte del litoral sin embargo, predomina el sustrato de bajo relieve, constituído por un pavimento calcáreo erosionado que no corresponde con la morfología de un proceso de acreción arrecifal reciente y que de acuerdo con Land (com. pers.), debe corresponder al Pleistoceno. Por lo que es probable que la topografía de bajo relieve y la baja inclinación de la pendiente, resulten de un proceso erosivo por oleaje durante los cambios de nivel del mar que antecedieron al Holoceno; en forma similar a como fueron formadas las terrazas submarinas que, entre 10 y 20 m de profundidad, se encuentran en el Caribe (Logan, 1969). La presencia de profundos cortes basales (Notches) en la base de los parapetos que dan lugar al alto relieve, indican procesos erosivos por oleaje en períodos interestadiales. Ambiente de Alta Severidad.- Uno de los efectos que resultan de la suave pendiente de la zona frontal, es la acumulación de sedimento libre producido por la contínua degradación de múltiples organismos arrecifales (Stoddart, 1969). Sedimento que en ausencia de una trama arrecifal que lo estabilice, es fácilmente resuspendido, transportado y vuelto a depositar por efecto del oleaje. Esta dinámica del sedimento genera condiciones de alta severidad ambiental para los corales escleractíneos, debido a la naturaleza mucosa de su epitelio que les sirve para la captura de partículas alimenticias por adhesión (Muscatine, 1973); por lo que cuando hay sedimento en suspensión, el epitelio del coral atrapa los granos de sedimento al igual que si fueran alimento. La acumulación de este material no asimilable, sobre la superficie de la colonia puede llegar a bloquear el intercambio agua-tejido del coral, afectando seriamente el metabolismo del par simbionte. La importancia de este evento en la sobrevivencia de los corales es muy alta, al grado de que los corales tienen mecanismos específicos para eliminar el sedimento acumulado (Hubbard y Pockock, 1972; Kohleminen, 1973; Bak y Elgershvizen, 1976). Este proceso sin embargo, es lento y energéticamente costoso, ya que entre otras cosas, implica la producción masiva de mucus, que es muy rico en material protéico (Benson y Muscatine, 1974; Duchklow y Mitchell, 1979). Si la tasa de sedimentación sobre el corallum es más elevada que la capacidad del coral para deshacerse del sedimento acumulado, la parte directamente afectada o incluso toda la colonia, morirá eventualmente. Si la tasa de sedimentación no llega a niveles letales, pero es relativamente alta para la capacidad de rechazo del coral, la pérdida constante de energía que éste sufre al mantener el mecanismo de limpieza puede afectar su desarrollo. Lo que resulta en colonias pequeñas y relativamente frágiles a otras causas naturales de "stress". Durante tormentas, el sedimento libre es fácilmente resuspendido y transportado rápida y violentamente, produciendo un efecto abrasivo sobre los tejidos blandos de los corales (Hubbard y Pockock, 1972; Bak y Elgershvizen, 1976). Dependiendo de la magnitud de la pérdida de tejido, el resultado puede ser destrucción parcial o total de la colonia. Ambos efectos, el producido por oleaje cotidiano y el producido por tormentas, tienen un denominador común, son más intensos cerca del fondo que a cierta distancia del mismo; lo que implica que la severidad ambiental corresponde principalmente a un efecto de fondo, afectando a colonias pequeñas o recién formadas. La importancia de este efecto de fondo es tal, que en términos de áreas restringidas puede mantener a la fauna escleractínea, en un estado de desarrollo incipiente, como ya ha sido observado (Loya, 1976). Sin embargo, con respecto al área de estudio que es muy extensa, debe de considerarse la posibilidad de que aleatoriamente existieran habitats protegidos, los que favorecerían eventualmente un desarrollo normal de los corales. Además, ha transcurrido un largo período desde la etapa formativa, durante el cual es probable que hayan existido cambios en las condiciones climáticas, determinando ciclos de severidad ambiental variable. Por lo que sería de esperarse que en los más de 5000 años que lleva inundada la zona frontal del arrecife del área de estudio, hubieran existido numerosas ocasiones en las que algunas colonias alcanzaran una talla mínima, que las hiciera menos sensibles a este efecto de fondo. Conforme la colonia continúa creciendo a partir de ese umbral, el efecto del sedimento libre en la sobrevivencia de la colonia sería cada vez menor. El crecimiento sostenido de esas colonias "Invulnerables" (sensu Connell, 1975) al efecto de fondo, iría modificando gradualmente el ambiente y disminuyendo la severidad ambiental en su área de influencia, favoreciendo por tanto el desarrollo de otras colonias. Este proceso, teóricamente factible, sería muy lento al principio, pero conforme transcurre el tiempo daría lugar a una transformación gradual de la comunidad coralina de la zona frontal, cuyo desarrollo se volvería más acelerado gradualmente hasta alcanzar a formar una trama arrecifal bien constituída. Eventos Catastroficos.- Sin embargo, el que los altos desarrollos coralinos en la zona frontal estén restringidos a la presencia de crestas calcáreas previamente formadas y el que predomine la comunidad poco desarrollada del sustrato de bajo relieve, indica que existen otros factores que determinan que "escapes" a la situación predominante, sean eventualmente destruídos. Los procesos de destrucción natural más probables en este caso, corresponderían a procesos de bioerosión y al impacto de ciclones sobre la incipiente estructura arrecifal. Bioerosión.- La variedad de mecanismos y la multitud de organismos que participan en los procesos de bioerosión (desde bacterias hasta peces; Hutchings, 1986), determinan que la magnitud de los procesos bioerosivos en la dinámica arrecifal sea tan elevada, que de no haber un activo proceso de crecimiento que compense la destrucción biológica, la trama arrecifal puede ser debilitada al grado de que un arrecife sea eventualmente destruído (Stoddart, 1969; Ginsburg 1983). De hecho, se considera que debido a la magnitud de estos procesos, en la historia de un arrecife no existe un estado único de crecimiento, sino que el arrecife crece en unas épocas -sobre todo durante transgresiones marinas o subsidencia del litoral- y es parcialmente destruído en otras; en un balance dinámico a largo plazo (Hopley, 1982; Davies, 1983). Risk y MacGeachy (1978) han descrito secuencias sucesionales de biohoradadores que resultan en la destrucción de más del 70% de los cabezos de coral estudiados, debilitando considerablemente la trama arrecifal. Estos resultados corresponden únicamente a la acción de biohoradadores y no incluyen el efecto de organismos raspadores y rebajadores, que contribuyen significativamente al proceso destructivo (Ver a Hutchings, 1986; para una discusión completa sobre el tema). Aparte de la producción de sedimento, los procesos de biohoradación resultan en la fragmentación de la colonia por debilitamiento de su estructura. Una vez fragmentada la colonia pueden suceder una de dos cosas: a) Si la colonia se encuentra en un arrecife bien constituído, existe una relativamente alta probabilidad de sobrevivencia para los fragmentos que pueden quedar atrapados e inmobilizados en la trama arrecifal; llegando a generar eventualmente nuevas colonias (Highsmith, 1982). b) Si como es el caso, en la zona de estudio, esto sucede en condiciones donde no existe una trama arrecifal bien constituída, los fragmentos resultantes quedan libres sobre el fondo y sujetos a la acción del oleaje de tormenta, con una alta probabilidad. de ser eventualmente destruídos. Además, estos fragmentos pueden quedar por abajo del la talla crítica que hacía a la colonia "invulnerable" (sensu Connell, 1975) al efecto de fondo. Uno de los resultados de esta acción bioerosiva en el área de estudio, sería la de retrasar aún más, el ya de por sí lento desarrollo de la fauna escleractínea y por ende de la trama arrecifal en la zona frontal. Ciclones.- Este proceso de destrucción biológica está íntimamente asociado con la destrucción física provocada por ciclones y tormentas de alta intensidad, durante los que colonias previamente debilitadas o fragmentadas por procesos bioerosivos, son rápidamente destruídas por el oleaje que producen estos eventos meteorológicos. Se ha observado que incluso en arrecifes muy bien desarrollados, los ciclones tiene invariablemente un efecto desvastador sobre la comunidad coralina desde la superficie hasta 20 m de profundidad o más, dependiendo de la topografía arrecifal; mientras que la extención de daño catastrófico puede alcanzar hasta más de 50 km, dependiendo del ángulo de incidencia y de la trayectoria del ciclón, con respecto a la dimensión y disposición de la formación arrecifal afectada (Stoddart, 1963; Woodley et al., 1981; Dollar, 1982; Mah y Stearn, 1986; Harmelin-Vivien y Laboute, 1986). La devastación que produce el ciclón resulta de varios procesos, el más importante de los cuales parece ser la erosión por sedimento, grava y pedazos de coral, que son producidos y desplazados violentamente por el oleaje de largo periódo (Rutzler y Macintyre, 1982), y que en general sólo pueden soportar colonias muy robustas de corales masivos (Stoddart, 1963; Woodleyet al., 1981); lo que aunado a la abrasión sobre las partes blandas de colonias que lograron mantenerse en pie, resulta en una situación catastrófica que destruye la estructura de la comunidad de corales escleractíneos. El oleaje producido por un ciclón no solamente puede destruir a la comunidad coralina, sino a las estructuras arrecifales propiamente dichas, tales como los sistemas de macizos y canales en la zona frontal (Stoddart, 1963; 1974; Hernández-Avilaet al., 1977; Harmelin-Vivien y Laboute, 1986). En el caso de los arrecifes de la zona de estudio, que tienen una suave pendiente a sotavento y que se extienden hasta los 25 m de profundídad como máximo, el efecto de un ciclón seria todavia mayor que en los arrecifes bien constituídos y con fuerte relieve (Suhayda y Roberts, 1977). Este efecto ha sido comprobado localmente por el daño causado a la comunidad coralina de Punta Maroma por el ciclón Allen en 1981. La gran mayoría de las colonias deAcropora palmata de la zona frontal (1 a 4 m de profundidad) fueron destruídas, algunas de las cuales alcanzaban dimensiones considerables. Si bien Acropora palmata es un coral ramificado y por lo tanto relativamente fácil de romper, esta destrucción fué causada por oleaje generado a gran distancia, a más de 160km de los litorales de Quintana Roo. El tiempo de recuperación de la comunidad coralina a estas catástrofes, parece tomar muchos años y está en función de frecuencia de eventos catastróficos durante ese período; mientras que en caso de destrucción de la estructura arrecifal, la reconstrucción puede ser imposible con las bajas tasas de acreción arrecifal actuales (Davies et al., 1985; Neuman y Macintyre, 1985). A la fecha, no se ha logrado obtener información suficiente que permita predecir razonablemente el tiempo teórico de recuperación de una comunidad afectada por un ciclón. Por ejemplo, Stoddart (1963) después de analizar el efecto de huracán "Hattie" en los arrecifes beliceños estimó en 20 a 25 años ese período. Un monitoreo constante del proceso de recuperación de esos arrecifes, lleva a este mismo autor once años después, a afirmar que el tiempo de recuperación sería mucho más largo de lo que él había estimado originalmente (Stoddart, 1974). Por lo que este tipo de eventos puede tener una fuerte influencia en la estructuración de una comunidad coralina, particularmente si la frecuencia de ciclones y tormentas intensas es alta. A este respecto Gentry (1971), considera que la probabilidad de que la trayectoria de un ciclón pase por la costa de Quintana Roo es del orden de 50% por año, basado en un análisis de 100 años (ver Jaúregui et al., 1980). Asumiendo que la periodicidad y trayectorias de ciclones y tormentas tropicales a la fecha conocida (Fig. 7), reflejan un patrón promedio de lo que sucedió durante los últimos miles de años (a partir de cuando se alcanzó un nivel del mar cercano al que prevalece actualmente, hace unos 5000 años; Ginsburg y Choi, 1983; Digerfeldt y Hendry, 1987), puede considerarse que la frecuencia de impactos producidos por ciclones en el área de estudio, sería relativamente elevada. Más aún, sabiendo que se requieren períodos del orden de cientos de años para transformar la topografía arrecifal y que el tiempo de recuperación de la comunidad coralina a eventos catastróficos producidos por ciclones, es mayor que 25 años en arrecifes bien formados. Todos estos argumentos permiten proponer que el bajo desarrollo coralino en la zona frontal de los arrecifes del área de estudio (Fig. 8), proviene de determinantes históricas que han propiciado condiciones de alta severidad ambiental para los corales escleractíneos; lo que aunado a naturales procesos destructivos, determinan que el desarrollo de la incipiente trama arrecifal sea muy lento. Desarrollo que cuando eventualmente alcanza magnitudes importantes, es interrumpido por el efecto de ciclones y tormentas tropicales de alta intensidad, que en virtud de la lentitud del proceso de crecimiento, adquieren una alta frecuencia relativa. 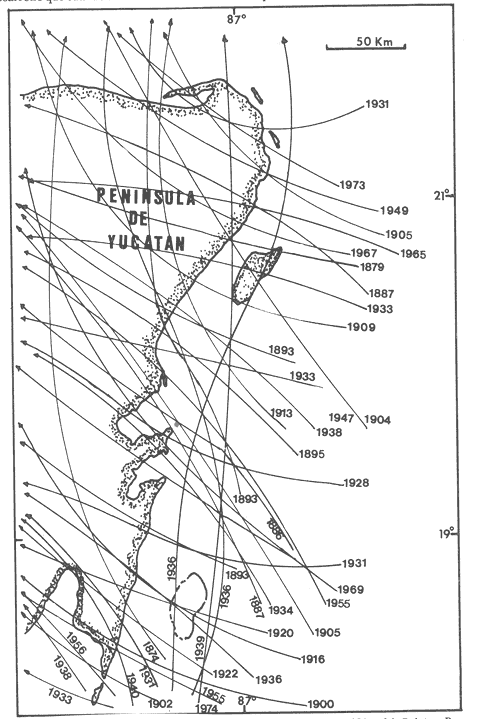 Figura 7. Trayectoria de Ciclones y Tormentas Tropicales que han Iniciado en el Litoral de Quintana Roo, en los Ultimos 100 años. Basado en Gentry (1971) y Jaúregui el al., (1980)  Figura 8. Representación Esquemática de la Secuencia de Desarrollo de los Arrecifes de 1 Area de Estudio.(Ver texto) ----------- = Acreción Arrecifal; Porcentaje de cobertura; _._._.=Tamaño medio de, la colonia de coral. AgradecimientosNumerosas personas han discutido y comentado las diferentes partes de la temática de este trabajo a través de su larga gestación, y a todas ellas quedo muy reconocido por su valiosa contribución. En particular a R. Ginsburg, I, Macintyre, así como a Eduardo Martín. ADDENDUM:Mientras este artículo se encontraba en prensa, la zona de estudio a que se refiere este trabajo fue severamente afectada por el huracán Gilberto, uno de los más intensos de este siglo. Los efectos de este evento en el desarrollo de la comunidad coralina están siendo evaluados y los resultados aparecerán en una proxima publicación. Con base en la información a la fecha obtenida, puede afirmarse que el modelo propuesto se ve confirmado. LITERATURABAK, R.P.M. y J.H.B.M. ELGERSHVIZEN, Patterns of oil sediments rejection in corals.Mar.Biol.,1976.105-113.37(2): BRADBURY, R.H. y P.C. YOUNG,Coral interactions and community structure: an analysis of spatial pattern. Mar Ecol. Prog Ser., 1983. 265- 271.11: BENSON, A.A. y L. MUSCATINE,Wax in coral mucus: energy transfer from corals to reef fishes. Limnol. Oceanogr. 1974.810-814.19: BUDDEMEIR, R.W. y R.A. KINZIE, Coral Growth. Oceanogr. Mar. Biol. Ann. Rev.,1976.183-125.14: CONNELL, J.H., Ecology and Evolution of Communities. The Belknap Press, Some mechanisms pruducing structure in natural communities In: M.L. Cody y J.M. Diamond (Eds.) Harvard Univ,1975.p460-490. DARWIN, C. The structure and Distribution of coral Reefs.Univ. Calif. Press.1842. DAVIES, PJ., Perspectives on coral reefs. AIMS,Reef growth. [In]: DJ. Barnes (Ed.) Australia,1983. 69-106. DAVIES, PJ., MARSHALL, J.F. y D. HOPLEY, Proceedings of the Fifth International Coral Reef Symposium, Relationships between reef growth and sea level in the Great Barrier Reef. In: (B. Salvat, Ed.) Tahiti,1995.105-110.3: DIGERFELDT,G. y M.D. HENDRY,An 8000 year Holocene sealevel record from Jamaica:implications for interpretation of Caribean reef and coastal history. Coral Reefs,1987.165-170.5(4): DOLLAR, S.J., Wave stress and coral community structure in Hawaii. Coral reefs. 1982.71-82.1(2): DUCKLOW, H.W. y R. MITCHELL, Limnol. Oceanogr., Bacterial populations and adaptations in the mucus layers on living corals.1979.715-725.24: DUSTAN, P., Marine Bioloy, Growth and form on the reef-building coral Montastrea annularis.1975.101-107.33: GENTRY, R.C., Hurricanes: one of the major features of air-sea interactions in the Caribbean Sea. Symposium on Investigations and Resources of the Caribbean Sea.UNESCO,1971. p. 79-88. GINSBURG, R.N., Geological and biological roles of cavities in coral reefs.Perspectives on coral reefs. In: DJ. Barnes (Ed.) AIMS, Australia,1983.148-153. GINSBURG, R.N. y D. CHOI, Proceeding of the Great Barrier Reef Conference,Quaternary reefs of Belize, Central America In: Baker et. al., (Eds.) Townsville.1983.p27-42. GLEASON, H.A., Ecology, On the Relationship between speces and area.1922.158-162.3: GOLDBERG, W.M., Bull. Mar. Sci.,The ecology of the Coral-Octocoral communities of the Southeast Florida coast: Geomorphology, species composition and zonation.1973.465-488.23(3): HARMELIN-VIVIEN, M.L. y P. LABOUTE, Catastrophic impact of hurricanes on atoll outer reef slopes in the Tuamotu (French Polynesia). Coral reefs., 1986.55-62.5(2): HERNANDEZ-AVILA, M.L, H.H. ROBERTS y LJ. ROUSE, Proceedings of the third International Reef Simposium.Hurricane generated waves and coastal boulder rampart formation. In: D.L Taylor (Ed.)Miami1977.72-78.2: HIGHSMITH, R.C.,Reproduction by fragmentation in corals. Mar. Ecol. Peog. Sedes,1982.207-226.7: HOPLEY, D., The geomorphology of the Great Barrier Reef. Wiley and sons, Inc.1982.453p. HUBBARD, J.A. y Y.P. POCKOCK, Geol. Rundsch., Sediment Rejection by recent scleractinian corals: A key to paleoenvironmental reconstruction.1972.598-626.61: HUBBARD, D.K, St. Croix, USVI. Coral Reefs, Sediment as a control of reef development:1986.117-126.5(3): HUTCHINGS, P.A., Biological destruction on Coral Reefs: A review. Coral Reefs1986.239-252.4(4): JAUREGUI, E., J. VIDAL y F. CRUZ., Problematica y Perspectivas de Quintana Roo.Los ciclones y tormentas tropicales en Quintana RooUNAM-CIQRO, 1980. P.47-64.período 1871-1978. P JORDAN, E., An. Centro Cienc. del Mar y Limnol. Estructura y composición de arrecifes coralinos en la región noreste de la Península de Yucatán, México.Univ.Nal.Autón. México,1979.69-86.6(1): JORDAN, E., Tesis Doctorado,UACPyP, Arrecifes coralinos del noreste de la Peninsula de Yucatán: Estructura comunitaria, un estimador del desarrollo arrecifal.UNAM,1980.118pp. JORDAN, E., (a, En prensa) Gorgonian community structure and reef zonation patterns on Yucatán Coral reefs. Bull. Mar. Sci. JORDAN, E., An. Centro Cienc. del Mar y Limnol.(b, En prensa). Arrecifes Profundos de Cozumel. Univ. Nal. Autón. México. JORDAN, E., Arrecifes coralinos del Caribe Mexicano: Su potencial de Uso. Informe final,CONACyT-UNAM.1988.200p. JORDAN, E., M. MERINO, 0. MORENO y E. MARTIN, Proceedings of the fourth International Coral Reef Symposium, Community Structure of Coral Reefs in the Mexican Caribbean. In: E. Gómez (Ed.) Manila.1981.303-308.2: JORDAN, E y E. MARTIN, Atoll Res. Bull.,Chinchorro: Morphology and composition of a Caribbean Atoll.1987.310-20. KOLEHMAINEN, S.E. Final Report Mission 8. PRINUL, Siltation experiments on corals in situ. Puerto Rico,1973. 20 p. LOGAN, B.W., Mem. Am. Assoc. Petrol. Geol., Coral Reefs and Banks, Yucatán Shelf, México. In: Logan et al, (Eds.)1969.129-196.11: MAH, AJ. y C.W. STEARN, The effect of Hurricane Allen on the Bellairs fringing reef, Barbados. Coral Reefs,1986.169-176.4(3): MARSH, L.M., R.H. BRADBURY y R.E. REICHELT, Determination of the physical parameters of coral distributions using line transect data. Coral Reefs. 1994.175-180.2(4): MACINTYRE, I.G., Am. Assoc. Petrol. Geol. Bull., Submerged reefs of the Eastem Caribean.1972.720-738.56: MUSCATINE, L, Bioloy and Geoloy of Coral Reefs. Nutrition of Corals. In: Jones y Endean (Eds.) Academic Press.1973, pp. 77-116. NEUMANN, A.C. y I. MACINTYRE, Proceedings of the Fifth International Coral Reef Symposium,Reef responses to sea level rises: keep-up, catch-up or give-up. In: (B. Salvat, Ed.) Tahiti,1985.105-110.3: LIDDELL, W.D. y S.L OLHORST, Bull. Mar. Sci., Patterns of reef community structure, North Jamaica.1987.311-329.40(2): LOYA, Y., Marine Biology, Community Structure and Species Diversity of Hermatypic corals al Eilat, Red Sea.1972.100-123.13(2): LOYA, Y., Bull Mar. Sci, Effects of water turbidity and sedimentation on the community structure of Puerto Rican corals.1976.450-466.26(4): LOYA, Y., Coral Reefs. Research Methods.Monographs on Oceanographic Methodology Plotless and transect methods In: D. Stoddart and M.E. Johanness (Eds.) UNESCO,1978.197-218.5: PIELOU, E.C., Wiley Interscience, Ecological Diversity. N.Y.1975.165 pp. RISK, MJ. y J.K. MACGEACHY, Aspects of bioerosion of Modem Caribbean reefs, R. Biol. Trop.,1978.85-105.26 (supl. 1): ROBERTS, H.H., S.F. MURRAY Y J.N. SUHAYDA, Proceedings of the third International Reef Symposium. Physical processes in a fore reef shelf enviroment. In: D.L. Taylor (Ed.)Miami,1977. 507-516.2: RUTZLER, K. y I.G. MACINTYRE, The Atlantic Barrier Reef Ecosystem at Carrie Bow Cay, Belize. The habitat distribution and community structure of the barrier reef complex at Carrie Bow Cay, Belize. In: (Rutzler y Macintyre (Eds.) Smithsonian Inst. Press.1982.9-45.1: SCHLANGER, W., Geol. Soc. Am. Bull, The paradox of drowned reefs and carbonate platforms.1981.197-211.92: SOKAL, R.S. y F.J. ROHLF, Biometry. Freeman Co.1981. STEARN, C.W. y T.P. COFFIN, Proceedings of the third International Reef Symposium.Carbonate Budget of a fringing reef, Barbados. In: D.L. Taylor (Ed.)Miami,2:471-476.1977. STODDART, D.R., Atoll Res. Bull, Effects of hurricane Hattie on the British Honduras reefs and cays,1963.1-42.95: Oct. 30-31,1961. STODDART, D.R., Biological Reviews, Ecology and Morphology of recent Coral reefs.1969.433-498.44: STODDART, D.R. Proceedings of the Second International Reef Symposium, Post-hurricane effects on British Honduras reefs: Resurvey of 1972. In: Cameron et al., (Eds.) Australia.1974.473-483.2: SUHAYDA J.N. y H.H. ROBERTS, Proceedings of the Third International Reef Symposium.Wave action and sediment transport in fringing reefs In: D.L. Taylor (Ed.)Miami,1977.571-576.2: ROGERS, C.S., y Y.H. SUCHANEK y F.A. PECORA, Bull Mar. Sci,Effect of Hurricane David and Frederick on shallow Acropora palmata reef commuinities: St. Croix, U.S. Virgin Islands. 1982.532-548.32: WEINBERG, S., A comparison of coral reef survey methods. Bidjr. Dierkd. 1981.199-218.491: WOODLEY, J.D., E.A. cHORNESKY, P.A. CLIFFORD, J.B.C. JACKSON, L.S. KAUFMAN, N, KNOWLTON, J.C. LANG, M.P. PEARSON, J.W. PORTER, M.C. ROONEY, K.W. RYLAARSDAM, V.J. TUNNICLIFFE, C.M. WAHLE, J.L WULFF, J.S.G. CURTIS, M.D. DALLMEYER, B.P. JUPP, M.A.R. KOHEL, J.NEIGEL y E.M. SIDES,Hurricane Allen's impact on Jamaican Coral reefs. Science1981.749-755.214: ZAR, J.H., Biostatistical analysis, Pretice-Hall Co. 1974.
|