|
MOLUSCOS DE LA PLATAFORMA CONTINENTAL DE NAYARIT: SISTEMÁTICA Y
ECOLOGÍA (cuatro campañas oceanográficas)
Trabajo
recibido el 9 de diciembre de 1987 y aceptado para su publicación el 4
de mayo de 1988.
M. Reguero
A. Garcia-Cubas
Instituto de Ciencias del Mar y Limnología, UNAM.
Laboratorio de Malacología. Contribución No. 642 del Instituto de
Ciencias del Mar y Limnología, UNAM.
Los trabajos
orientados al conocimiento de la malacofauna del Pacífico Mexicano son
escasos, particularmente en la Plataforma Continental del Estado de Nayarit.
Este estudio tiene como objetivo la obtención de expresiones
cualitativas y cuantitativas, de la composición y distribución de
las diferentes especies que conforman la comunidad de moluscos
bentónicos en el área, así como las relaciones entre tipos
de nutrición y habitat para cada una de las familias. Para tal efecto,
se llevaron a cabo 4 campañas oceanográficas a bordo del B/O "El
Puma" en los años 1982 a 1984, utilizando redes de arrastre
convencionales para captura de macroorganismos bentónicos. El muestreo
se efectuó en un área de barrido de 1'497,177 m² repartida
en 54 estaciones, a profundidades que variaron desde 15 hasta 122 m, con
registros contínuos de temperatura, salinidad y oxígeno disuelto
y análisis de sedimentos. Se cuantificaron 26,600 individuos, de los
cuales 3,262 fueron capturados vivos y 23,338 correspondieron a conchas. Tales
organismos se ubicaron sistemáticamente en 5 clases, 61 familias, 112
géneros y 208 especies; predominando la Clase Gastropoda, tanto en
porcentaje de ejemplares capturados vivos como en número de especies. En
lo referente a abundancia, los Bivalvos representaron el 79 % del total de
organismos recolectados. La convergencia de diversas especies típicas,
tanto de la Provincia Panámica como de la Californiana, confirma el
carácter transicional de la zona de Nayarit. Con base en valores de
densidad absoluta (ind/km²) se estableció una distribución
zonal de los taxa, resultando una mayor concentración de las poblaciones
en la región Noroccidental, debida posiblemente a la confluencia de
corrientes y a la naturaleza del fondo. Para complementar el análisis
sobre estructura de la comunidad de moluscos, se emplearon índices de
diversidad, equitatividad y riqueza de especies, encontrándose los
valores más altos en sedimentos de arena y en la campaña
efectuada en septiembre-octubre de 1984. La caracterización del
área estuvo marcada por la ocurrencia preferencial de 19 especies (15
Bivalvia y 4 Gastropoda), de las cuales Solenosteira gatesiy
Argopecten circularis fueron las más importantes por su amplio rango de
distribución local y mayor abundancia relativa. Asimismo, se registraron
9 especies de moluscos no mencionadas en aguas nacionales y se ampliaron los
límites de distribución geográfica para 19
especies.
Work aimed at the
knowledge of Mexican Pacific malacofauna is rare, specifically with regards to
the continental shelf of the State of Nayarit. The objetive of this study is to
obtain qualitative and quantitative data on the composition and distribution of
the various species which make up the benthic mollusk community in the area, as
well as the relationships among types of nutrition and habitats for each
family. Hence, four oceanographic cruises were performed aboard R/V "El Puma"
from 1982 trough 1984, using conventional trawling nets to catch benthic
macroorganisms. Sampling was done within a sweep area of 1'497,177m²
divided in 54 stations, with depths varying from 15 to 122 m, with continuous
recording of temperature, salinity, and dissolved oxygen as well as performing
sedimentary analysis. A total of 26,600 specimens were quantified, out of which
3,262 were captured alive and 23,338 were shells. Such organisms were
systematically arranged in 5 classes, 61 families, 112 genera, and 208 species.
The class Gastropoda was predominant, both in percentage of specimens captured
alive and in number of species. Abundance-wise, bivalves accounted for 79 % of
the total organisms sampled. The convergence of several typical species - in
the Panamic Province as well as in the Californian Province- confirms the
transitional nature of the Nayarit area. Based on absolute density values
(specimens/km²), a taxa zone distribution was established, observing a
greater concentration of population in the north-westem region, perhaps due to
the confluence of currents and to the nature of the sea-bottom. In order to
supplement the analysis on the mollusk community structure, species diversity,
equitability, and richness indexes were used, observing the highest values in
sandy bottoms and in the cruise which took place during september-october 1984.
The characterization of the area highlighted the preferencial occurrence of 19
species (15 Bivalvia and 4 Gastropoda), of which Solenosteira
gatesiand Argopecten circularis were the most relevant due to their
wide range of local distribution and greater relative abundance. Likewise, 9
mollusks species, not mentioned in domestic waters, were recorded, and the
geographical distribution limits of 19 species were
expanded.
La necesidad de conocer los recursos del mar adquiere en la actualidad el carácter de inaplazable, ante la problemática de suministrar alimento a la creciente población humana. No obstante, para que tales recursos sean susceptibles de utilización, es preciso hacer estudios regionales orientados a la elaboración de inventarios de comunidades faunísticas que proporcionen información sobre diversidad, abundancia y distribución de las especies presentes en una localidad determinada, con el objeto de integrar finalmente un atlas de los recursos marinos nacionales. Ubicado en este contexto y tomando en cuenta que el área de Nayarit reviste gran interés por tratarse de una zona limítrofe entre dos provincias malacológicas (la Californiana y la Panámica) (Parker, 1964), el presente trabajo pretende contribuir al conocimiento de los moluscos de esta región; su importancia, tanto biológica como económica, es bien reconocida, pues además del sitio que ocupan en las cadenas tróficas, el hombre los ha utilizado durante siglos como alimento, alcanzando algunos de ellos un alto nivel de comercialización. Los principales objetivos de este estudio se concretan a investigar las relaciones que existen entre las poblaciones de moluscos y su ambiente marino en el área donde tradicionalmente la flota camaronera captura este recurso, en tanto que los moluscos son un componente importante de la fauna de acompañamiento del camarón. Entre los problemas de mayor interés, figura el de identificar las especies de moluscos que habitan el área de la plataforma continental de Nayarit, su distribución e importancia relativa, algunos factores de orden físicoquímico y biológico que las afectan, así como los sustratos sedimentarios en los que habitan. Al respecto, se observan las variaciones en temperatura, salinidad y oxígeno disuelto en las aguas marinas. ANTECEDENTESLas referencias bibliográficas para esta región del país resultan particularmente escasas, en especial aquéllas que aluden a aspectos relacionados con ciencias marinas. La mayoría de las citas son informes de expediciones científicas en los que se hace mención a especies aisladas de moluscos, o bien son catálogos de la fauna malacológica que habita en las costas del Pacífico oriental. Entre los más importantes puede citarse a Hertlein y Strong (1940-1951) y Keen (1971). En la recopilación bibliográfica sobre el Golfo de California, elaborada por Schwartzlose y Hendrickson (1983), se incluyen alrededor de 40 referencias para el área de Nayarit que tratan diversos tópicos geológicos, geográficos, pesqueros y algunos biológicos sobre aves, peces y crustáceos. Los relacionados con moluscos son monoespecíficos y se refieren casi exclusivamente a cultivo ostrícola en lagunas costeras. El trabajo de Parker (1964) aporta datos sobre comunidades bentónicas en la plataforma continental de la zona de estudio. Dicho autor describe distintos ambientes de acuerdo con la composición faunística, tipo de sedimento, profundidad, temperatura del agua y oxígeno disuelto. Recientemente Hendrickx (1985)In:Yáñez-Arancibia (Ed.), trata sobre macroinvertebrados bentónicos acompañantes del camarón en el área del Golfo de California y discute la importancia de 31 especies de moluscos, como recursos pesqueros potenciales. DESCRIPCION DEL ÁREAEl Estado de Nayarit se encuentra situado en la parte Sur de la Sierra Madre Occidental y en la Llanura Costera del Océano Pacífico. Limita al Norte con los Estados de Durango y Sinaloa, al Este y Sur con Jalisco y al Oeste con el Océano Pacífico. Por su extensión ocupa el 23er. lugar en el país, abarcando una superficie de 27,621 km². Se localiza entre los 20º34' y 23°00' de Latitud Norte y los 103º58' y 105º45' de Longitud Oeste y ocupa una superficie de 27,621 Km² (Valencia Huerta, 1980). El área de estudio se restringe a la zona ubicada entre las coordenadas geográficas 21º15'36" y 22º46'00" de Latitud Norte y los 105º15'00" y 106º17'48" de Longitud Oeste, abarcando la mayor parte de la plataforma continental del Estado de Nayarit, la cual en general es estrecha hacia el Norte y Sur de la línea de costa, ensanchándose frente a las Islas Marías (Fig.1) HIDROGRAFÍAEl litoral nayarita ocupa una extensión aproximada de 175 km de perfiles bajos donde se desarrollan complejos sistemas lagunares y esteros, con una superficie del orden de las 92,400 Ha (S.R.H.,1971) en cuya porción Norte se ubican las Lagunas de Teacapán, Agua Brava y Mezcaltitán, así como el estero de Camichín. Las Bahías de San Blas y Jaltemba están al sur de la desembocadura del Río Santiago. La punta Mita cierra la Bahía de Banderas en su extremo norte. Todos los ríos de Nayarit se encuentran en la vertiente del Océano Pacífico. El Acaponeta, el San Pedro Mezquital y el Huaynamota, afluente del Santiago, nacen en el estado de Durango. El Río Teacapán o Cañas se localiza en la porción Norte y una parte de su curso forma el límite estatal con Sinaloa. El río Bejuco desemboca en la Laguna Agua Brava. El Río Grande de Santiago (de la cuenca Lerma-Chapala-Santiago) penetra a Nayarit procedente de Jalisco. En el Sur, el río Ameca que nace en Jalisco forma límite con dicho Estado (Valencia Huerta, 1980). La mayoría de estos ríos drenan en el área grandes volúmenes de agua dulce, aunque se considera que en la actualidad dichos aportes han decrecido debido a las captaciones de agua a lo largo de sus cauces por represas e irrigación de tierras laborables. Por lo tanto, los datos que se enlistan a continuación reportados por Curray, Emmel y Crampton (1969), deben tomarse con reservas.  Rios HIDROLOGÍA
TemperaturaEn la carta de isotermas de fondo elaborada por Roden (1958) y Roden y Groves (1959)In: Parker (1964) se presentan los resultados de observaciones realizadas durante un período de 20 años, señalándose para el área de Nayarit valores medios de temperatura que varían de 14ºC en zonas próximas a la costa, hasta 10ºC al occidente de las Islas Marías; mientras que en las aguas someras de la zona de San Blas se estima entre 22 y 30ºC SalinidadAlvarez-Borrego (1983) reporta una distribución horizontal de la salinidad a 10 m de profundidad que varía de 34.80 %o a 35.20 %o entre los 109° y 106° de Longitud Oeste. Oxígeno disueltoLas concentraciones de este gas a profundidades de 100 a 200 m son alrededor de 0.5 ml/litro; mientras que a profundidades menores son de 3 ml/litro en promedio (Parker, 1964). MareasEl rango medio de marcas en primavera es de 1.25 m en Mazatlán y alrededor de 0.98 m en San Blas, el promedio es de 0.85 m y 0.70 m, respectivamente. Las mareas son mixtas semidiurnas (Curray, Emmel y Crampton, 1969). CorrientesParker (1964), Schwartzlose y Hendrickson (1983) y Alvarez-Borrego (1983) delimitan la entrada al Golfo de California trazando una línea que conecta La Paz con Topolobampo y otra entre Cabo San Lucas y Cabo Corrientes, incluyendo así el área de la plataforma continental en Nayarit. Esta es una zona transicional que tiene una estructura oceanográfica muy complicada y dinámica en la que la influencia del Golfo sobre el Océano Pacífico adyacente es pequeña. El flujo hacia adentro y fuera del Golfo ha sido estimado como 1.19x10 6 m³ S-¹ y 1.17x10 6 m³ s-¹, respectivamente (Roden, 1958; In:Alvarez-Borrego,1983). En la entrada del Golfo de California hay 3 clases de aguas superficiales: agua de la corriente fría de California de baja salinidad (S %<- 34.60), la cual fluye hacia Figura 1. Localización del área de estudio y estaciones de muestreo. el Sur a lo largo de la costa Oeste de Baja California; agua caliente del Pacífico tropical oriental de salinidad intermedia (34.65 <- S %o <- 34.85), la cual fluye en el área desde el Sureste; y agua caliente de alta salinidad (S %o >- 34.90) del Golfo de California (Roden y Groves, 1959; Griffths, 1968; Stevenson, 1970;In:Alvarez-Borrego, 1983). 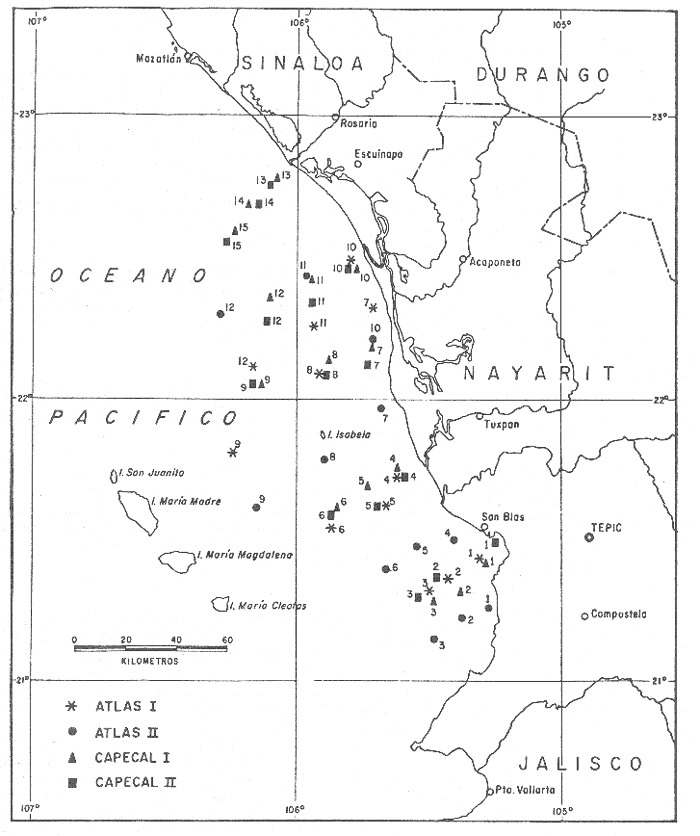 Figura 1. Localización del área de estudio y estaciones de muestreo. Se concluye que la circulación de superficie durante el invierno se caracteriza por corrientes hacia el Sur hasta Cabo Corrientes y en el verano existe un flujo hacia el Norte a lo largo de la costa de México y entra al Golfo (Roden,1958; In: Alvarez-Borrego, 1983). ClimaEl clima de la región varía de subtropical a tropical, de tipo AW o (w) (e) según el sistema climático de Köppen modificado por García (1973), con una temperatura media anual alrededor de los 25ºC El régimen pluviométrico anual va de 850 mm hacia el extremo Norte en Mazatlán, aproximadamente 1,200 mm en Tepic y hasta 1,660 mm en la costa Sur cerca de San Blas. La época de lluvias se inicia hacia el final del verano y principio de otoño, acompañada frecuentemente de tormentas tropicales o chubascos procedentes del Sur. Los vientos dominantes durante los meses de invierno son del Noroeste y los del Oeste a Suroeste en el verano. Hay un régimen de brisa marina por la tarde que decrece después de la puesta del sol. MATERIALES Y MÉTODOSEl estudio es producto de 4 campañas oceanográficas a bordo del B/O "El Puma". Dos de ellas, denominadas ATLAS-I y ATLAS-II, se realizaron del 7 al 21 de febrero y del 15 al 28 de abril de 1982, respectivamente y forman parte del proyecto "Atlas Oceanográfico de los recursos biológicos demersales de la plataforma continental de los Estados de Guerrero y Nayarit". Las otras 2 campañas, conocidas bajo el nombre de CAPECAL-I y CAPECAL-II, se llevaron a cabo del 22 de mayo al 10 de junio de 1983 la primera , y del 18 de septiembre al 6 de octubre de 1984 la segunda, correspondiendo ambas al proyecto "Calamares y Peces del Golfo de California". Para efectos de muestreos biológicos e hidrológicos, se establecieron series de transectos perpendiculares a la costa con 3 estaciones en cada uno, tomando en cuenta las isobatas de los 20, 50 y 85 m como promedio considerando además el tipo de fondo y los caracteres geográficos del litoral para el desarrollo adecuado de las maniobras. Se realizaron un total de 54 estaciones distribuidas de la manera siguiente: 12 en ATLAS-I, 12 en ATLASII, 15 en CAPECAL-I y 15 en CAPECAL-II (fig. 1). Los valores de salinidad, oxígeno disuelto y temperatura del agua en las primeras tres campañas se obtuvieron utilizando botellas muestreadoras tipo Niskin, colocadas a distintos niveles de la profundidad registrada mediante el empleo de una ecosonda. En CAPECAL-II se usó el equipo electrónico denominado CTD) (conductividad, temperatura, profundidad) a fin de obtener los datos mencionados. En cada estación se tomó una muestra de 100 ml de sedimento por medio de una draga Smith McIntyre. El análisis granulométrico se hizo de acuerdo con la clasificación propuesta por Shepard y Moore (1955) y se elaboró un mapa de distribución de sedimentos en el área (Fig. 2), con base en los datos aportados por el Laboratorio de Geología Marina de la "Estación Mazatlán". La recolecta del material biológico se efectuó mediante el empleo de una red camaronera de 80 pies, con una luz de malla de 3.8 cm en el copo, con el fin de capturar la fauna malacológica acompañante del camarón. El tiempo de arrastre fue de 30 min a una velocidad promedio de 2.5 nudos, cubriendo un área de barrido de 27,725.5 m² por estación. Los ejemplares capturados en los arrastres fueron discriminados por clase, preservados en formol al 10% y etiquetados para su traslado al Laboratorio de Malacología del Instituto de Ciencias del Mar y Limnología de la UNAM. En el Laboratorio se lavaron las muestras con agua corriente y se secaron a temperaturas inferiores a 70ºC, separando las formas colectadas vivas de los exoesqueletos para su posterior identificación y cuantificación a nivel específico. En el caso de los bivalvos con conchas desarticuladas se adoptó el criterio de García-Cubas (1963) quien considera cada valva como un organismo. El material identificado quedó incorporado a la colección malacológica del Instituto de Ciencias del Mar y Limnología de la UNAM. 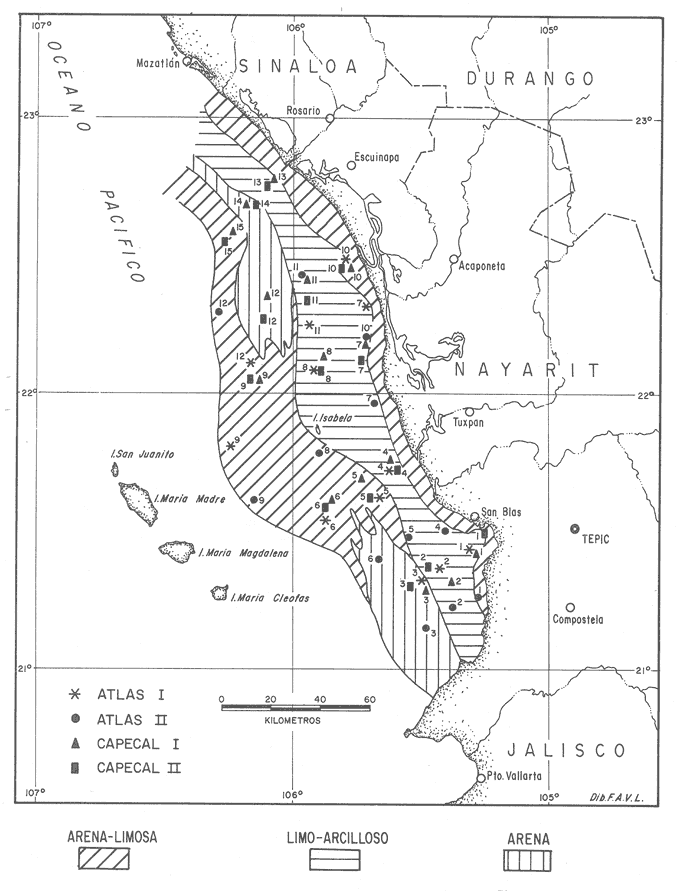 Figura 2. Distribución de las estaciones según el tipo de sedimento. Las comunidades de moluscos están caracterizadas por varios componentes que fueron determinados con base en la abundancia, frecuencia y densidad relativas de las especies, de donde se derivó el valor de importancia relativo. Tales criterios se complementan con anotaciones de distribución geográfica y local, hábitos alimenticios y habitat (Keen, 1971; Andrew, 1971; Pourchon, 1968; García-Cubas, 1981). El índice de diversidad o medida de información que aporta el sistema, fue calculado mediante las expresiones matemáticas propuestas por Margalef (1957) y Shannon-Wiener (1963): H'n = -Σ Pi ln Pi; "nat"/ind. (1) donde: Pi = ni/n (proporción de individuos) ni = número de individuos N = total de individuos en una colecta. La fórmula de Shannon-Wiener fue separada en sus dos componentes, de acuerdo con lo señalado por Lloyd y Ghelardi (1964). Así, el elemento de "equitatividad", que es una medida de la abundancia relativa y representa la proporción de la diversidad máxima en iguales condiciones de abundancia de las especies, se obtuvo utilizando el enunciado de Pielou (1975): J' = H'/Hmax = H'/ln S (2) El índice de "riqueza de especies" se considera como otro componente de la diversidad y es una función del logaritmo de la extensión de la muestra. La expresión matemática que lo define (Margalef, 1957) es la siguiente: D = (S-1) / ln N (3) donde: S = número total de especies en una colecta N = total de individuos de todas las colectas Los índices señalados anteriormente se calcularon sobre la base de los logaritmos naturales y se aplicaron exclusivamente a poblaciones vivas, de acuedo con el tipo de sustrato en el que se encontraron. RESULTADOS
HIDROLOGÍALas mediciones efectuadas sobre temperatura, salinidad y oxígeno disuelto en el área variaron de 13.9 a 30.0ºC, de 33.7 a 34.9 %o y de 0.00 a 4.69 ml/1, respectivamente. Los valores de estos parámetros se detallan en los cuadros de concentración por campaña (Tablas 1-4), incluyendo posicionamiento y profundidad para cada una de las estaciones de muestreo. 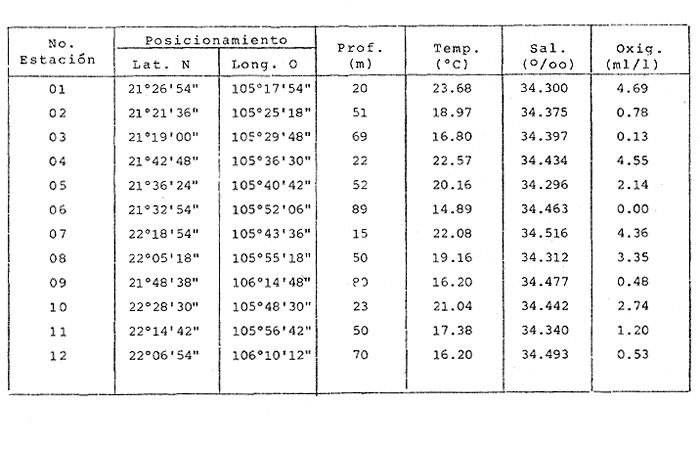 TABLA 1 UBICACIÓN DE ESTACIONES Y PARAMETROS HIDROLÓGICOS. CAMPAÑA ATLAS-I 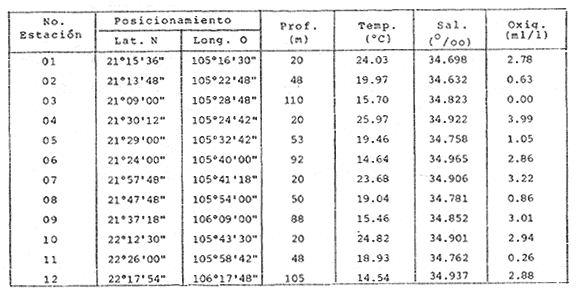 TABLA 2 UBICACIÓN DE ESTACIONES Y PARÁMETROS HIDROLÓGICOS CAMPAÑA ATLAS-II 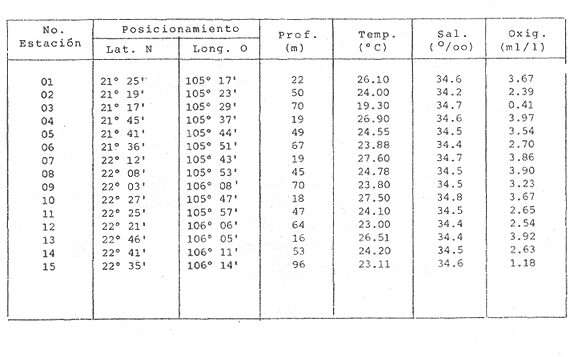 TABLA 3 UBICACIÓN DE ESTACIONES Y PARÁMETROS HIDROLÓGICOS. CAMPAÑA CAPECAL-I 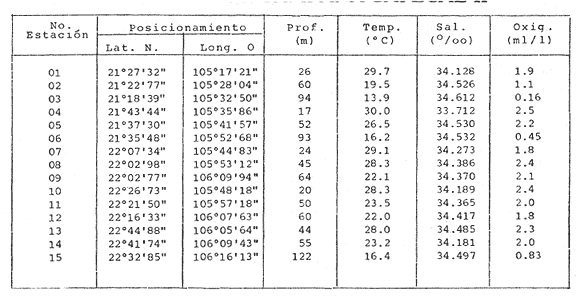 TABLA 4 UBICACIÓN DE ESTACIONES Y PARÁMETROS HIDROLÓGICOS. CAMPAÑA CAPECAL-II SedimentosComo resultado del análisis de sedimentos se observó que las estaciones se encuentran distribuidas de la manera siguente: 28 en sustrato de tipo limo- arcilloso, 17 en arena limosa y 9 en arena (Tabla 5 y Fig. 2). Consideraciones a la sistemáticaLa identificación de la malacofauna se efectuó de acuerdo con Keen (1971) quien adopta para la clase Bivalvia el sistema de clasificación propuesto por Newell (1963) In: Mooreet al. (1969); para la clase Gastropoda, el criterio de Thiele (1935), el cual es seguido por Moore et al. (1960); para Polyplacophora, el de Smith (1961, 1963); para Scaphopoda, el desarrollado por Emerson (1962); y para la clase Cephalopoda, la revisión hecha por Dall (1908) y Robson (1932). En la lista sistemática se adoptó el siguiente formato: 1) El número que aparece a la izquierda entre paréntesis indica la secuencia de las especies identificadas en este estudio. 2) Dado que en este trabajo no se incluyen las ilustraciones correspondientes a cada una de las especies y con el fin de facilitar el acceso a las mismas, se cita a la derecha del nombre científico, enseguida de la inicial K, el número asignado por Keen (1971). 3) La simbología empleada para indicar la distribución en el área de estudio tiene el siguiente significado: AI = ATLAS-I, AII = ATLAS-II, CI = CAPECAL-I, CII = CAPECAL-II. Los números entre paréntesis se refieren a las estaciones de muestreo en las que se presentó la especie. 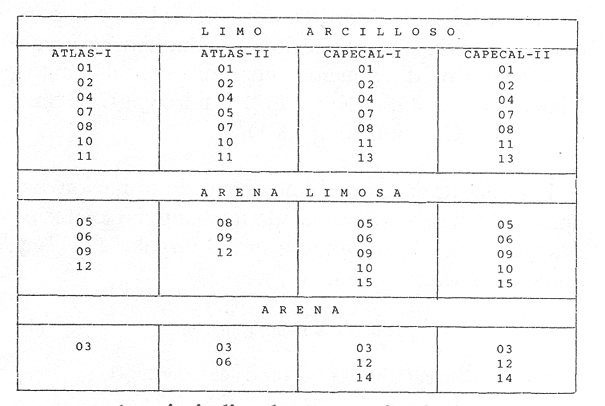 TABLA 5 DISTRIBUCIÓN DE ESTACIONES EN RELACIÓN AL SUSTRATO Los organismos quedaron ubicados sistemáticamente en 5 clases, 61 familias, 112 géneros y 208 especies. 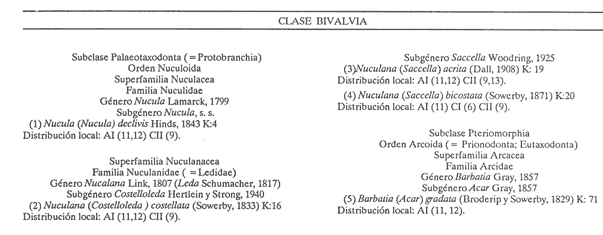 SISTEMETICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA  SISTEMATICA ANÁLISIS CUANTITATIVOEn las cuatro campañas oceanográficas se realizaron 54 estaciones de muestreo, de las cuales en 48 aparecieron moluscos. El total de individuos recolectados fue de 26,600; 3,662 (12.263%) se capturaron vivos y 23,338 (87.736%) correspondieron a conchas. En el ecosistema analizado el mayor número de especies corresponde a la Clase Gastropoda (102 especies), seguida de la Clase Bivalvia (95 especies); mientras que en las Clases Scaphopoda, Polyplacophora y Cephalopoda se presentaron solamente 6, 4 y 1 especies respectivamente. De las 208 especies identificadas en las cuatro campañas, 83 (39.9%) tuvieron representantes vivos. Tales formas se encontraron en tres de las cinco Clases citadas: Gastropoda 55 especies (66.265%), Bivalvia 24 especies (28.915%) y Polyplacophora 4 especies (4.819%). Por lo que se refiere a abundancia, el orden se invierte, ya que la Clase Bivalvia representó el 79.259% (21,083 ejemplares) del total de organismos colectados, Gastropoda el 19.684% (5,236 ejemplares) y en las 3 clases restantes el valor porcentual resultó inferior a la unidad: Scaphopoda 0.695% (185 ejemplares), Cephalopoda 0.195% (52 ejemplares) y Polyplacophora 0.165% (44 ejemplares). Cabe señalar, sin embargo, que el porcentaje de gasterópodos capturados vivos se eleva a un 86.143% (2,810 individuos) sobre 12.507% (408 individuos) de bivalvos y 1.348% (44 individuos) de poliplacóforos. Los resultados respecto a densidad absoluta expresada en número de individuos por unidad de área (DA ind/Km²) , abundancia (AR), frecuencia (FR), densidad (DR) y valor de importancia relativos (VIR), así como número de individuos (No. Ind.) por especie se encuentran concentrados en la figura 3, separando los valores para población total de los que se refieren a población viva. El análisis cuantitativo de la comunidad total vierte como resultado que el área de estudio se encuentra caracterizada por 19 especies: 15 bivalvos (Glycymeris canoa, Argopecten circularis, anadarareinharti, Pitar mexicanus, Pegophysema edentuloides, Trachycardium belcheri, Chione amathusia, Chione kellettii, Lucina fenestrata, Semele laevis, anadara formosa, Mulinia coloradoensis, Cardita spurca beebei, anadara mazatlanica, Plicatula spondylopsis) y 4 gasterópodos(Solenosteira gatesi, Calyptraea mamillaris, Bursa nana y Distorsio decussata) que en conjunto representan alrededor del 70% respecto al total de individuos recolectados en los cuatro cruceros (Figura. 4). No obstante, la comunidad viva que tipifica el área está integrada por 6 especies de gasterópodos(Solenosteira gatesi, Bursa nana, Polinesis uber, Distorsio decussata Fusinus dupetitthoaursi, Conus virgatus) y 2 de bivalvos(Noetia delgada y plicatula spondylopsis ). Es conveniente señalar, sin embargo, que las especies de los géneros Calyptrea, Crucibulum y Chaetopleura, con valores altos de importancia relativa, se excluyen como representativas de la comunidad viva en fondos suaves, en virtud de que se trata de formas epifaunales de sustrato duro -incluyendo conchas de otros moluscos-, las cuales eventualmente se desprenden del mismo en el momento de la colecta. En las figuras 5-8 se muestra de una manera gráfica la relación entre índices de diversidad y sus componentes de equitatividad y riqueza de especies para cada una de las estaciones de muestreo en las que se registraron poblaciones vivas, agrupando las estaciones por campaña según el tipo de sustrato en el que se ubican y eliminando aquellas en las que estos índices tuvieron valores de cero porque los organismos recolectados vivos pertenecían a una misma especie.  Figura 4. Especies características del área de estudio: 1) Glycymeris canoa, 2) argopecten circularis, 3) Andara reinharti, 4) Pitar mexicanus, 5) Pegophysema edentuloides, 6) Trachycardium belcheri, 7) Chione Amadiusia, 8) Chione kellettii, 9) Lucina fenestrata, 10) Semele laevis, 11) Anadara formosa, 12) Mulinia coloradoensis, 13) Cardita spurca beebei, 14) Anadara mazadanica, 15) Plicatula spondylopsis, 16) Solenosteira gatesi, 17) calyptraea mamillaris, 18) Bursa nana, 19) Distorsio decussata. Se observó que los valores más altos de diversidad coinciden, en general, con estaciones localizadas en la región noroccidental de la zona de estudio, en sedimentos de tipo limo-arcilloso para los cruceros ATLAS y en arena-limosa para CAPECAL, presentándose los siguientes rangos por campaña: ATLAS-I, de 0.525 a 1.940 nats/ind; ATLAS-II, de 0.376 a 2.086 nats/ind; CAPECAL- I, de 0 a 1.883 nats/ind y CAPECAL-II, de 0 a 2.305 nats/ind. Así, la mínima diversidad se detectó en las estaciones 01, 07 de CAPECAL-I y 02 de CAPECAL-II y la máxima en la estación 09 de CAPECAL-II, ubicadas en sustrato, limo-arcilloso las primeras y en arena-limosa la última. 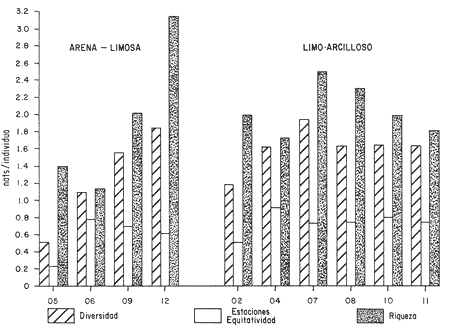 Figura 5. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas en ATLAS-I, según el tipo de sustrato. 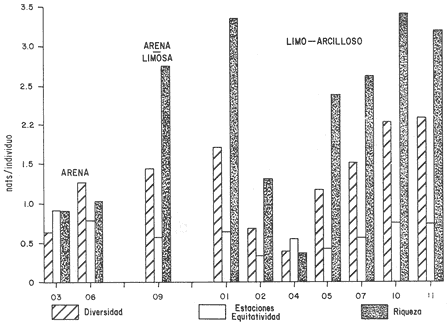 Figura 6. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas en ATLAS-II, según el tipo de sustrato. 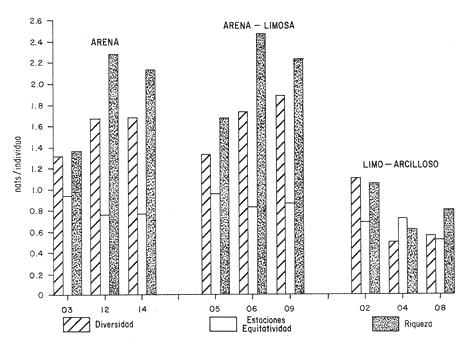 Figura 7. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas en CAPECAL-I. según el tipo de sustrato. 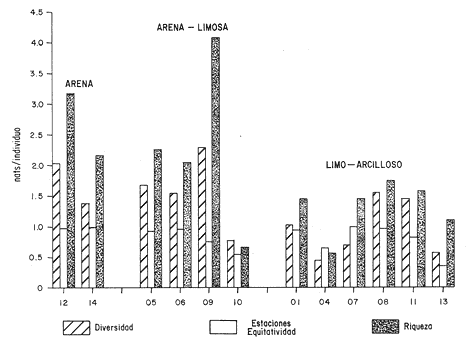 Figura 8. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas en CAPECAL-II, según el tipo de sustrato. Este comportamiento no se mantiene al conjuntar los 4 cruceros como una sola colecta y considerar exclusivamente las variaciones según el tipo de sedimento (Fig. 9), ya que la diversidad es similar en los tres ambientes (arena H'n = 2.53; arena-limosa H'n = 2.35; limo-arcilloso H'n = 2.34). Sin embargo, los valores más altos para los índices de diversidad y equitatividad se registraron en sustrato arenoso, aún cuando la riqueza específica fue mayor en sedimento limo-arcilloso. 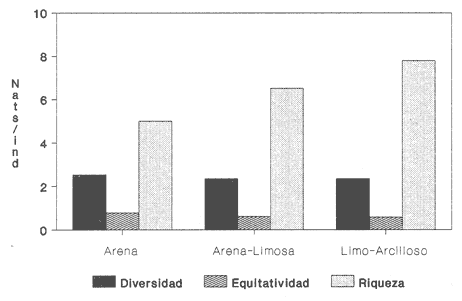 Figura 9. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas, según el tipo de sustrato. Por otra parte, el análisis espacio-temporal (Fig. 10) muestra diferencias en la diversidad por campaña, observándose el siguiente orden decreciente: CAPECAL-II (2.508 nats/ind ), ATLAS-II ( 2.279 nats/ind ), CAPECAL-I ( 2.219 nats/ind ) y ATLAS-I ( 2.174 nats/ind). NUTRICIÓN Y HABITATDe acuerdo con la información reportada en la bibliografía (Purchon, 1968; Andrews, 1971; García~Cubas, 1981), las Familias de moluscos presentan relaciones diversas entre tipos de nutrición y habitat. Tales diferencias se consignan en las Figs. 11-17, de donde se infiere que la forma de nutrición más frecuente en los Bivalvos corresponde a la filtración de partículas orgánicas en suspensión, presente en 17 de las 26 Familias identificadas (65.38%), seguida de la recolección de materia orgánica depositada en la que se ubican 8 Familias (30.76%) y, por último, la carroñera con una Familia (3.84%). Referente a su relación con el sustrato, 17 Familias (65.38%) son infaunales, 7 epifaunales (26.92%) y 2 seminfaunales (7.69%) aunque algunas pueden presentar diversas alternativas alimenticias y de habitat. En los Gasterópodos el tipo de nutrición dominante es el carnívoro, registrado en 19 de las 30 Familias identificadas (63.33%), le siguen en orden de importancia las formas carnívoro-carroñeras con 5 Familias (16.66%), los filtradores de partículas orgánicas en suspensión con 3 Familias (10%), los que se alimentan de materia orgánica depositada con 2 Familias (6.66%) y una Familia herbívora (3.33%). En relación al habitat, 24 Familias resultaron epifaunales (80%), 3 seminfaunales (10%) y 3 infaunales (10%). Entre los Gasterópodos también se encuentran Familias que pueden ubicarse alternadamente en los distintos tipos tanto de nutrición como de habitat. Todos los Poliplacóforos son ramoneadores microherbívoros y epifaunales. Los Escafópodos, por su parte, son seminfaunales y se nutren de materia orgánica depositada. Finalmente, la Familia de Cefalópodos identificada es de hábitos carnívoros y pelágica. 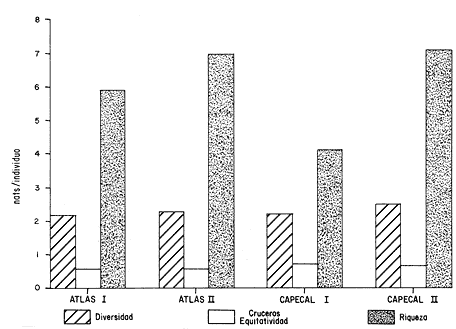 Figura 10. Indices de diversidad (nats/ind), equitatividad y riqueza de especies para poblaciones vivas, por campaña oceanográfica. DISCUSIÓN Y CONCLUSIONESLos resultados obtenidos de temperatura del agua concuerdan con los reportados por Roden (1958) y Roden y Groves (1959)In:Parker (1964) para el área de estudio, presentándose una gradación en relación inversa a la profundidad. Esta similitud en los datos podría indicar que en los últimos años no ha habido ninguna alteración significativa de las isotermas de fondo. La estructura halina vertical del área de estudio es extremadamente compleja, debido a la mezcla de las tres masas de agua que confluyen en la entrada del Golfo de California. Los resultados obtenidos en este trabajo reflejan un comportamiento similar al registrado por Roden y Emilsson (en prensa). En la capa superior (20 m) se registra una salinidad de 34.5 %o como promedio y el acarreo de agua de alta salinidad procedente del Golfo, hacia la entrada del mismo, es mayor en el lado oeste, donde la advección es más pronunciada y la salinidad aumenta. Entre los 44 y 60 m de profundidad hay una capa de salinidad mínima (34.4 ‰ que se origina por la mezcla de agua de la corriente de California y la del mismo Golfo. 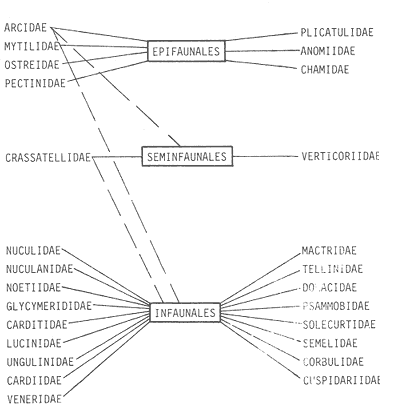 Figura 11. Habitat de las Familias de Bivalvos identificadas. 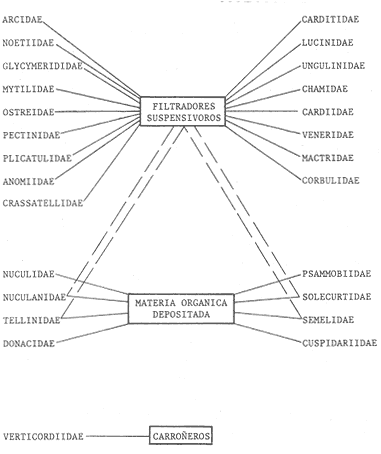 Figura 12. Tipos de nutrición en las Familias de Bivalvos identificadas. 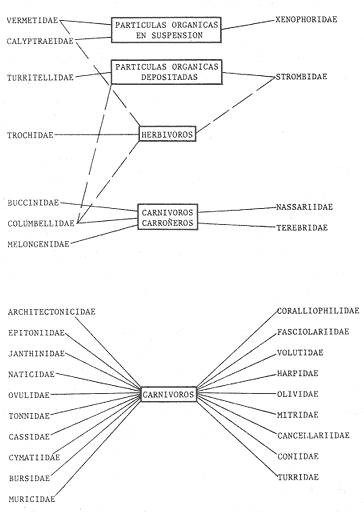 Figura 13. Tipos de nutrición en las Familias de Gasterópodos identificadas. 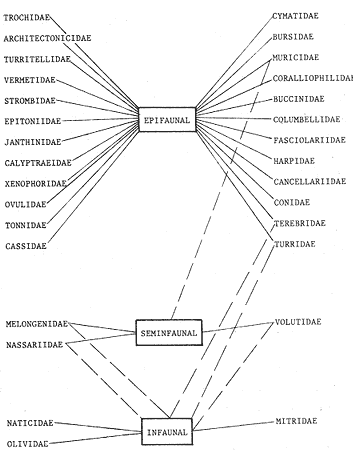 Figura 14. Habitat de las Familias de Gasterópodos identificadas.  Figura 15. Tipo de nutrición, y habitat de las Familias de Poliplacóforos identificadas.  Figura 16. Tipo de nutrición, y habitat de las Familias de Escafópodos identificadas.  Figura 17. Tipo de nutrición, y habitat de las Familias de Cefalópodos identificadas. En el nivel subsuperficial, que se extiende entre los 64 y 122 m de profundidad, hay una capa con valores promedio de salinidad de 34.6 ‰ que, según Roden y Emilsson (en prensa) es característica del Pacífico Oriental Tropical e indica un flujo de agua hacia el norte que puede confundirse en la parte occidental del Golfo de California con la capa superior de alta salinidad. Este comportamiento halino es atribuido a los frentes de Cabo San Lucas y Cabo Corrientes, cuya intensidad varía con la estación del año y es mayor al final de la primavera (Roden y Emilsson, en prensa), tal como se observa en los datos registrados durante la Campaña ATLAS-II, efectuada durante el mes de abril. La relación entre oxígeno disuelto y profundidad se da en general de manera inversa. Así, la concentración del gas disminuye en tanto que la profundidad aumenta, coincidiendo las zonas anóxicas y de concentración mínima de oxígeno con las mencionadas por Parker (1964) para la plataforma continental del Estado de Nayarit. La predominancia específica de la Clase Gastropoda puede explicarse en función del arte de pesca ya que los organismos de esta clase son en su mayoría epifaunales, mientras que un gran número de bivalvos son semi-infaunales o infaunales. Entonces, la probabilidad de ser capturados por la red de arrastre es mayor para los gasterópodos que para los bivalvos. Por otra parte, el hecho de que los gasterópodos sean principalmente de hábitos carnívoros, los ubica en una posición ventajosa respecto a las otras clases. Aspecto que se manifiesta en la gran diferencia porcentual de individuos vivos en la colecta. La abundancia numérica de los bivalvos puede ser atribuida, en cambio, al criterio adoptado para su cuantificación, pues ante la imposibilidad de articular valvas aisladas, cada una fue considerada como un ejemplar, elevándose por tanto su proporción real. Por lo que se refiere a la distribución de las especies, señalada por Keen (1971), Dushane y Poorman (1967), Dushane y Sphon (1968) y Lesser (1984), se amplía el rango Sur para 6 especies de Gasterópodos(Solenosteira gatesi, Trajana perideris, Mitrella dorma, Fusinus (Aptyxis) cinereus, Kylix Paziana y Drillia (Drillia) cunninghamae),3 de Bivalvos (Mulinia coloradoensis, Tellina (Laciolina) ochracea y Semele guaymasensis) y tres de Poliplacóforos(Chaetopleura euryplax, Ch. mixta y Radsiella sp). El rango de distribución Norte se modifica para 2 especies de Gasterópodos(Murex (Murex) recurvirostris recurvirostris y Subcancilla phorminx), 4 de Bivalvos (Pitar (Lamelliconcha) callicomatus, P. (L.)unicolor, Macoma (Psammacoma) siliqua siliqua y Semele elliptica) y una especie de Escafópodo(Tesseracme tesseragonum). Se encontraron también nuevos registros para aguas nacionales, 3 de ellos corresponden a Gasterópodos(Turritella cingulata, Strombina (?Cotonopsis) turrita y Subcancilla gigantea) y 6 Bivalvos (Nuculana (Saccella) bicostata, Anadara (Anadara) adamsi, Pitar (Hyphantosoma) hertleini, Tellina (Eurytellina) ebumea ebumea, T. (Merisca) brevirostris y T. (Phyllodella) insculpta). La mayor riqueza de poblaciones vivas en sustrato de tipo limo-arcilloso puede deberse a la propia distribución de los sedimentos en el área, lo cual ocasionó que allí quedara concentrado el mayor número de estaciones de muestreo; o bien a que este tipo de sedimentos constituyen verdaderas trampas de detritus, lo que hace posible soportar poblaciones que se nutren de ellos. En el análisis espacio-temporal se observó una diferencia mínima en cuanto a índices de diversidad, presentándose el valor más alto en la campaña CAPECAL-II y el menor en ATLAS-I. Aspecto que puede ser explicado en términos de la variación de la productividad y, por tanto, de disponibilidad de alimento y factores ambientales según las épocas en las que se realizaron los muestreos que corresponden a verano-otoño e invierno, respectivamente, pues, de acuerdo con Zeitschel (1969), a finales de agosto suele producirse una mezcla de las aguas superficiales y las profundas y a principios del otoño tiene lugar una restauración de la fertilidad del fondo al descender los productos de la descomposición de animales y vegetales que murieron al finalizar el verano. En algunos estudios, como el de Parker (1964), se hace un análisis de las comunidades faunísticas en función de la profundidad en que habitan. En el presente trabajo se caracterizó cada uno de los niveles, tomando en cuenta sólo a la comunidad viva integrada por especies con un valor de importancia relativa superior a la unidad. Así, en el estrato somero (15-26 m) se encontraron 5 especies exclusivas del mismo: 1 bivalvo (Lunarca brevifrons) y 4 gasterópodos (Melongena patula, Cancellaria decussata, Cancellaria balboae y Conus perplexus). La población viva del nivel medio (44-60 m) estuvo representada por una sola especie de la Clase Bivalvia (Chione kellettii). En el estrato profundo (64-122 m) se registraron 2 especies: un bivalvo (Barbatia gradata)y un gasterópodo (Polystira nobilis). Además, se encontraron especies comunes a los niveles somero y medio (Argopecten circularis, Pitar mexicanus, Nassarius catallus, Conus recurvus, Conus virgatus, Polystira picta); al medio y profundo(Plicatula spondylopsis, Cardita spurca beebei, Chama mexicana, Architectonica nobilis, Murex recurvirostris recurvirostris, Murex recuvirostris tricoronis, Hexaplex brassica, Fusinus dupetitthouarsi, Kylix paziana, Polystira oxytropis, Knefastia tuberculifera), y otras que están presentes en los tres estratos de profundidad(Noetía delgada, Polinices uber, Distorsio decussata, Bursa nana, Solenosteira gatesi). Conviene señalar, sin embargo, que la malacofauna registrada como muerta en el momento de la colecta, se encontró distribuida en los 3 niveles muestreados indiscriminadamente y que los patrones de abundancia y distribución de las 19 especies dominantes de la comunidad total (vivos y muertos) indican una tendencia a concentrarse en la región noroccidental de la zona de estudio, a profundidades que van de los 44 a los 66 m, lo cual puede estar relacionado con el movimiento de las masas de agua en la entrada del Golfo de California. Las especies de bivalvos que actualmente son capturadas, ya sea en la pesca de subsistencia o en la industrializada, pertenecen fundamentalmente a los génerosAnadara, Argopecten, Pitar, Dosinia y Chione.Entre los gasterópodos, las especies de interés comercial, tanto por su concha como por su carne, se ubican principalmente en los géneros Strombus, Hexaplex y Murex (Baqueiro et al., 1982). Finalmente, en la fauna característica del área se encuentran especies de moluscos que podrían ser utilizadas como un recurso alimenticio y aunque algunos no alcanzan la talla comercial óptima, su aprovechamiento se justificaría por su abundancia específica. Tal es el caso de Solenosteira gatesi y Bursa nana, las cuales podrían constituir un recurso alternativo en el futuro.
Cuadro 1 AgradecimientosLos autores manifiestan su reconocimiento a Mario Gutiérrez E. y Alberto Castro del Río, por la aportación del análisis sedimentológico. Asímismo, agradecen a Esteban Monroy S., Fernando Vega López y Xavier Chiappa Carrara la elaboración del material gráfico. LITERATURAÁLVAREZ-BORREGO, S., Gulf of California. Estuaries and enclosed seas, 26 Chapter 17. Elsevier Scientific Publishing Co., In: Ketchum, H.B. (Eds.), New York:1983.427-449. ANDREWS, J., Sea shells of the Texas coast.University of Texas Press, Austin,1971. 298 p. BAQUEIRO, E., J.A. MASSO y H. GUAJARDO, Distribución y abundancia de moluscos de importancia comercial en Baja California Sur.Instituto Nacional de la Pesca, México,1982. 62 p. CURRAY, J.R.,F.J. EMMEL y P.J.S. CRAMPTON, , Lagunas Costeras, un simposio (Coastal Lagoons, a Symposium).Mem. Simp. Inter. Lagunas Costeras Holocene history of a strand plain, lagoonal coast, Nayarit, Mexico. In: Ayala-Castañares, A. y F.B. Phleger (Eds.) UNAM-UNESCO, México . UCSI (S) - UNAM (MAR). 1969.63-100 p. 28-30 noviembre 1967. DALL, W.H., The mollusca and Brachiopoda.Reports on the dredging operations off the west coast of Central America to the Galapagos, to the west coast of Mexico, and in the Gulf of California... XIV. Bull. Mus. Comp. Zool., Harvard, 1908. 205-487, pls. 1-22. 43 (6): DUSHANE,H. y R. POORMAN, A checklist of mollusks for Guaymas, Sonora, Mexico. Veliger, 1967.413-41.9(4): DUSHANE,H. y G.G.SPHON, A checklist of intertidal mollusks for Bahia Willard and the southwestern portion of Bahia San Luis Gonzaga State of Baja California, Mexico. Veliger, 1968.233-46.10(3): EMERSON, W.K.,Jour.Paleont., A classification of the Scaphopod mollusks.1962. 461-82, pls, 76-80,2 text figs36(3): GARCÍA-CUBAS, A.,Sistemática y distribución de los micromoluscos de la Laguna de Términos, Campeche, México. Bol Inst. Geol. Univ. Nal Autón. México, 1963. 1-55,24 figs., 4 láms. 67(4): GARCÍA-CUBAS, A.,Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de Términos, Campeche). Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México,Publ. Esp.1981.1-182.5: GARCÍA E., Modificaciones al sistema de Clasificación Climatica de Kóppen Inst. Geogr. Univ. Nal. Autón. México,1973 246 p. HENDRICKX, M. E.,Recursos Pesqueros Potenciales de México: La Pesca Acompañante del Camarón. Diversidad de los Macroinvertebrados Bentónicos acompañantes del Camarón en el Area del Golfo de California y su Importancia como Recurso Potencial, In: Yañez- Arancibia, A. (Ed.) Progr. Univ. de Alimentos, Inst. Cienc. del Mar y Limnol., Inst. Nal. de Pesca. UNAM, D.F.,México,1985. 95-148.748 p. Cap. 3: HERTLEIN,L.G. y A.M. STRONG, ,Moolusks from the west coast of Mexico and Central America. Eastern Pacific expeditions of the New York Zoological Society.Pts. I-X. ZoologicalNew York.1940-51. KEEN. M.A., Sea Shells of Tropical West America.Marine Mollusks from Baja California to Peru.Stanford University Press,2a. ed.,Stanford,1971.1065 p., 22 láms. KEEN. M.A., y E. COAN, Marine Molluscan Genera of Western North America. An Illustrated Key. Stanford University Press, 2a. ed.,Stanford,1974. 208 p. LESSER, H., Tesis Profesional. Prospección sistemática y ecológica de los moluscos bentónicos de la plataforma continental del Estado de Guerrero, México,Fac. de Ciencias, Univ. Nal. Autón. México,1984.107 p. LLOYD, M. y RJ. GHELARDI,J. Anim. Ecol., At table for calculating the "Equitability" component of species diversity.1964.217-227.33: MARGALEF,D.R. La teoría de la información en ecología.Mem. R Acad Cienc. Artes.,Barcelona1958.79 p. 32(13) MARGALEF,D.R.Ecología. Ed. Omega,Barcelona,1974. 951 p. MOORE, R.C. (Ed.), Treatise on Invertebrate Paleontology. Part I. Mollusca 1 (J. Brookes Knight et al.) (Mollusca, General features: Scaphopoda; Amphineura; Monoplacophora; Archaegastropoda).1960. 351 p., 216 text figs. MOORE, R.C. (Ed.),Treatise on Invertebrates Paleontology. Part N: Bivalvia. 3 vols.1969.952 p., ill. Vols. 1-2, PARKER, R.H., Zoogeography and ecology of some macro-invertebrates, particulary mollusks, in the Gulf of California and the continental slope of Mexico. Vidensk. Medd. fra. Dansk naturh. Foren. Bd. 1964.126, 178 p., pls. i-xv, 29 figs PIELOU, E.C., Ecological diversity. Ed. Wiley New York,1975. , 165 P PURCHON, R.D., The Biology of the Mollusca. Pergamon Press, New York 1968., 560p. ROBSON, G.C., A monograph of the Recent Cephalopoda, based on the collections in the British Museum (Natural History). Part I. Octopodinae, London:Part II: Octopoda, excluding Octopodinae. Ibid, British Museum Publ.,1929-32.236.p.,pls. i-vii (1929);359 p., 6 pls. text. figs.(Enero 1932). RODEN, G. I. y I. EMILSSON, en prensa. Simposio "El Golfo de California" (Simposium "The Gulf of California"). Oceanografía física del Golfo de California (Physical oceanography of the Gulf of California ). In: Ayala-Castañares, A., F. B. Phleger, R. Schwartzlose y A. Laguarda (Eds). Univ. Nal. Autón. México. (En español e Inglés). SCHWARTZLOSE, R. A. y J. P, HENDRICKSON, Bibliografía del Golfo de California: Ciencias Marinas (hasta el final de 1981). Inst. Cienc. del Mar y Linmol. Univ. Nal. Autón. México,Publ., Esp.1983.1-212.7: SECRETARÍA DE RECURSOS HIDRÁULICOS (Ed.), Plan Nacional de desarrollo pesquero en lagunas litorales. SRH, México,1971.12p. SHANNON, E. C. y N. WIENER, , The Mathematical Theory of Communication. Urbana, University of Illinois Press1963,117 p. SHEPARD, F. P. y D. G. MOORE, Bull. American Assoc. Petrol. Geol Central Coast Sedimentation: Characteristics of sedimentary environment, Recent History and Diagenesis.1955.1463-1593,75 figs39 (8): SMITH, A. G., Proc. California Acad Sci., Four species of chitons from the Panamic province.ser. 4,1961. 81-90. pls. 8-9.30 (4): SMITH, A. G., A revised list of chitons from Guadalupe Island, Mexico (Mollusca: Polyplacophora). Veliger,1963.147-49.5(4): THIELE, J., Handbuch der Systematischen Weichtierkunde Jena,1934-35.779-1154, figs. 784-789.2: VALENCIA HUERTA, A.,Monografía del Estado de Nayarit.SARH, México,1980.62 p. ZEITSCHEL, B., Primary productivity in the Gulf of California, Mar. Biol.,1969201-207.3 (3):
|