|
CARACTERÍSTICAS MORFO-FUNCIONALES DE Chirostoma Consocium,
Ch.Chapalae Y Ch. Ocotlanae DEL LAGO DE CHAPALA, MÉXICO (PISCES.A
THERINIDAE)
Trabajo recibido el 8 de octubre de 1986 y aceptado para su
publicación el 5 de abril de 1988.
A. Rodríguez Ruíz
C.Granado Lorencio
Departamento
de Ecología. Universidad de Sevilla. Apartado 1095,41080 Sevilla,
España
En el presente trabajo. se realiza el
estudio morfo-funcional. de tres especies del Genero
Chirostoma. coexistentes en el Lago de Chapala. Se estudiaron 179 ejemplares,
correspondientes aCh.consocium (61), Ch.
chapalae (66) yCh. ocotlanae (52), sobre las
cuales se efectuaron un total de 14 variables biométricas y 3 variables
merísticas de marcado carácter funcional. A la matriz de datos,
se aplicaron tratamientos matemáticos descriptivos (X y SD) y
multivariantes (APC). Los resultados obtenidos muestran la existencia de
diferencias morfológicas interespecíficas, que confieren a cada
especie adaptaciones exclusivas en la explotación de los recursos del
ecosistema, permitiendo asi su coexistencia.
Chirostoma,Chapala,Variables morfofuncionales, coexistencia
de especies.
Palabras claves:Chirostoma,
Chapala, variable morfo-funcionales, coexistencia de especies
In
this paper, a morpho-functional study of three species of Genus
Chirostoma that coexist at Chapala Lake (Mexico) is given.
We studied 14 biometric variables and 3 meristic variables, of on enhaced
functional character, of 179 specimens of Ch. consocium
(61), Ch. chapalae (66) and Ch. ocotlanae
(52). Descriptive mathematical analysis (X and SD) and multivariable analysis
(PCA) were applied to the data matrix. The results demonstrate the existence of
interespecific morphological differences which allow the unique adaptation of
each species to exploit ecosystem resources, thus permiting their coexistence.
Chirostoina, Chapala, morpho-functional variable, species
coexistence.
Key words:Chirostoma,
Chapala, morpho-functional variable, species coexistence.
La coexistencia de especies congenéricas en ecosistemas acuáticos estables ha constituido, desde hace muchos años, un atrayente objeto de estudio. Muchos autores han puesto de manifiesto las divergencias anatómicas (Matthes, 1963; Keast y Webbs, 1966; Alexander, 1967; Gatz, 1979, 1981; Liem, 1980; Adamicka, 1983), tróficas (Etnier, 1971; Liem y Osse, 1975; Werner y Hall, 1977; Werner, 1979; De Silva et al, 1980; Gladfelter y Jhonson, 1983; Bengston, 1984) y de segregación espacial (De Martini, 1969; Zaret y Rand, 1971; Moyle, 1973; Greenwood, 1974; Werner, 1977; Van Ojen, 1982; Brabrand, 1984) que intervienen en el mantenimiento de estas asociaciones faunísticas, a veces altamente diversificadas (caso de los cíclidos en los lagos africanos), fruto de una radiación adaptativa de las especies, tendente a evitar posibles fenómenos competitivos en ambientes con factores limitantes (Principio de exclusion competitiva, Gause, 1934). Por ello, la detección de diferencias interespecíficas aporta un esclarecimiento en la comprensión de la historia evolutiva de las comunidades animales. El Lago de Chapala constituye un claro ejemplo de lo expuesto anteriormente, al presentar una ictiofauna caracterizada por familias endémicas, en las cuales, algunos de sus géneros, están representados por más de diez especies (Atherinidae, Goodidae y Cyprinidae) (Barbour,1973;Arregui, 1979). Esta masa de agua, situada en la zona central de México, ocupa el primer lugar, por extensión, del país, con una superficie de 1109 km 2 (Ziesler y Ardizzone, 1979). Su origen se remonta al Pleistoceno medio, como consecuencia de la actividad volcánica y los movimientos tectónicos acontecidos durante dicho periodo (elevación y dislocación), dando lugar a una configuración de pequeños lagos repartidos irregularmente por la cuenca (Fig.1) (Barbour, 1973). Se considera como un lago tropical, de circulación holomíctica (Hutchinson, 1957), sin estratificación marcada, somero (profundidad media, 7.4 m) y de escasa transparencia. El trabajo se ha basado en un exahustivo análisis de variables biométricas con marcado carácter funcional, en tres especies del GéneroChirostoma (Ch. consocium, Ch. Chapalae y Ch. ocot1anae), único género de la Familia Atherinidae que se encuentra en el Lago. La elección de dichas variables se realizó con el fin de establecer el papel asumido por cada especie dentro de la compleja red de interacciones que presenta la comunidad íctica del Lago de Chapala. 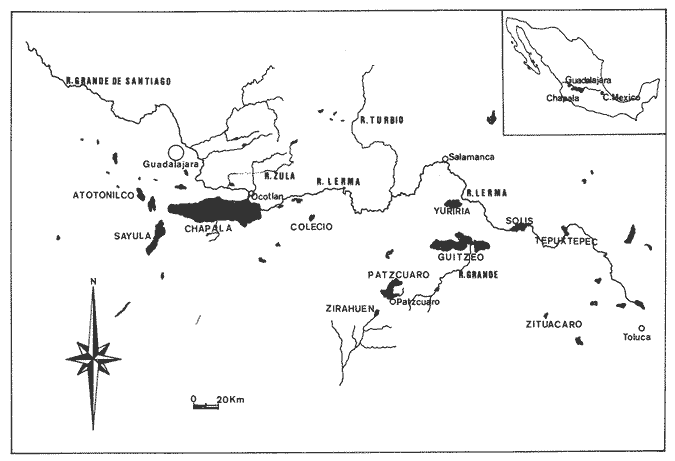 Figura 1. Localización del área de estudio. O Población. MATERIAL Y MÉTODOSLos ejemplares de estudio se obtuvieron a partir de las pescas comerciales realizadas por la población ribereña del lago, mediante capturas con chinchorro. Fueron seleccionados un total de 179 ejemplares correspondientes a: 61 Ch.consocium, 66 Ch. chapalae y 52 Ch. ocotlanae. Sobre cada uno de los individuos, se realizó una serie de medidas en el cuerpo (Tabla 1 y Figura 2) con calibre de precisión (1 mm). Los caracteres morfométricos contínuos, en peces, se incrementan con el tamaño. Este crecimiento de las variables, es distinto en cada una de las especies, estando influenciados por condiciones extrínsecas (Kux y Libosvársky, 1981). Por esta razón, se establecieron una serie de índices biométricos (Tabla 2). A la matriz de los índices biométricos, se le aplicó tratamientos matemáticos tanto descriptivos como discriminantes. Las medias para cada uno de los parámetros obtenidos, fueron comparados mediante el estadístico Student (Sokal y Roffif, 1979), aplicándole el programa BMDP3D (Dixon, 1981). Se utilizó también, análisis de componentes principales (APC), mediante el programa BMDP4M (Dixon, 1975). Este tratamiento, es utilizado por numerosos autores, para el estudio biométrico de las especies (Girona, 1981; Libosvársky et al, 1981; Guillén, 1982; Libosvársky, 1982; Libosvársky y Kux, 1982; Castelló, 1983; Granado, 1983; y otros), ya que el uso de técnicas de análisis multivariantes, tiene como objeto hallar las relaciones que ligan unos caracteres que determinan la variabilidad morfológica de cada especie y su posible relación con el medio. 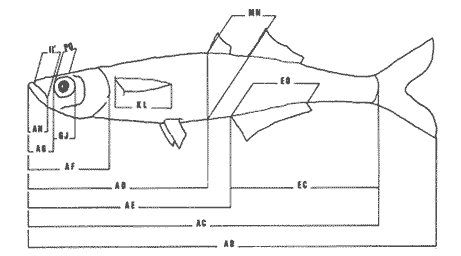 Figura 2. Medidas biométricas realizadas en cada ejemplar.  TABLA 1 VARIABLES EFECTUADOS EN CADA UNO DE LOS INDIVIDUOS 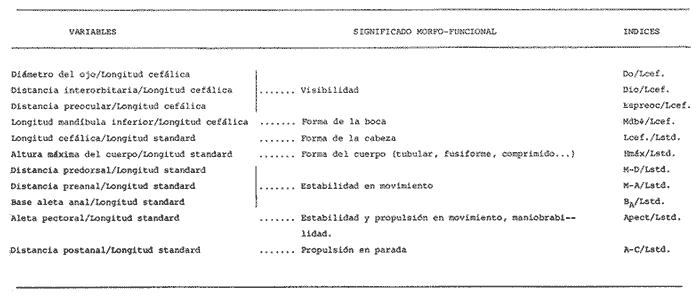 TABLA 2 RELACIÓN DE LOS NDICES UTILIZADOS Y SIGNIFICADO MORFO-FUNCIONAL RESULTADOSEn la Figura 3, se han representado las medias e intervalos de confianza (95%) para los índices de la región cefálica. Ch. consocium y Ch. ocotlanae, si bien muestran una gran semejanza, el diámetro de las órbitas oculares, en la primera, son mayores y más separados entre sí (t-test, P 0.01). Ch. ocotlnae se caracteriza por la asimetría de sus mandíbulas, a diferencia de la simetría mandibular presentada por Ch. consocium. La especie que se separa más nítidamente es Ch. chapalae, que se caracteriza por presentar un diámetro de los ojos menor y más distanciados entre sí (t- test,P 0.01); la posición de éstos, está más retrasada (t-test, P 0.01) y la longitud de su mandíbula inferior es mayor (t-test,P 0.01). 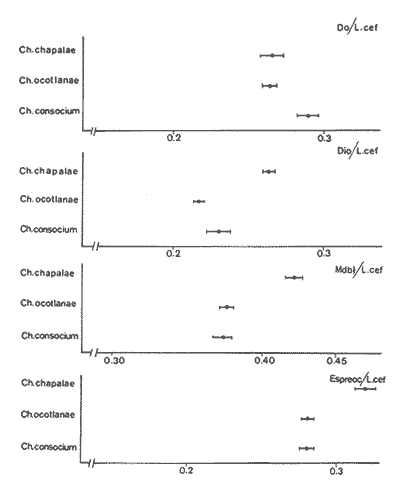 Figura 3. Media e intervalos de confianza (95%) para las variables relacionadas con la región cefálica en Ch. chapalae, Ch. ocotlanae y Ch. consocium. En las figuras 4 y 5, se representan las medias e intervalos de confianza (95%) para los índices relacionados con la forma del cuerpo, capacidad de estabilidad y de movimiento de las especies. Los resultados obtenidos muestran separadas a Ch. chapalae y Ch.consocium, encontrándose Ch. ocotloanae en una situación intermedia. Ch. consocium, se presenta como la especie con una menor longitud de las aletas pectorales (t-test,P 0.01); la ubicación tanto de su primera aleta dorsal como la anal están localizadas más cerca de la región cefálica que en ninguna de las otras especies (t-test,P 0.05 y P 0.01); la longitud del pedúnculo caudal es mayor (t-test,P 0.01) y su cuerpo es menos fusiforme (t- test,P 0.01). Tales resultados evidencian una menor capacidad de maniobra, agilidad y estabilidad en la locomoción para Ch. consocium. En una situación opuesta, se encuentra Ch. chapalae. Esta especie presenta mayor longitud de aletas pectorales (t-test,P 0.01), y por lo tanto una mayor agilidad en las maniobras y estabilidad permanente. La primera aleta dorsal, y la aleta anal se localizan más alejadas de la región cefálica (t~test,P < 0.01), así como posee una gran base la aleta anal (t-test P < 0.01), índices que le confieren un mayor equilibrio cuando los individuos se desplazan en una dirección durante un movimiento brusco. 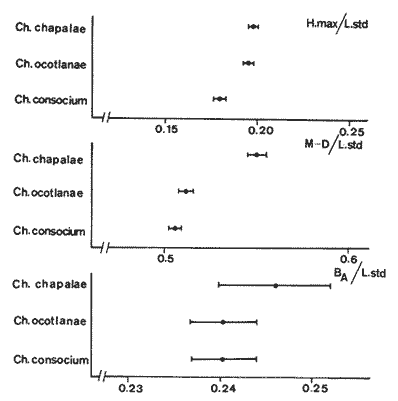 Figura 4. Media e intervalos de confianza (95%) para la forma del cuerpo, así como para las variables que determinan la estabilidad de los individuos en Ch.Chapalae, Ch. ocotlanae y Ch. Consocium. 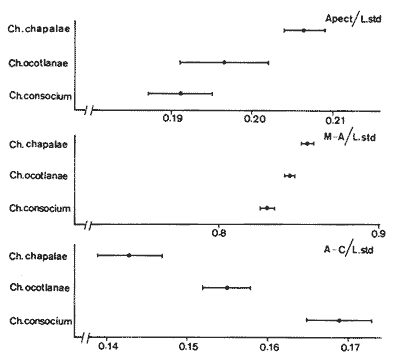 Figura 5. Media e intervalos de confianza (95%) para las variables que determinan la movilidad, maniobra, equilibrio y capacidad de propulsión en Ch. chapalae, Ch. ocotlanae y Ch. Consocium. 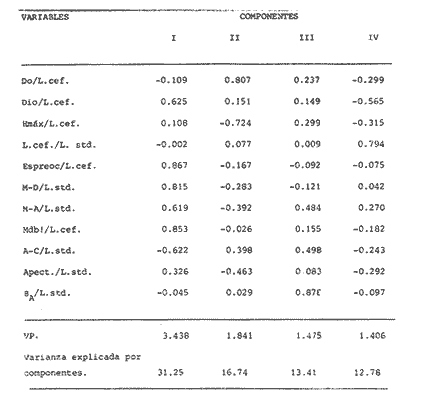 TABLA 3 CARGA DE CADA ÍNDICE BIOMÉTRICO SOBRE LOS 4 COMPONENTES PRINCIPALES CON EIGEN VALORES MAYORES A UNO 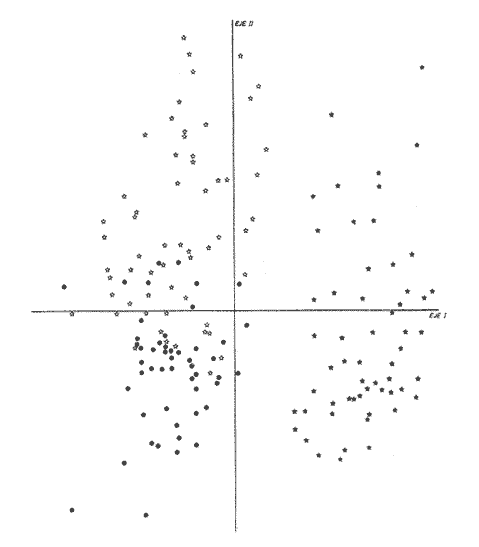 Figura 6. Distribución en el espacio definido por los ejes I y II del análisis de componentes principales sobre los ejemplares de CH chapale (*), Ch ocotlanae (.) y Ch. consocium (*) . (ver texto) Ch. consocium, se presenta como la especie que posee un espacio preocular, mandíbula inferior y distancia interorbitaria menor junto con un mayor diámetro ocular. Su cuerpo es menos fusiforme y está localizada tanto su primera aleta dorsal como la anal, más cerca de la región cefálica que en el resto de las especies estudiadas. Ademas la longitud de su penduculo caudal es mas grande que en las otras. Ch . ocotlanae, se encuentra en una posición intermedia entre las dos especies, asemejándose por sus características, más a Ch. consocium que aCh. chapalae. DISCUSIÓNLas tres especies estudiadas del Género Chirostoma, presentan diferencias importantes en relación a variables de marcado carácter funcional, confiriéndoles singulares adaptaciones al medio en el que se localizan. Dicha separación morfofuncional puede haber sido un mecanismo desarrollado con el fin de reducir los fenómenos de competencia interespecífica, generando ventajas selectivas, de una especie respecto a otras, en la explotación de recursos (Keast y Webbs, 1966; - Barbour, 1973; Werner y Hall, 1977; Van Oijen, 1982). Si se combinan la forma del cuerpo, características de las aletas ( tipo de movimiento), grado de maniobrabilidad, poder natatorio y velocidad, posición, estructura y características bucales se podrá comprender mejor cual es el posible modo de vida, tendencias alimenticias y tipo de hábitat utilizado por las diferentes especies (Keast y Webbs, 1966). Por otro lado, las variables consideradas indican claramente adaptaciones que permiten explotar recursos tróficos diversos. A la luz de los resultados es Ch. chapalae, la especie que se separa más nitidamente de las otras. Esta especie, debido a las características que encontramos para los distintos órganos de locomoción del cuerpo, se diferencia de Ch. consocium y Ch. ocotlanae, por presentar una elevada agilidad en sus movimientos y muy hábil en sus maniobras, debido a que posee un cuerpo bastante fusiforme, unido a sus aletas pectorales que poseen una gran longitud. Además, esta especie, presenta de forma significativa una posición retrasada de la primera aleta dorsal y aleta anal respecto a la región cefálica, así como una base de esta última aleta, mucho mayor. Este conjunto de caracteres, junto a la disposición cefálica de sus ojos, posición dorso-terminal de la boca y su peculiar dentición, definen a Ch. chapalae, como una especie de gran capacidad de equilibrio y estabilidad, con las características necesarias para desenvolverse en una forma de vida muy superficial, especialmente adaptada a una alimentación de tipo planctónico EnCh. consocium y Ch. ocotlanae, aunque muy parecidas en forma y morfología, sus estructuras anatómicas incluyen una serie de formas distintivas del cuerpo, habilidad en el movimiento y configuración de sus mandíbulas que están correlacionadas con diferentes modos de vida, grado de maniobrabilidad, poder natatorio y velocidad, así como niveles de alimentación (Keast y Webbs, 1966). Tanto las características definidas por la forma del cuerpo, como la posición dorso-terminal de la boca, junto con la elevada densidad denticular de sus mandíbulas, son las apropiadas para las especies con tendencias hacia una alimentación de presa de tamaño pequeño (tipo planctónico). Sus pequeños y comprimidos cuerpos maxímizan la velocidad de giro, mientras que minimizan la potencia requerida para el mismo. Combinando este aspecto, con las características de las aletas, la morfología resultante les permite ajustarse a una variedad de planos a través de una ondulación simple de aletas o remado (Werner, 1977; Werner y Hall, 1977). Además, tanto los numerosos y pequeños dientes que poseen sus mandíbulas, como la presencia de su elevada densidad de branquispinas en rastrillo, cumplen una función de filtro mecánico muy eficiente, para la captura de presas pequeñas (Drenner et al, 1984). A pesar de su gran semejanza, Ch. ocotlanae, se presenta como una especie más ágil y hábil en los mecanismos de navegación que Ch. consocium. Estos rasgos funcionales vienen determinados por sus características morfológicas. Ch. ocotlanae posee un cuerpo, más fusiforme que Ch. consocium, además la ubicación tanto de la primera aleta dorsal como de la anal se encuentran más distanciadas de la región cefálica, por ello esta especie tiene mayor capacidad de estabilidad y equilibrio para la navegación. Este hecho viene también apoyado, por la mayor longitud de la base de la aleta anal que actúa con efecto compensatorio (Keast y Webbs, 1966). La longitud de las aletas pectorales en Ch. ocotlanae son mayores que en Ch. consocium; dicha característica le proporciona a la primera, una mayor capacidad de maniobra en sus movimientos a la vez que aporta un mayor equilibrio en la navegación, así como una mayor capacidad de propulsión normal de los individuos. Si observamos los resultados obtenidos para estas variables en Ch. consocium, se aprecia que la longitud de las aletas pectorales es inferior, por tanto, menor será la capacidad de maniobra, estabilidad y propulsión de los individuos. Por otro lado, la primera aleta dorsal y la aleta anal, se encuentran en una posición más cercana de la región cefálica; lo que indica que, la capacidad de equilibrio tiene que ser menor que en las otras especies estudiadas, junto con, la menor base que presenta la aleta anal. Si bien, ambas especies poseen características muy determinantes de una alimentación de tipo planetónica (posición dorso-terminal de la boca, alta densidad denticular tanto de la mandibula inferior como de la superior y alta densidad de branquispinas), debido a las características funcionales, Ch. ocotlanae, tendrá mayor capacidad de explotación de presas más evasivas. Las especies estrechamente emparentadas y en especial las congenéricas , a menudo son muy semejantes morfológica, fisiológica, etológica y ecológicamente. Las aquí estudiadas, son un claro ejemplo de ésto. A consecuencia de ello, para que estas especies puedan coexistir en el mismo ecosistema, se tienen que dar diferencias ecológicas sutiles, pero importantes entre ellas, siendo su estudio de vital importancia para definir el papel que representa una especie en su comunidad y sus interacciones con otras (Pianka, 1982). En una cuenca de la naturaleza del Lago de Chapala, de gran extensión y antigüedad, es de esperar que la coexistencia del gran número de especies endémicas que en él se localizan, sea el resultado de una evolución radial y una perfecta adaptación en base a explotar la gran abundancia de recursos (espaciales y tróficos) existentes en el medio, más que el resultado de un tiempo de aislamiento (Barbour, 1973). LITERATURAADAMICK A, P.The Hauge. Contributions to the functional anatorny of the feeding apparatus of five cyprindis of Parakrama Samudra (Sri Lanka): In Limnológy, of Parakrama Samudra-Sri Lanka. Schiemer, F.(ed). (1983). 171-175. ALEXANDER, R. McN. J. Zool., Lond The functions and mechanisms of the protusible upper jaws of some Acanthopterygian fish.(1967).43-64. 151: ARREGUIN, F. Plan piscípcola Chapala. Tesis. Universidad Autónoma Guadalajara. Guadalajara, Jal México.(1979).p:102. BARBOUP, C. D.Copeia A biogegraphical history of Chirostoma (Pisces:Atherinidae): A species flock from the Mexican Plateau.(1973).533-556. 3: BENGSTON,D. A. Mar. Ecol Prog. Ser.Resource partitioning, by Menidia menidia bery1lina (Osteichtyes: Atherinidae). (1984).21-30. 18: BRABRAND, A. Pol Arch. Hydrobiol. Microhabitat segregation between bream (Abramis brama L.) and white bream (Blicca bjoerkna L.) in a mesotropic lake SE Norway.(1984). 99-108. 31 (2): CASTELLO, V. Utilización de técnicas multivariantes en el estudio de la variabilidad morfológica de una población de barbos (Barbus barbus sclateri G.) (Pisces:Cyprinidae) en el Sur de España. Historia Natural (1983). 1-15.3 (1): DE MARTINI, E.E. The Wasmann Journal of Biology,A correlative study of the ecology and comparative feeding mechanism morphology of the Embiotocidae (Surf-Fishes) as evidence of the family's adaptive radiation into available ecological niches1969177-247.27 (2): DE SILVA, S.S.; CAMARANATUNGA ,P.R.T. y DE SILVA, C.D. Neth. Jour. Zool. Food, feeding ecology and morphological features associated with feeding of four co-occuring cyprinids (Pisces: Cyprinidac). (1980).54-73.30 (1): DIXON, W.J. Biomedical Computers Programs. BMDP. University of California Press USA. (1975).p:791. DIXON, WJ. Statistical Software. BMDP. University of Califronia Press. USA(1981). p: 344. DRENNER, R.W.; MUMMERT, J.R. DeNOYELLES, F. y KETTLE, D. Limnol. Oceanogr.Selective particle ingestion by a filter-feeding fish and its impact on phytoptankton community structure. (1984). 941-948. 29 (5): ETNIER, D.A. Trans. Amer. Fish. Soc. Food of three species of sunfishes (Lepomis, Centrarchidae) and their Minnesota Lake. (1971).124-128. 1: GATZ, A.J., Jr. Ecology Community organization in fishes as indicated by morphological features. (1979).711-718.60 (4): GATZ, A.J., Jr. Amer. Midl. Nat. Morphologically inferred niche differentation in stream fishes. (1981).10-20. 106 (1): GAUSE, G.E The struggle for existence. Baltimore, Denver.(1934). GIRONA, R. Tesis de Licenciatura. Ecología de la población de Carpin del embalse de Arrocampo (Cáceres). Universidad de Sevilla. (1981). p:89. GLADFELTER, W.B. y JHONSON, W. S EcologyFeeding niche separation in a guild of tropical reef fishcs (Holocentridae).(1983).552-563. 64 (3): GRANADO, C. Tesis Doctoral. Ecología de la comunidad ictica del embalse de Arrocampo (Río Tajo. Cáceres). Universidad de Sevilla.(1983). p: 364. GREENWOOD, P.H. Bull. Br. Mus. Nat. HisL (Zool). Suppl, The cichlid fishes of Lake Victoria, East Africa: The biology and evolution of species flock. (1974). 1-134.6: GUILLEN. E. Tesis de Licenciatura. Estudio ecológico de la ictifauna del embalse de Torrejón (Río Tajo, Parque Natural de Monfragüe, Cáceres) Universidad de Sevilla.(1982). p:199. HUTCHINSON, G.E Atretise on Limnology Wiley. p: (1957).741. Vol. I. KEAST, A. y WEBBS, D. J. Fish. Res.Bd Canada. Mouth and body form relative to feeding ecology in the fish faune of small lake, Lakc Opinicon, Ontario.(1966).1845-1877 23 (12): KUX,Z. y LIBOSVARSKY J. Folia ZooL, Variable morphological characters of Gobio gobio (Cyprinidae) examined by principal component analysis. (1981). 229-240. 30 (3): LIBOSVARSKY, J. Folia Zool Note on comparasion of morphornctric characters in fishes of unequal body size. (1982).169-174.31 (2): LIBOSVARSKY, J.; VOLSKIS, R.;WLODEK, J. y GAJDUSEX.J.Folia Zool, Comparasion of morphometric characters of vimba and bream using correlation matrices. (1981). 75-82.30 (1): LIEM, K,F. Amer. Zool Adaptative significance of intra and interespecific differences in the feeding repertoires of Cichlid fishes. (1980).295- 314. 20: LIEM, K.E y OSSE, J. W. M. Amer Zool. Biological Versatility.Evolution and food explotation in African Cichlid fishes.(1975).427-45415 (2): MATTIHES, II. Bijdr. Dierk A comparative study of the feeding mechanisms of some african cyprinidae (Pisces: Cypriniformes). (1963). 1-36.33: MOYLE, P.B. Trans. Amer. Fish. Soc. Ecological segregation among three species of minnows (Cyprinidae) in a Minnesota Lake.(1973).794-805. 4: PIANKA, E.R. Ecología evolutiva. Ed. Omega. Barcelona(1982). p 365. SOKAL, R.R. y ROHLF, FJ.Biometria. Prinicipios y métodos estadísticos en la investigación biológica. Ed. H.Blume. Madrid.(1979) p:832. VAN OIJEN, M.J.P. Neth. Zool. Ecological diferentation among the piscivorous Haplochromine Cichilds of Lake Victoria (East Africa) (1982).336-363.32 (3): WERNER R, E. E. Amer.Natur., III Species packing and niche complementary in there sunfishes.(1977). 553-578. (979). WERNER. E.E. Sport. Fish. Ins. Niche partitioning by food size in fish comunities From:Predator-Prey sistem in fisheries management R.II.Stroud and H. Clepper (Eds.). Washington, D. C. (1979).311-322. WERNER, E.E. y HALL,D.J Ecology Compctition and habitat shift in two sunfishes (CEntrarchidae). (1977).869-876. 58 ZARET,T.M. y RAND,A.S. EcologyCompetition in tropical stream fishes: Support for the competitive exclusion principle. (1971). 336-342.52: ZIESLER,R. y ARDIZZONE,G.D. Documento técnico. Las agas continentales de América Latina. FAO.ROMA. COPESCAL. (1979). p:171.1
|