|
ARRECIFES PROFUNDOS EN LA ISLA DE COZUMEL, MÉXICO.
Trabajo
recibido el 11 de junio de 1987 y aceptado para su publicación el 16 de
diciembre de 1987.
Eric Jordán Dahlgren
Estación Puerto Morelos, Instituto de
Ciencias del Mar y Limnología, UNAM. Contribución No. 634 del
Instituto de Ciencias del Mar y Limnología, UNAM.
Se estudia la morfología y composición biótica
de la comunidad coralina de los arrecifes profundos de Cozumel. Estos arrecifes
que se encuentran a más de 20 m de profundidad estan densamente poblados
por una comunidad coralina en la que los corales escleractineos no son los
controladores de substrato más importantes. Un alto grado de
bioerosión sobre la superficie arrecifal, determina que la mayor parte
de la superficie disponible se encuentre en condiciones de baja
iluminación y penumbra, donde organismos no simbióticos son
más eficientes que los corales escleractincos. El aporte de terrigenos
sobre estos arrecifes es nulo, debido a la naturaleza karstica del substrato
insular, a la vez que estan inmersos en un medio oceánico por el paso de
la corriente de Yucatán. Su profundo emplazamiento a sotavento de la
isla determina un alto nivel de protección a las olas generadas por
temporales, protegiendo así la debilitada estructura arrecifal y
favoreciendo una lenta tasa de destrucción. Se propone una
hipótesis sobre el origen y desarrollo posterior de esos arrecifes
profundos, basada en las tasas de ascenso del mar durante la última
transgresión marina y en condiciones ambientales estables en los
últimos 5000 años.
The morphology and general community
structure of deep setting reefs on the lee of Cozumel island is studied. These
submerged reefs, more than 20 m bellow the water surface, are colonized by a
dense coral community in which scleractinian corals are restricted to high
ilumination areas and whose overall importance is lower than that of
non-symbiotic organisms. This predominance of non-symbiotic organisms upon the
reef formation as a whole, obeys to a high degree of bioerosion on the
calcareous reef structure resulting in abundant shade and penumbra areas were
scleractininan corals are unable to grow. Land influence on these submerged
reefs is negligible due to the karstic nature of the island substrate and the
flow of oceanic waters carried by the Yucatan current. The reefs are also well
protected from the disruptive effect of long period waves produced by
hurricanes and storms, due to the deep,leward basement upon which these reefs
are found. On this basis, an hypothesis on the origin and further development
of these reefs is presented, which assumes that the actual structure is a
result of the high rates of sea level ascent during the Holocenic transgression
and very stable environmental conditions for the past 5000
years.
Una notoria característica de los arrecifes coralinos modernos es la amplia variabilidad que presentan tanto en su morfología, como en los patrones de distribución y abundancia de la comunidad coralina que los coloniza. Esta variabilidad es resultado de un complejo proceso de interacciones tanto geológicas como biológicas, situación que dificulta la posibilidad de establecer una relación funcional entre el arrecife y la comunidad coralina del presente. En términos del proceso de desarrollo de un arrecife moderno, existen dos componentes temporales principales: Uno de largo plazo determinado por la historia geológica de una localidad (Stoddart,1969; Hopley,1982); y otro de mediano plazo, que resulta de cambios en la estabilidad climática y tasas de ascenso del nivel del mar, durante la transgresión Holocénica (Davies, 1977; James y Ginsburg, 1979; Shin et a1,1982; Davies y Montaggioni,1985). En la organización de la comunidad coralina que en la actualidad habita en la formación arrecifal, intervienen multitud de interacciones ecológicas cuya temporalidad se estima en decenas de años, y las que reflejan en mayor o menor medida los determinantes geológicos que afectan a la estructura arrecifal. A la vez, otros factores externos al sistema juegan un importante papel en el desarrollo y mantenimiento de la estructura comunitaria y del propio arrecife coralino, tales como: La cantidad y calidad de aportes terrígenos por descargas continentales o insulares (Stoddart, 1969; Purdy, 1974); la interacción física, y de intercambio de materiales y energía entre ecosistemas costeros tropicales: manglares, seibadales y arrecifes coratinos (Ogden y Gladfelter, 1983); la frecuencia y magnitud de ciclones y tormentas tropicales (Stoddart, 1974; Roberts, 1974). A la fecha sin embargo, es difícil evaluar (excepto en situaciones catastróficas) la importancia real que estos factores externos tienen en cada caso específico. Todos estos factores, tanto históricos como recientes, aunados a la compleja dinámica de la comunidad coralina determinan un mosaico de microambientes, donde diferentes secuencias sucesionales son la norma más que la excepción (Jordán, en prensa). Las diferentes expresiones que la comunidad coralina tiene a nivel local reflejan por tanto, no sólo el resultado de las interacciones ecológicas del presente, sino también un complejo componente histórico, independientemente de que una comunidad compleja tiene diferentes vías de integración (May, 1974). A la fecha y en la región del Caribe, la mayor parte de la información que se utiliza para entender la dinámica de la comunidad coralina de un arrecife dado, se obtiene de análisis de estructura comunitaria, en pequeñas secciones arrecifales, básicamente dirigidos a la fauna escleractinea (especies clave en la integración de la comunidad) y de gorgonaceos, los que corresponden al taxon más aparente en muchos arrecifes del Caribe (Porter, 1972; Kinzie, 1973; Goldberg, 1973; Bak, 1977; Behety, 1975; Loya, 1976; Jordán, 1979; Jordán et al, 1981; Muzik, 1982; Lasker y Coffroth, 1983). O a interpretaciones de desarrollo estructural basados en la historia geológica del sistema (Newell y Rigby, 1957; Stoddart, 1969; Shinn et al., 1982; Ginsburg y Choi, 1983). No es sino muy recientemente y gracias a los trabajos anteriores, que se comprende que este tipo de estudios, muy valiosos en su contexto, aportan poca información de carácter realmente útil para evaluar la condicion y grado de desarrollo de un arrecife en términos de generar alternativas racionales de manejo: básicamente por que minimizan la importancia (o la excluyen) de otros componentes bióticos e históricos, que es necesario considerar para lograr una aproximación realista a las condiciones del sistema en estudio. Esta situación determina que la evaluación del grado de desarrollo y condición de arrecifes coralinos, rcquiera analizar la problemática desde un contexto más amplio que incluya áreas espacialmente significativas y otros componentes de la comunidad coralina, y particularmente enfocar el análisis desde un contexto histórico, en el que procesos acaecidos mucho tiempo antes, pueden ser determinantes en la estructura actual de un arrecife coralino. En este trabajo se pretende analizar la naturaleza de los arrecifes profundos de la isla de Cozumel que son particularmente favorables a este tipo de análisis global, particularmente por la situación profunda y a sotavento que presentan, por la naturaleza karstica del substrato y la permanente influencia de la corriente de Yucatán a su paso por el canal de Cozumel. Esto determina que los efectos de aporte insular, los de interacción con otros sistemas y los que producen los ciclones sean practicamente nulos, por lo que se puede considerar para los fines de este trabajo que la condición actual que presentan estos arrecifes, ha sido determinada por procesos históricos y la dinámica comunitaria interna, en condiciones ambientales que han sido estables desde hace más de 5000 años. ÁREA DE ESTUDIOLa isla de Cozumel se encuentra localizada a los 20°28' de latitud norte y a los 86°55' de latitud oeste, frente a la costa oriental de la Península de Yucatán (Fig.1). Esta isla alargada, con su eje mayor en sentido sur-norte, es la isla de mayor tamaño en aguas mexicanas. La isla de Cozumel se encuentra en un ambiente netamente marino, que se caracteriza por un movimiento constante de las masas de agua, las que tienen un bajo contenido de nutrientes, temperaturas por arriba de los 23°C y presentan una elevada transparencia. Este ambiente es determinado por el flujo permanente de la corriente de Yucatán (Wust,1963; Merino,en prensa), que a lo largo del margen de sotavento, alcanza velocidades mayores a tres o cuatro nudos en el verano, (Fig. 2). No existe indicación de que el origen de la isla se deba a un proceso de acumulación de calizas arrecifalcs sobre fallas subsidentes, como sucede en el caso del Banco Chinchorro (Jordán y Martín, 1987), y los atolones del complejo Beliceño (James y Ginsburg,1979). Probablemente el origen de la isla corresponde a un desprendimiento del margen oriental de la Península, durante la formación de la cuenca de Yucatán, entre el Mesozoico tardío y el Cenozoico temprano (Uchupi, 1973). 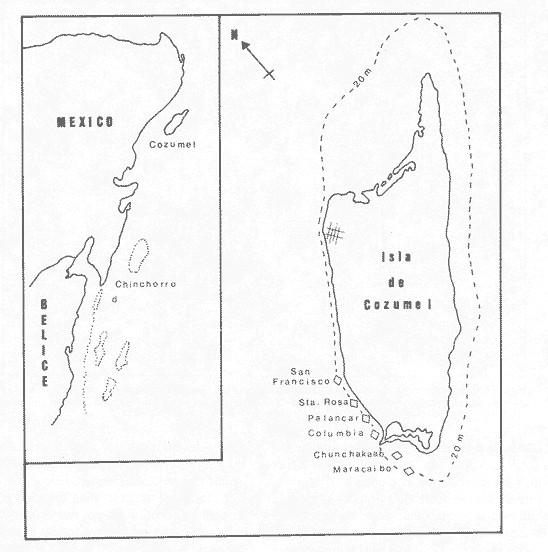 Figura 1. Localización del área de estudioy situación de los arrecifes profundos de la isla de Cozumel, Nótese que estos arrecifes se encuentran exclusivamente en la parte sur y a sotavento de la isla. La isla, constituída por calizas sedimentarias, probablemente se encuentra al igual que toda la Península de Yucatán, sobre un basamento metamórfico del Paleozoico, cubierto por más de 500 m de depósitos, fundamentalmente lechos rojos del Jurásico que subyacen una sucesión de areniscas y calizas depositadas durante el Cretácico, y el Paleoceno, y sobre las que son comunes afloramientos del Terciario (Aguayoet al., 1978). La naturaleza karstica de la isla impide la formación de ríos en su superficie, ya que toda el agua de lluvia percola a través de fracturas y fisuras en el terreno, hasta el nivel freático; por lo que los escurrimientos hacia el mar practicamente no acarrean sólidos en solución. La caractrerística geomorfológica mas notoria de la isla, es la presencia de una terraza submarina, que en el margen occidental es corta y termina entre 20 y 30 m de profundidad, dando lugar al talud insular que se precipita en una pendiente cercana a la vertical, hasta profundidades mayores de 400 m. El origen de esta terraza que constituye una plataforma insular, no ha sido determinado, pero es muy probable que corresponda a una terraza de erosión, muy comunes en todo el Caribe y que fueron formadas por exposición alternada a ambientes submarinos y subaéreos durante las transgresiones marinas del Pleistoceno (Logan et al, 1969). 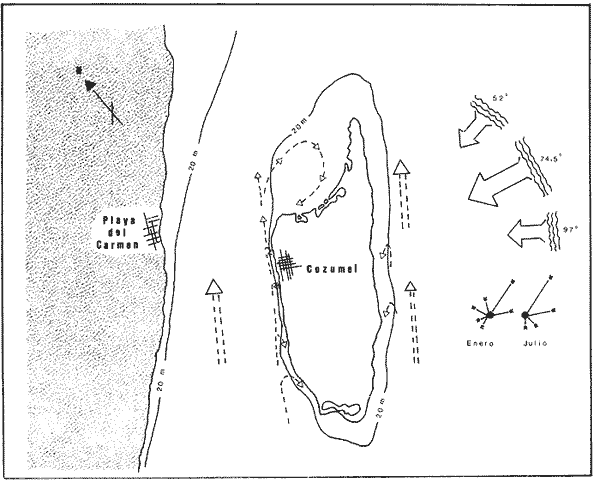 Figura 2. Corrientes, vientos y oleaje predominante sobre la isla Cozumel. Datos tomados de Pilot Charts y derroteros de navegación. Estimación de la Dirección del oleaje predominante basado en Burke 1982. Sobre el borde de esta terraza submarina, en la porción sur y a sotavento de la isla, se encuentra una serie de formaciones arrecifales, que siguiendo el contorno del borde, forman una especie de parapeto arrecifal sobre el talud insular y que constituye los arrecifes profundos de Cozumel (Fig.3). Estas formaciones arrecifales son discontinuas y están integradas por numerosas estructuras más o menos aisladas entre sí, de dimensión variable y que en conjunto se extienden por más de 9 km, siempre sobre el borde de la plataforma insular en la porción suroeste de la isla. Hacia la región central y norte de la plataforma, estas formaciones arrecifales desaparecen y el substrato calcáreo es colonizado por una rica y diversa comunidad coralina, sin presentar una acreción significativa. Localmente se denomina arrecife a grandes áreas arrecifales constituidas por numerosas formaciones como es el caso de los arrecifes de Palancar, Colombia, Maracaibo, etc. En este trabajo el término arrecife se emplea para indicar una zona donde ha habido acreción arrecifal coralina y puede referirse según el sentido del texto, a una sola estructura o al conjunto de estructuras de un área arrecifal dada, cuyo nombre corresponde con la toponimia local (Fig.1). METODOLOGÍAMorfología Arrecifal.- La distribución de las areas arrecifales de Cozumel es bien conocida por los lugareños y concuerda con observaciones realizadas en fotografías aéreas y vuelos ex-profeso sobre el área de estudio. Un mapa turístico (Hajoysky,1974) y una guía de buceo de Cozumel (Lewbel y Martin 1984) fueron utilizados corno base general para ubicar las zonas con formaciones arrecifales (Fig.1). Las cartas de navegación publicadas sobre Cozumel solo incluyen los arrecifes superficiales que constituyen un peligro para la navegación (Cartas de la Secretaría de Marina: 900, 1977; 925, 1978; 924, 1982). 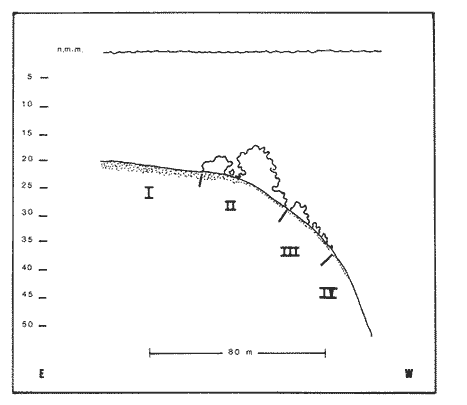 Figura 3. Morfología general de los arrecifes profundos de Cozumel. I.- Plataforma insular, normalmente cubierta por una capa de arena de espesor variable. II.- Zona de desarrollo arrecifal en el borde de la plataforma insular. III.- Zona de pedacería arrecifal al inicio del talud. IV.- Talud insular. La escala de profundidades es en metros. En virtud de que cada área arrecifal esta constituida por numerosas estructuras arrecifales, se inició el estudio morfológico con un reconocimiento general que permitiera tipificar las diferentes formaciones arrecifales. El reconocimiento se llevó a cabo en dos etapas: 1) Sondeo batimétrico en cada área arrecifal, con el objeto de definir la geomorfología general, mediante transectos perpendiculares a la costa, empezando por la orilla y terminando en el límite de detección de la ecosonda (más de 200 m de profundidad). 2) Reconocimiento submarino, por buceo de deriva a lo largo del borde registrando características morfológicas básicas de cada tipo de formación como dimensiones, orientación, ubicación y particularmente la similitud o disimilitud con otras formaciones arrecifales adyacentes. Estos registros fueron controlados por tiempo de deriva bajo el agua y seguimiento en superficie de boyas señaladoras de la posición de los buzos. A intervalos fijos se determinó la posición de las boyas por triangulación (sextante), con objetos o rasgos fisiográficos de posición conocida a lo largo del litoral (Jordán,1987), posiciones que eran posteriormente correlacionadas con los tiempos de deriva para definir la ubicación de los registros submarinos. Análisis comunitario.- El análisis de la estructura de la comunidad coralina se basa en la asignación de atributos morfológico-funcionales a la biota que controla el substrato arrecifal, lo que permite agrupar a las especies dominamtes en tipos estructurales básicos, independientemente de su identidad específica (Bradbury et al, 1986). Los atributos son definidos en términos del papel primario que las especies que constituyen la comuinidad de superficie juegan en el mantenimiento de la estructura arrecifal, reconociendose tres tipos básicos: 1) Constructores arrecifales principalmente corales escleractineos y algunas esponjas de ambientes profundos. 2) Organismos de carácter temporal o definitivo que compiten por el espacio, pero no contribuyen a la acreción arrecifal, tales como algas, esponjas erectas, anemonas coloniales y gorgonaceos. 3) Organismos biohoradadores (detectables en superficie), básicamente esponjas en placa y perforantes. Así, en cada formación arrecifal el censo consiste en medir el porcentaje de la superficie que corresponde a los siguientes atributos: 1) Corales (corales escleractinios y especies del hidrocoralMillepora). 2) Macroalgas y algas filamentosas. 3) Esponjas. 4) Otra biota (Gorgonaceos, anemonas coloniales, etc.) 5) Substrato abiótico, que incluye calizas expuestas, grava y arena. El atributo corales es dividido, además, en cinco subcategorías de acuerdo con el papel que juegan en la constitución arrecifal: A) Acropora palmata, especie constructora primaria de rápido crecimiento. B) Masivos, incluyendo aquí aMontastrea annularis, el otro constructor arrecifal fundamental en el Caribe. C) Incrustantes o en placa. D) Foliosos. E) Ramificados menores. La proporción de substrato ocupada por cada atributo se estimó mediante el muestreo de transectos unldimensionales, en sentido perpendicular a la estructura arrecifal (Loya, 1978; Jordán, 1980). El transecto es delimitado mediante la extensión de una cadena plástica que colocada sobre el fondo, sigue todo contorno del arrecife, en sentido perpendicular al borde de la plataforma. Esta disposición del transecto permite censar los tres ambientes principales del sustrato arrecifal: base, lados, superficie superior (Jordán, 1987). La cadena está constituída por eslabones de 2.57 cm cada uno, lo que permite estimar directamente la proporcionalidad de cada atributo. En virtud de que las formaciones arrecifales bajo estudio son relativamente profundas, el tiempo de trabajo útil bajo el agua es menor a dos horas por día por buzo, lo que determina una baja eficiencia de muestreo y un elevadísimo costo de operación. Por esa razón el sistema de muestreo fue establecido arbitrariamente, con base en la experiencia de muestreos cuantitativos previos (Jordán, 1980; 1987; Jordán et al, 1981), censos similares (Jordán y Martín, a; b; 1987) y consideraciones logísticas. Con base en el reconocimiento general, se seleccionó una estructura arrecifal "representativa" para cada área arrecifal importante: Chunchakaab, Colombia, Palancar y Santa Rosa. En cada una de ellas, el muestreo consistió en la realización de cinco transectos por cada estructura arrecifal analizada, fijando el límite de profundidad para el censado en 40 m (el reconocimiento sin embargo, se extendió hasta los 60 m de profundidad). El carácter de estas mediciones es por tanto semicuantitativo, pero el amplio reconocimiento preliminar asegura una apropiada representación de la biota que controla el substrato en cada formación arrecifal estudiada. A diferencia de los análisis tradicionales de estructura comunitatia en los que la discriminación específica es fundamental y un elevado grado de detalle es priorizado a la amplitud del muestreo, en este trabajo el método de muestreo se basa en cubrir la mayor área posible con el mínimo detalle necesario, pero incluyendo a toda la biota de superficie que es capaz de controlar el sustrato (Kinzie y Snider, 1978; Bradburyet al., 1986). Se elaboró además, una lista preliminar de las especies de corales identificados durante el muestreo. La identificación se realizó básicamente "in situ" y sólo aquellas especies de identificación dudosa fueron colectadas para su posterior identificación en el laboratorio, de acuerdo con Wells (1957) y Goreau y Wells (1967). RESULTADOSLos arrecifes estudiados se encuentran a profundidades por lo regular mayores de 20m y pueden alcanzar hasta 5 m de altura sobre el nivel de la plataforma en el lado interno del arrecife (hacia la costa) y, en el lado externo, descender hasta más de 40 m de profundidad sobre el talud insular (Tabla 1). Sin embargo, el límite inferior de las estructuras es dificil de precisar, ya que sobre la pendiente del talud insular, el ambiente arrecifal se puede extender hasta más de 60 m de profundidad, debido a la presencia de abundantes desprendimientos de la caliza arrecifal que proveen de un sustrato duro sobre la capa arenosa a diversas especies de la comunidad coralina profunda. A partir de esa profundidad, la inclinación del talud se aproxima a la vertical impidiendo la acumulación de arena y de los desprendimientos arrecifales. La abrupta caida del talud insular favorece un drenaje natural del sedimento libre, permitiendo que las formaciones coralinas que se desarrollan en estas áreas alcancen dimensiones considerablemente mayores que las de otras formaciones arrecifales de la isla, que crecieron en la parte interna de la plataforma insular. Una excepción a este planteamiento es el arrecife de Chunchakaab, que aunque no se encuentra directamente sobre el borde del talud, presentan características similares a las de los arrecifes profundos (Fig.4). Aún cuando estos arrecifes presentan una estructura interna bien consolidada, apreciable en las paredes internas de cavernas y fisuras de la estructura arrecifal, la costra calcárea superficial que constituye el sustrato sobre el que se desarrolla la comunidad coralina se encuentra horadada y erosionada en alto grado. Las paredes laterales, las áreas internas expuestas y las zonas basales del arrecife son las que muestran el mayor grado de deterioro, y estas áreas corresponden a más del 60 o 70% de la superficie total de las formaciones arrecifales. 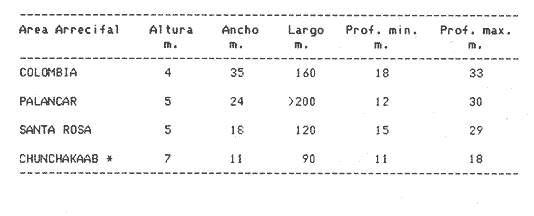 TABLA 1 DIMENSIONES DE LOS ARRECIFES DE BORDE COZUMEL MAS DIMENSIONES CORRESPONDEN A LAS DEFORMACIONES ARRECIFALES AISLADAS, REPRESENTATIVAS DE TODAS LAS QUE CONSTITUYEN EL ÁREA ARRECIFAL INDICADA Y NO SE REFIERE A TODA LA FORMACIÓN DEL BORDE, LOCALMENTE CONOCIDAS COMO ARRECIFES Y QUE SE EXTIENDEN POR MUCHOS CIENTOS DE METROS (VER TEXTO). CHUNCHAKAAB *, ESTE ARRECIFE SE ENCUENTRA SOBRE LA PLATAFORMA Y NO EN EL BORDE DE LA MISMA, VER FIG. 1. LAS DIMENSIONES (ALTURA, ANCHO, ETC). INDICAN LOS VALORES MÁXIMOS DE LA FORMACIÓN ARRECIFAL, ANALIZADA EN EL ÁREA ESPECIFICADA  Figura 4. Vista de la parte superior de los arrecifes hacia el talud insular. Se puede apreciar una amplia hendidura en el macizo arrecifal y los bordes de la misma densamente colonizados por biota coralina. Al fondo se observa la capa arenosa, salpicada con pedacería de coral. Una característica de los arrecifes profundos, es una elevada densidad poblacional entre los organismos que controlan el sustrato arrecifal donde la abundancia de corales escleractineos es relativamente baja, pudiéndose apreciar que algas y esponjas juegan un papel muy importante en todos los casos (Fig.5). Especialmente notorio es que los sustratos libres tengan un área tan reducida, sugiriendo que las condiciones ambientales son en extremo favorables para la flora coralina que aunque en forma esporádica, controla todos los sustratos duros no colonizados permanentemente por otros organismos (Dahl,1973). En la misma figura 5, se puede apreciar que los Acroporidos están prácticamente ausentes y que con excepción de Chunchakaab (que corresponde a un arrecife de plataforma y más somero que los restantes), los corales masivos tienen una baja representación en favor de formas incrustantes y foliadas. La virtual ausencia de Acropora palmata y la escasa abundancia deMontastrea annularis y otras especies de corallum masivo, son notorias, particularmente por el importante papel que estas especies juegan en la integración de la estructura arrecifal (Shinn, 1963; Shinn et al, 1982; Newell y Rigby, 1957; Dunstan, 1975). 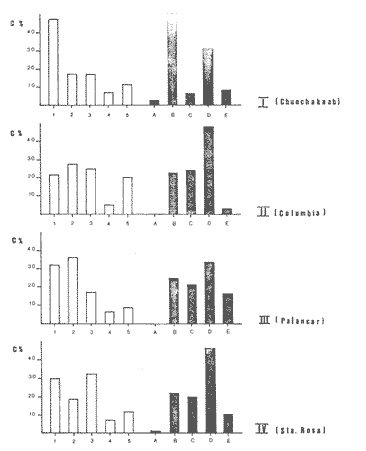 Figura 5. Porcentaje de cobertura del sustrato arrecifal en los arrecifes profundos de Cozumel. Los números corresponden a los diferentes atributos morfológico-funcionales seleccionados (ver texto): 1.- Corales; 2.- Algas; 3.- Esponjas; 4.- Otros organismos; 5.- Abiótico. Las letras corresponden a subcategorías morfológicas del atributo corales: A.- Ramificados estructurales, básicamente Acropora palmata; B. Corales de esqueleto masivo; C.- Incrustaciones o en placa; D.- Foliados; E.- Ramificados menores. Las desviaciones de las medias indicadas son en todos los casos muy amplias y se han omitido con propósitos de claridad. Aún cuando hay una alta riqueza específica de corales escleractineos (Tabla 2), con coberturas muy elevadas en zonas específicas y que grandes colonias de corales sean comunes en estos arrecifes, en términos de la superficie total de la estructura arrecifal la importancia de los corales escleractineos es reducida. Es solo en la parte superior y bien iluminada de los arrecifes, donde la cobertura del substrato tiende a ser dominada por corales escleractineos, cuya distribución en parche permite sin embargo, que parte del sustrato superior se encuentre colonizado por otros organísmos (Tabla 3). Los espacios no ocupados por corales o esponjas, en éstas partes superiores, son normalmente dominados por feofitas carnosas, comoPokokiella variegata y algas calcáreas del género Halimeda.  TABLA 2 LISTA PRELIMINAR DE ESPECIES DE ESCLERACTINESOS QUE HABITAN EN LOS ARRECIFES PROFUNDOS DE COZUMEL. LAS ESTIMACIONES DE ABUNDANCIA SON CUALITATIVAS Y NO PRETENDEN REPRE SENTAR LA ABUNDANCIA REAL DE ESTAS ESPECIES EL ÁREA DE ESTUDIO. + + + = MUY ABUNDANTE; + + = COMUN; + = PRESENTES; - = AUSENTE. *= ESTE HIDROCORAL SE INCLUYE POR SU FORMA DE CRECIMIENTO CALCÁREA En los abundantes microambientes de sombra y penumbra que resultan de la degradación de la costa arrecifal, la biota controladora del substrato está compuesta principalmente por esponjas, algas, gorgonaceos y otros alcionarios no simbióticos. La comunidad coralina que coloniza las paredes calcáreas del talud por debajo de la zona arrecifal es similar a la de los ambientes de penumbra, y las especies dominantes corresponden a esponjas y alcionarios no simbióticos. 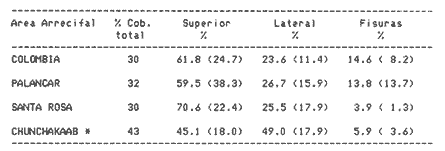 TABLA 3 PORCENTAJE DE COBERTURA POR CORALES ESCLE RACTINEOS EN ARRECIFES DE BORDE. PORCENTAJE EN SUPERIOR, LATERAL Y FISURAS, SE REFIERE AL PORCENTAJE DE LA COBERTURA TOTAL QUE FUE MEDIDO EN LAS DIFERENTES PARTES DE LA ESTRUCTURA ARRECIFAL. EL VALOR ENTRE PARENTESIS INDICA UNA DESVIACIÓN ESTANDARD. CHUCHAKAAB * ESTE ARRECIFE SE ENCUENTRA SOBRE LA PLATAFORMA Y NO EN EL BORDE DE LA MISMA, VER FIG. 1 Esta comunidad domina a todo lo largo del talud insular de sotavento, particularmente en el área central y norte de la isla, donde no hay arrecifes profundos y los escleractineos se restringen a unas cuantas colonias aisladas sobre el borde del talud. (Fig. 4). La reducida densidad poblacional que presentan los corales escleractineos en los arrecifes profundos, puede ser interpretada como el resultado de una baja capacidad competitiva por el sustrato, a la vez que su mayor abundancia en las partes altas y bien iluminadas del arrecife, sugiere que esta situación es generada por la predominancia de ambientes de penumbra y sombra; donde las condiciones de baja iluminación son poco favorables al desarrollo de corales simbióticos. Esto se debe a que aún cuando los corales son politróficos (Muscatine y Porter, 1977), estos obtienen la mayor parte de la energía que requieren para satisfacer sus altas demandas metabólicas, a partir de la fotosíntesis de las zooxantelas (Muscatine et al, 1981; Porter, 1984); las que además, son responsables de acelerar las tasas de calcificación en el coral (Goreau, 1963; Gladfelter, 1985). Para que las zooxantelas puedan producir la cantidad de energía necesaria, requieren de condiciones de alta calidad de iluminación, toda vez que el punto de compensación del par simbionte, se encuentra en niveles más altos de iluminación, que el del componente vegetal aislado (Trench, 1971). Por lo que la abundancia de ambientes de penumbra y sombra, aunado a la localización de los arrecifes por debajo de los 20m de profundidad, determina que el largo tirante de agua filtre la mayor parte del espectro visible aprovechable por las zooxantelas, reduciendo así su eficiencia fotosintética. La figura 6 muestra que a 20m de profundidad la tasa de crecimiento de corales de aguas intermedias, es menor al 50% de la observada a 10m (Dunstan, 1975). 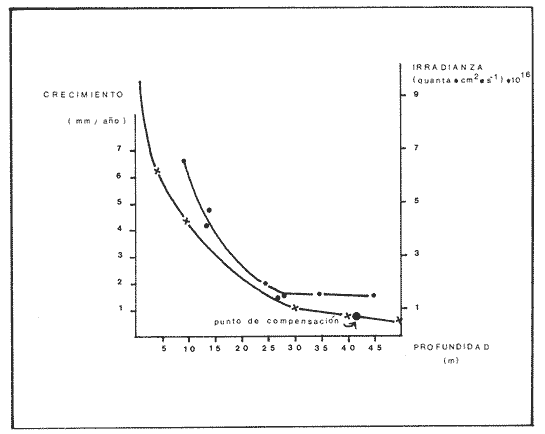 Figura 6. Curva de crecimiento del coral masivo Montastrea annularis, en función de la profundidad (curva inferior), como resultado de la disminución en la eficiencia fotosintética de las zooxanthelas por atenuación de la calidad de la luz al atravesar la columna de agua. La curva representa condiciones de crecimiento en aguas oceánicas con escasa influencia continental. Tomado de Dustan (1975). Esto es lo que determina que sea básicamente en las partes superiores del arrecife, o en las zonas con mayor iluminación lateral, donde los corales alcancen a cubrir áreas significativas de la superficie arrecifal. En el resto de la estructura arrecifal, la densidad poblacional de organismos no simbióticos tiende a ser mayor que la de los corales. Muchos de esos organismos son biohoradadores en mayor o menor medida (Schroeder y Zankl, 1974; Rutzler, 1975). La importancia de los corales escleractineos en la construcción, consolidación y mantenimiento de las estructuras arrecifales es bien conocida, por lo que su reducida densidad poblacional indica una limitada capacidad para balancear los procesos naturales de degradación y horadación que se dan en el seno de la estructura arrecifal, tanto por agentes biológicos como físicos (Stoddart, 1969; Zankl y Schroeder, 1972; Tunnicliffe, 1975). Además de la dominancia de organismos no simbióticos en las superficies poco iluminadas, existen muchos otros biohoradadores de hábitos crípticos que no son registrados por el método de muestreo empleado, y es necesario considerar que la contínua expansión poblacional de estos organismos debilita la estructura arrecifal internamente, a la vez que fomenta la formación de mayor cantidad de ambientes de penumbra, generando un circuito cerrado en el que los procesos destructores tienen una mayor importancia relativa que los procesos constructores, los que dependen fundamentalmente del crecimiento de corales escleractineos. Esta situación, de una preponderancia de los procesos destructores, ha perdurado por largo tiempo como lo indica la abundante pedacería arrecifal, la superficie desgarrada y altamente horadada de los arrecifes, así como la presencia de múltiples cavernas y huecos en la estructura arrecifal (Fig.7).  Figura 7. Composición de la biota coralina en la parte más iluminada de los arrecifes profundos. Se aprecia la predominancia de corales con formas placoides, foliadas y ramificadas menores. Observesé que las colonias de coral tienden a ser de tamaños reducidos y el substrato restante es ocupado por otros organismos como esponjas y algas. Profundidad: 18 m. Por tanto, la condición en que se encuentran los arrecifes profundos sugiere que, después de un activo proceso de formación, evidenciado por la magnitud de las estructuras arrecifales y su alto grado de consolidación, se inició un lento proceso degenerativo, que en las áreas más iluminadas podría ser localmente balanceado por procesos constructivos, pero no revertido en toda la formación arrecifal. DISCUSIÓNCon base en estos resultados es posible especular fundamentadamente sobre el origen y desarrollo ulterior de los arrecifes profundos de Cozumel. Esta discusión se basa en que estos arrecifes fueron incapaces de mantener una tasa de crecimiento lo suficientemente elevada, que permitiera compensar la velocidad de ascenso del nivel del mar durante la transgresión Holocénica, quedando como arrecifes sumergidos (Macintyre et al, 1972; Neumann y Macintyre, 1985). Si bien las condiciones en que actualmente se encuentran estos arrecifes no indica un proceso de crecimiento activo, es evidente que las estructuras que hoy en día existen fueron formadas cuando las condiciones ambientales eran más adecuadas para un proceso de acreción arrecifal. En ésta argumentación se asume que el período formativo de los arrecifes profundos fué, durante la transgresión Holocénica y posterior a la formación de la terraza insular básicamente por el reducido tamaño de las estructuras arrecifales, su localización en el borde de la plataforma y el alto grado de horadación, que solo puede corresponder a procesos bioerosivos en medio submarino (Schroeder y Zankl, 1974). De acuerdo con Bloom et al, (1974), hace 125,000 años el nivel del mar tenía un nivel ligeramente mayor al actual, a partir del cual desciende durante la última época glacial, con varios ascensos durante los períodos interestadiales ninguno de los cuales es mayor de -20m. Se estima que hace 20,000 años el nivel del mar se encontraba aproximadamente unos 135m por debajo del nivel actual, y a partir de entonces la transgresión es ininterrumpida hasta alcanzar el nivel actual del mar. Entre 11,000 y 10,000 años antes de la época presente, el nivel del mar alcanzó el borde de la plataforma insular (Ginsburg y Choi, 1983), por lo que el período formativo de los arrecifes profundos de Cozumel debe haberse iniciado poco después. Este proceso formativo, sin embargo, debe haber sido relativamente lento desde un principio, como sugieren observaciones aisladas en el interior de cavernas y huecos de los arrecifes del borde, donde se apreció que la estructura interna de los arrecifes esta constituida por calizas cementadas y restos de corales masivos; sin encontrar evidencia de una trama de estructuras ramificadas que indique la presencia de corales formadores de rápido crecimiento, particularmente Acropora palmata. La importancia de éste hecho, reside en que la ausencia (o reducida densidad de esta esppecie en la trama estructural implicaría lentas tasas de acreción en la formación de arrecifes Holocénicos (Shinn, 1963; Stoddart, 1969; Shinn et al, 1982). Si bien estas observaciones aisladas no constituyen una evidencia formal de la ausencia de esta especie en la estructura primaria de estos arrecifes, la posición a sotavento de los mismos, en un ambiente con bajo nivel de energía, sugiere que las condiciones dominantes sean poco favorables a un amplio desarrollo poblacional de A. palmata; especie adaptada a los altos niveles de energía de las zonas de rompiente (Geister, 1977). De hecho, hoy en día, en las formaciones arrecifales someras sobre la plataforma de sotavento de Cozumel, entre 10 y 5m de profundidad es raro encontrar colonias de esta especie (Jordán, 1987). De acuerdo con Dillion y Vedder (1973), y Uchupi (1973), en los últimos 10,000 años la plataforma de Yucatán ha sido relativamente estable con respecto a la magnitud de los cambios de nivel del mar, por lo que la dinámica del desarrollo arrecifal estuvo básicamente determinada por el ascenso del nivel del mar. Ahora bien, suponiendo que la acreción resultante durante ese período de crecimiento produjera estructuras de hasta 10m de altura antes de quedar sumergidas a demasiada profundidad, se puede asumir que estos arrecifes deben haber dejado de crecer activamente hace más de 5,000 años, cuando el nivel del mar se encontraba a menos de dos metros por debajo del nivel actual. Por tanto, el período formativo debe haber sido del orden de 5,000 a 6,000 años, dando tasas de acreción media de 2m por cada 1,000 años, como se expresa en la figura 8. 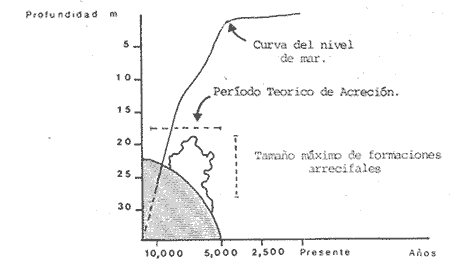 Figura 8. Diagrama que muestra la posible relación entre la estructura arrecifal y la curva de ascenso del nivel del mar fué compilada por Ginsburgy Choi (1983), con base en fechamientos de carbon-14 en turba y carbonatos de la zona arrecifal de Belice. Esta curva es más apropiada para el área de Cozumel, que la curva de ascenso de Bermudas (Beumann, 1971), o la del Atlántico Occidentral (Lightly et al, 1982). Esta tasa de crecimiento arrecifal es claramente inferior a la velocidad de ascenso del nivel del mar, estimado en más de 5.5m por cada 1,000 años. Entre hace 11,000 y 5,000 años, el ascenso no fué regular y durante ese período hubo tasas de ascenso más aceleradas, cercanas a los 10m/1,000 años (Fig.8). En general, el ascenso del nivel del mar desde la parte superior del talud (-35m) hasta los -2m de hace 5,000 años, equivale a un incremento del nivel del mar mayor a 32m en solo 6,000 años (Ginsburg y Choi, 1983). Fechamientos de corales con Carbono-14, realizados por Shinn et al, (1983) en el complejo arrecifal de Belice, señalan tasas de acreción arrecifal durante el Holoceno, de entre 1 y 6 m por cada 1,000 años. Macintyre et al, (1977) reportan tasas de acreción de más de 5 m/1,000 años para el Arrecife de Alacranes, en la misma época. Sin embargo, estas elevadas tasas de acreción, que permitieron a los arrecifes en cuestión compensar el ascenso del nivel del mar, solo han sido reportadas para arrecifes en la zona de barlovento y en los que el constructor arrecifal predominante es A. palmata, especie que suponemos ausente de la estructura arrecifal primaria en los arrecifes profundos de Cozumel. La existencia de estructuras arrecifales sumergidas, incapaces de mantener el ritmo de ascenso del nivel del mar, es relativamente común (Macintyre, 1972; Neumann y Macintyre, 1985). Por ejemplo, en el Banco Chinchorro existen estructuras arrecifales a sotavento que semejan los complejos sistemas de macizos y canales de barlovento (pero que a diferencia de aquellos, solo se levantan uno o dos metros sobre el sustrato Pleistocenico) y que también se caracterizan por la ausencia de A.palmata (Jordán y Martín, 1987). Estos arrecifes de sotavento en Chinchorro, a profundidades entre 8 y 25 m, se encuentran únicamente en los extremos sur y norte, y se generan por refracción del oleaje dominante sobre los extremos del Banco. Quizas entonces, se deba también a un proceso de refracción durante la transgresión Holocénica, lo que haya dado lugar a que los arrecifes profundos de Cozumel se desarrollaran sobre el extremo sur de la isla. Ahora bien, asumiendo que este esquema de formación de los arrecifes profundos de Cozumel es correcto, el proceso de degradación debe haberse iniciado hace varios miles de años, y conforme transcurre el tiempo y la superficie degradada aumenta, el proceso se acelera paulatinamente. Sin embargo, este proceso es aún lento, como atestigua el tamaño todavía relativamente grande de las estructuras arrecifales, las que no pueden haber sido mucho mayores en su momento de crecimiento más activo o habrían sido capaces de mantener la tasa de ascenso del nivel del mar y no quedar rezagadas. Es probable que la lentitud del proceso se deba a que aún cuando los corales escleractíneos se restringen a las áreas más iluminadas de la estructura arrecifal, su desarrollo es aún lo suficientemente activo como para compensar en parte la degradación de la estructura. Esto seguramente se debe en gran medida al flujo de la corriente de Yucatán que manteniendo una alta dinámica, favorece un sano crecimiento en los corales, cuyo desarrollo es básicamente limitado por la abundancia de ambientes de penumbra y sombra, y no por efecto del sedimento libre sobre la plataforma insular (Hubbard y Pockock, 1972; Hubbard, 1986), o por la falta de movimiento de agua (Hubbard, 1974; Jokiel, 1978). Lo que aunado a su localización profunda y a sotavento, limita la tasa de degradación de la estructura arrecifal a la velocidad de los procesos de bioerosión, sin que intervengan esfuerzos mecánicos por efecto del oleaje de tormentas y ciclones, que seguramente acelerarían la tasa de destrucción de estas debilitadas estructuras. Esta hipótesis acerca del origen de los arrecifes profundos de Cozumel, solo puede ser verificada con perforaciones y fechamientos de los diferentes estratos de acreción en esas formaciones arrecifales. Lo que es incuestionable, es que estos arrecifes se encuentran en un claro proceso de destrucción, aunque es tranquilizante reconocer que este proceso -si no es acelerado por actividades humanas- aún tiene que transcurrir por algunos cientos o miles de años, antes de minimizar el incomparable paisaje submarino que ofrecen los arrecifes profundos de Cozumel. AgradecimientosEste estudio implicó operaciones de buceo difíciles, prolongadas y a veces un tanto arriesgadas, y no hubiera sido posible llevarlo a cabo sin la entusiasta y comprometida cooperación de los integrantes del grupo de buceo que realizó el muestreo: Eduardo Martín Chávez, Alejandro González de la Parra, Martín Sánchez Segura y Luz María Guzmán Fernández. Mi reconocimiento a Robert Ginsburg, Patricia Briones y Eduardo Martín por sus comentarios y discusiones sobre este trabajo. A Francisco Escobar de la Llata por la elaboración del material gráfico y a todos los compañeros que laboran en la Estación Puerto Morelos, que ayudaron de una u otra forma. Muchas otras personas, empresas e instituciones facilitaron la realización de este trabajo, en particular quiero destacar al Director del Parque Ecológico Chankanaab que nos facilitó el alojamiento en la isla de Cozumel. Al Club Naútico de Cozumel, quien amablemente nos facilitó su Marina para nuestras embarcaciones. A la compañía Caribbean Divers, quien nos ayudó en muchas cuestiones logísticas durante el desarrollo de este estudio. A la Coordinación de la Reserva Arrecifes de Cozumel, SEDUE. Este estudio fue realizado con apoyo del Conacyt otorgado al autor a través del proyecto PCCBBNA-021928. LITERATURAAGUAYO, J.E., R.BELLO, M del VECHIOJ.ARUJO y M.BASAÑEEstudio sedimentológico en el área Tulum-Cancún Isla Mujeres, estado de Quintana Roo,México. Informe de la Subdirección de Tecnología de Exploración del IMP197833 BAK, R.P. M., Coral Reefs and their zonation in the Netherlands Antilles. Reefs and related carbonates Ecology and Sedimentology-. Stud. Geol. 4. American Ass. Petrol. Geol. In Frost, Weiss and Saunders (Eds.): 1977 BEHETY, P.A., Nuevos reportes de gorgonaceos (Coelenterata para Cuba. Acad. Ciencias Cuba Ser. Oceanológica No.3319759 BLOOM A.L., W.S. BROEKER, J. CHAPPELL, R.K. MATTHEWS y K.J. MESOLEL Quaternary sea level fluctuations on a tectonic coast: new Th-230/U-234 dates from New Guinca.Quat.Res., 1974185-205 4 BRADBURY, R.H., Y. LOYA, R.E. REICHELT y W.T.WILLIAMSPattems in the structural typology of bentic communities on two coral reefs of the Central Great Barrier Reef. Coral reefs, 1986161-168 4 DAHL, A.L Marine Biology, Surface arca in ecological analysis: Quantification of Benthic Coral Reef algac.1973239-249 23 DAVIES, P.J., Modern Reef Growth-great Barrier Reef. Proc. Third Int. Coral Reef Symp.,1977325-330 DAVIES, P.J. y L.MONTAGGIONIReef growth and sea-level change: The environmental signature. Proc. Fifth Int. Coral Reef Symp.,1985477-511 3 DILLON, W.P. and J.G. VEDDER Bull.Geol.Soc.Am.Structure and Development of the Continental Margin of British Honduras. 19732713-2732 84 DUNSTAN,P., Growth and form in the reef-building coral Montastrea annularis. Mar. Biol. 1975101-107 33 GEISTER, J., The influence of wave exposure on the ecological zonation of caribbean coral reefs. Proc. Third Int.Coral Reef Symp.,197723-29 1 GINSBURG R.N. and D.CHOI., Quaternary reefs of Belize, Central America. proc. Great Barrier Reef Conf.James Cook Univ. Press. Townsville. 1983 GLADFELTER, E.H., Metabolism, calcification and carbon production: Organisms-level studies. Environmental signature.Proc.Fifth Int.Coral Reef Symp., 1985527-539 3 GOLDBERG, W.M., Bull.Mar.Sci. The ecology of the coral-octocoral communities off the southeast Florida coast: Geomorphology, species composition and zonation. 1973465-48823 3 GOREAU, T.F., Calcium carbonate deposition by coralline algae and corals in relation to their role as reef builders. Ann. New York. Acad. Sci., 196367-90 109 GOREAU,T.F., and J.W.WELLS, Bull Mar.SciThe shallow Water Scleractinia of Jamaica.Revised List of Species and their Vertical Distribution Range.1967442-45317 2 HAJOVSKY, Mapa turístico de los Arrecifes de Cozumel. 1974 HOPLEY,D., The geomorphology of the Great Barrier Real. Wiley and Sons.,Inc. 1982453 HUBBARD, J.A.E.B., Scleractinian coral behaviour in a calibrated current experiment: an index to their distribution pattems. Proc. Fourth Int.Coral Reef Symp.,1974107-126 2 HUBBARD,J.A.E.B., and YA. POCKOCK Sediment rejection by recent scleractinian corals: A key to paleoenvironmental reconstruction. Geol. Rundsch.,1972598-626 61 HUBBARD, D.K, Coral Reef Sediment as a control of reef development: St. Croix, U.S.V.I. 1986117-1265 3 JAMES, N.P. and R.N. GINSBURG The Seaward Margin of Belize Barrier and Atoll Reefs. Int. Assoc. of Sedimentologists. Blackwell Scientific Publ. Special Publ. 3.1979191 JOKIEL, P.L., Effects of water motion on reef corals. Jour.Exp.Mar.Biol. Ecol., 197887-97 35 JORDAN, E., An.Centro Cienc. del Mar y Limnol., Univ. Nal. Autónom. México Estructura y Composición de Arrecifes Coralinos en la Región Noreste de la Península de Yucatán, México.197969-866 1 JORDAN,E., Arrecifes Coralinos del Noreste de la Península de Yucatán: Estructura Comunitaria, un Estimador del Desarrollo Arrecifal. Ph. D. Dissertation. Univ. Nal. Autónom. México., 1980148 JORDAN, E., Informe final del proyecto "Arrecifes coralinos del Caribe Mexicano: Su potencial de uso". CONACyT-ICMI,UNAM1987 JORDAN, E., (En prensa). Gorgonian community structure and Reef zonation patterns on Yucatan Coral Reefs. Bulletin of Marine Science. JORDAN, E., M.MERINO, O.MORENO, and E. MARTIN Community Structure of Coral Reefis in the Mexican Caribbean.Proc.Fourth International Coral Reef Symposium,Manila,Phillipines.303-308. 2 JORDAN, E., and E.MARTIN, Atoll Res. Bulletin.Chinchorro: Morphology and composition of a Caribbean Atoll. 1987a KINZIE,R.A., Bull.Mar.Sci., The Zonation of West Indian Gorgonians.197393-15523 1 KINZIE, R and P.SNIDER, Coral reefs: research methods. UNESCO. Monographs on Oceanographic Methodology A simulation study of coral reef survey methods In: D.Stoddart & R.Johannes (Eds.). 1978231-250 LASKER,H.R., and M.A.COFFROTHMar. Ecol. Prog. Ser., Octocoral Distributions at Carric Bow Cay, Belize.198321-28 13 LEWBEL G.S. y L.R. MARTIN, Diving and snorkeling guide to Cozumel. Pisces Book Co. New York.1984 LIGHTLY,R.G.,I.G.MACINTYRE y R.STUCKENRATH Acropora palmata reef framework: a reliable indicator of sea level in the Western atlantic for the past 10,000 years. Coral reefs, 1982125-130 1 LOGAN, B.W., Coral reefs and Banks, Yucatan Shelf Mexico. In: Logan,Harding,Alan Williams y Snead (Eds.). Mem.ASSOC. Petrol. Geol.1969129-196 11 LOYA, Y., Bull. Mar. Ser., Effects of water turbidity and sedimentation on the community structure of Puerto Rican corals. 1976450-46626 4 LOYA, Y., Methodology, 5. Coral reefs: research methods. UNESCO Monographs on Oceanographic In: D.Stoddart & R.Johannes (Eds.). Plotless and transect methods. Pp.197-218 MACINTYREJ.G., Am. Assoc. Petrol. Geol. Bull.Submerged reefs of Eastern Caribbean1972720-738 MACINTYRE, I.G., R.B.BURKE, and R. STUCKENRATH, Geology, Thickest recorded Holocene reef section, Isla Perez core hole, Alacran reef, Mexico.1977749-754 5 MAY, R.M., Stability and Complexity in Model Ecosystems. Monographs in Population Biology, 6. Princeton Univ.Press.1974 MERINO,M., An. Inst. Cienc. del Mar y Limnol, Univ. Nal. Autón. México, Aspectos de la Circulación Costera del Caribe Mexicano con base en Observaciones Utilizando Tarjetas de Deriva.198631-4613 2 MUSCATINE L y J.W.PORTER Bioscience,Reefs Corals: Mutualistic Symbiosis, adapted to nutrient poor environments. 1977454-46027 7 MUSCATINE L., L.R. McCLOSKEY y R.E. MARIAN Limnol. and Oceanogr Estimating the daily contribution of carbon from zooxanthellae to animal respiration. 1981601-611 26 MUZIKK. rrie Bow Cay, Belize, I. Structure and Communities Octocorallia (Cnidaria)from. Carric Bow Cay, Belize. The Atlantic Barrier Reef Ecosystem at Ca In: Rutzler K., and I.G. Macintyre. (ed.) 1982303-310 NEUMANN, A.C., Quaternaria Quaternary Sea level data from Bermuda. 197141-43 15 NEUMANN, A.C.,AND I.G. MACINTYRE Reef response to sea level rise: Keep up, Catch-up or Give-up. Proc. Fifth Int.Coral Reef Symp.,1985105-110 1 NEWELL, N.D. y J.K.RIGBY, Geological studies on the Great Bahama Bank In: Regional Aspects of Carbonate Deposition. Spec. Publ. Soc. Econ. Paleont. Mineral 195715-79 5 OGDEN J.C. y E.H. GLADFELTER Coral reefs, seagrass beds and mangroves: Their interaction on the coastal zones of the Caribbean. UNESCO Reports in Marine Science198323 PORTER, J.M., Ecology, Patterns of Species Diversity in Caribbean Coral reefs. 1972668-67353 4 REISWIG, II. M., Bulletin of Marine SciencePopulation dynamics of three Jamaican Demospongiac.1973191-226 23 PURDY, G.E., Bull. Am. Ass. Petrol. Geol.Karst-Determined Facies Patterns in British Honduras: Holocene Carbonate Sedimentation Model. 1974825-855 58 ROBERTS, H.H., Variability of reefs in regard to vhanges in wave power around an island. Proc. Second Int.Coral Reef Symp1974497-512 2 RUTZLER, K Oceología The role of burrowing sponges in Bioerosion. 1975203-216 19 SCHROEDER, J.H. y H. ZANKL., Dynamic Reef formation: a sedimentological concept based on studies of Recent Bermuda and Bahamas Reefs. Proc. Fourth Int.Coral Reef Symp., 1974413-428 2 SHINN, E. Spur and Groove formation on the Florida reef tract. J.Sed.Petrol., 1963291-303 33 SHINN, E.A., J.H. HUDSON, R.B.HALLEY, B. LIDZ, D.M. ROBBIN and I.G. MACINTYREGeology and sediment aecumulation rates at Carric Bow Cay, Belize. The Atlantie Barrier Reef Ecosystem at Carrie Bow Cay, Belize, I.63-76. In: Rutzler and Macintyre (eds)., Smithsonian Inst. Pres. 1982 STODDART, D.R., Ecology and morphology of recent coral reefs. Biol. Rev., 1969433-498 44 STODDART, D.R., Post-Huricane Changes on The British Honduras Reefs: Re- survey, 19721974 Cameron, A. M., V.M. Campbeil, A.B. Cribb, R. Endean, J. S. Jell, O.A.Jones, P. Mather, and F.H. Talbot, (ed). Proc.Second International Coral Reef Symposium, 2:473-483. Cameron, A. M., V.M. Campbeil, A.B. Cribb, R. Endean, J. S. Jell, O.A.Jones, P. Mather, and F.H. Talbot, (ed). Brisbane Australia: The Great Barrier Reef Committee. TRENCH, R.K, The psicology and biochemistry of zooxanthellae symbiotic with marine coelenterates. I. The assimilation of photosynthetic products of zooxanthellae by two marine cocienterates. Proc.R.Soc.Lond1971225-235 177 TUNNICLIFFE, V.J. Disseration. Biological and Physical processes affecting the survival of a stony coral, Acropora cervicomis. Yale University, New Have, USA1980 UCHUPI,E., Am. Assoc. Petrol. Geol. Bull., Eastem Yucatan continental margin and Western Caribbean Tectonies. 19731075-1085 57 WELLS, J.W., Scleractinia. Treatise on Invertebrate Paleontology. Part F. F328-F444. In: Moore, R.C. (ed.) Geol.Soc.America and Univ.Kansas Press. 1956 WUST,G., Stratification and Circulation in The Antillean-Caribbean Basins.. Columbia University Press, New York 1964
|