|
SIZE, WEIGHT AND CALORIC CONTENT
ESTIMATES OF Calocaris mecandreae (CRUSTACEA: DECAPODA) FROM CHELAE BIOMETRY
FOR STUDIES ON FISH ECOLOGY
Trabajo recibido el 22 de agosto de 1986 y
aceptado para su publicación el 5 de diciembre de 1986.
J.A. Calderón P.
Instituto de Ciencias del Mar y Limnología, Estación
Mazatlán, Universidad Nacional Autónoma de México.
Contribución No. 623 del Instituto de Ciencias del Mar y
Limnología, UNAM.
Se hizo un estudio de la
relación entre las medidas de los quelípedos y el tamaño
del camarón excavador Calocaris macandreae de una
población del Mar de Irlanda al Oeste de la Isla de Man. La linearidad
de la relación parece estar alterada debido a la alta incidencia de
quelas regeneradas. Sin embargo, es posible obtener una estimación
adecuada del tamaño al usar una combinación de las medidas de la
quela, principalmente la proporción entre las longitudes de propodio y
palma, con referencia a esta última, dado que las primeras etapas del
proceso de regeneración no son isométricas. Se hicieron
ecuaciones de longitud del caparazón contra peso para cada una de las
estaciones del año, ya se encontraron diferencias significativas en peso
entre el verano y el invierno. El contenido calórico varió entre
individuos inmaduros y maduros con ovarios parcial o totalmente desarrollados
de 20.1 a 23.1 Kj/g peso seco sin ceniza, respectivamente. El contenido de
cenizas fue más alto en el primer grupo (49.1%) que en el segundo
(43.5%). Se expone un ejemplo práctico para el uso de esta
información en el análisis del contenido estomacal en
peces.
A study on the relationship between
cheliped measurements and size of the fossorial shrimp Calocaris
macandreae from an Irish Sea population off the west coast of the
Isle of Man was carried out. The linearity of the relationship appears to be
disrupted by the high occurrence of regenerated chela. Nevertheless, an
adequate estimate of size is feasible by using a combination of chelae
measurements, mainly the ratio between propodium. and Palm lenghts, with
reference to the latter, since the earlier stages of the regeneration process
are not isometric. Carapace length versus dry weight equations for each of the
seasons were made as significant differences in weight between Summer and
Winter samples were found. Caloric content was lower in immature individuals
than in those having half of fully developed overies, 20.1 and 32.1 Kj/g
ash-free dry weight, respectively. Ash content has higher in the former group
(49.1%) than in the latter (43.5%). A practical example for the use of this
information in the analysis of fish stomach content is given.
The importance of Calcaris mecandreae Bell is mainly ecological. It is a substantial source of food for some demersal fishes such as cod (Gadus morhua), pouting (Trisopterus luscus) poor cod (Trisopterus minutus) and other gadoids (Amstrong, 1980; Buchanan, 1963; Elmhirst, 1912) and the commercially important prawn Nephrops norvegicus (Oakley, 1978). In dealing with Calocaris found in the stomach content of a fish under study, one may encounter that measurements of those specimens are difficult if not impossible, because their soft exoskeleton is demaged due to the digestive process. Frequently, the only parts which remain relatively intact are the chelipeds, providing that they are not from a recently moulted individual. If a relationship between any cheliped lengths and the size of the animal can be found, weight and caloric values could be estimated more accurately once the appropriate relationships are determined. The aim of this paper is to provide that information which could be used in finding out the contribution ofC. macandreae to the diet of a fish population under study, in terms of weight and energy. MATERIAL AND METHODSSpecimens of Calocaris macandreae Bell were collected between December 13, 1977 and August 15, 1980 from an Irish Sea area located at approximately 17 Km to the Northwest of Bradda Head, Isle of Man (54°12' N, 4°50'W) on board the 20 in research trawler "Cuma" of the Department of Marine Biology of the University of Liverpool. The sampling gear included two mm mesh, 2.5 in beam trawls, and a 0.5 m² modified Baird grab (Baird, 1959; Buchanan, 1963). For the preservation of samples, 10% neutral buffered saline formalin was used. Length measurements to the nearest 0.1 min. were carried out by means of a calibrated Wild stereomicroscope at a 6X magnification; focal variation was mantained to a minimum by focusing first at a higher magnification. The definition of carapace length given by Buchanan (1963) was used, i.e. from the posterior lateral margin of the carapace to the anterior margin located at the base of the second antenna (Fig. 1A). The relationship between carapace and total length is defined by the equation: total length = 2.5 + 2.8 carapace length (Calderón-Pérez, 1981). When present, the chelipeds were removed and two length measurements (on each of them) were carried out: Propodium length (PRL), from the tip of the fixed finger to the posterior margin on the dactylopodite side (Fig. 1B) and Palm length (PL) from the tip of the protuberance located near the propodium-dactylopodite articulation to the posterior margin of the propodium on the fixed finger side (Fig. 1C). Whole specimens and individual chelae were dried in an oven for 24 hr and weighed in Mettler analytical balance to the nearest 1 mg. The relationship between wet and dry weight is given by the equation. D = 0.247 W - 2.0105 Where D = Dry weight and W = Wet weight (Calderón-Pérez, 1981). 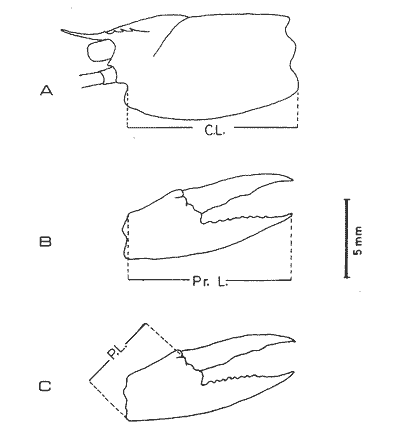 Figure 1. Length measurements taken on Calocaris macandreae. A, Carapace length, Propodium length; C, Palm length. Ash content was determined in 200 specimens placed individually, in pre-weighed porcelain containers, in a muffle furnace at a temperature of 550°C for a period of at least 12 hours. The samples were then transfered to a dessicator to cool down for 2 hours and weighed in the analytical balance. The caloric value of Calcaris mecandreae was determined by means of a Gallenkamp ballistic bomb calorimeter calibrated with benzoic acid. Since the amount of matter required for one determination exceeded that of one individual, a total of 195 specimens were pooled into four different groups, using as the separation criterion the size of the ova. Five replicated were made in three of the groups and six in one of them. RESULTSIn figure 2A the relationship between carapace and mean propodium lengths is shown, in figure (2B) that of C.L. and mean palm length is displayed and in figure (2Q) carapace length is plotted against mean cheliped weight. The high standard deviation values from the means, and the overlapping of the ranges, indicate that the distribution of the cheliped length values for each interval of C.L. is not normal and the mean is of little use to find a relevant relationship. 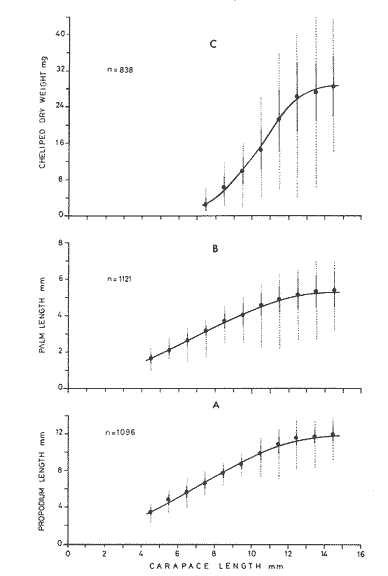 Figure 2. Relationship between carapace length and the three chela measurements: A, mean propodium length; B, mean palm length abd c, mean dry weight. Vertical continuous lines indicates standard deviation whilst the superimposed broken lines represent the range. In figure (3) propodim and palm length frequency distributions for such C.L. intervals show several modes, particularly among the bigger size classes which explain the high values of the dispersion parameters. An explanation for these non-normal distributions of chelae length and weight seems to lie in the fact thatC. macandreae has an autotomy-prone nature and most of the chelae examined were at different stages of regeneration. Considering that a given cheliped length value is common to several C.L. intervals, an attempt to infere an accurate carapace length value from either of the two chela length measurements would be virtually impossible. On the other hand, both measurements may be used to solve the problem as it seems that the regeneration process in C. macandreae is nor isometric, i.e., the propodium length increases factor than the palm length, thus the ratio propodium/palm lengths between normal chelae and those at different stages of regeneration is different. A normal chela i.e. on the extreme right of the distribution, has a ratio of less than 2.10 except for individuals smaller than 8.0 mm C.L., whereas regenerated chelae have ratios between 2.10 and 3.23. Observed ratio figures from matching propodium and palm modes are set out on Table 1 for the carapace length intervals of 1 mm between 5.0 and 15.0 mm. The relationship between C.L. in mm, and dry weight in mg is given by the equation. 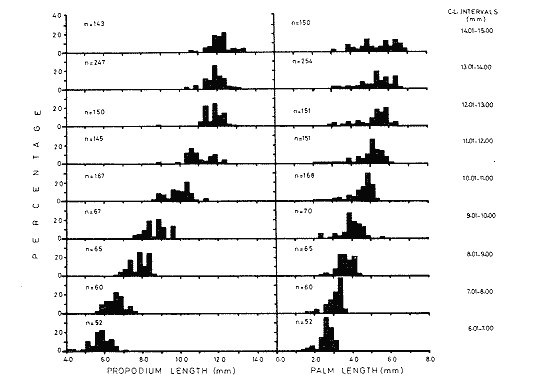 Figure 3. Frequency distributions of propodium and palm lengths for each interval of carapace length. In dry weight = 3.117; In carapace length -2.74 (Eq. 1). This equation was made Considering the results of weight from all seasons although there are significant differences in weight between Summer and Winter as displayed in Table (2) where Student (t) tests showed that the differences of mean values in all but three intervals of carapace length were significant. Thus it is pertinent to give the appropriate equation for each of the seasons: Winter: In dry weight = 3.073; In carapace length 2.5490 (Eq. 2); Spring: In dry weight = 2.8149; In carapace length 1.8305 (Eq. 3); Summer: In dry weight = 2.978; In carapace length -2.1496 (Eq. 4); Autumn: In dry weight = 3.1750; In carapace length -2.7520 (Eq. 5). Observed and predicted dry weight values from equations for Summer and Winter are shown in Tables 3 and 4 respectively. Confidence interval at the 95% level is also given. Conversion of weight values into energy units can be made by using Table 5 in which the caloric content of four different groups of individuals is displayed. The ash content of the same groups is set out on Table 6. 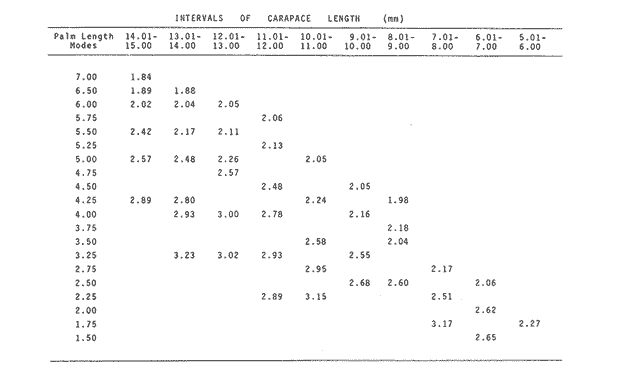 TABLE 1 RATIO OF MATCHING PROPODIUM AND PALM MODES HAVING PALM LENGTH AS REFERENCE FOR EACH INTERVAL OF CARAPACE LENGTH 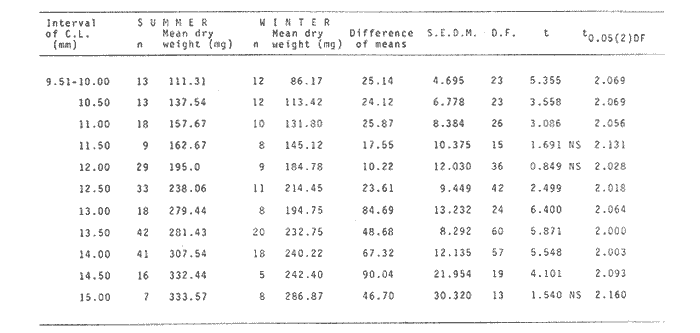 TABLE 2 STUDENT (t) TEST FOR DIFFERENCE OF MEAN WEIGHTS PER INTERVAL OF CARAPACE LENGTH BETWEEN SUMMER AND WINTER, ABBREVIATIONS: S.E.D.M., STANDARD ERROR OF DIFFERENCE OF MEANS; D.F., DREGREES OF FREEDOM; N.S., NON-SIGNIFICANT DISCUSSIONInterpolation of size and the consequent values of weight and caloric content from chelae measurement is not a straight forward operation as originally proposed. Besides natural variation within each size class, the high incidence of regenerated chelae following autotomy, disrupts the linearity of the relationship which is only evident when palm length modes of extreme right ratios are considered against the corresponding intervals of carapace length, both displayed in Table 1. The use of the ratio propodium/palm length is therefore essential to discriminate normal from regenerated chelae. 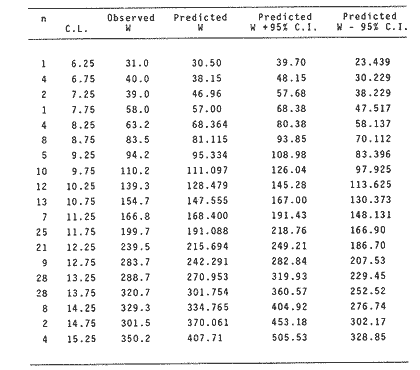 TABLE 3 COMPARISON BETWEEN PREDICTED WEIGHT VALUES USING THE EQUATION IN W = 2.9066 LN C.L. - 1.9087 AND 013 SERVED SUMMER MEAN WEIGHT VALUES FOR 19 INTERVALS OF CARAPACE LENGTH IN Calocaris macandreae. C.L. CONSIDERED IS THE MID POINT OF THE CLASS INTERVAL. C.I., CONFIDENCE INTERVAL 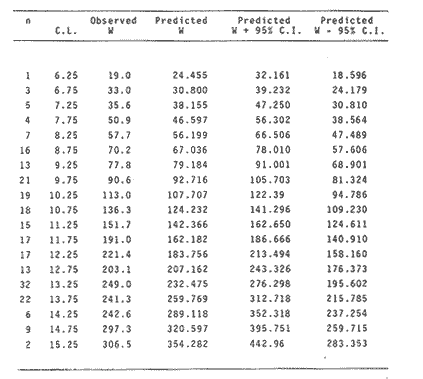 TABLE 4 COMPARISON BETWEEN PREDICTED DRY WEIGHT VALUES USING THE EQUATION IN W = 2.9969 LN L.C. - 2.2952 AND 013SERVED WINTER MEAN WEIGHT VALUES FOR 19 INTERVALS OF CARAPACE LENGTH I IN Calocaris macandreae. C.L. CONSIDERED IS THE MID POINT OF THE CLASS INTERVAL. C.I., CONFIDENCE INTERVAL The increase in ratio towards the first stages of regeneration suggest that C. macandreae requires a longer though slender chela rather than an isometric but shorter one. This might be related to its burrowing habits which involves the use of both chelipeds (Buchanan, 1963; Elmhirst, 1912; Rünnstrom, 1925) and for that purpose, the sooner both chelae attain symmetry the more efficient the digging and burrow improvements will be Most of the intervals of carapace length, especially those in the range 12.0 to 15.0 mm show some degree of overlapping in weight, as evidenced in Tables 3 and 4 by the confidence intervals, although a general pattern of increase with size is followed when mean values are considered. Student (t) test showed that it is more convenient to use the appropriate equation for each season rather than the overall equation since weight differences between Summer and Winter are significant in most cases. An important contribution to the body weight of Calcaris is given by the ovary which in the Summer and Autumn months may increased more than 50 mg, apart from the normal body weight gain due to the increased food availability typical of these months in the water column by plankton production and in the benthos as detritus derived from the former (meadows and Cambell, 1978; Pomeroy, 1980). In the Winter and early Spring months, ovaries are spent in addition to the food depletion which accounts for the lower mean weights in all but three intervals of C.L. The caloric content range of 20.16 - 23.1 Kj/g ash-free dry weight in C. macandreae is similar to values reported for other benthic crustaceans such as Penaeus indicus (Read and Caulton, 1980) in which a figure of 24.0 Kj/g ash-free dry weight is given; juvenile Macrobrachium rosenbergii; 20.06 Kj/g ash-free dry weight (Nelson et al. , 1977) and the mean value of four entomostraceans, Artemia sp, Leptodora kindti, Tigriopus californicus and Calanus helgolandicus, 24.4 Kj/g ashfree dry weight (Slobodkin and Richman, 1961). 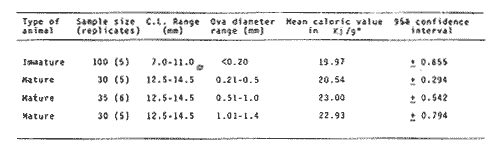 TABLE 5 MEAN CALORIC VALUES OF FOUR DIFFERENT TYPES OF Calocaris INDIVIDUALS 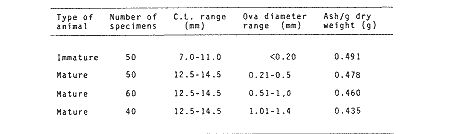 TABLE 6 ASH CONTENT VALUES OF FOUR DIFFERENT TYPES OF Calocaris INDIVIDUALS The separation of groups of individuals based on ova size as an index of ovary weight seems justified for the ash content analysis although for caloric content it does not seem to be the case since both groups of ova size 0.51 to 1.00 min and 1.01 to 1.40 produced similar results; in fact the value of the former was slightly higher than that of the latter. This results was not totally unexpected for the energy required for vitellogenesis must be stored in the hepatopancreas whilst the ova are underdeveloped. Lawrence et al. (1979) have shown that during the ovary development in Penaeus spp there is a transfer of material; mostly lipids, from the hepatopancreas to the ovary; similarly. Adiyodi and Adiyodi (1971) found a large in the ovary simultaneous to a substantial decrease of lipids, in the hepatopancreas of the crabParathelphusa hydrodromous. It is obvious that for the researcher regarding Calocaris macandreae as a food item on the stomach content of a fish, the above considerations are pointless since he will not have the elements to decide which value should be applied except for the fact that individuals having ovaries with ova of size 0.51 to 1.40 min are more frequently found in the Spring and Summer months; i.e., from sixty to eighty percent in the samples, whereas in the Winter months their proportion varies from thirty to fifty percent. In the Autumn months the proportion of those individuals is low (less than 30 percent) although the proportion of berried animals could be higher than 60 percent (Calderón-Pérez, 1981), thus the proportion of individuals with a high caloric content is about 90 percent. In spite of the problems to infere size from chelae measurement, a reasonable estimate is still possible in most cases. Let us consider a practical example; both chelipeds of C. macandreae were its only intact parts found in the stomach content of a cod sampled in August; palm and propodium length were 5.0 mm. and 12.3 mm respectively in both chelae. From figure 3 and Table I it will be noticed that this palm length is common to four intervals of carapace length in the range 10.01 15.0 mm, the ratio propodium/palm however, clearly indicates that these chelae belong to the interval 13.01 14.0 mm; 2.46 compared to 2.48. Taking a mean value of 13.5 min of C.L., weight can now be estimated by means of equations 4 which correspond to the Summer months. The result is 0.286 g of dry weight, to express this figure in energy units, ash content has to be substracted from that amount (45%) remaining 0.1573 g of ash-free dry weight which is then multiplied by the caloric equivalent of either group of mature individuals having ova of size 0.51 to 1.00 min or 1.01 to 1.40 (23 Kj/g ash-free dry weight). The results is 3.6 Kj. The same procedure can be applied to all chelae except in those having a palm length of 6.0 mm which in Table 1 correspond to three intervals of C.L. and the ratio is practically the same. In this particular case, the class interval with the highest frequency of individuals will be the best choice since its probabilities are higher. In the Irish Sea, that interval of C.L. was 13.01 - 14.00 mm (Calderón-Pérez, 1981); else where, however, it might be slightly different (Buchanan, 1963; Rünnstrom, 1925). AgradecimientosThis study was carried out at the Department of Marine Biology of the University of Liverpool, Port Erin, Isle of Man. I want to express my gratitude to E. Naylor for providing laboratory facilities; to B. Bowers for his advice, encouragement and critical reviews of previous drafts of the manuscript; to the skipper and Crew of the R.V. "Cuma" for their help at sea and to all members of the staff who helped me in different ways. My thanks are also due to A. Ayala-Castañares for his support and encouragement and to E. Macías Regalado for his disinterested help and for the translation of Rünnstrom's Paper. Finantial support was provided by the British Council and the National Council of Science and Technology of Mexico (Consejo Nacional de Ciencia y Tecnología). LITERATURAADIYODI, R.G. and K.G. ADIYODI Indian J. Exp. Biol. Lipid metabolism in relation to reproduction and moulting in the crab. Parathelphusa hydrodromous (Herbst): Cholesterol and unsaturated fatty acids.1971. 514 -515. 9: ARMSTRONG, M.J. Ph. D.Thesis The feeding ecology of a demersal fish community over a muddy substrate off the west coast of the Isle of Man. University of Liverpool.1980. BAIRD, R.H. Shellfish Committee Bo. A preliminary account of a new half square metre bottom sampler. I.C.E.S., C.M. 1958 1958.70 BUCHANAN, J.B. J. Mar. Biol. Ass. U.K The ecology of Calocaris macandreae (Crustacea: Thalassinidea).1963.729-747. 43 CALDERON-PEREZ, J.A. Ph. D.ThesisSome aspects of the biology of Calocaris mecandreae Bell (Crustacea: Decapoda, Thalassinidea) in Isle of Man waters. University of Liverpool1981237. ELMHIRST, R. Report of the Scottish Marine Biological Association 1935.1934-1935. LAWRENCE, A.L., D. WARD, S. MISSLER, A. BROWN, J. McVEY and B.S. MIDDLEDITCH Proc. World Maricul. Soc. Organ indices and biochemical levels of oca from penaeid shrimp mantained in captivity versus those captured in the wild. 1979.453-463. 10: MEADOWS, P.S. and J.I. CAMPBELL An Introduction to Marine Science. Blackie and Sons Ltd.London1978. 176 p. NELSON, S.G., H.W. LI and A.W. KNIGHT Comp. Biochem. Physiol. Caloric, carbon and nitrogen metabolism of juvenile Macrobrachium rosenbergii (De Man) (Crustacea: Palaemonidae) with regard to trophic position.1977.319-327.58. POMEROY, L.RFundamentals of Aquatic Ecosystems. Detritus and its role as a food source. In: Barnes, R.S.K. and K.H. Mann (Eds.). Blackwell Scientific PublicationsOxford1980.84-102. OAKLEY, S.G. Ph. D. Thesis. Food, feeding behaviour and some aspects of the ecology of Nephrops norvegicus in the Irish Sea.University of Liverpool.1978. READ, G.H.L. and M.S. CAULTON Comp. Biochem. Physiol.Changes in mass and chemical composition during the moult cycle and overian development in inmature and mature Penaeus indicus Milne Edwards. 1980.431-437. 664: RUNNSTROM, S. Bergens Mus. Shr. , ME bdBeitrag zur Kenntnis einiger hermaphroditischen decapoden Crustaceen. 1925.1-115.3 (2) SLOBODKING, L.B. and S. RICHMAN Nature Calories/gm in species of animals. 1961299. 191
|