|
DETERMINACIÓN DE LOS ELEMENTOS DEL BALANCE
ENERGÉTICO EN EL PESCADO BLANCO ADULTO, Chirostoma estor J. (PISCES,
ATHERINIDAE)
Trabajo recibido el 27 de mayo de 1986 aceptado para su
publicación el 27 de enero de 1987.
S. Espina
J. Latoumerie
A. Sánchez
M. E. Vega
Laboratorio de Ecofisiología, Depto.
Biología, Fac. Ciencias, Universidad Nacional Autónoma de
México.
Los especímenes adultos de Chirostoma
estor (N= 70) se colectaron en el mes de Enero en el Lago de
Pátzcuaro. Estos organismos se obtuvieron por desove artificial y se
mantuvieron en corrales de malla donde se les proporcionó, como dicta
suplementaria, alimento balanceado para peces. El objetivo del trabajo fue
medir cada uno de los elementos del balance energético para estimar el
campo de crecimiento de la especie, representados en la ecuación: C = P
+ R + F + U. Los símbolos representan el consumo de alimento, el campo
de crecimiento y producción, la tasa respiratoria y las pérdidas
energéticas en heces y productos nitrogenados. Los resultados muestran
que los peces ingirieron del alimento al 2.3 % de su peso corporal y de la
energía incorporada, asimilaron alrededor del 90%; de esta
fracción el 75% correspondió a crecimiento. El 15% de la
energía incorporada se utilizó en procesos metabólicos. El
consumo de O2 se relacionó con el peso corporal,
como es usual en peces: R (µl O2/h = 170
W0.91. El alto valor del campo de crecimiento,
caracteriza a C. estor como un eficiente convertidor de
energía. Sin embargo, el crecimiento en peso fue reducido, lo que se
puede atribuir a la baja temperatura del lago en esta época del
año (14 °C) y a las condiciones de cautiverio. Se discuten los
resultados en cuento a su incidencia en el cultivo de la especie.
Chirostoma estor were collected in
January from net confinement at Pátzcuaro Lake, Michoacán;
México, where fishes were kept after hand spawning and maintained with
manufactured food. Laboratory work aimed to measure the elments of the
energetic balance equation: C = P + R + F + U; in order to determine the specie
scope for growth. The symbols represent food consumption, production,
respiratory metabolism and losses in faeces and nitrogen products,
respectively. Results showed fishes fed at 2.3% corporal weight and from this
they assimilated 90%. Oxygen uptake for metabolic processes was 15% of energy
incorporated. Gas consumtion dependence from body weight was R = 170
W0.91 (µl O2/h). Fishes
scope for growth was high, up to 75%. Thus C. estor is
considered as an efficient energy converter. However increase weight is slow
what is probably due to winter temperature (14 °C) and confinement
conditions. Results are discussed in reference to aquacultural practices
improvement.
En el Lago de Pátzcuaro, Michoacán, coexisten varias especies de aterínidos del género Chirostoma, como C. bapioni, C. grandocule, C. patzcuaro y C. estor. Este último, denominado pescado blanco, destaca por su importancia comercial. En las últimas décadas ha disminuido considerablemente la población de pescado blanco en el lago, probablemente por el cambio en las condiciones del vaso y a la intensa captura a que se ha sometido a la especie (Ancona et al., 1940; Henderson, 1974). Esta situación ha motivado el estudio exhaustivo de la biología del recurso con objeto de racionalizar la captura e iniciar prácticas acuaculturales (Solórzano, 1955, 1963; Rosas, 1970; Lara, 1974). Recientemente, se han efectuado estudios con el fin de conocer los requerimientos energéticos del pescado blanco y poder predecir el campo de crecimiento, cuando se alimenta con dicta manufacturada comercial (Espina et al., 1980; Vega, 1982) ya que la mayoría de los trabajos realizados con enfoque bioenergético, se han efectuado en peces carnívoros mantenidos con alimento fresco (Beamish, 1964; Brett, 1976; Niimi y Beamish, 1974; Mercy-Gerald, 1976) y en la carpa herbívora, Cteinopharygodon idella (Fisher, 1970, 1972a, b; Fisher y Lyakhnovich, 1973). En relación a los hábitos alimenticios, Chirostoma estor es omnívoro y tolera el alimento manufacturado tanto en condiciones de laboratorio, como en los encierros en los que se mantienen los peces en cultivo en el Lago de Pátzcuaro (Vega et al., 1981). Así, si se conoce el valor de la dieta y la cantidad ingerida por el organismo, es posible predecir el crecimiento en condiciones experimentales determinadas. Varios autores han propuesto modelos tendientes a simplificar la complejidad de los procesos inherentes al crecimiento en peces (Paloheimo y Dickie, 1956, 1966; Warren y Davis, 1967; Duncan y Klekowski, 1975); producto de dichos trabajos es la ecuación balanceada energéticamente: C = P + R + F + U Donde C es la energía contenida en el alimento consumido, R es el consumo de oxígeno, F es la energía perdida en las heces, U la pérdida en productos nitrogenados y P es producción o crecimiento. Todos los términos de la ecuación se refieren a tasas y se expresan en términos calóricos, esto es cal./día. Las investigaciones sobre el balance energético son especialmente importantes ya que dan cuenta de la incidencia en el crecimiento de la energía contenida en el alimento, como asimismo de los otros destinos de ésta en el cuerpo del animal objeto de estudio. Dada la relevancia que tiene esta especie autóctona, como recurso en el país, es conveniente llevar a cabo este tipo de investigaciones, para poder implementar técnicas acuaculturales nuevas o mejorar las existentes. Por lo anteriormente expuesto, es propósito de este trabajo, determinar los diferentes elementos que constituyen el balance energético en Chirostoma estor adulto, mantenido con alimento balanceado comercial. Asimismo, se pretende conocer la eficiencia de utilización de la energía contenida en dicho alimento. MATERIAL Y MÉTODOSLos organismos (N= 66) utilizados en este estudio, de 1.9 a 4.56 g de peso corporal fueron proporcionados en la Estación Piscícola de Pátzcuaro, donde se obtuvieron mediante desoye artificial. En el Lago de Pátzcuaro, los peces se mantuvieron en corrales de malla de 4 x 4 m y 2.2 m de profundidad, donde se les proporcionó alimento balanceado. La captura se efectuó en dichos corrales en el mes de Enero. La temperatura del agua tanto dentro como fuera del encierro, fue de 14 °C, el pH de 8.4 y el oxígeno disuelto 5.6 ppm. Estos factores del medio, se midieron con un electrodo mixto Ysi-54 ARC (±0.5C y ±0.1 ppm 02) y el pH con un potenciómetro Cole Palmer (±0.01). Los peces se trasladaron al laboratorio en un cilindro de 40 litros acondicionado con doble bolsa de polietileno. En la bolsa interna se dispusieron los peces con agua del medio, se insufló oxígeno y se cerró herméticamente; en la bolsa externa se colocó hielo con el fin de mantener la temperatura a 14 °C. Ya en el laboratorio, los peces se mantuvieron en acuarios de 60 litros con aireación constante y filtro químico. El agua se recambió parcialmente cada día, hasta que el pH alcanzó el valor de 7.5 que es el pH del agua desclorinizada del laboratorio. La temperatura alcanzó gradualmente 20 °C ± 1.0 °C. El fotoperoido se mantuvo en 12 horas de luz. El periodo de aclimatación fue de 20 días y durante este lapso, los organismos se mantuvieron con dieta balanceada comercial. El alimento se proporcionó al 3% del peso corporal, en dos raciones diarias. El periodo de alimentación se limitó a dos horas (Vega, 1982). La tasa de consumo de alimento se midió por método gravimétrico, esto es, por diferencia entre el peso seco del alimento suministrado y el peso seco del remanente en los acuarios el cual se colectó mediante sifón, se decantó y se secó a 60 °C en estufa Blue-M. El análisis del alimento reveló un contenido de 25.73% de proteínas, 6.12% de grasas y 50.81% de carbohidratos. El contenido calórico de la dieta fue de 5,048.2 calorías por gramo de alimento. Después de retirar el alimento de los acuarios y recambiar el agua, las heces producidas durante 24 horas, se obtuvieron por filtración; se secaron y se pesaron en balanza analítica. El valor calórico (3,854 cal./g de heces)se determinó con bomba calorimétrica Parr. La tasa de excreción nitrogenada se consideró como el 15% de la proteína usada por los organismos. La proteína usada se calculó por diferencia entre la proteína consumida y la depositada, que representa en la mayoría de los peces, el 16% de la proteína ingerida (Niimi y Beamish, 1974). Para transformar los datos, se utilizó el factor de 5.7 cal./mg. Este valor es un promedio entre el valor calórico del amoníaco y el de la urea, principales productos de la excreción nitrogenada en los peces ya que se conoce que el pescado blanco de Pátzcuaro, excreta urea en forma apreciable (Espina et al.,1980). La tasa de consumo de oxígeno de rutina se midió individualmente en respirómetro cerrado, en una submuestra de 16 animales. Se utilizaron cámaras de vidrio de 4 litros con agua desclorinizada a 20 °C y aireación constante. A cada cámara se había fijado una manguera de plástico para tomar las muestras de agua. Los animales permanecieron en los respirómetros 24 horas previas al experimento, con objeto de disminuir el "stress" por manipulación. En este lapso los peces estuvieron en ayunas, con el fin de evitar alteraciones en la tasa metabólica, debidas a la ingestión de alimento (Phillips, 1972). Al iniciar la medición se suspendió la aireación y se sellaron las cámaras con aceite vegetal; se tomaron muestras iniciales de agua en frascos de 125 ml y al cabo de una hora, se tomaron las muestras finales. En estas muestras se cuantificó la concentración de oxígeno y el consumo del gas se obtuvo por diferencia entre ambas muestras. Para trasformarlo en sus equivalentes calóricos, se utilizó el factor 3.36 cal/mg O2 (Brett, 1976). Al finalizar estos experimentos se sacrificaron los peces y se pesaron. De los mismos acuarios se tomó una muestra adicional de 12 individuos, en los que se determinó el estado de madurez gonádica, mediante técnicas histológicas. Como la frecuencia de los pesos en la muestra de peces capturados presentó una distribución trimodal, se agruparon los datos en clases-pesos. El balance energético se basó en la ecuación mencionada: C = P + R + F + U, cuyos símbolos para consumo, producción, respiración, evacuación de heces y excreción nitrogenada, corresponden a los aceptados por International Biological Program (Duncan y Klekowski, 1975). Asimismo, esta nomenclatura se utilizó para el cálculo de los índices de asimilación y eficiencias de crecimiento. Los valores de los elementos de la ecuación balanceada, se calcularon para las marcas de clase de cada clase-peso, utilizando los valores obtenidos en las distintas submuestras, como sigue: consumo de alimento, 3% del peso corporal (g); heces, 8.0% de la ingesta (g) y la excreción nitrogenada considerando el consumo de alimento (g) por el factor 0.257 que corresponde al valor centesimal proteico de la ración administrada y luego utilizando las conversiones por el método descrito anteriormente. El consumo de oxígeno se obtuvo de la ecuación de la recta de regresión de la captación de gas (ml 02/h) en función del peso corporal (g) ajustada por el método usual de cuadrados mínimos. Las diferencias entre los resultados, se estimaron estadísticamente significativas ( p < 0.05 ) mediante las pruebas no paramétricas de Mann- Whitney y Kruskal Wallis (Zar, 1974). RESULTADOS Y DISCUSIÓNEl pescado blanco de Pátzcuaro, Chirostoma estor Jordan, es una especie que muy fácilmente pierde su estado estable fisiológico ante estímulos externos como son la manipulación o el propio forcejeo de los peces en la red de captura. El 100% de sobrevivencia obtenido en este trabajo, tanto en el momento de la colecta como durante el traslado, la manutención y la experimentación a que fueron sometidos los especímenes, se debe probablemente al extremo cuidado que se tuvo de no tocar a los peces, al hecho de mantener la temperatura del agua similar a la que tenían en los corrales y proporcionarles oxígeno, durante el traslado. Al respecto, se conoce que los animales acuáticos, toleran mejor cualquier "stress", si disponen de oxígeno en abundancia (Kinne, 1971). Estas características del pescado blanco, normó el diseño experimental que se siguió en este estudio. Se utilizaron submuestras tomadas al azar para cada una de las mediciones, las cuales no se volvieron a emplear. Para trasladar los peces de los acuarios de manutención a las cámaras respirométricas, se capturaron con un vaso de precipitado. La distribución de los datos correspondientes al peso y las frecuencias respectivas, se presentan en la figura 1. Las medidas se contrastaron mediante la prueba de Kruskal-Wallis y los resultados indicaron que las diferencias encontradas son altamente significativas (p < 0.001 ). Los datos agrupados en a tienen un intervalo de peso de 1.8 a 2.8 g; en b de 2.5 a 3.9 g y c de 3.55 a 4.75 g. Los cálculos del balance energético se efectuaron a partir de las marcas de clase de los intervalos de peso considerados. 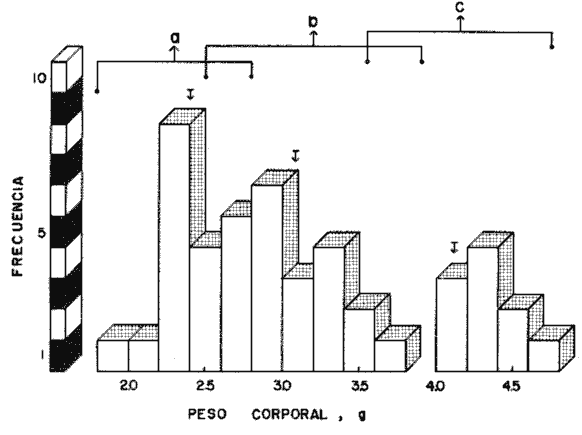 Figura 1. Distribución de frecuencias del peso corporal de Chirostoma estor. Agrupaciones consideradas en relación al peso: a, b, c. Las flechas indican las medias de las distribuciones. Cabe destacar que tanto los animales pequeños, con pesos promedios de 2.31 g y 3.08 g como los de mayor tamaño de 4.12 g, son peces maduros sexualmente. Los cortes histológicos que se realizaron en las gónadas de hembras y machos mostraron ovocitos con diferentes grados de vitelogénesis y asimismo, testículos en diferentes etapas de maduración. Ahora bien, como los especímenes se alimentaron ad libitum (3% peso corporal) en dos raciones diarias, se puede afirmar que efectivamente se alimentaron a saciedad (Beamish et al., 1975). La ingestión de alimento fue estadísticamente diferente; los más pequeños consumieron 40% menos que los más grandes y 22% menos que los de peso intermedio de la muestra (Tablas 1 y 2). Este comportamiento es común en los peces y se ha confirmado para especies tales como Micropterus salmoides y Notropis athedonoides (Beamish et al., 1975). Así, es posible considerar que los ejemplares adultos de C. estor, utilizados en este estudio, se encuentran aún en la fase activa de crecimiento. En el intervalo de peso medio (1.90 a 4.65 g) los organismos consumieron el 2.3% del alimento suministrado. Este valor concuerda con lo señalado por varios autores para otras especies; Oysostoina aurata consume entre el 2.3% y el 5.2% de su peso en alimento (Sabaut y Luquet, 1973); Ophicophalus punctatus, de 1.61% a 4.08% p.c./día (Mercy-Gerald, 1976). Cabe destacar que este autor encontró en su trabajo una amplia gama de consumo en relación al peso y el dato mencionado, que concuerda con el obtenido para C. estor, corresponde a peces de mucho mayor tamaño (30 g). TABLA 1 BALANCE ENÉRGETICO DE Chirostoma estor ALIMENTADOS CON DIETA COMERCIAL (C=P+R+F+U). VALORES PROMEDIO EN mg/día/individuo±ERROR ESTANDAR 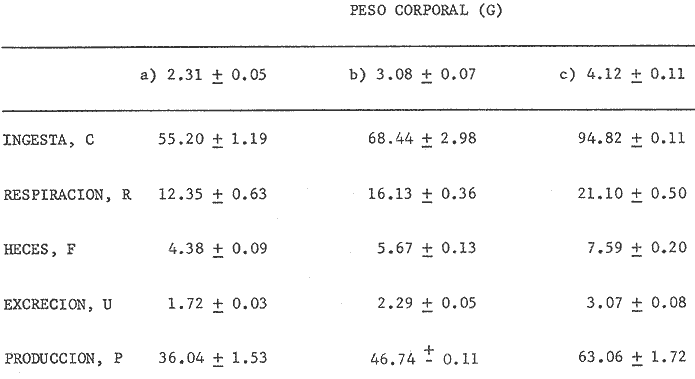 TABLA 2 BALANCE ENERGÉTICO DE Chirosloma estor ALIMENTA DOS CON DIETA COMERCIAL (C= P+R+F+U). VALORES PROMEDIO EN Cal/día/individuo, Y COMO POR CENTAJE DEL CONSUMO DE ALIMENTO 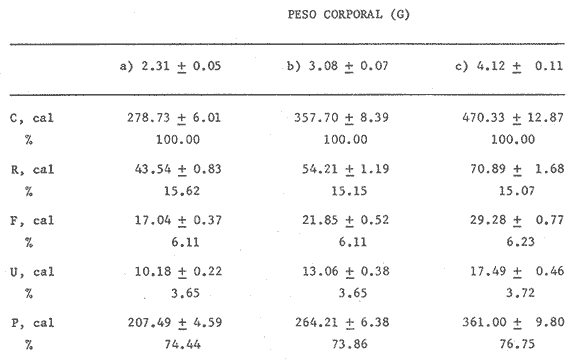 El valor calórico de la ración consumida fue de 278.73, 357.70 y 470.33 caUdía para los tres grupos considerados (Tabla 2). Estas diferencias fueron estadísticamente significativas (p < 0.05). Ahora bien, los productos fecales se obtuvieron por recolecciones sucesivas de materia sólida sin considerar la porción en solución de las mismas. Esta metodología es generalmente aceptada ya que las substancias solubles son escasas y la medición conlleva menos del 4% de error en la estimación de las heces totales (Elliot, 1976), lo que está dentro del error experimental en este trabajo. Chirostonza estor, tiene una tasa de evacuación de heces, correspondiente al 8% de su peso corporal por día. Esta tasa es baja en comparación con salmónidos alimentados con dieta comercial, que alcanza valores entre el 10% y el 20% (Beamish, et al., 1975), en tanto que es comparable con el 11 % determinado para Salmo trutta (Elliot, 1976). El valor calórico de las heces fue de 3,854.63 cal/g. Las heces producidas alcanzan valores de 17.04, 21.85 y 28.28 cal./día (Tabla 2). Estos resultados son comparables con los de Mercy-Gerald (1976) para Ophiocephalus punctatus y por Pandian y Ranghuraman (1972) tanto para O. striatus como para Micropterus cyprinoides. Asimismo, de los datos presentados se puede inferir que los animales de todas las categorías de peso, pierdan alrededor del 60% de la energía consumida en el alimento, vía evacuación de productos fecales. Por otra parte, al calcular la tasa de pérdida en heces a partir de la ecuación F = C-A, donde F representa el valor calórico de las heces, C el del consumo de alimento y A la asimilación, las pérdidas son mayores 1.6 a 1.8 veces (Tabla 3). Como se puede observar, los datos corresponden a 27.20, 35 a 39 y 38 a 47 cal./día, en las diferentes clases pesos. Estos resultados son del mismo orden que en la carpa herbívora Ctenopipharyngodon idella, alimentadas con dicta de procedencia animal y mantenidas a 22 °C (Fisher, 1972a). Ahora bien, la energía asimilada es la energía fisiológicamente útil y se puede calcular mediante los diferentes métodos mostrados en la Tabla 3 (Brett y Groves, 1979). Los peces de mayor tamaño asimilaron más que los medianos y pequeños, lo cual está en relación al mayor consumo de alimento ya que en relación a la ingesta, las pérdidas en heces fueron similares en los tres grupos (Tabla 3). No se encontraron diferencias significativas entre los valores obtenidos por los diferentes métodos de cálculo (p > 0.05). Asimismo, la eficiencia de asimilación del 90%, fue similar en todas las clases pesos consideradas. En la mayoría de los peces de agua dulce, la eficiencia de asimilación es del 70 al 80%. Sin embargo, el resultado obtenido para C. estor es consistente tomando en cuenta que el exponente de la recta de regresión de la tasa metabólica en función del peso, es también un índice de asimilación (Paloheimo y Dickie, 1966); para esta especie el exponente fue de 0.91. La energía perdida en productos de excreción varía con el peso del pez, con la temperatura y con la estación. No obstante cuando las pérdidas en productos nitrogenados se expresan como proporción de la captación de energía, el efecto del peso no es aparente. Tal es el caso del pescado blanco cuyas pérdidas corresponden al 3.6% de la energía consumida en el alimento, independiente del peso corporal (Tabla 3), lo que concuerda con los valores obtenidos para peces aclimatados a 20 °C (Elliot, 1976). TABLA 3 ASIMILACIÓN Y EFICIENCIA DE UTILIZACIÓN DE LA ENERGÍA CONSUMIDA POR Chirostoma estor. * = HECES CALCULADAS COMO SE SEÑALA EN**. 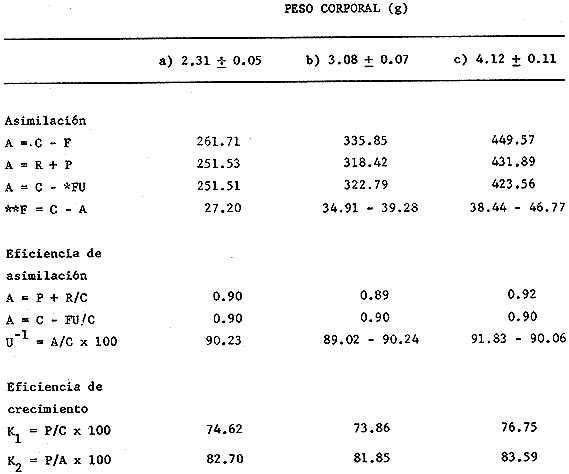 Los especímenes consumieron 14.18 ± 0.21, 18.11 ± 0.41 y 24.37 ± 0.64 mg de proteína por día. Estos resultados son reflejo de una alta eficiencia de utilización de las proteínas, aproximadamente 86%. El consumo de oxígeno, para una submuestra de 25 animales, se calculó en función del peso. La recta de regresión resultante, que expresa la dependencia de la captación de oxígeno del peso corporal es: R = 170 W0.91 Donde R es el consumo de oxígeno (µl/h) y W es el peso vivo de los peces (g). Esta dependencia se presenta en la figura 2. El intervalo de peso considerado fue de 2.2 a 3.2 g. Ahora bien, el promedio de la tasa de captación calculado para los datos agrupados fue de 375, 479 y 610 µl/h, respectivamente (Tablas 2 y 3). La relación entre el consumo de oxígeno (R) y el peso corporal, ha sido ampliamente estudiada en peces y existe una fórmula general (Fisher, 1972a): R = 370 W0.81 Donde R corresponde apl 02/h y W se expresa en g. 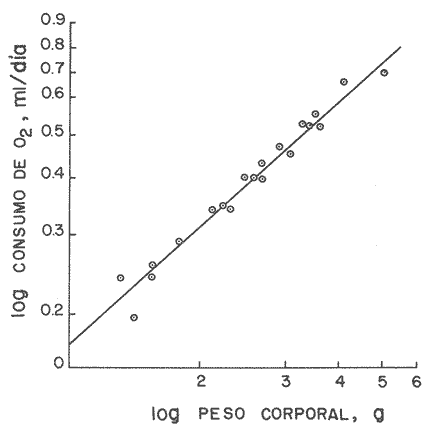 Figura 2. Dependencia del consumo de oxígeno del peso corporal. R = 0.17 W0.91; r = 0.90. Las tasas de consumo de oxígeno obtenidas en este estudio son inferiores (aprox. 40%) a las calculadas utilizando la fórmula general mencionada (Tabla 4). Tal discrepancia es atribuible a la hora del día en que se hicieron las mediciones ya que no se dispuso de los datos de captación de oxígeno en un ciclo de 24 horas, por lo que pudo haber coincidido con un mínimo en la curva. Sin embargo, estas diferencias no son esencialmente importantes en los estudios de bioenergética, dada la variabilidad interespecífica reportada para los peces (Fisher, 1972a). TABLA 4 COMPARACIÓN ENTRE LOS VALORES DE CONSUMO DE OXíGENO OBTENIDOS EN ÉSTE TRABAJO CON LOS CALCULADOS MEDIANTE LA FÓRMULA GENERAL (WINBERG, 1%1). VALORES EN µlO2/hora 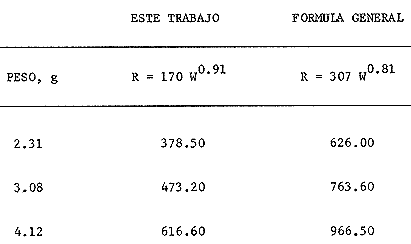 El exponente de la ecuación potencial fue de 0.91 para el pescado blanco; como se dijo anteriormente este dato se considera un índice de la eficiencia de asimilación. Así, si la dependencia del peso tuviera un valor de intersección en 307 µl O2 1-1 como en la ecuación general, el valor de la pendiente se reduciría, lo cual sería inconsistente con los valores de asimilación obtenidos. Brett (1972) señala que la pendiente de dicha recta, tiene un valor de 0.8 en un intervalo de 0.76 a 0.86 para varias especies de peces, tanto de agua dulce como de ambientes marinos; no obstante, otros autores reportan un dato similar al encontrado para C. estor en especies dulceacuícolas (Beamish, 1964; Beamish y Mookherjii 1964; Fisher, 1972a; Fisher y Lyakhnovich, 1973). En relación al consumo de oxígeno también pueden ocurrir desviaciones respecto a la ecuación general, ya que la tasa metabólica no sólo es dependiente del peso, sino que también lo es del conjunto de factores que conforman el ambiente ecológico de las especies. De la determinación de los parámetros discutidos, fue posible calcular el balance energético de Chirostoma estor adulto (Tabla 1). Los datos promedio de las clases-peso (cal./día) se presentan en la Tabla 1. El crecimiento de los peces en los dos primeros grupos fue de 86 cal./día/mg de peso corporal y el del tercer grupo fue de 88 cal./día/g. Así, no se encontraron diferencias en los primeros; en contraste, los peces de mayor tamaño crecieron significativamente y a una tasa mayor (p < 0.05). En relación a la energía ingerida, también la incorporaron en forma diferencial y con la misma tendencia; los de tamaño pequeño e intermedio incorporaron un 74% y los de mayor talla, un 77%. Resultados similares se obtuvieron considerando la eficiencia bruta de crecimiento, K1 (Tabla 3). Dicho índice define esa parte de la energía ingerida que ha sido utilizada para crecimiento (Fisher, 1972a, 1972b; Paloheimo y Dickie, 1966), en el pescado blanco se obtuvieron valores de 74.6, 73.86 y 76.75 para las diferentes clases-pesos. Por otra parte, el índice K2 que refleja la eficiencia con que la energía asimilada se incorpora al pez como crecimiento (Tabla 3), es un 10% mayor. El campo de crecimiento de Chirostoma estor alimentado con dieta balanceada comercial, fue en los peces pequeños, de 36 mg/día. En los medianos de 46.7 mg/día y en los de mayor tamaño de 63.1 mg/día. Así, aunque la eficiencia de crecimiento es regularmente alta con esta dieta y la asimilación de la misma es en promedio 83% (también alta), se podría definir a Chirostoma estor como un eficiente convertidor de energía, pero de lento crecimiento. Este escaso crecimiento provocaría el enanismo de esta población en cautiverio, lo que podría atribuirse a la baja temperatura del lago en la estación de invierno (14 °C), a la alta densidad en que se criaron los peces en los corrales o bien a la escasez de alimento. Si bien en el laboratorio, se mantuvieron con dieta ad libitum, el tiempo de permanencia de los peces en el mismo, fue de sólo 30 días. La coherencia de estos resultados puede comprobarse estimando el valor calórico de la ración, para producir el crecimiento observado (Solomon y Bradfleld, 1972): Energía de la ración = 1.25 (energía metabólica + energía de producción). Con base en esta ecuación, los valores teóricos calculados para las diferentes clases-pesos fueron: a) 314, b) 398 y c) 540 cal./día. Aunque existe una diferencia de aproximadamente 11% entre los valores estimados y los observados (Tabla 2), estas diferencias no fueron estadísticamente significativas (p > 0.05). Por tanto, la energía captada en el alimento da cuenta de la producción observada en las condiciones prevalecientes en el laboratorio. Este tipo de estudios ecofisiológicos con enfoque energético muestran que controlando el medio, especialmente en relación a la temperatura y al alimento, se puede alcanzar un crecimiento óptimo en varias especies de peces (Wallace, 1973). Así, para mejorar las biotecnias existentes involucradas con esta especie, es posible recomendar que se proporcione a los peces alimento balanceado a saciedad, en dos raciones diarias al 3% del peso corporal y que en lo posible, se administren sobre 500 cal./día./individuo. Que se cubran los estanques con plástico, con el fin de aumentar la temperatura del agua en los meses fríos. Conclusiones- La supervivencia de los organismos, durante el traslado y la manutención en el laboratorio, fue del 100%. - Los peces consumen el equivalente al 2.3% de su peso corporal cuando se alimentan con una dieta balanceada comercial. El valor calórico del consumo fue de 280 - 470 cal./día/individuo. - La evacuación del las heces correspondió al 8% del peso corporal por da, con un valor calórico de 3,854.6 cal./g. Los organismos perdieron entre 17 y 30 cal./día en materia fecal, equivalente al 6% de la energía consumida en el alimento. En productos de excreción, perdieron alrededor del 4% de la energía captada. - Los especímenes, presentaron una alta eficiencia de asimilación (90%) del alimento administrado, en todas las clases-pesos consideradas. - La ecuación que describe el consumo de oxígeno (µl/h) en relación al peso corporal (g) es R = 170 W0.91 - La utilización del oxígeno fue del orden del 15% de la energía ingerida en las tres clases-pesos. - El balance energético de Chirostoina estor fue positivo y la producción calculada fue 90 cal./día/mg de peso corporal. - La eficiencia bruta de crecimiento obtenida, alcanzó un 75% y la eficiencia neta un 80%. Así, el campo de crecimiento se estimó entre 40 y 60 mg/día. - De los resultados obtenidos se puede inferir que los organismos objeto de este estudio, son eficientes convertidores de energía pero de crecimiento lento, probablemente debido a la baja temperatura del lago (14 °C) en la época de invierno y a la escasez de alimento administrado en el encierro, entre otras causas. - Es posible traducir la alta eficiencia de asimilación del alimento balanceado, en una mayor tasa de crecimiento, controlando las variables temperatura y alimentación. AgradecimientosLos autores agradecen a R. Sumano, de la Delegación de Pesca en Pátzcuaro, Michoacán, el apoyo brindado para la captura de los peces. A J. Tolosa, quién realizó el estudio histológico de las gónadas de los organismos. A H. Troncoso por los análisis bromatológico y calórico de alimento suministrado a los peces. LITERATURAANCONA, I., BATALLA, M.A., CABALLERO, C., HOFFMAN, C., LLAMAS, R., MARTIN DEL CAMPO, R., OCHOTERENA, L, RIOJA, F., ROCA, J., SAMANO, A., VEGA, C. y F. VILLAGRAN, An. Inst. Biol, Méx, Prospecto biológico del Lago de Pátzcuaro. 1940. 418-499. 11 (2): BEAMISH, F.W.H., Col. J. Zool, Respiration of fishes with special emphasis on standard oxygen consumption. II. Influence of weight and temperature on respiration of several species. 1964. 177-187. 42: BEAMISH, F.W.H. y P.S. MOOKHERALL, Can. J. Zool., Respiration of fishes with special emphasis on standard oxygen consumption. I. Influence of weight and temperature on respiration of goldfish Carassius auratus. 1964. 161-175. 42: BEAMISH, F. W. H., NIIMI, A. J. y P. F. LETT, Comparadve Physiloff Funcdonal Aspeas of Structural Materíais. Bioenergetics of teleost fishes: environmental influences. Bolis, L, MadreU, H.P. y K. Schmidt-Nielsen (Eds.) North Holland, Publ. Co. 1975. 189-209. BRETT, J. R., Res. Physiol., The metabolic demand for oMen in fish particularly salminids, and a comparision with others vertebrates. 1972. 151-170. 14: BRETT, J. R., J. Fish Res. Bd. Canada, Scope for metabolism and growth of sockeye salmon, 0ncorhynchus nerka, and some related energetics. 1976. 307-313. 33: BRETT, J. R. y T. D. D. GROVES, Fish Physiology, 3. Physiological Energetics. In: Hoar, W.S., Randall, D.J. y J.R. Brett (Eds.). Academic Press. New York: 1979. 280-352. DUNCAN, A. y R. Z. KELKOWSKI, Methods for Ecological Bioenergetics IBP-24. Parameters of an energy budget. Grodzinski, W., Meltowski, R.Z. and A. Duncan (Eds.) Blackweil Sci.Publ. Oxford: 1975. 97-147. ELLIOT, J. M., J. Animal, Ecol., Energy losses in the waste productos of brown trout (Salmo trutta L). 1976. 561-580. 45: ESPINA, S., DÍAZ, F., LATOURNERIE, J., SÁNCHEZ, A., ROSAS, C. y M.E. VEGA, IV Cong. Nal. Zool. Ensenada, BCN Res: Estudios preliminares para determinar la eficiencia de crecimiento del pescado blanco de Pátzcuaro, Chirostoma estor (Atherinidae). 1980. 84. FISHER; Z., Pol. Arch Hydrobiol., The elements of energy balance in grass casp (Ctenopharyngodon idella Val.) Part I. 1970. 421-434. 17: FISHER, Z., Pol, Arch Hydrobiol., The elements of energy balance in grass carp (Ctenopharyngodon idella Val.). Part II. Fish fed with animal food. 1972a. 65-82. 19: FISHER; Z, Pol. Arch Hydrobiol, The elements of energy balance in grass carp (Ctenopharynogodon idolla Val.). Part III. Assimilability of proteins, carbohydrates and lipids by fish fed with plant and animal food. 1972b. 83-95. 19: FISHER, Z., y V.P. LYAKHNOVICH, Pol. Arch. Hydrobiol., Biology and bíoencrgetics of grass carp (Ctenopharyngodon idella Val.) 1973. 524-557. 20: HENDERSON, F., Progr. Inv. Fom. Pesq. Programa de evaluación de recursos para apoyar el desarrollo pesquero en aguas mexicanas. , 8 Contribuciones al estudio de pesquerías en México. MEXICO/PNUD/FA0 CEPM; 1974 KINNE, O., Maríne Ecology. Part 2. Kinne, O. (Ed.) Wiley Interscience, London: 1971. 821-874. Vol. I, LARA, V.A., Simp. FA0/Carpas: Acuicultura en América Latina Aspectos del cultivo extensivo e intensivo del pescado blanco de Pátzcuaro (Chirostoma Jordan 1879). 1974. 113-116. 1: MERCY-GERALD, V., Hydrobiologia, The effect of size on the comsumption, absorption and conversion of food in 0phiocephalus punclatus Bloch. 1976. 77-85. 49: NIIMI, A. J. y F. W. H. BEAMISH, Can. J. Zool., Bioenergetics and growth of largemouth bass (Micropterus salmoides) in relation to body weight and temperature. 1974. 447-456. 52: PALOHEIMO, H. E. y L. M. DICKIE, Fish Res. Bd. Canada, Food and growth in fishes. I-A growth curve derived from experimental data. J. 1965. 522-542. 22 (2): PANDIAN, T. J. y R. RAGHURAMAN, Tilapia mossambica. Mar. Biol., Effects of fecding rate on conversion efriciency and chemical composition of the fish 1972. 129-136. 12: PHILLIPS, M. A., Fish Nutrition. Calorie and energy requireincnt. In: Halver, J. E.(Ed.). Academic Press, New York: 1972. 2-27. ROSAS, M., Pescado blanco (Chirstoma estor). Inst. Nac. Inv. Biológico-Pesqueras. 1970. 79 p. SABAUT, J. J. y P. LUQUET, Mar. Biol., Nutitional requireiments of the gilthcad bream Chtysophys aurata. Quantitative protein requirements. 1973. 50-54. 18: SALMON, D. H. y A. E. BRADHELD, J. Animal, Ecol., The cncrgeties of feeding, metabolism and growh of perch (Perca fluviatillis L.). 1972. 699-718. 41: SOLORZANO, A. P., La pesca en el Lago de Pátzcuaro, Mich. y su Importancia Económica y Regional. Sría. de Marina. Dir. Gral. de Pesca. 1955. 58 p. SOLORZANO, A. P., Algunos aspectos biológicos del pescado blanco del Lago de Pátzcuaro, Mich., Chirostoma estor. Dir. Gral. de Pesca e Ind. Conexas. Inst. Nal. de Inv. Biológico Pesqueras: 1963. 1-15. VEGA, M. E., DIAZ, F., LATOURNERIE, J., SÁNCHEZ, A. y S. ESPINA, V. Congr. Nal. Zool. Estudio comparado sobre la eficiencia de crecimiento en Chirostorna nestor juveniles y adultos. Cuernavaca, Morelos México. 1981. VEGA, M. E., Tesis Prof. Biología. Fac. Ciencias, Balance energético de Chirostorna estor, captación de alimento y eficiencia de la extracción de energía en juveniles y adultos. Univ. Nal. Autón. México. 1982. 36p. WALLACE, J.E., J. Comp. Biochem Physiol, Observaciones on the relationship between the food consumption and metabolic rate of Blenniusius pholis. 1973. 293-306. 45A: WARREN, C.E. y G.E. DAVIS, The Biological Basis of Feshwater Fish Production. Laboratory studies on the feeding, bioenergetics and growth of fish. Gerking, S.D. (Ed.) Blackwell Sci. Publ., Oxford: 1967. 175-214. WINBERG, G.G. Vop. Ikhtiol, New data on intensity of metabolism in fish. 1961. 158-165. l (18): ZAR; H.J., Biostatiscal Analysis. Pretince-Hall, Inc. Englewood Cliffs, New York. 1974. 690 p.
|