|
ANÁLISIS DE LA FRECUENCIA Y HABITOS ALIMENTICIOS DE
LA "CIGALA", Nephrops norvegicus (LINNEO, 1758) EN EL MEDITERRANEO OCCIDENTAL
(CRUSTACEA: NEPHROPSIDAE)
Trabajo recibido e120 de marzo de 1986 y
aceptado para su publicación el 11 de marzo de 1987.
Asunción Gual-Frau
Instituto de Investigaciones Pesqueras, Barcelona,
España.
Manuel Gallardo-Cabello
Universidad Nacional Autónoma de
México. Instituto de Ciencias del Mar y Limnología. A.P. 70-305.
04510 México, D.F. Contribución 614 del Instituto de Ciencias del
Mar y Limnología, UNAM.
En el presente trabajo, se analiza el régimen
alimentario de la cigala Nephrops norvegicus del
Mediterráneo Occidental, en relación al sexo, clases de tallas,
profundidad y período estacional. La dieta no presentó
diferencias importantes de acuerdo al sexo ni a las clases de tallas. Los
organismos más abundantes en el contenido estomacal fueron:
crustáceos, equinodermos, poliquetos, moluscos y peces. La
alimentación es más activa y la dieta más variada durante
la primavera y el otoño a una profundidad de 150 m. La cigala es un
depredador macrófago, eurífago y oportunista.
This paper deals with data about Nephrops
norvegicus diet aecording to: sex, length categories, depth and
seasonal patterns in the Wester Meditarranean Sea. The main types of food founs
in the foregut contents, regardless of sex or size were: crustaceans,
echinoderms, polychaetes, molluscus and fishes. The increase of active feeding
and the higherdiet diversity index ocurred during springand autumn in depths of
150 m. Norway lobster is an euriphage and opportunist predator.
El estudio de las relaciones tróficas de los organismos que integran un ecosistema determinado, es fundamental para el conocimiento del papel (nicho) que éstos juegan en la dinámica de la comunidad. El estudio de la alimentación de una especie, por medio del análisis del contenido estomacal, permite conocer su régimen alimenticio, a través del cual puede determinarse la posición trófica que ocupa en el ecosistema y sus posibles relaciones de dependencia con otras especies ó grupos. De la misma forma, pueden conocerse las posibles variaciones de la dicta, en función de determinados parámetros como pueden ser la profundidad y épocas del año, lo que puede indicar, en primer lugar, su localización en el espacio al menos durante el momento de la alimentación y en segundo lugar su localización "funcional" como un componente más de la biocenosis. Por otra parte, el estudio del régimen alimenticio de una especie no selectiva y altamente eurífaga, puede proporcionar una idea aproximada de su entorno. En el presente trabajo, se realiza una análisis sobre el régimen y hábitos alimenticios de la cigala, Nephrops norvegicus en el Mediterráneo Occidental, en relación al periodo estacional y la profundidad, aportándose por primera vez, datos en la zona de estudios mencionada, con respecto a los índices de diversidad en número de componentes de la dieta y repleción gástrica, disponibilidad del alimento, periodos de mayor o menor actividad alimentaria, migraciones verticales y ritmos de alimentación de periodicidad nictemeral (circadiana). MATERIAL Y METODOSLos ejemplares de la cigala, Nephrops norvegicus se obtuvieron frente a las costas de Cataluña - Mediterráneo Occidental - entre Blanes y Vilanova i la Geltrú. En la figura 1 se muestra la zona de estudios. 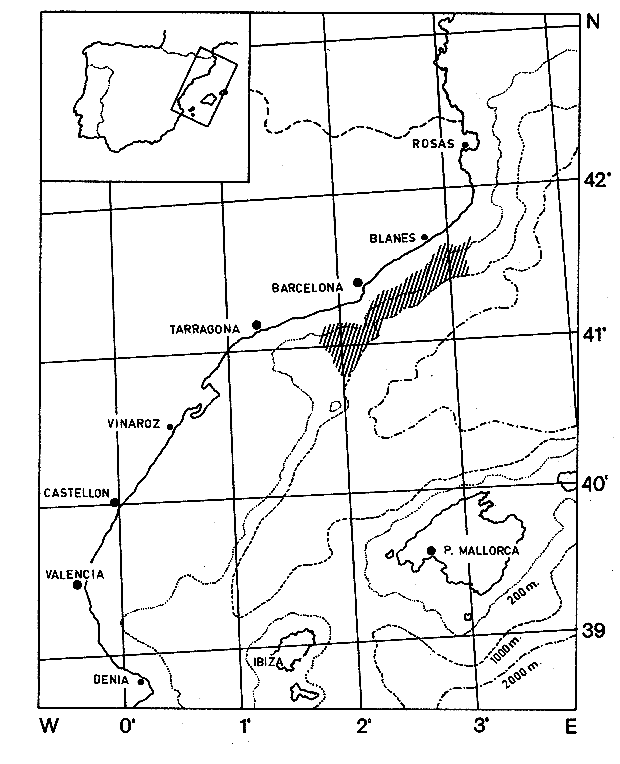 Figura 1. Mar Catalán y situación de las áreas de estudios. Los datos biológicos se obtuvieron de un total de 552 ejemplares, procedentes de los muestreos biológicos y de la captura comercial, realizados de octubre de 1976 a junio de 1978. No se realizaron capturas durante los meses de marzo y noviembre de 1977, así como en enero, marzo y mayo de 1978. Los muestreos se llevaron a cabo efectuando arrastres de una hora de duración a distintas profundidades, el primero a 150 m y el segundo a 300 m, utilizando para ello, una red comercial con un copo de 22 mm y un sobrecopo de 3 min. de abertura de malla. En la Tabla 1 se muestra el número y porcentaje de los ejemplares capturados mensualmente, en relación a las profundidades mencionadas. De cada ejemplar se obtuvo: la longitud total, la longitud del cefalotórax, el peso y el sexo. Los estómagos fueron extraídos y conservados en alcohol al 70%, siendo clasificados de acuerdo al tamaño de cada ejemplar, según el criterio establecido por el "Working Group of Nephrops Stock" (ICES), que establece clases de tallas con intervalos de 5 mm de longitud del caparazón. 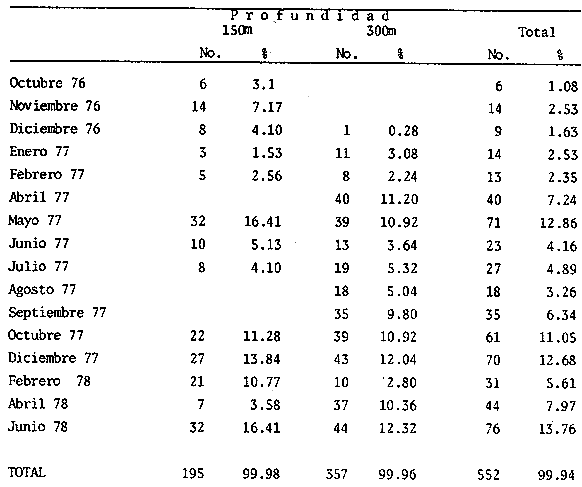 TABLA 1 NÚMERO DE EJEMPLARES CAPTURADOS DURANTE LOS MUESTREOS MENSUALES EN RELACIÓN A LA PROFUNDIDAD. Los estados de repleción gástrica, en que se clasificaron los estómagos fueron los siguientes: I. vacío, II. casi vacío, III. casi lleno y IV. lleno. El coeficiente de repleción gástrica se obtuvo dividiendo el número total de estómagos llenos entre el número total de estómagos. Las presas fueron identificadas con la ayuda de un microscópio esteroscópico. Resultó de gran utilidad la comparación de las estructuras (restos) encontrados en el contenido estomacal de la cigala, con los de ejemplares bentónicos completos, procedentes de la misma zona de estudios. Dichos organismos, obtenidos con redes de arrastre de fondo y dragas, proporcionaron una valiosa información sobre la fauna de acompañamiento. Aquellas presas cuya identificación resultó dudosa son reportadas a nivel de phylum, clase o familia según sea su caso. Los resultados que se presentan a continuación, se expresan como la frecuencia de ocurrencia, es decir, el porcentaje de los estómagos (excluyendo los vacíos) que presentaron un tipo particular de organismo. No se consideró el peso del estómago debido a la desviación que puede presentarse en la obtención de este parámetro, dada la cantidad de jugos gástricos existentes en el tracto digestivo y la acumulación de material no digerible en el contenido estomacal (Thomas y Davidson, 1962). Asimismo, no se obtuvo la longitud o peso de los organismos debido a que en la mayoría de los casos, sólo fue ingerida una parte del cuerpo de la presa. El índice de diversidad en número de componentes de la dieta, fue obtenido por medio de la aplicación de la ecuación de Levins (1968): B = 1/Σ Pi² Donde Σ Pi², es la sumatoria de los cuadrados de la frecuencia relativa de las especies encontradas. RESULTADOS Y DISCUSIÓN
ANÁLISIS DEL CONTENIDO ESTOMACALDel número total de ejemplares examinados (552) el 91.5% presentó un estado de repleción gástrica que osciló entre 25% y 100%. Solamente 47 individuos (el 8.5%) presentaron el estómago completamente vacío. Se identificaron un total de 1350 organismos, de los cuales solamente 178 (el 13%) pudieron ser reconocidos a nivel de especie. Lagardére (1977) logró identificar 90 especies (el 8.20%) de un total de 1097 organismos encontrados en el contenido estomacal de 125 ejemplares de Nephrops norvegicus del Golfo de Gascuña, Atlántico Nororiental. El bajo porcentaje de presas identificadas al nivel mencionado, indica la enorme dificultad para reconocer los organismos del contenido estomacal de este crustáceo. Ello se debe fundamentalmente a que la especie en estudio "mastica" el alimento antes de su ingestión (Höglund, 1942), a la función trituradora del molino gástrico (Yonge, 1924) y a la degradación del alimento por los ácidos digestivos, lo que impide encontrar, la mayoría de las veces restos de organismos en buen estado y fácil identificación. Siendo ésta la causa de que en la mayoría de los casos solo puedan identificarse las presas a nivel de clase o familia, sin que sea posible determinar una posición taxonómica inferior. Es importante señalar que el estado de trituración en que se encuentra una presa en el contenido estomacal, depende del tiempo de permanencia en el estómago del predador, así como de su propia naturaleza. De esta forma, organismos que poseen partes duras, compuestas por quitina o substancias calcáreas ofrecen un mayor tiempo de conservación - y mayor posibilidad de identificación - aunque su ingestión no haya sido reciente. Por lo contrario, las especies que carecen de estructuras duras son digeridas más rápidamente que las anteriores, presentando grandes dificultades para su identificación, la cual en ciertas ocasiones es imposible. Debido a lo anterior, cabe esperar que en los resultados obtenidos los organismos más lábiles no estén debidamente representados. Así mismo, debe tomarse en cuenta, que en numerosos casos es difícil determinar si los restos que aparecen en el contenido estomacal corresponden a uno ó más individuos, como es el caso de los ofiuroideos. En otras ocasiones, como es el caso de los crustáceos decápodos, no suele presentarse más de un ejemplar en el contenido estomacal, acompañado con restos de otros phyla u órdenes. De acuerdo con lo anterior, el análisis que se presenta debe considerarse como una aproximación a la dicta de Nephrops norvegicus. IDENTIFICACIÓN DE LAS PRESASEn el presente estudio no se encontraron diferencias significativas en la dieta de la cigala, en relación al sexo o a los distintos grupos de edad o clases de tallas, coincidiendo con las observaciones realizadas por Thomas et al. (1962). Los resultados, obtenidos, se muestran en la Tabla 2, expresados como el índice de ocurrencia (IO) de los organismos encontrados en el contenido estomacal de Nephrops norvegicus. La elaboración de dicha Tabla se hizo siguiendo los criterios de diversos autores para las categorías taxonómicas de cada grupo, de la manera siguiente: crustáceos (Bowinan et al., 1982); poliquetos (Fauchald, 1977); moluscos (Brookes et al., 1960); peces (Mc. Neill, 1975); foraminíferos (Levine et al., 1980) y otros grupos (Barnes 1968). 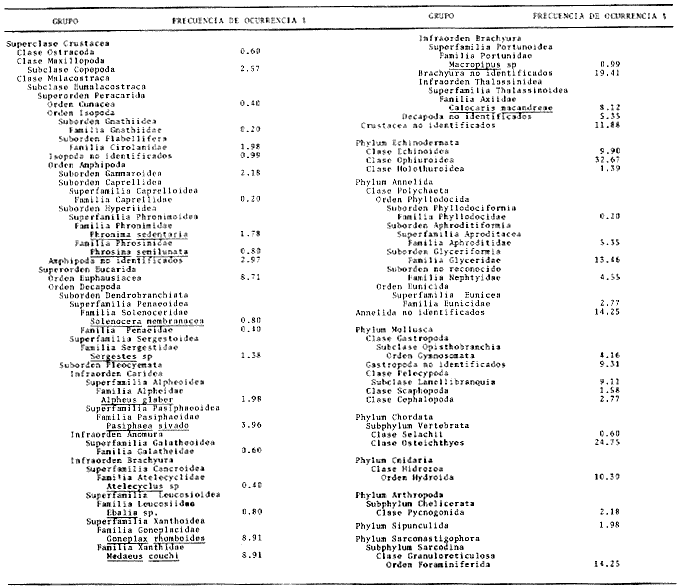 TABLA 2 FRECUENCIA DE OCURRENCIA DE LOS ORGANISMOS ENCONTRADOS EN EL CONITENIDO ESTOMACAL DE Nephros norvegicus. En la Tabla 3 se muestran los valores del IO de los distintos grupos de organismos encontrados. Crustáceos.Los crustáceos constituyeron el grupo más abundante de los organismos que se presentaron en el contenido estomacal de la cigala. Del número total de presas encontradas, los crustáceos ocuparon el 37% y fueron los organismos que presentaron el máximo valor (97.23%) de IO (Tabla 3). Thomas et al. (1962) encuentran que dicho grupo es el más altamente representado en el contenido estomacal de Nephrops norvegicus en el Mar del Norte, con un valor del IO del 76%. Lagardère (1977) reporta que los crustáceos fueron el segundo grupo más abundante en la dieta de la cigala del Golfo de Gascuna, representando el 24.15% del total de las presas encontradas. En el presente estudio, se halló un total de 514 crustáceos, de los cuales 83 individuos (16.14%) no pudieron ser identificados a un nivel taxonómico inferior al de superclase.  TABLA 3 VALORES DE FRECUENCIA DE OCURRENCIA DE LAS PRESAS ENCONTRADAS EN EL CONTENIDO ESTOMACAL DE Nephros norvegicus. El resto, 431 ejemplares (83.85%) pertenecen a tres clases, cinco órdenes, 17 familias, 12 géneros y 8 especies. En las Tablas 2 y 4, se observan los valores del IO para los distintos grupos de crustáceos.  TABLA 4 VALORES DE FRECUENCIA DE OCURRENCIA DE CRUSTÁCEOS ENCONTRADOS EN EL CONTENIDO ESTOMACAL DE Nephros norvegicus. De acuerdo con lo anterior, se observa que los cumáceos y los ostrácodos presentan los valores más bajos del IO, 0.40% y 0.60%, respectivamente, ocupando el 0.46% y el 0.69% del total de los crustáceos identificados. Lagardère (1977) encuentra que dichos grupos son los menos representativos, ocupando cada uno de ellos, el 1.88% del total de los crustáceos. Por otra parte, Thomas et al. (1962) reportan a los ostrácodos como el segundo grupo más importante en la dieta de la cigala, alcanzando un valor del IO del 15%. Para los cumáceos dichos autores encuentran un valor de IO de 6.8%. Los copépodos constituyeron el 3.01% del total de los crustáceos identificados, con un valor del IO del 2.57%. Thomas et al. (1962) encuentran que este grupo es uno de los mejores representados, con un valor del IO del 10.20%. Asimismo, Lagardére (1977) reporta que los copépodos constituyen el segundo grupo más importante de los crustáceos en la dieta de N. norvegicus, ocupando un porcentaje del 24.15%. Los isópodos representaron el 3.71% del total de los crustáceos identificados, con un valor del IO del 3.17%. La familia Cirolanidae fue la más abundante, presentando un valor del IO del 1.98%. Así mismo, Lagardère (1977) encuentra que este grupo es el cuarto mejor representado en la dieta de la cigala, ocupando sin embargo, un porcentaje mayor (13.20%) al encontrado en el presente trabajo. Los anfipodos constituyeron el tercer grupo más abundante, con un valor del IO del 7.92%, ocupando el 9.28% del total de crustáceos identificados. Thomas et al. (1962) reportan resultados semejantes encontrando un valor del IO del 7%. Lagardère (1977) encuentra para este grupo un valor del 4.9% en relación al total de los crustáceos. En el presente estudio, los anfipodos mejor representados fueron los gamaroideos, con un valor del IO del 2.18%, así como las especies Phronima sedentaria y Phrosina semilunata, con valores del 10 del 1.78% y del 0.80%, respectivamente. El segundo grupo mejor representado en la dieta de la cigala es el de los eufausiáceos, con un valor del IO del 8.71%, ocupando el 10.21% del total de los crustáceos. Lagardère (1977) reporta este grupo como el tercero más importante, con un porcentaje del 16.22%. Por otra parte, Thomas et al. (1962) no encuentran ningún representante de este grupo en el contenido estomacal de N. norvegicus. Los decápodos constituyeron el grupo más importante, con un valor del IO del 61.98%, ocupando el 72.62% del número total de crustáceos identificados. Lagardère (1977) reporta para este grupo un valor del 36.6% del total de los crustáceos y Thomas et al. (1962) encuentran un valor del IO del 27%. Aunque en dichos estudios los decápodos no alcanzan cifras tan elevadas como en el presente análisis, constituyen, no obstante, el grupo más abundante en la dieta de N. norvegicus. El grupo de los decápodos estuvo representado por dos subórdenes Dendrobranchiata y Pleocyemata. Este último se halla representado por cuatro infraórdenes: Caridea, Anomura, Brachyura y Thalassinidea. Del número total de decápodos identificados, el 4.55% corresponde a organismos del subórden Dendrobranchiata y el 95.45% a individuos del subórden Pleocyemata. El grupo Dendrobranchiata presentó un valor del IO del 2.58%. El género Sergestes fue el mejor representado, ocupando el 53.85% del total del organismos identificados para este grupo. Le siguen en importancia Solenocera membranacea y los peneidos con valores del IO del 0.80% y del 0.40% y porcentajes del 30.77% y 15.38%, respectivamente. El grupo Pleocymeata presentó un valor del IO del 54.08%. El grupo de los brachiuros, fue el mejor representado, ocupando el 57.71% del número total de organismos de este grupo, con un valor del IO del 39.42%. Le siguen en importancia los talasinideos, los carideos y los anomuros (familia: Galatheidae) con valores del IO del 8.12%, 5.94% y del 0.60% y porcentajes del 23.42%, 17.14% y 1.71%, respectivamente. Los carideos estuvieron representados por, Pasiphaea sivado y Alpheus glaver, especies que alcanzaron valores del IO del 3.96% y 1.98%, respectivamente. Los brachiuros más importantes están representados por las siguientes especies: Medaeus couchi y Goneplax rhomboides ocupando cada una de ellas el 44.55%, con un valor del IO del 8.91%. EquinodermosEl segundo grupo de mayor abundancia en el contenido estomacal de Nephrops norvegicus, estuvo representado por los equinodermos. Dichos organismos representaron el 16.73% del total de las presas encontradas, con un valor del IO del 43.96% (Tabla 3). Thomas et al. (1962) reportan para este grupo un valor del IO del 50%, no obstante dichos organismos ocupan el cuarto lugar de importancia en la dieta de la cigala. Lagardère (1977) encuentra que los equinodermos ocupan el quinto lugar de importancia en el régimen alimentario de N. norvegicus, con un porcentaje del 7.11% del total de presas identificadas. En el presente análisis, todos los equinodermos encontrados pudieron ser identificados perteneciendo a las clases: Echinoidea, Ophiuroidea y Holothuroidea. En la Tabla 2, se observan los valores de IO para los distintos grupos de equinodermos. El grupo de los ofiuroideos, fue el más abundante, ocupando el 74.32% del total de equinodermos identificados con un valor del IO del 32.67%. Le siguen en importancia los equinoideos, con un porcentaje del 22-52% y un valor del IO del 9.9%. El grupo menos abundante fue el de los holoturoideos, con un porcentaje del 3.15% y un valor del IO del 1.39%. Lagardère (1977) encuentra resultados semejantes a los antes mencionados: ofiuroideos, 79.48%; equinodeos, 12.82% y holoturoideos, 7.69%. Por otra parte, Thomas et al. (1962) encuentran que el grupo mejor representado en la dieta de la cigala, fue el de los equinoideos, con un valor del IO del 28%; le siguen en importancia los ofiuroideos, con un valor del IO del 16%. Los holoturoideos estuvieron escasamente representados, con un valor del IO del 0.14%. PoliquetosLos poliquetos constituyeron el tercer grupo más abundante en la dieta de Nephrops norvegicus. Estos organismos ocuparon el 15.44% del total de presas encontradas. El valor del IO fue del 40.59% (Tabla 3). Lagardère (1977) reporta a este grupo como el más abundante en el contenido estomacal de la cigala, ocupando el 40.65% del total del presas identificadas. Thomas et al. (1962) encuentran que el valor del IO para los poliquetos fue del 66%, siendo el segundo grupo mejor representado. En este estudio, del total de los ejemplares hallados, el 65% pudo ser identificado a nivel de familia, el 7.3% como poliquetos sedentarios. El 27.8% restante no pudo ser reconocido. Todas las familias identificadas pertenecen a los poliquetos errantes y constituyen el 90% del total de los organismos que pudieron ser reconocidos como errantia o sedentaria. No obstante, Thomas et al. (1962) y Lagardère (1977) encuentran que de los organismos identificados, es mayor la proporción de poliquetos sedentarios (70% en Thomas et al., 1962 y 58% en Lagardère, 1977) en relación a las formas errantes (30% y 41%). En la Tabla 2, se presentan los valores del IO obtenidos para las familias de poliquetos encontradas en el presente estudio. Los organismos más abundantes corresponden a los miembros de la familia Glyceridae, que ocupan el 46% del total de poliquetos encontrados, con un valor del IO del 13.46%. Similarmente, Thomas et al. (1962) y Lagardère (1977) reportan a esta familia como la que representa más ampliamente a los poliquetos errantes: IO: 9% (en Thomas, et al., 1962) y 22.86% (en Lagardère, 1977), del total de poliquetos encontrados. Le siguen en importancia las familias: Aphroditidae: 18% del total de poliquetos encontrados, IO: 5.35% (13.45% en Lagardère, 1977). Neplityidae: 15.5%, IO: 4.55% (2% en Thomas et al., 1962, la tercera más importante) y Eunicidae: 9.4%, IO: 2.77%. La familia Phyllodocidae estuvo escasamente representada, con un valor del IO del 0.20%, ocupando un porcentaje del 0.68% del total de los poliquetos encontrados. MoluscosLos moluscos representaron el 10,24% del total del organismos identificados, con un valor del IO del 26.93%. (Tabla 3). Thomas et al. (1962) reportan para este grupo un valor del IO del 64% y Lagardère (1977) encuentra un porcentaje del 17.59% del total de presas identificadas. Ambos autores consideran a los moluscos como el tercer grupo más abundante en la dieta de la cigala. Los organismos identificados en el presente estudio pertenecen a las clases: Gastropoda, Pelecypoda, Escaphopoda y Cephalopoda. En la Tabla 2, se presentan los valores del IO para las clases encontradas. La clase Gastropoda fue la más abundante, con un valor del IO del 13.47%, ocupando el 50% del total de moluscos identificados. El 31% de los gastrópodos encontrados pertenecen al orden Gymnosomata y el 69% no pudieron ser identificados en una categoría taxonómica inferior. La subclase Lammelibranquia de la clase Pelecypoda, ocupó el 33.82% del total, con un valor del IO del 9.11%. Le siguen en importancia la clase Cephalopoda, con un porcentaje del 10.29% y un valor del IO del 2.77% y la clase Escaphopoda, con un porcentaje del 5.88% y un valor del IO del 1.58%. Thomas et al. (1962) y Lagardère (1977), encuentran a la clase Gastropoda como la mejor representada, con un valor del IO del 84% (en Thomas) y un porcentaje del 43.52% del total de moluscos (en Lagardère) le siguen en importancia las clases Gastropoda, Scaphopoda y Cephalopoda en ambos autores. PecesLos peces ocuparon el 9.64% del total de presas identificadas, con un valor del IO del 25.34% (Tabla 3). Thomas et al. (1962) encuentran un valor del IO del 13% y Lagardère (1977) reporta que los peces ocuparon el 9.57% del total de presas identificadas. El 98% de los peces identificados en el presente estudio, pertenecen a la clase Osteichthyes y el 2% a la clase Selachii (Tabla 2). HidrozoariosLos hidrozoarios representaron el 3.92% del total del presas identificadas, con un valor del IO del 10.30% (Tabla 3). Otros autores encontraron que los hidrozoarios son menos abundantes en la dieta de Nephrops norvegicus. Thomas et al. (1962) reportan, para este grupo, un valor del IO del 1.3%. Lagardère (1977) encontró que los hidrozoarios representan tan solo el 0.1% del total del presas identificadas en el contenido estomacal de la cigala. Otros gruposLos pignogónidos y los sinpuncúlidos fueron poco abundantes en el contenido estomacal de la cigala. Dichos grupos ocuparon porcentajes de 0.83% y 0.75%, respectivamente, del total del presas encontradas. Los valores del IO fueron del 2.18% para los pignogónidos y del 1.98% para los sinpuncúlidos (Tabla 3). Otros autores no reportan organismos de los grupos mencionados en el contenido estomacal de Nephrops norvegicus. El 14.25% de los estómagos examinados contenía foraminíferos y el 10% material de fondo. Se considera que dichos restos son de ingestión involuntaria, ya sea por hallarse adheridos al alimento o bien, formando parte del contenido estomacal de la propia presa. VARIACIONES EN LA COMPOSICIÓN DE LA DIETA EN RELACIÓN AL CICLO ESTACIONAL.Durante el estudio del régimen alimenticio de Nephrops norvegicus se observaron algunas diferencias importantes con respecto al ciclo estacional. Para el análisis de este fenómeno, se elaboraron las Tablas 5 y 6 así como la figura 2, en donde se muestran los valores del índice de ocurrencia (IO), para cada uno de los grupos. 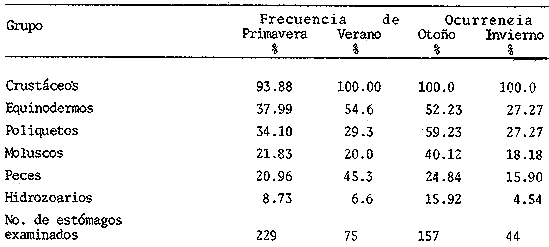 TABLAS 5 VALORES DE FRECUENCIA DE OCURRENCIA DE LAS PRESAS ENCONTRADAS EN EL CONTENIDO ESTOMACAL DE Nephros norvegicus. EN RELACIÓN A LA PERIODICIDAD ESTACIONAL.  TABLA 6 VALORES DE FRECUENCIA. DE OCURRENCIA DE LOS CRUSTÁCEOS ENCONTRADOS EN EL CONTENIDO ESTOMACAL DE Nephrops novegicus. EN RELACIÓN A LA PERIODICIDAD ESTACIONAL. Durante los meses de primavera, el valor del IO de los crustáceos es del 93.88%, siendo inferior al encontrado durante las otras estaciones. No obstante, la diversidad de número de componentes de la dieta es mayor debido a la presencia de organismos que no aparecen durante las otras estaciones, como es el caso de los cumáceos, ostrácodos, anfipodos (familia: Caprellidae) y decápodos (como Solenocera membranacea, Ebalia sp y peneidos). En esta época el IO alcanza los mayores valores para copépodos (3.72%), eufausiáceos (12.10%), anfipodos gamaroideos (3.25%) y Phrosina semilunata (1.39%). El IO disminuye para los decápodos (54%) en relación al verano y al otoño, debido posiblemente, al incremento de presas de otros grupos de crustáceos. No obstante, algunas especies de decápodos alcanzan sus mayores valores durante esta época, como es el caso de Alpheus glaber. Los valores del IO encontrados para los otros grupos son los siguientes: equinodermos (37.99%), poliquetos (34.10%), moluscos (21.83%), peces (20.96%) e hidrozoarios (8.73%). Durante el verano, el IO de los crustáceos es del 100%, pero disminuye el número de componentes de la dicta en relación al período anterior. El 10 desciende para los anfípodos gamaroideos y los eufausiáceos, pero aumenta para los isópodos (familia Cirolanidae) y los decápodos, alcanzando valores del 4.0% y del 88.0% para cada uno de estos grupos. Aparecen decápodos que no se presentaron durante la primavera, como es el caso de Sergestes sp, Macropipus sp y Atelecyclus sp. Durante este período, aumenta la importancia de Medaeus couchi, Goneplax rhomboides y Calocaris macoidreae en relación a la estación anterior. Durante el verano se alcanzan los mayores valores del IO para estas especies. Por otra parte, disminuye el IO de Alpheus glaber. Durante esta época aumenta el IO de Pasiphaea sivado. Franqueville (1971) señala este período, como la época de mayor abundancia de dicho decápodo en el Mediterraneo Occidental. Los valores del IO aumentan para equinodermos (54.6 y peces (45.3%) y disminuyen para poliquetos (29.3%), moluscos (20%) e hidrozoarios (6.6%). 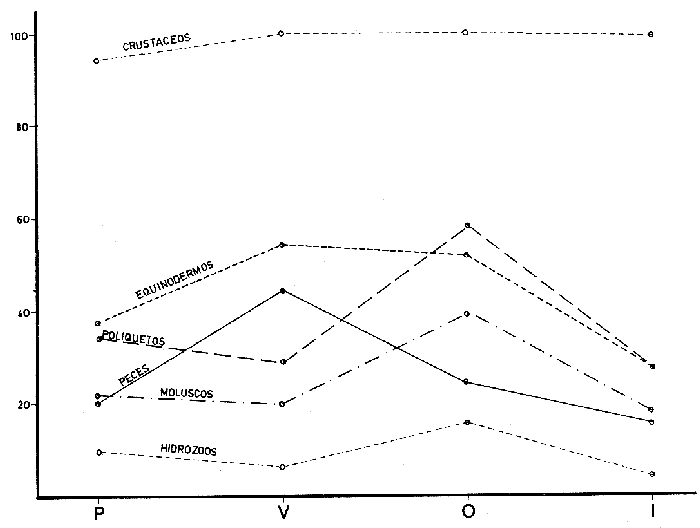 Figura 2. Relación entre los valores del Indice de Ocurrencia de los organismos encontrados en el contenido estomacal de Nephrops norvegicus con respecto al ciclo estacional. Durante el otoño el valor del IO de los crustáceos es de 100% y el número de componentes de la dieta aumenta en relación a la estación anterior. Así, se incrementa la importancia de los copépodos, isópodos, eufausiáceos, anfipodos gamaroideos y de Phrosina semilunata. Aparecen por primera vez los isópodos de la familia Gnathiidae. Phronima sedentaria alcanza el mayor valor del IO (2.54%) durante esta época. Disminuye el valor del IO de los decápodos (71%) en relación al verano, debido posiblemente al incremento del número de organismos de otros grupos de crustáceos. Las especies de decápodos más importantes durante esta época son: Medaeus couchi, Goneplax rhomboides, Calocaris macandreae y Pasiphaea sivado, así como el género Sergestes. Aumenta el valor del IO de los poliquetos (59.23%) y moluscos (40.12%). Disminuye la abundancia de los equinodermos (52.23%) y peces (24.84%). Durante esta época los hidrozoarios alcanzan el mayor valor del IO (15.92%). En el invierno el valor del IO de los crustáceos es del 100%. El número de componentes de la dieta desciende considerablemente en comparación a las otras estaciones. En este periodo, aumenta la importancia de los isópodos en relación al otoño. Disminuye el IO de los decápodos al 46%, estando representados, únicamente por Calocaris macandreae (9.1 %), Goneplax rhomboides (9.1%) y Medaeus couchi (6.81%). Durante esta época descienden los valores del 10 para equinodermos (27.27%), poliquetos (27.27%), moluscos (18.18%), peces (15.9%) e hidrozoarios (4.54%). Las épocas de más activa alimentación corresponden a la primavera y al otoño, períodos en que la dieta es más variada en relación a las demás estaciones. En la Tabla 7, se muestran los valores del índice de diversidad en número de componentes de la dieta y del índice de repleción gástrica.  TABLA 7 INDICES DE DIVERSIDAD EN NÚMERO DE COMPONENTES DE LA DIETA Y DE LA REPLECIÓN GÁSTRICA POR ESTACIONES. El índice de diversidad en número de componentes de la dieta alcanza su mayor valor durante la primavera, presentándose un período de activa alimentación, durante el cual aumenta el índice de repleción gástrica en relación a otras estaciones. Esta época podría relacionarse con una mayor disponibilidad del alimento debido a que durante el mes de abril, surgen en la superficie del Mediterráneo aguas frías y fértiles de menor salinidad, - de origen Atlántico- que penetran a través del estrecho de Gibraltar aumentando la productividad de las aguas marinas (Margalef y Castellvi, 1967). En el verano la dieta es menos variada y el índice de repleción gástrica decrece -posiblemente por una disminución de la productividad, asociada al establecimiento de la termoclina- lo que impide la mezcla de las aguas superficiales con las capas frías, más fértiles, de mayor profundidad (Margalef y Castellvi 1967). Durante los meses del otoño, se presenta un segundo período de activa alimentación, aumentando los índices de diversidad de la dieta y de repleción gástrica. Lo anterior puede deberse a que durante el mes de noviembre, las aguas superficiales comienzan a enfriarse y desaparece la termoclina, lo que trae como consecuencia la mezcla vertical de las aguas y el enriquecimiento de las mismas (Margalef y Castel1vi, 1967). Durante el invierno los valores descienden en relación a los meses anteriores. VARIACIONES EN LA COMPOSICIÓN DE LA DIETA CON RELACIÓN A LA PROFUNDIDADEn el presente estudio se observaron algunas diferencias significativas en la dieta de la cigala, con relación a la profundidad. En las Tablas 8 y 9, así como en las figuras 3 y 4, se observan los valores del índice de ocurrencia (IO), obtenidos para cada grupo, con relación a las profunidades de 150 m y 300 m.  TABLAS 8 VALORES DE FRECUENCIA DE OCURRENCIA DE ORGANISMOS ENCONTRADOS EN EL CONTENIDO ESTOMACAL DE Nephrops novegicus EN RELACIÓN A LA PROFUNDIDAD. En el caso de los crustáceos, se observa que el IO alcanza su mayor valor (100 %) a una profundidad de 300 m y que desciende ligeramente (92.39 %) a una profundidad de 150 m. A menor profundidad la dieta de Nephrops norvegicus es más variada y está formada por organismos que no se presentaron a los 300 m, como es el caso de cumáceos, ostrácodos, copepódos, isópodos (familia Gnathiidae) y anfípodos (familia Caprellidae y ejemplares de Phrosina semilunata). Son mayores los valores de los isópodos (5.27%) anfipodos (familia Gammaroidea, 4.7% y Phrosina sedentaria, 3.52%) eufausiáceos (15.29%), decápodos (Sergestes sp, 2.35%; Galatheidae, 1.17%; Ebalia sp, 1.17%; y Macropipus sp, 1.76%), poliquetos (42.93%), moluscos (30.43%), peces (31.52%) e hidrozoarios (15.76%). Los equinodermos mostraron ser igualmente abundantes en el contenido estomacal de Nephrops norvegicus, tanto los colectados a 150 m como a 300 m de profundidad. En profundidades de 300 m se presentan organismos diferentes a los encontrados en profundidades menores, como es el caso de algunos decápodos (peneidos, 0.62% y Atelecyclus sp, 0.62%). Asimismo, a dicha profundidad, descienden los valores de los isópodos (2.18%), anfípodos (3.41%), eufausiáceos (5.61%) y decápodos del suborden Dendrobranchiata (2.48%). No obstante, se incrementa la importancia de Solenocera membranacea (0.93%). 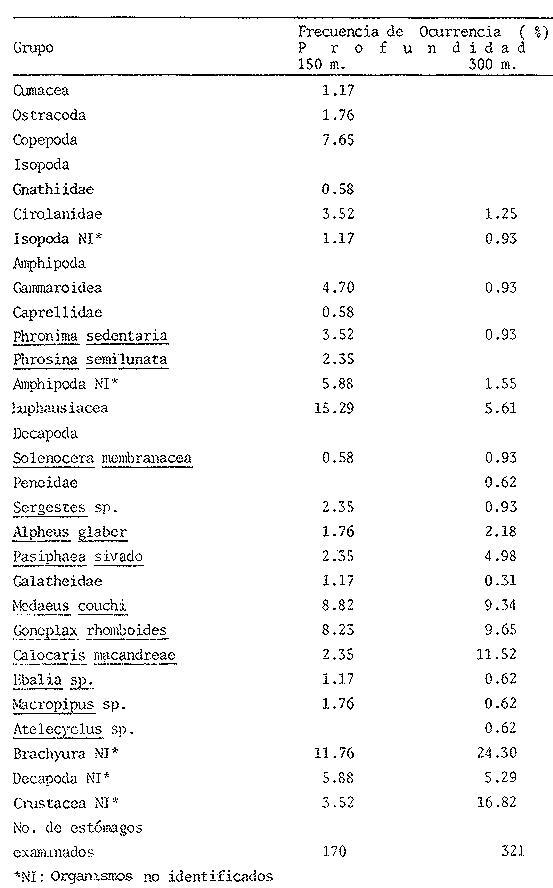 TABLA 9 VALORES DE LA FRECUENCIA DE OCURRENCIA DE CRUSTÁCEOS ENCONTRADOS EN EL CONTENIDO ESTOMACAL DE Nephrops norvegicus EN RELACIÓN A LA PROFUNDIDAD. Por otra parte aumenta el índice de ocurrencia para los decápodos carideos como son Alpheus glaber (2.18%) y Pasiphaea sivado (4.98%). Los decápodos del grupo Pleocymeata alcanzan sus máximos valores, como es el caso de Medaeus couchi (9.34%), Goneplax rhomboides (9.65%) y Calocaris macandreae (11.52%). Esta última especie se localiza preferentemente a los 300 m de profundidad (Zariquiey, 1935). A los 150 m de profundidad, la dieta es más variada y la alimentación más activa. En la Tabla 10, se muestran los valores del índice de repleción gástrica y de diversidad en número de componentes de la dieta. El índice de diversidad en número de componentes de la dieta, alcanza su mayor valor, a los 150 m de profundidad. La alimentación es más activa y aumenta el índice de repleción gástrica. Es mayor la proporción de presas pelágicas o bentopelágicas como son: copépodos, anfipodos, eufausíáceos y Sergestes sp. A los 300 m de profundidad descienden los valores de los índices de diversidad en número de componentes de la dieta y de repleción gástrica. Es mayor la proporción de organismos que viven en los fondos como son: Calocaris macandreae, Goneplax rhomboides y Medaeus couchi. RITMOS ALIMENTARIOSEn el presente estudio se observaron diferencias en el número de individuos capturados durante los muestreos, con relación a la profundidad y durante el ciclo estacional. En la Tabla 11 se muestran los resultados obtenidos. Durante la primavera, las poblaciones de la cigala son más abundantes a una mayor profundidad. Este fenómeno se acentúa notablemente durante los meses del verano, en los que solo el 10% de los individuos habitan a una profundidad de 150 m. Durante el otoño y el invierno, las poblaciones se distribuyen en igual proporcion en ambas profundidades. Höglund (1942) obtiene resultados semejantes en sus estudios sobre la cigala en el Atlántico Norte. Dicho autor reporta migraciones verticales en áreas de menor profundidad durante el invierno y de mayor profundidad durante el verano. En el presente estudio, se considera que dichos movimientos están relacionados con la dinámica de las masas de agua durante el ciclo anual -como es el caso del establecimiento o la ruptura de la termoclina- con la consecuente estratificación o mezcla de dichas masas de agua. Margalef (1972) reporta que las termoclinas constituyen zonas de mayor abundancia para algunos organismos, o bien, barreras inferiores o superiores a las migraciones de ciertas especies. Por lo que la formación de la termoclina durante el verano, puede disminuir las migraciones verticales de la cigala (de mayor a menor profundidad) o bien los movimientos migratorios (de menor a mayor profundidad) de las presas batipelágicas que sirven de alimento a esta especie. Contrariamente, durante el transcurso de las otras estaciones -en que se produce la mezcla de las aguas, desciende la temperatura y aumenta la salinidad en las capas superiores- la distribución de Nephrops norvegicus tiende a ser homogénea, presentándose períodos en que la alimentación es más activa. 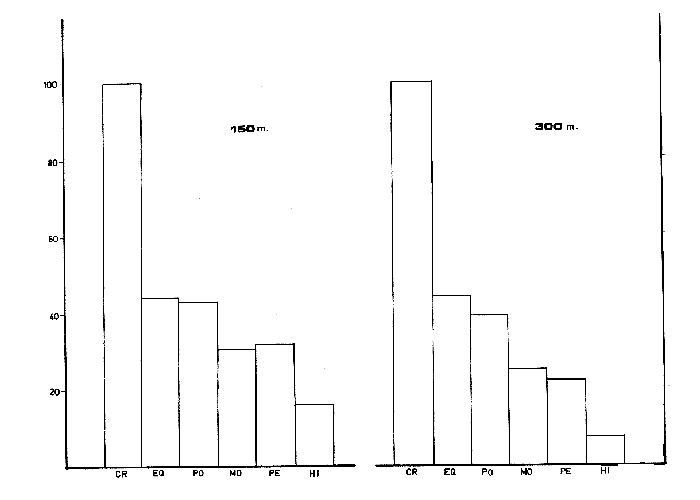 Figura 3. Relación de los valores del Indice de Ocurrencia de los organismos encontrados en el contenido estomacal de Nephrops norvegicus con respecto a la profundidad (CR = crustáceos; EQ = equinodermos; PO = poliquetos; MO = moluscos; PE = peces y HI = hidrozoarios). Por otra parte, también debe considerarse que las migraciones verticales de la cigala pueden corresponder a fenómenos fisiológicos más complejos, en los que influyen ritmos endógenos con un cierto grado de independencia de parámetros físicos y químicos. En relación a ritmos de actividad alimentaria diaria, algunos autores determinan que la cigala presenta una actividad máxima durante las horas del alba y del anochecer a profundidades de menos de 100 m. Dichas observaciones están basadas en que Nephrops norvegicus abandona su madriguera (túnel subterráneo de 40 cm de longitud, aproximadamente) para buscar su alimento. En el transcurso de estas horas, en que los organismos se encuentran sobre la superficie del fondo, pueden ser más fácilmente capturados por las redes de arrastre. Durante lapsos de tiempo de mayor intensidad luminosa u obscuridad, las capturas son menos abundantes (Chapman et al., 1971; Höglund et al., 1965; Fontaine et al., 1969; y Simpson, 1961). En el presente estudio, como ya se ha mencionado, los organismos fueron obtenidos a profundidades de 150 y 300 m, en las cuales la luz ya no juega un papel importante. Las máximas capturas fueron obtenidas a una profundidad de 300 m entre las 12 y 14 horas. Es posible señalar que el ritmo alimentario diario de la cigala, esté íntimamente relacionado con las migraciones nictemerales de las presas, existiendo una sincronización de las horas de mayor actividad entre depredador-presa. A poca profundidad, es posible que Nephrops norvegicus se alimente durante las horas nocturnas, debido a las mayores concentraciones del macroplancton en las capas superiores. A mayor profundidad es posible que los horarios de mayor actividad alimentaria de las capas superficiales, se desplacen en función de las migraciones verticales de las presas a capas más profundas durante el día. 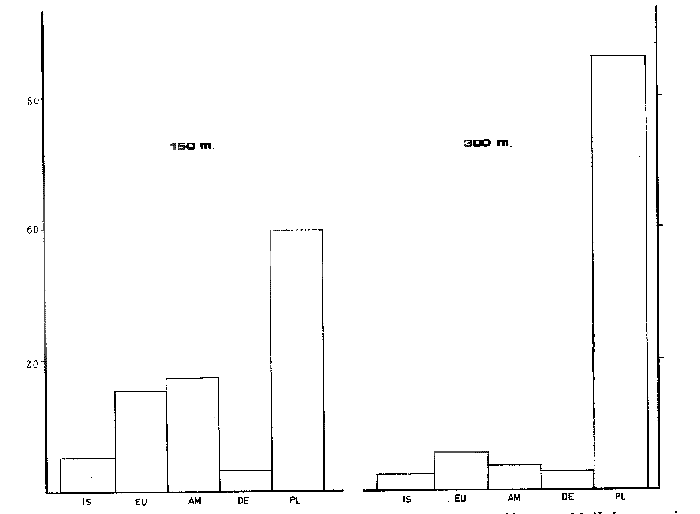 Figura 4. Relación de los valores del Indice de Ocurrencia de los organismos encontrados en el contenido estomacal de Nephrops norvegicus con respecto a la profundidad (IS = Isopoda; EU = Euphausiaecea; AM = Amphipoda; DE = Dendrobranpcehiata y PL = Pleocymeata).  TABLA 10 INDICES DE DIVERSIDAD EN NÚMERO DE COMPONENTES DE LA DIETA Y DE REPLECIÓN GÁSTRICA EN RELACIÓN A LA PROFUNDIDAD. HÁBITOS ALIMENTICIOSDe acuerdo con los resultados obtenidos en el presente estudio, se puede decir que Nephrops norvegicus es básicamente un depredador macrófago, oportunista y que presenta una dieta altamente eurífaga, constituida por los recursos batipelágicos y bentónicos disponibles en los fondos de arena y fango del Mar Catalán  TABLA 11 PORCENTAJE DE E.TEMPLARES CAPTURADOS EN RELACION A LAS ESTACIONES Y A LA PROFUNDIDAD. Secundariamente, cuando el alimento escasea, se comporta como un ramoneador, alimentándose de organismos que desprende de rocas y conchas, como es el caso de los hidrozoarios. Como la mayoría de los decápodos, la necrofágia ocupa un lugar importante en el régimen alimenticio de la cigala, en función de la disponibilidad del alimento. Debido a que los hábitos alimenticios de Nephrops norvegicus no son selectivos, los resultados obtenidos en el presente análisis pueden representar, de una manera aproximada, la abundancia estacional y/o batimétrica de los distintos organismos o grupos que habitan en los fondos arenosos del bentos, en las áreas y profundidades consideradas. ConclusionesEl régimen alimenticio de Nephrops norvegicus no presentó diferencias significativas en relación al sexo y a las clases de tallas. Los crustáceos fueron los organismos más abundantes en el contenido estomacal de la cigala, durante todas las estaciones del año y a diferentes profundidades. Le siguen en importancia los equinodermos, los poliquetos, los moluscos y los peces, principalmente. Las especies identificadas con mayor frecuencia fueron los decápodos: Genoplax rhomboides, Medaeus couchi y Calocaris macandreae. Las épocas en que la alimentación es más activa, corresponden a la primavera y al otoño, periodos en que es mayor el índice de diversidad en el número de componentes de la dieta. A los 150 m de profundidad, la dieta es más variada y la alimentación más activa, siendo mayor la proporción de presas pelágicas y bentopelágicas que a 300 m. La proporción de organismos que viven en los fondos es mayor conforme aumenta la profundidad. Durante los meses de la primavera y el verano, las poblaciones de Nephrops norvegicus son más abundantes a los 300 m que a los 150 m de profundidad. Durante el otoño y el invierno dichas poblaciones se distribuyen en igual proporción en ambas profundidades. Las máximas capturas fueron obtenidas durante las 12 y 14 horas a profundidades de 300 m. Nephrops norvegicus es un depredador macrófago, eurífago y oportunista. Secundariamente se comporta como un ramoneador. AgradecimientosA Ramón Margalef, Catedrático de la Universidad Central del Barcelona, por su apoyo para la realización del presente estudio. A A. Laguarda Figueras, Investigador del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, por la revisión del presente trabajo. A F. Sardá, J. Camp, C. Baz y J. Lleonera, miembros del grupo de recursos del Instituto de Investigaciones Pesqueras de Barcelona, por su cooperación para la publicación del mismo. A Francisco Xavier Chiappa Carrara, A. Durán y A. 01medo, por la ayuda prestada para la realización del presente trabajo. LITERATURABARNES, R., Zoología de los Invertebrados. Ed. Interamericana, S.A. México. 1968. 761 p. BOWMAN, T. y L ABELE, The Bioloy of Crustacea. 1. Classification of recent crustacea. Abele, L (Ed.). New York: Academic Press 1982. 1-27. BROOKES, J., L R. COY, A. KEEN, A. G. SMITH, P- L BATTEN, E. L YOCHELSON, N. H. LUDBROOK R. ROBERTSON, C. M. YONGE y R. C. MOORE, Mollusca 1. Treatise on Invertebrate Paleontology. Part I. R. C. Moore (Ed.). University of Kansas Press. Laurence, Kansas. 1960. 1351 p. CHAPMAN G.J. y A.L. RICE, Nephrops novegicus Mar, Biol., Some direct observations on the ecology and behaviour of the Norway lobstu, 1971. 321-329. 10 (4) FAUCHALD, K., National History Museum of Los Angeles Conty. The Polychacta wonns. definítions and keys to the orders, families and genera. Sciences Series 1977. 1-90. 28: FIGUEIREDO, M. J. y H. J. THOMAS, Oceanogr Mar. Biol Ann Rev., Nephrops norvegicus (L 1758) Leach-a review. 1967. 371-407. 5: FONTAINE, B. y N. WARLUZEL, Rev. Trav. Inst Peches Marit., Biologie de la Langoustine. Nephrops norvegicus (L.) 1969. 223-246. 33 (2): FRANQUEVILLE, C. Tethys Macroplancton profond (Invertebrés) de la Mediterranée Nord-occidentale. 1971. 11-56. 3 (1): HÖGLUND, H. Nephrops norvegicus (L). F. Fiske. Nor., Havskräftan eller Kejsarhummern, 1942. 293-296. 1: HÖGLUND, H. y B.I. DYBERN, Coast. Int. Explor. Mer., Diurnal and seasonal variations in the catch-composition of Nephrops norvegicus (L.) at the Swedish west 1965. 1-9. 146: LAGARDÈRE, J.P., Bull Centr. Et Rech. Scient. Biarritz Recherches sur la distribution verticale et sur l'alimentation des crustacés décapodes benthiques de la pente cotinentale du golfé de Gascogne. Analyse des groupements carcinologiques. 1977. 387-439. 11 (4): LEVINE, N.D., J.C. CORLISS, F.E.G. COX, G. DROUX, J. GRAIN, B. M. HONIGBERG, G. F. LEEDAI.E A. R. LOEBLICH, J. LOM, D. LYNN, E. G. MERINFELD, F. C. PAGE, G. POLJANSKY, V. SPRANGUE, J. VAVRA y F. G. WALLACE, Protozool, A newly revised classification of the Protozoa. 1980. 37-58. 27 (1): LEVINS, R., Evolution in Changing Environments. The theory of the niche. R. H. MacArthur (Ed.). Princeton University Press: 1968. 39-66. MARGALEF, R., Ecologia Marina. Ritmos, fluctuaciones y sucesión. Fundación La Salle (Ed.). Edit. Dossat. Caracas: 1972. 454-492. MARGALEF, R. Y J. CASTELVI, Inv. Pesq., Fitoplancton y producción primaria de la costa catalana, de julio de 1966 a julio de 1967. 1967. 491-502. 31 (3): MC NEILL, A., The Chordates. Cambridge Univ. Gran Bretaña: 1975. 20. SIMPSOM, A.C., Intem Council Expl. Sea. Diurnal variation in the catches of Nephrops norvegicus. 1961. 1-2. 102: THOMAS, H. J. y C. Davidson, Mar. Res. The food of the Norway lobster. 1962. 5-15. 3: YONGE, C. M. Bris. J. Exptl. Biol., The mechanism of feeding, digestion and assimilation in Nephrops novegicus. 1924. 343-389. 1: ZARIQUIEY, R., Bull. Inst. Cat. Hist. Nat., Crustáceos del Mediterráneo (Decápodos Macruros) Nephrops norvejícus, Linné, var. meridionalis. 1935. 1-7. 35:
|