|
ABUNDANCIA DE ALGUNAS ESPECIES DE ANCHOAS EN LA LAGUNA DE
TÉRMINOS (MÉXICO), ESTIMADA A TRAVÉS DE LA CAPTURA DE
HUEVOS
Trabajo recibido el 25 de febrero de 1986 y aceptado para su
publicación el 3 de abril de 1987.
César Flores Coto
Alberto Ocaña Luna
Alfonso Luna Calvo
Faustino Zavala García
Universidad Nacional Autónoma de
México. Instituto de Ciencias del Mar y Limnología. A.P. 70-305,
04510 México, D.F. Contribución 612 del Instituto de Ciencias del
Mar y Limnología, UNAM.
Dentro del programa del Laboratorio de Zooplancton del
ICMI (UNAM), tendiente al conocimiento de las comunidades
ictioptanctónicas de las lagunas costeras del Golfo de México se
llevan a cabo una serie de análisis preliminares de evaluación de
adultos que desovan en ellas; a través de los productos de desove. En
este trabajo se atiende a los engraulidos de la Laguna de Términos,
Campeche. El análisis está basado en 229 muestras de zoopiancton
colectadas de junio de 1981 a mayo de 1982. Las especies registradas fueron
Cetengraulis edentulus, Anchoa mitchilli y A.
hespesetus y su biomasa se estimó como el cociente de la
producción anual de huevos entre el producto de la fecundidad relativa y
la proporción de hembras en la población. Los resultados arrojan
para C. edentulus una fecundidad relativa (Fr) de 585.5
ov/gr, proporción de hembras (K) de 0.59, talla mínima de madurez
gonádica (mg) de 97.5 mm y una biomasa, bajo dos métodos
empleados, de 10,303 y 11,034 toneladas métricas. Para A.
mitchilli Fr = 823.8, K= 0.48 y mg = 37 mm y una biomasa de 7,046 y
7,967 ton. métricas. A. hepsetus tuvo valores de Fr =
1,298, K = 0-50, mg = 85.5 mm y una biomasa de 15.87 y 16.44 ton.
métricas. La gran biomasa de engraulidos confirma el carácter de
dominante de este grupo en las lagunas costeras del Golfo de México y lo
muestra como un recurso potencial con un alto significado ecológico en
la dinámica de estos sistemas.
Comprised in the program concerning to knowledge of
the ichthyoplankton communities from coastal lagooris of the Gulf of Mexico,
realized by the zooplankton laboratory from the ICMI, (UNAM), some preliminary
evaluation analysis of the adults that spawn in them are being made in basis on
the spawn products. This paper deals with the Engraulids from the Terminos
Lagoon Campeche. The analysis is based upon 229 zooplankton samples collected
from june 1981 to may 1982. The registered species were: Cetengraulis
adentulus, Anchoa mitchilli and A. hespselus and
their biomass was estimated as the quotient of annual egg production between
the product of the relative fecundity and the proportion of females of the
population. The resoults show that for C. edentulus the
relative fecundity (Fr) was 585.5 ov/gr, females proportion (K) of 0-59,
minimum size of gonadic maturity (mg) of 97.5 mm and a biomass using two
different methods, of 10,303 and 11,034 metric tons. For A.
mitchilli Fr = 823.8, K = 0.48, mg = 37 mm and a biomass of 7,046 and
7,967 metric tons. A. hepsetus showed values of Fr = 1,298,
K = 0.50, mg = 85-5 mm with a biornass of 15.87 and 16.44 metric tons. The high
biomass of engraulids confirms the dominant character of this group in the
coastal lagoons of the Gulf of Mexico and shows that this is a potential
resource with a significatively high ecological importance in the dinamic of
this system.
El estudio de las etapas de desarrollo de los peces y los censos de huevos y larvas se muestran como métodos accesibles y económicos para obtener información de las comunidades ícticas, sobre áreas, épocas y densidad de desove de las especies y constituyen además una vía para el cálculo de la biomasa de individuos adultos. Dentro del programa del Laboratorio de Zooplancton del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, tendiente al conocimiento de las comunidades ictioplanctónicas de las Lagunas costeras del Golfo de México se ha orientado parte de la información obtenida durante varias campañas para iniciar, por medio del censo de huevos, una serie de análisis preliminares de evaluación de la biomasa de adultos que desovan en las lagunas y que presentan un recurso pesquero actual o potencial. A la fecha sólo se han terminado un par de trabajos el primero sobre Archosargus rhomboidalis (Chavance et al., 1984) y el segundo sobre Bairdiella chrysours (Sánchez Iturbe y Flores Coto, 1986) ambos de la Laguna de Términos. Algunas especies de engraulidos constituyen una importante riqueza, particularmente en aguas costeras, como es el caso de Engraulis ringens la anchoveta peruana, Cetengraulis edentulus en el oriente de Venezuela y Engraulis mordax o anchoveta norteña en el noreste de México. En el Golfo de México diversos estudios han sido enfocados a evaluar la abundancia y potencial para las pesquerías de especies de anchoas, anchovetas y sardinas, (Houde, 1976; 1977a; 1977b; 1977c). De acuerdo a Olvera-Limas y Cid del Prado (1983), en el Banco de Campeche, este grupo de peces está considerado como uno de los recursos pesqueros potenciales más abundantes, los cuales pueden llegar a constituir pesquerías que ofrezcan posibilidades de diversificar la pesca en esa región, ya que a la fecha la explotación es a nivel regional e incide sobre larvas y juveniles, vendiéndose el producto secado y salado en forma de charal o como harina de pescado. Para el presente trabajo se ha escogido a las especies de anchoas que desovan en la Laguna de Términos para la evaluación de su biomasa desovante, considerando que algunas de ellas representan un recurso pesquero potencial y por presentar los engraulidos la mayor abundancia larvaria entre toda la comunidad ictioplantónica. ÁREA DE ESTUDIOLa Laguna de Términos está situada al sur del Golfo de México (Fig. 1). Diversos aspectos descriptivos, geológicos, físicos, químicos y biológicos, pueden encontrarse en: Phleger y Ayala-Castañares (1971), Gómez Aguirre (1974), Vázquez-Botello (1978), Flores Coto y Alvarez-Cadena (1980), Mancilla-Peraza y Vargas-Flores (1980), Yáñez-Arancibia et al. (1980), Reséndez (1981a, 1981b), entre otros. Los aspectos más importantes de su fisiografía se pueden resumir así: Es un cuerpo elíptico con alrededor de 70 km en su parte más larga y 28 en la más ancha. Se conecta al mar por dos bocas. Su profundidad media es de cerca de 2.5 m, con sus mayores profundidades (3.5 m) en la parte central y canales de las bocas que alcanzan más de 10 m. En su parte interna descargan tres importantes ríos, Candelaria, Chumpan y Palizada. MATERIALES Y MÉTODOSSe estableció una red de 24 estaciones de muestreo distribuidas homogéneamente (Fig. 1). Las colectas se efectuaron mensualmente desde agosto de 1981 hasta mayo de 1982, cubriéndose el área en tres días. Se utilizó una red tipo trapecio con malla de 250 um, manga de 1.5 m, con diámetro de boca y copo de 50 y 10 cm respectivamente. En la boca se colocó un flujómetro. Los arrastres fueron horizontales durante cinco minutos, en trayectoria circular, entre 10 y 40 cm bajo la superficie del agua. La salinidad se midió con refractómetro y la temperatura, con un termómetro de cubeta. Las muestras se fijaron con una solución de formalina al 5% neutralizada con borato de sodio; posteriormente en el laboratorio se extrajeron el total de huevos, separándose los correspondientes a la familia Engraulidae, para su determinación específica. Los datos se estandarizaron para expresarse como huevos por m³. La abundancia de las especies o biomasa adulta se estimó de acuerdo con Houde (1977a): B = PA/fk donde: B = Biomasa adulta; PA = Producción anual de huevos; f = Fecundidad relativa; K = Proporción de hembras en la población La metodología utilizada en el procesamiento de datos para la obtención de la producción anual de huevos, se basó en los trabajos de Sette y Ahlstrom (1948), Kramer et al., (1972), Houde (1977a), Smith y Richardson (1977) y particularmente los de Chavance et al. (1984) y Sánchez Iturbe y Flores-Coto(1986), quienes trabajaron sobre otras especies en la misma laguna, trasformando del método de polígonos de Sette y Ahlstrom (1948) el valor de área (m²) a volumen (m³). 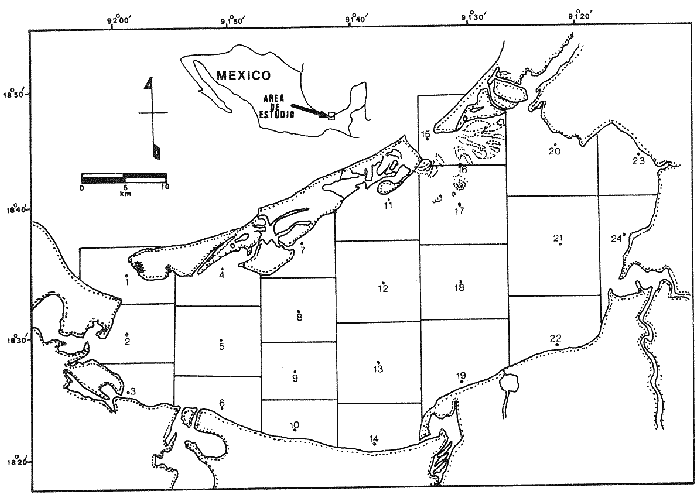 Figura 1. Laguna de Términos. Delimitación de areas que representan las estaciones de muestreo. La producción anual requirió de la estimación del número de huevos en cada estación, el cálculo de la abundancia en la laguna y la producción diaria y por campaña. La estimación de la abundancia de huevos en cada periodo de colecta se calculó por dos métodos (Chavance et al., 1984), en el primero se tomó la densidad promedio por metro cúbico en la laguna y se multiplicó por el volumen total de esta, estimado 3 en 4.1 x 109 m, que resulta de la sumatoria de volúmenes de cada estación de muestreo asumiendo una profundidad de 2.5 m (Tabla 1). En el segundo método se calculó multiplicando la densidad de huevos de cada estación por el volumen que representa, efectuando la sumatoria del total de estaciones.  TABLA 1 VOLUMEN ESTIMADO PARA VADA ESTACIÓN DE MUESTREO. La producción diaria se obtuvo dividiendo la abundancia entre el tiempo de residencia de los huevos. La producción por campaña se obtuvo multiplicando el valor de la producción diaria por el número de días que represente la campaña. El tiempo de campaña se estimó sumando a los días de muestreo, la mitad de días desde que fue realizada la campaña precedente y la mitad de días antes del inicio de la siguiente; así la producción anual se obtuvo como la suma de la producción de las diferentes campañas. RESULTADOSDurante los 10 meses de muestreo se colectaron un total de 79,823 huevos de engraulidos, encontrándose 5 morfotipos, de los cuales sólo tres pudieron ser determinados a nivel específico, en base a las descripciones de Simpson (1965), Mansueti y Hardy (1967) y Jones et al. (1978). Del total de huevos 40,155 correspondieron a Anchoa mitchilli, 229 a A. hepsetus, 36,102 a Cetengraulis edentulus y 3,337 indeterminados. FECUNDIDAD Y PROPORCIÓN DE SEXOSComo un paso necesario para la evaluación de la abundancia de las especies, se calculó la proporción de hembras en la población de cada especie, así como sus fecundidades absoluta y relativa. Para A. mitchilli se disectaron un total de 250 especímenes, encontrándose 129 machos y 121 hembras, lo que arroja una proporción de hembras de 0.48; de éstas se sacaron las gónadas y sólo seis estuvieron en condiciones de poder ser analizadas para la obtención de la fecundidad absoluta, la que se determinó para cada una, como el número de ovocitos mayores de 132 µm, considerando a los de menor talla como inmaduros. De A. hepsetus se disectaron un total de 51 especímenes de los que fueron 25 machos y 26 hembras, lo que arroja una proporción de hembras de 0.5098; de las que se extrajeron las gónadas, encontrándose sólo 7 maduras. La fecundidad absoluta se determinó como el número de ovocitos mayores de 165 µm. Se disectaron 126 especímenes de C. edentulus, encontrándose 75 hembras y 51 machos, lo que corresponde a una proporción de 0.59 hembras; de éstas sólo 5 estuvieron en condiciones de ser analizadas para la obtención de la fecundidad absoluta, la que se determinó como el número de ovocitos mayores de 165 µm. Los resultados individuales de fecundidad absoluta y relativa son anotados en la Tabla 2, y a fin de obtener la talla mínima de madurez gonádica y la fecundidad relativa por especie, se realizaron regresiones lineales entre la fecundidad absoluta y longitud patrón y peso lleno (Fig. 2). Las tallas mínimas de madurez gonádica y la fecundidad relativa por especie, fueron respectivamente : 37.5 mm y 823.8 ov/gr para A. mitchilli, 85.5 mm y 1298 ov/gr para A. hepsetus y 97.5 mm 586.5 ov/gr para C. edentulus. 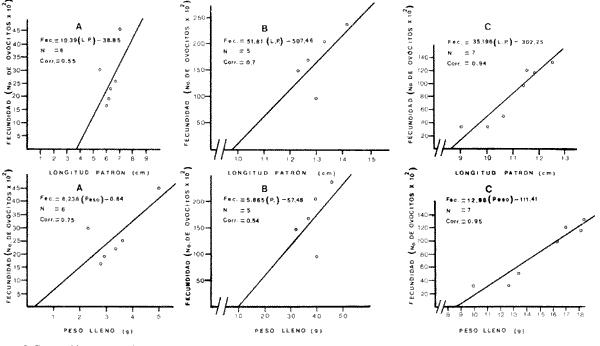 Figura 2. Correlación entre el número de ovocitos y la longitud patrón y peso lleno de las especies Anchoa mitchilli (A), Centengraulis adentulus (13) y Anchoa hepsetus (C). La fecundidad relativa a A. mitchilli, 823.8 ov/gr, fue un valor muy próximo al promedio de fecundidades individuales de 807.7 ov/gr (Tabla 2) y ambos resultaron muy similares al registrado por Gaspar-Dillanes y Sánchez-Iturbe (1985) para la misma especie, de la Laguna de Tamiahua. La talla mínima de madurez, 373 mm, resultó también muy similar a lo señalado para otras áreas, como en Tamiahua con 38.5 mm (Gaspar-Dillanes y Sánchez-Iturbe, 1985), o en las costas atlánticas de los Estados Unidos para donde Jones et al. (1978) registran tallas entre 34 y 40 mm. La talla mínima de madurez de A. hepsetus, 85.5 no difiere mucho de los 75 mm que señalan Jones et al. (1978), para las costas atlánticas de los Estados Unidos. Simpson et al. (1965), señalan la presencia de especímenes de C. edetulus sexualmente maduros de 32 a 40 mm, en aguas venezolanas, tallas mucho menores que la de 95 mm, obtenida para esta especie en la Laguna de Términos. 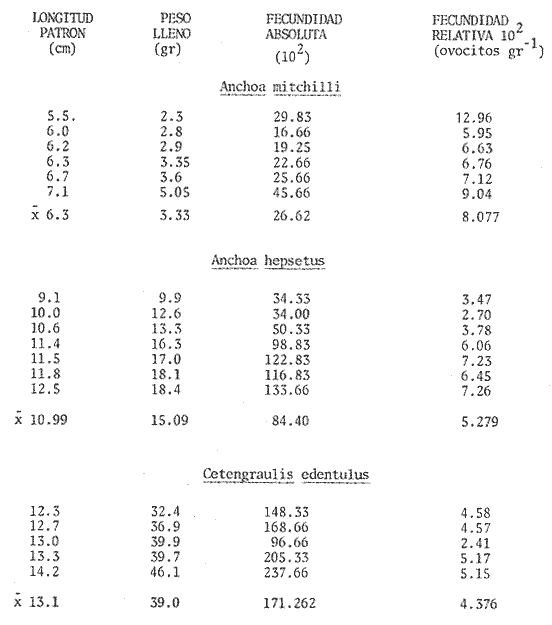 TABLA 2 FECUNDIDAD ABSOLUTA Y RELATIVA DE LAS HEMBRAS ANALIZADAS DE Anchoa mitchilli, A. hepsetus y centengraulis edentulus. 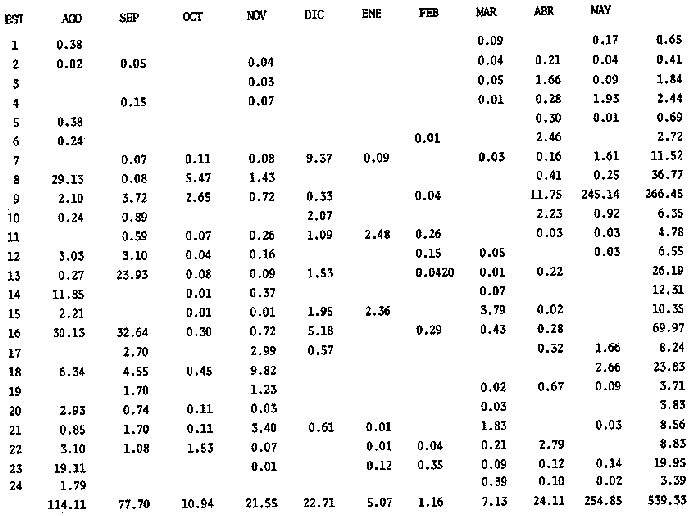 TABLA 3 Anchoa mitchilli, NÚMERO DE HUEVOS POR m³ DURANTE CADA CAMPAÑA Y ESTACIÓN DE MUESTREO ESTIMACIÓN DE LA BIOMASA DESOVANTEA partir de los registros primarios de huevos de cada especie, estandarizados a huevos/m³, (Tablas 3, 4 y 5), y siguiendo la metodología descrita, se calcularon la abundancia mensual (Ab), producción diaria (Pd) y por campaña (Pc), para obtener finalmente la producción anual de huevos (PA), que representa la sumatoria de todas las campañas (Tablas 6, 7 y 8). La abundancia de Anchoa mitchilli en la laguna, estimada como biomasa desovante, se hizo en base a los siguientes valores: producción anual de huevos (PA) que bajo los dos métodos fue de 2.7861 x 1012 y 3.1505 x 1012; fecundidad relativa Fr = 823.8 ovocitos/gr de hembra; y la proporción de hembras en la población adulta, K= 0.48, con la que la biomasa estimada (B) fue de 7,046 y 7,967 Ton métricas según el primero y segundo método respectivamente Para Anchoa hepsetus los valores fueron: PA = 1.0298 x 1010 y 1.067 x 1010, bajo los dos métodos; Fr = 1298 huevos/gr; K = 0.5098; B = 15.87 y 16.44 Ton métricas. En el caso de Cetengraulis edentulus los valores fueron: PA = 3.5966 x 1012 y 3.8519 x 1012; Fr = 586.5 ov/gr; K = 0.5952; B = 10303 y 11034 Ton métricas. 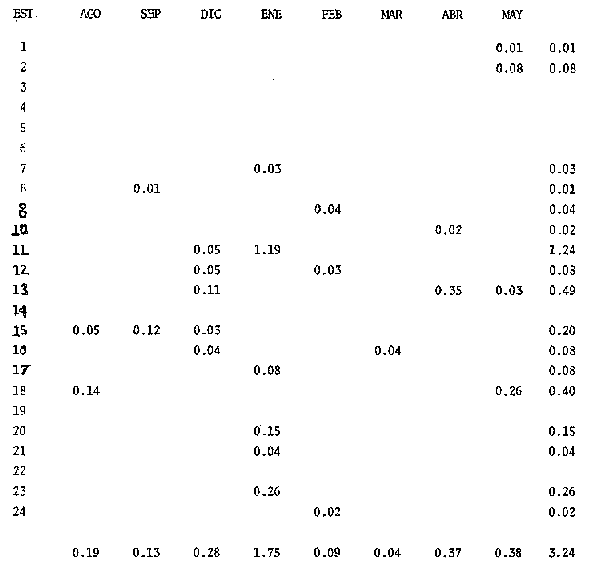 TABLA 4 Anchoa hepsetus, NÚMERO DE HUEVOS POR m³ DURANTE CADA CAMPAÑA Y ESTACIÓN DE MUESTREO 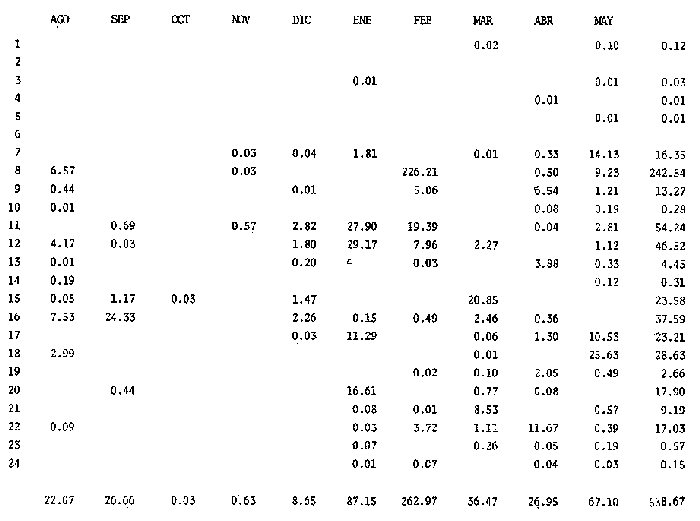 TABLA 5 Centengraulis edentulus, NÚMERO DE HUEVOS POR m³ DURANTE CADA CAMPAÑA Y ESTACIÓN DE MUESTREO 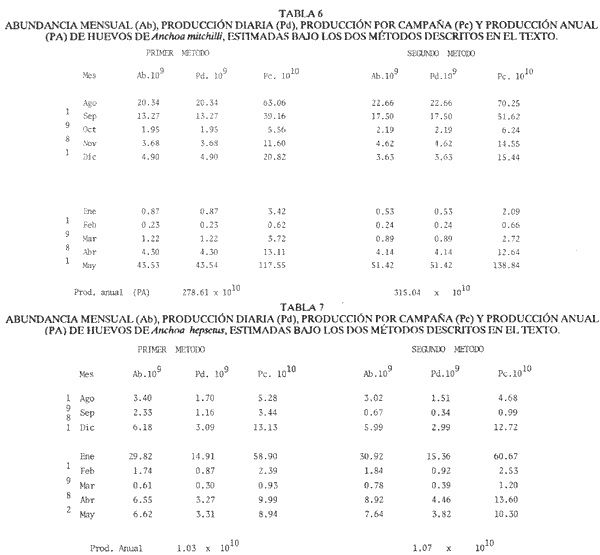 TABLA 6 ABUNDANCIA MENSUAL (Ab), PRODUCCIÓN DIARIA (Pd), PRODUCCIÓN POR CAMPAÑA (Pc) Y PRODUCCIÓN ANUAL (Pa) DE HUEVOS DE Anchoa mitchilli, ESTIMADAS BAJO LOS DOS MÉTODOS DESCRITOS EN EL TEXTO. 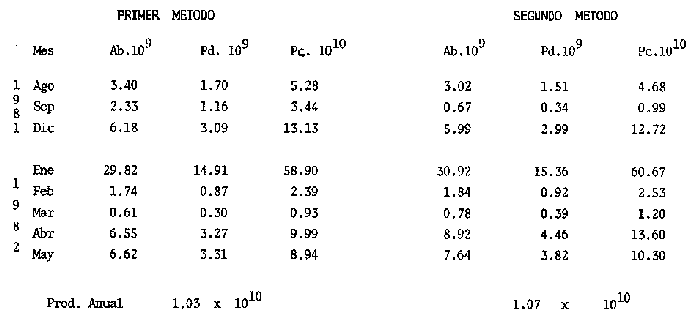 TABLA 7 ABUNDANCIA MENSUAL (Ab), PRODUCCIÓN DIARIA (Pd), PRODUCCIÓN POR CAMPAÑA (Pc), Y PRODUCCIÓN ANUAL (PA), DE HUEVOS DE Anchoa hepsetus, ESTIMADAS BAJO LOS MÉTODOS DESCRITOS EN EL TEXTO. DISCUSIÓNLos resultados del presente trabajo señalan que Cetengraulis edentulus aunque con la fecundidad relativa más baja de las tres especies, tuvo la mayor biomasa, lo que aunado a sus antecedentes, en los que se anota como abundante y ampliamente distribuida en la Laguna de Términos, particularmente en los sistemas fluviolagunares conectados a esta (Amezcua-Linares et al., 1980; Reséndez-Medina, 1981, Yáñez-Arancibia y Lara-Domínguez, 1983); la muestran como un recurso potencial importante. Anchoa mitchilli con una biomasa menor es también un recurso potencial considerable. A. hepsetus con una fecundidad relativa alta, tuvo no obstante una biomasa tan baja, que puede considerarse despreciable para la Laguna de Términos. Su escasez corresponde con sus antecedentes que indican, se trata de una especie escasa, o visitante ocasional de la Laguna (Amezcua-Linares et al. 1980; Reséndez-Medina, 1981; Yáñez-Arancibia y Lara-Domínguez 1983), que tiende a vivir en aguas de alta salinidad (Castro, 1978) y es más común en zonas alejadas de la costa (Hoese y Moore, 1979). En la Tabla 9 se resumen los resultados de diversos estudios para la estimación de biomasa a través de la captura de huevos, que se han desarrollado en el Golfo de México, en la plataforma continental de Florida (USA) como en Lagunas costeras de México (Houde, 1976; Chavance et al., 1983; Gaspar-Dillanes y Sánchez-Iturbe, 1985; Sánchez-Iturbe y Flores Coto, 1986). 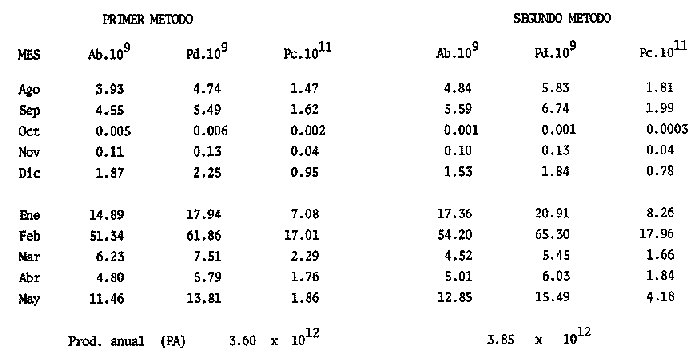 TABLA 8 ABUNDANCIA MENSUAL (Ab), PRODUCCIÓN DIARIA (Pd), PRODUCCIÓN POR CAMPAÑA (Pc), Y PRODUCCIÓN ANUAL (PA), DE HUEVOS DE Cetengraulis edentulus, ESTIMADAS BAJO LOS DOS MÉTODOS DESCRITOS EN EL TEXTO.  TABLA 9 BIOMASA DESOVANTE DE ALGUNAS ESPECIES EN EL GOLFO DE MÉXICO ESTIMADAS ATRAVÉS DE LA CAPTURA DE HUEVOS. *Valores promedio: Chavance et al. (1983); 2. Sánchez-Iturbe y Flores Coto (1986); 3. éste estudio; 4. Gaspar Dinamarqueses Sánchez-Iturbe (1985); S. Houde (1976). Aunque la comparación que puede hacerse de estos resultados no es absoluta, ya que las áreas son distintas tanto en tamaño como en su dinámica y ecología, permiten al menos algunas consideraciones. Los valores registrados por Houde (1976) para la plataforma de Florida corresponden a un área de poco más de un millón de kilómetros cuadrados, por lo que la biomasa de las Lagunas de Términos y Tamiahua, con alrededor de 1650 km² y 750 km² respectivamente, representan proporcionalmente un valor mucho mas alto. El que la evaluación haya sido sobre queridos en la plataforma y engraulidos en las lagunas no es fortuito, ya que la dominancia de engraulidos en lagunas costeras y estuarios, ya en su forma adulta, como juveniles o en sus etapas planctónicas, es un hecho común en el Golfo de México (Betchell y Copeland, 1970; Guillén y Landry, 1981; Reis y Dean, 1981; Sabins y Truesdale, 1974; Subrahmanyan y Coultas, 1980; Yáñez-Arancibia et al., 1980; Flores Coto, et al., 1983), por lo que su biomasa no sólo significa un gran potencial pesquero sino que ecológicamente debe representar uno de los eslabones más importantes de la cadena trófica en estos ecosistemas. Los engraulidos, los más de ellos de pequeña talla, planctófagos como carácter básico de su alimentación, representan un eficiente (además de numeroso) sistema concentrador de energía dispersa en los pequeños organismos del micro y mesozooplancton y por tanto no accesible a los carnívoros, muchos de los cuales representan recursos pesqueros actuales. Por otro lado, los engraulidos constituyen una vía importante de exportación del exceso de producción de las lagunas, ya por su salida a las áreas costeras o a través de la captura directa que hagan de ellos en sus diversas etapas, aquellas especies que penetran a las lagunas para alimentarse. Particularmente para la Laguna de Términos, Yáñez-Arancibia et al. (1980) señalan que de 127 especies de peces registrados a través de varios años, el 45% corresponde a visitantes ocasionales y 45% a visitantes cíclicos y de ellos la mayor parte son carnívoros de segundo o tercer orden y que utilizan la laguna como área de alimentación. De muchas de tales especies, como Synodus foetens, Strongylura marina, Hyporhamphus unifscialus, Bairdiella chrysoura, Cynoscion nebulosus y Trachinotus falcatus, entre otras, se sabe que un alto porcentaje de su dieta los constituyen los engraulidos (Carr y Adams, 1973). En resumen puede decirse que la gran biomasa de engraulidos registrada a través de la captura de huevos, los confirma como dominantes en lagunas costeras, en este caso en la Laguna de Términos y muestra a C. edentulus y A. mitchilli como un recurso potencial con un alto significado ecológico en la dinámica de estos sistemes. Los clupeidos por su parte parecen más abundantes en las zonas neríficas costeras, tanto en la plataforma de Florida (Houde 1976), como en los de Yucatán (0lvera Limas y Cid del Prado, 1983) y Campeche (San Vicente-Añorve, 1985). AgradecimientosLos autores desean expresar su agradecimiento a R. Román por las facilidades otorgadas durante el trabajo de campo a las autoridades del IMCL por el financiamiento de este proyecto y a E. López por mecanografiar el manuscrito. LITERATURAAMEZCUA-LINARES, F. y A. YÁÑEZ-ARANCIBIA, An. Centro. Cienc. del Mar y Limnol. Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces. Univ. Nal. Autón. México, 1980. 69-118. 7 (1): BAGENAL, T.B. y E. BRAUN, Methods for Assessment of Fish Production in Fresh Waters. IBP. Eggs and early life history In: T. Bagenal (Ed.). Black weil Scientific Pubis. Handbook, 3.3a. ed Oxford, London: 1978. 165-201. BETCHELL, T. J. y B. J. COPELAND, Cont. Mar. Sci., Fish species diversity indices as indicators of pollution in Galveston Bay, Texas. 1970. 103- 132. 15: CARR, W. E. S. y C. A. ADAMS, Trans. Amer. Fish. Soc., Food habits of juveniles marine fishes occupying, seagrass beds in the estuarine zone near Crystal River, Florida. 1973. 511-540. 102 (3): CHAVENCE, P., C. FLORES y A. SÁNCHEZ, Trans. Amer. Fish. Soc. Early life history and adulta biomasas of sea brea in the Términos Lagos, Southern Gulf of México. 1984, 166-177. 113: FLORES-COTO, C. y J. ÁLVAREZ-CADENA, An. Centro Cienc. del Mar y Limnol Estudios preliminares sobre abundancia y distribución del ictiopiancton en la Laguna de Términos, Campeche. Univ. Nal. Autón. México, 1980. 67-78. 7 (2): FLORES-COTO, C., F. BARBA-TORRES y J. SÁNCHEZ-ROBLES, Trans. Amer. Fish. Soc., Seasonal diversity, abundance and distribution of ichthyoplanckton in Tamiahua Lagoon, Western Gulf of México 1983. 247-256. 112: GASPAR-DILLANES, M. T. y A. SÁNCHEZ-ITURBE, Estimación de la biomasa desovante de Anchoa mitcilli Cuvier y Valenciennes, 1848 (Pisces: Engraulidae) y determinación de algunos parámetros ecológicos y poblacionales a partir de estudios ictiopianctánicos, en la Laguna de Tamiahua, Veracruz, México (1984/1985) Biología de Campo, Arca: Ciencias del Mar (Ecología), Fac. Ciencias, Dept. Biología, Univ. Nal. Autón. México. 1985. GÓMEZ-AGUIRRE, S., An Centro Cienc. del Mar. y Limnol. Reconocimientos estacionales de hidrología y plancton en la Laguna de Términos, Campeche, México (1964-1965). Univ. Nal. Autón. México 1974. 61-81. 1 (1): GUILLÉN, G. J. y A. M. LANDRY, Procedings of the annual Conference Southeastern Associadon of Fish and Wildlife Agencies, Species composition and abundance of ichthyoplankton at beachfron and saltmarsh enviroments. 1981. 388-403. 34: HOUDE, E. D. Proc. Gulf Caribb. Fish. Inst. 28th. ánnu Sess.: Abundance and potencial for fasheries development of some Sardiine-like Fishes in the Eastern Gulf of México 1976. 77-82 HOUDE, E. D., Fish. Bull., Abundance and potencial yield of the round Herring Entrumeus teres and aspects of its early life history in the Eastem Gulf of Mexico. 1977a. 61-89. 75 (1): HOUDE E. D., Fish. Bull. Abundance and potencial yield of the atiantic thread herring Optisthomea oglinum, and aspects of its early life history in the eastern Gulf of Mexico. 1977b. 493-512. 75 (3): HOUDE, E. D., Fish. Bull., Abundance and potential yield of the scales sardine Haengula jaguana and aspects of its early life history in the eastern Gulf of México 1977c. 613-628. 75 (3): JONES, W. P., F. D. MARTIN y J. D. HARDY, Development of fahes of the Mid-Atlantic Bight. An atlas of egg, larval and juvenile stages. Acipenseridae throug Ictaluridae r. Power Plant Project Office of Biological Services, Fish and Wildlife Service, U.S. Department of the Interior. 1978. 366 p. Vol. I. KRAMER, D., M. J. KALIN, G. STEVENSE, J. R. THRAILKILL y J. R. ZWEFEL, Collecting and processing data of fish eggs and larvac in the California current region. NOAA Tech Rep., NMFS. Circ., 1972. 1-38. 370: MANCILLA-PERAZA, M. y M. VARGAS-FLORES, An. Centro Cienc. del Mar y Limnol. Los primeros estudios sobre la circulación y el flujo neto a través de la Laguna de Términos, Campeche. Univ. Nal. Autón. México, 1980. 1-12. 7 (2): MANSUETI, A. J. y J. D. HARDY, An Atlas of Egg. Larvae and Juvenil Stages, Part I. Development of Fishes of the Chesapeake Bay Región. Natural Resources Institute, University of Maryland, Baltimore, 1967. 202 p. OLVERA-LIMAS, R. M.y A. CID DEL PRADO, Identificación de larvas y juveniles de las familias Engraulidae y Clupcidac, de las pesquerías de Puerto de Celestún, Yucatán. Ciencia Pesquera, Inst. Nal. Pesca Sría. Pesc. México, 1983. 1-13. 4: PHLEGER F. B. y A. AYALA-CASTAÑARES, Bull. An. Petrol. Geol, Processes and history of Terminos Lagoon, Mexico. 1971. 2130-2140. 55 (2): REIS, R. R. Y J. M. DEAN, Estuaries, Temporal variation in the utilization of an itertidal creek by anchovy (Anchoa mitchilli). 1981. 16-23. 4 (1): RESÉNDEZ-MEDINA, A., Biótica Estudio de los peces de la Laguna de Términos, Campeche, México. I. 1981a. 239-291. 6 (3): RESÉNDEZ-MEDINA, A., Biótica Estudio de los peces de la Laguna de Términos, Campeche, México. II. Ultima parte. 1981b. 345-430. 6 (4) SABINS, D. S. y F. M. TRUESDALE, Proc. Twenty-eight Ann. S.E. Assoc. Game and Fish. Comm, White Sulfur Saring. W.V. : Diel and seasonal ocurrence of inmature fishes in a Louisiana tidal pass. In: W. A. Rogers (Ed.). 1974. 161-170. SÁNCHEZ-ITURBE, A. y C. FLORES-COTO, An. Inst. Ciénc. del Mar y Limnol. Estimación de la Biomasa de Bairdiella chysoura (lacepede, 1802) por medio del censo de huevos y algunos aspectos de sus primeros estadios de la vida en la Laguna de Términos, Campeche. (Pisces: Sciacnidac). Univ. Nal. Autón. México 1986. (en prensa). SETTE, 0. E. y E. H. AHLSTROM., Sean Found Journ Mar. Res., Estimations of abundance of eggs of the Pacific pilchard (Sardinops caerulea) off southern California during 1940 and 1941. 1948. 511-542. 7 (3): SIMPSON, G. J., Serie Biológica Estudio de las primeras etapas de la Rabo Amarillo Cetengraulis edentulus (Cuvier) en el Oriente de Venezuela. 1965. 1-24. 1 (1): SMITH, P. E., Bull, The increase in spawing biomass of Northern anchovy Engraulis mordax Fish. 1972. 849-874. 70: SMITH, P.E. y S.L. RICHARDSON, Fish. Tech. pap., Standard techniques for polagie fish egg and larval surveys. FAO 1977. 1-100. 175: SUBRAHMANYAN, C. B. y C. L. COULTAS, Bull. Mar. Sci, Studies on animals communities in two north Florida salt marshes. Part III. Seasonal fluctuation of fish and macroinvertebrates. 1980. 790-818. 30 (4): VÁZQUEZ-BOTELLO, A., An. Centro Cienc. del Mar y Limnol. Variaciones de los parámetros hidrológicos en las épocas de sequía y lluvias (mayo y noviembre, 1974) en la Laguna de Términos, Camp., México. Univ. Nal. Autón México 1978. 159-178 5 (1):. YÁÑEZ-ARANCIBIA, A., AMEZCUA-LINARES, F. and DAY, J.W. Jr., Estuarine Perspectives. Fish community structure and function in Terminos Lagoon, a tropical stuary in the southem Gulf of Mexico. In: V. Kennedy (Ed.). Academic Press Inc., New York: 1980. 465-482. YANEZ-ARANCIBIA, A. y A. L. LARA-DOMNGUEZ, An. Inst. Cienc. del Mar y Limnol. Dinámica ambiental de la Boca de Estero Pargo y estructura de sus comunidades de peces en cambios estacionales y ciclos de 24 hrs. (Laguna de Términos, Sur del Golfo de México). Univ. Nal. Autón. México 1983. 85-116. 10 (1):
|