|
HÁBITOS ALIMENTICIOS DE PECES DEPREDADORES DEL
SISTEMA LAGUNAR HUIZACHE-CAIMANERO, SINALOA, MÉXICO
Trabajo
recibido el 3 de noviembre de 1986 y aceptado para su publicación el 8
de junio de 1987.
Gilberto Díaz González
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología, Lab.
Contaminación Marina.
Luis A. Soto
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología, Lab. de
Ecología del Bentos. Contribución 611 del Instituto de Ciencias
del Mar y Limnología, UNAM.
En el periodo comprendido de febrero de 1977 a
noviembre de 1978, se estudiaron los hábitos alimenticios de las seis
especies de peces depredadores más importantes del sistema lagunar
Huizache-Caimanero, Sinaloa: Ariopsis caendescens
(Günther, 1864), Cynoxion xanthulus (Jordán y
Gilbert, 1882), Centropomus robalito (Jordán y
Gilbert, 1882), Centropomus nigrescens (Günther, 1864),
Elops affinis (Regan, 1909) y Polydactylus
approximans ("Lay y Bennet, 1849). Para cada especie se determinaron
sus preferencias alimenticias. Los componentes del espectro alimenticio se
analizaron en función del régimen de precipitación y se
demostró una relación entre éste y la composición
de la dieta. Se reconoció la existencia de tres especies netamente
piscívoras (A. caemiescem, C. xanthulus y C.
nigrescens) y tres que se alimentan exclusivamente de camarones
peneidos (C. robalito, E. affinis y P.
approximans).
During the period between February 1977 and November
1978 the feeding habits of six species of the most important fish predators of
the Huizache-Caimanero lagoon system in Sinaloa, Mexico were studied.
Ariopsis caerulescens (Günther 1864),
Cynoscion xanthulus (Jordan and Gilbert, 1882),
Centropomus robalito (Jordan and Gilbert, 1882),
Centropomus nigrescens (Günther, 1864), Elops
afinis (Regan, 1909), and Polydactylus approximans
(Lay and Bennet, 1849). The preferential diet was determined for each species.
The trophic feeding habits were analyzed in relation to the precipitation
regime. A significant relationship between this factor and the composition of
the species diet was established. A. caerulescens, C.
xanthulus, and C. nigrecens are recognized as
exclusive fish predators while C. robalito, E. affinis and
P. approximans constitute important penaeid shrimp predators
in the area studied.
Las lagunas costeras mexicanas cubren una superficie de aproximadamente 1.5 x 106 ha, de las cuales el 57% corresponde al litoral del Pacífico (Mendoza, 1972). Estas áreas protegidas representan sistemas ecológicos de alta productividad biológica que proporcionan refugio y alimentación a organismos marinos que cuyo ciclo biológico es estuarino-dependiente. Este es el caso del sistema lagunar Huizache-Caimanero, localizado en el suroeste del Estado de Sinaloa el cual, sostiene una pesquería de camarón de considerable importancia (Chapa y Soto, 1969); dicho sistema sirve de hábitat a cuatro especies de camarones peneidos: Penaeus vannamei, P. stylirostris, P. califomiensis y P. brevirostris. La primera destaca como la más importante desde el punto de vista comercial, ya que soporta la producción camaronera en un porcentaje elevado (Soto, 1969; Menz, 1976). Entre los autores que se han avocado al estudio de la ictiofauna del sistema Huizache-Caimanero (Carranza, 1969a,b, 1970; Carranza y Amezcua, 1971a y b; González,1972; Amezcua, 1977; Warburton, 1978, 1979) se ha reconocido como fundamental la evaluación de las relaciones entre peces y otros organismos, particularmente los camarones peneidos. A este respecto, se da especial énfasis en la necesidad de estudiar las relaciones tróficas y los efectos de la acción depredadora de peces sobre especies de importancia comercial, con el propósito de contribuir al manejo racional de los recursos pesqueros propios del sistema lagunar referido. El presente estudio contempla como objetivo esencial, el aportar información sobre los hábitos alimenticios de las seis especies de peces reconocidas como los principales depredadores en el sistema lagunar Huizache-Caimanero: Ariopsis caerulescens (Günther, 1864); Cynoscion xanthulus (Jordán y Gilbert 1882); Centropomus robalito (Jordán y Gilbert, 1882); Centropomus nigrescens (Günther, 1864); Elops affinis (Regan, 1909) y Polydactylus approximans (Lay y Bennet, 1849). Así mismo, se desea reconocer el espectro alimenticio de cada una de las especies anteriores a través de las temporadas de precipitación y estío propias del área estudiada y analizar la proporción en la que se presentan los componentes alimenticios predominantes en la dieta, con el propósito de evaluar la acción depredadora de las especies seleccionadas sobre los camarones peneidos; para tal propósito se requiere establecer la influencia de la actividad trófica de los peces, sus adaptaciones y la composición por tallas de las poblaciones. ÁREA DE ESTUDIOEl sistema lagunar Huizache-Caimanero está situado entre los 22° 50' hasta los 23° 10' de latitud norte y desde los 105° 55' hasta los 106° 18' de longitud oeste (Fig. 1), al noroeste delimitado por el Río Presidio y al sureste por el Río Baluarte. Este sistema se localiza sobre la costa del Pacífico Mexicano, aproximadamente 25 km al sur de la ciudad de Mazatlán. El sistema está compuesto por dos lagunas; la laguna del Huizache en el noroeste y la laguna de Caimanero al suroeste. Ambas lagunas se encuentran unidas por un estrechamiento llamado Pozo de la Hacienda; esta comunicación acuática no es permanente a través de todo el año, pues el sistema se seca aproximadamente desde el mes de marzo hasta las primeras lluvias del mes de julio (Fig. 2). En la estación de sequía (mayo y junio) las salinidades alcanzan hasta 55°/oo en la Laguna de Caimanero y 125‰ en Pozo de la Hacienda, debido a las grandes acumulaciones de sal en esta época sobre todo en la parte sureste de Caimanero; cuando las Lagunas están llenas (agosto a diciembre) existe un gradiente de salinidad que fluctúa de 1 - 20‰ en Huizache de donde entra la mayor parte de agua dulce al sistema, 5‰ en Pozo de la Hacienda y 12 - 15‰ al sur de Caimanero (Menz, 1976). 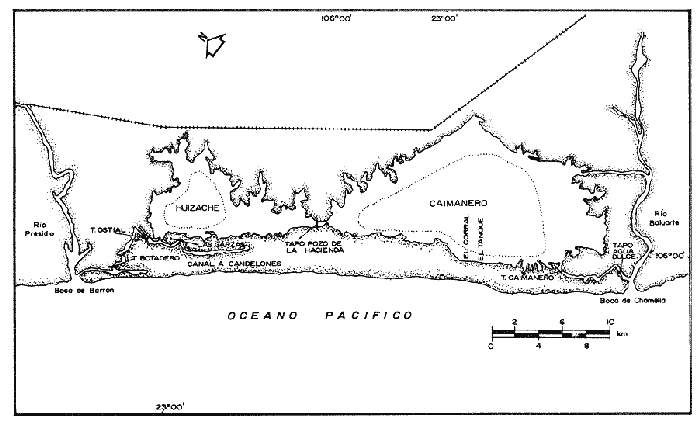 Figura 1. Sistema lagunar Huizache-Caimanero,Sinaloa, México. 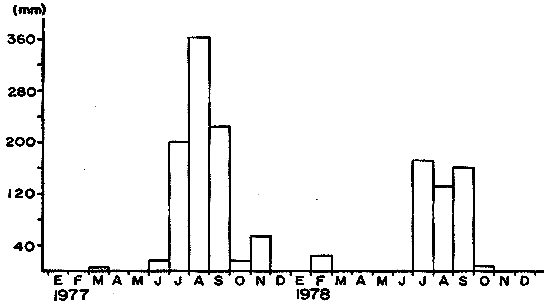 Figura 2. Régimen de precipitación durante el ciclo 1977-78 (Datos tomados de la SARH). La superficie aproximada de ambas lagunas al final de la época de precipitación es de 175 km2 (Soto, 1969; Menz, 1976; Paul, 1977; Moore, 1979) y 158 km² (Ayala et al., 1970); 134.3 km² correspondiendo a la Laguna de Caimanero y 40.7 km² a la Laguna de Huizache, respectivamente. De acuerdo con la modificación de García (1973), a partir del sistema de Koepen, el área presenta un clima cálido, subhúmedo y con precipitaciones en verano y en invierno, aunque en esta última estación las lluvias son mínimas, ya que están representadas por menos del 5% del total anual, superando los 750 mm. (Fig. 2). Se reconoce un régimen térmico invernal uniforme, no bien definido, con poca variación en la temperatura media mensual a través del año, la que siempre es superior a los 18° C. La diferencia entre el mes más cálido y el más frío oscila entre los 7 °C y 14 °C (Tamayo, 1949). La Laguna de Huizache tiene una dimensión de aproximadamente 6 km de ancho por 12 km de largo, con una profundidad máxima de 1.5 m. La Laguna de Caimanero mide 10 km de ancho por 20 km de largo con una profundidad máxima de 2.2 m. Al final de la época de sequía las cuencas lagunares pierden un 80 % de su área superficial y un 90 % de su volumen de agua (Moore 1979). La comunicación entre ambas lagunas y el mar se lleva a cabo a través de esteros, que son canales angostos y de profundidades variables dependiendo de la época y el estado de la marea; éstos están bordeados por mangles de las especies Rhizophora mangle, Laguncularia racemosa y Avicennia nitida. Las tres especies se encuentran asociadas y se extienden en ocasiones a más de 100 metros a ambos lados de los esteros (Chapa, 1966; Soto, 1969). El estero Ostial es el que comunica a la Laguna del Huizache y el Estero Agua Dulce a la Laguna de Caimanero. Los ríos Presidio y Baluarte descargan sus aguas a través de las bocas de Barron y Chametla, respectivamente. MATERIAL Y MÉTODOSDurante el periodo comprendido de febrero de 1977 a noviembre de 1978, se realizaron muestreos de ictiofauna con una frecuencia semanal, quincenal y mensual en nueve localidades del sistema Huizache-Caimanero. El tiempo de captura en cada localidad fue de 24 horas y el material obtenido se retiró cada dos horas. De las especies de Ariopsis caerulescens, Cynoscion xanthulus, Centropomus robalito, Centropomus nigrescens, Elops affinis y Polydactylus approximans colectadas, se analizaron los estómagos de 1,617 ejemplares, de los cuales 780 (47.24%) contenían alimento y 837 (51.76%) estaban vacíos. Para la obtención de las muestras, se utilizaron redes de diferentes aberturas de malla: a) dos redes agalleras (Red No. 1) de 40 metros de largo y 2.5 de alto, abertura 23 cm malla de hilo nylon y, (Red No. 2) de 50 metros de largo y 2.5 m de alto, con malla de hilo de seda y abertura de 5 cm y 9 cm; b) atarrayas de 5.0 m de diámetro y una abertura de malla de 4.0 cm. Los ejemplares obtenidos fueron identificados a nivel específico utilizando las claves propuestas por Jordán y Evermann (1896-1900), Berdegué (1954), Greenwood et al. (1966) y Castro (1978). Las mediciones de cada individuo se efectuaron en fresco con un ictiómetro convencional de 60 cm de largo. Se hicieron registros de la longitud total (LT) y la longitud standard o patrón (L. Std o LP). El peso en gramos se obtuvo con un dinamómetro; para la determinación del sexo se uso la escala de madurez de Nikolsky (1963). Los estómagos fueron conservados en formol al 5%, neutralizado con borato de sodio, para su análisis posterior. En la identificación del contenido estomacal se utilizaron los trabajos de Pérez (1970), Brusca (1973), Menz (1976) y Minello et al. (1984). La identificación del contenido estomacal fue hecha a nivel de especie, de género o de familia según las dificultades taxonómicas o el estado de digestión del organismo; la materia orgánica animal o vegetal no identificada fue considerada como detritus. El contenido estomacal fue analizado por el método de Frecuencia de Ocurrencia, en el cual cada estómago con alimento fue considerado como el 100% y los diferentes grupos reconocidos en el mismo estómago fueron visualmente estimados como una porción del volumen total del contenido estomacal del pez, esto se expresa como: F = n/NE x 100 Donde F = frecuencia, n = número de estómagos que contienen dicho alimento y NE = número total de estómagos analizados (Starck y Schroeden, 1970). Para el tratamiento estadístico de los datos se aplicaron las pruebas no- paramétricas de ξ, Tablas de Contingencia y de Kolmorov-Smirnov (Zar, 1974). RESULTADOS
Ariopsis caerulescens (Günther, 1864)
Espectro Alimenticio /Régimen de PrecipitaciónSe reconocieron 27 grupos alimenticios en la dieta de esta especie, cuya composición resulta influenciada por el régimen de sequía y de precipitación; dichos componentes alcanzan una mayor diversidad durante la época de precipitación. Esta información es presentada gráficamente en histogramas de frecuencia por mes de muestreo, así como en forma tabulada (Fig. 3A, Tabla l). Los grupos alimenticios más importantes durante la primera época de sequía (junio de 1977), fueron en orden de ocurrencia Callinectes spp, el díptero Chironomus sp, y las escamas de peces (Fig. 3A y Tabla 1); este último no se incluyó dentro del componente Restos de Peces, debido a su alta frecuencia de ocurrencia en A. caerulescens. Las escamas observadas en el contenido estomacal de esta especie se caracterizaron por su estructura y tamaño bien definido, lo cual elimina la posibilidad de que las escamas identificadas perteneciesen a peces ingeridos varias horas antes. En la segunda época de sequía (enero a junio de 1978), los grupos alimenticios en orden de importancia fueron: Restos de Peces, Escamas de Peces y Detritus (Fig. 3A y Tabla 1). Este último componente es importante en la dieta de los peces que habitan el área de estudio. De acuerdo a Blake et al. (1981), las fuentes principales de detritus dentro del sistema lagunar estudiado son: a) el aporte de los ríos durante la estación de precipitación, b) la descomposición de diatomeas, algas clorofitas Enteromorpha sp y Cladophora sp, macrofitas como Ruppia sp, y las hojas de los manglares. Durante esta época de los 88 individuos de A. caerulescens capturados, solo en 41 se reconoció material alimenticio (Fig. 3B). En la temporada de precipitación correspondiente a noviembre y diciembre de 1977, los grupos alimenticios reconocidos como predominantes fueron: Restos de Crustaceos, Callinectes spp, y Penaeus spp (Fig. 3A y Tabla 1). El primer componente alimenticio incluye los fragmentos pertenecientes a jaibas, camarones peneidos, langostinos, cangrejos, etc., que por su estado de digestión no fue posible identificar específicamente. No obstante la reducida abundancia de Callinectes spp en la época de precipitación, éste representó un importante componente en la dicta de A. caerulescens. Los elementos alimenticios sobresalientes registrados en la segunda época de precipitación (julio a noviembre de 1978), fueron los siguientes: Restos de Crustacea, Restos de Peces y Penaeus spp (Fig 3A y Tabla 1). Los peces indudablemente forman una parte importante de la dieta de A. caerulescens, como los demuestran la alta incidencia de grupos tales como: Escamas de Peces, Gerridae, Centropomidae, Aridae, A. mazatlanus, Gobionellus sp, Lutianidae, D. latifrons, Lile stolifera, Hemirhamphidae y Engraulidae. Con respecto a Penaeus spp, la ocurrencia de este grupo en el contenido estomacal de A. caerulescens se debe a la abundancia de los camarones juveniles en ésta época. De los 314 individuos registrados en esta temporada, solo en 207 se reconoció contenido estomacal (Fig. 3B). 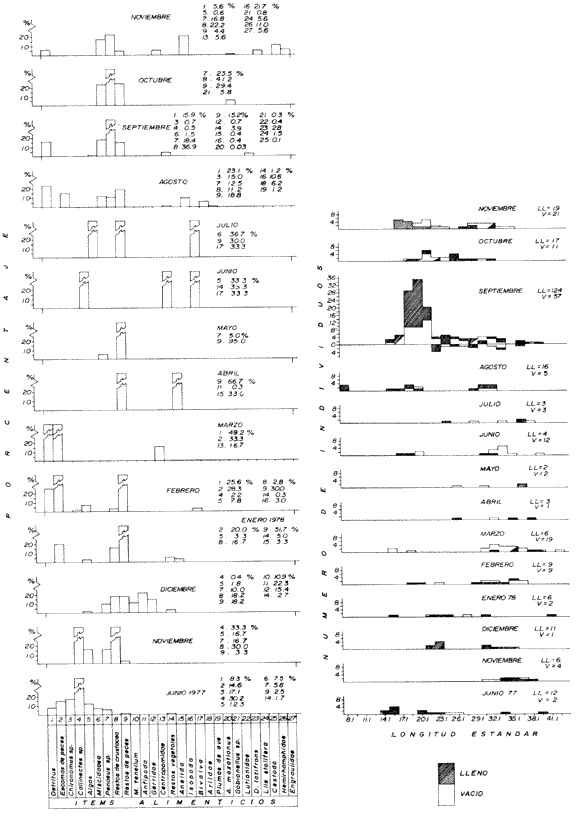 Figura 3. A. Espectro alimenticio de A. cacrulescens durante junio, 1977 -1978. En el margen derecho del histograma se indica el porcentaje de frecuencia de los grupos alimenticios predominantes. B. Relación entre la composición de tallas y la condición de llenado del estómago. Excepto septiembre, 1978 en que los individuos se capturaron con la red 2, el resto de la población se obtuvo con la red 1. 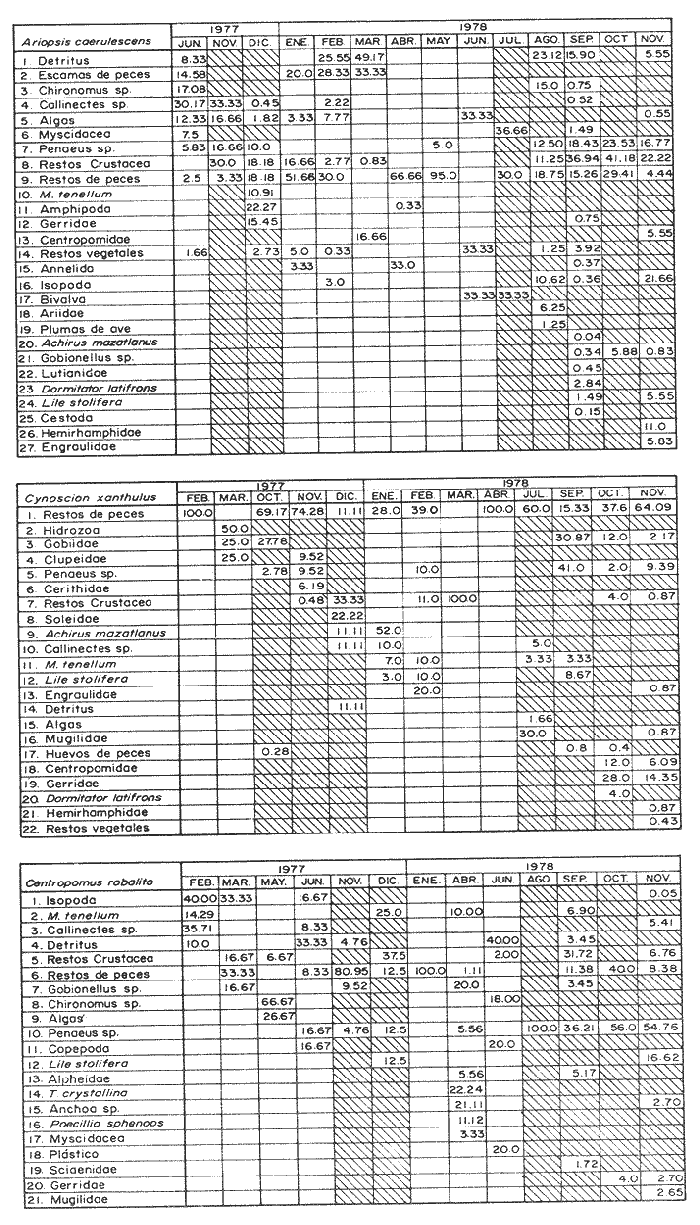 TABLA 1 ESPECTRO ALIMENTICIO DE LAS 6 ESPECIES DEPREDADORAS DURANTE LOS MESES DE ESTÍO Y PRECIPITACIÓN 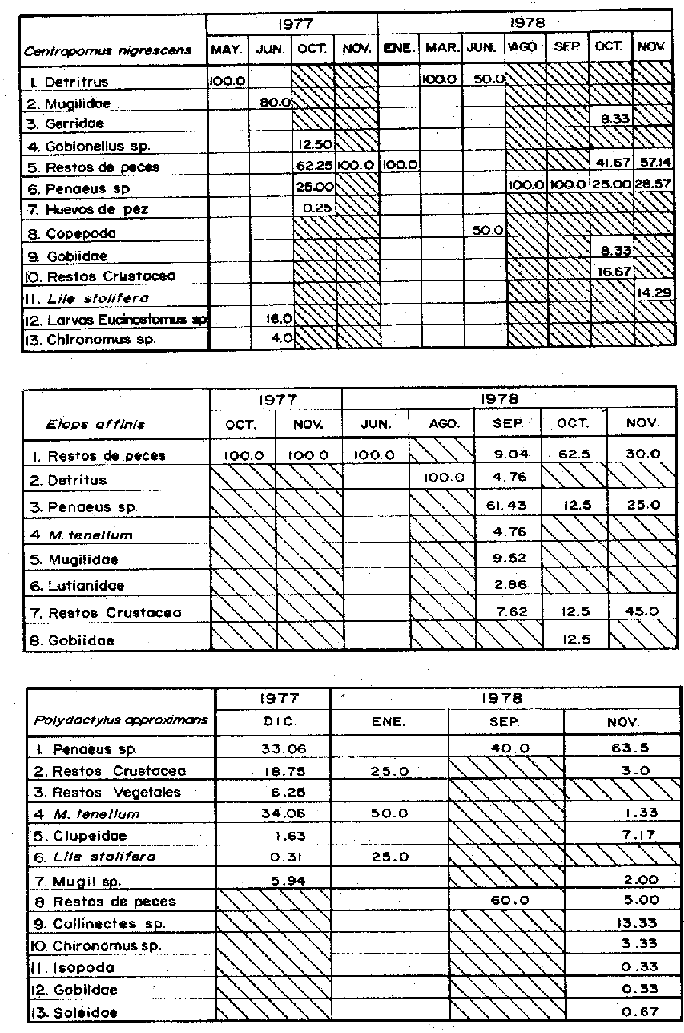 Dieta / Ciclo de VidaSe obtuvieron 84 individuos juveniles (8.1 cm - 18.6 cm. L. Std.) de los cuales, 64 individuos presentaron estómago con alimento y 20 sin contenido estomacal. En cambio, de los 318 individuos adultos (20.1 cm - 42.6 cm L. Std.), en 184 se reconocieron integrantes alimenticios y 134 individuos sin alimento (Fig. 3B). En el caso de los juveniles se observó una marcada tendencia a contener un volúmen estomacal considerable; de cada 10 individuos, 7 se obtuvieron con estómago lleno. En los individuos adultos la relación entre el llenado y vaciado del estómago no resultó tan evidente como en el caso anterior; solo de cada 10 especímenes, aproximadamente 6 presentaron contenido en sus estómagos. Sin embargo, la relación entre la talla de A. caerulescens y la condición de llenado del estómago, demostró que el llenado del estómago no depende de la talla de los individuos (Prueba de Contingencia 2x2, 0.001 < p < 0.005). 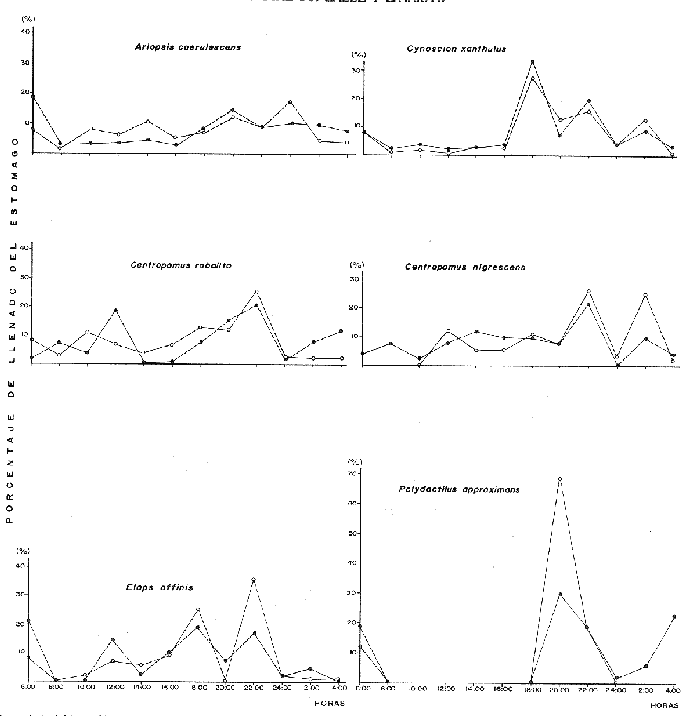 Figura 4. Actividad trófica de las 6 especies depredadoras estudiadas con relación a la hora del día o = estómagos vacíos, o = estómagos llenos. Como se indicó anteriormente, la dieta de A. caerulescens incluyó 27 grupos alimenticios, los cuales aparecen en forma azarosa a través del año. Esto se logró establecer mediante la aplicación de la prueba de Kolmorov-Smirnov, la cual rechazó la hipótesis nula en la que se asumió la distribución uniforme de la composición alimenticia (D = 0.48609* * observada, D0.05,315 = 0.04544 calculada). Si bien no se distingue una uniformidad en la ocurrencia de los grupos alimenticios, si se logra reconocer por su predominancia 5 componentes básicos en la dieta de A. Caerulenscens (Tabla 1): Restos de Crustáceo, Restos de Peces, Penaeus spp, Detritus y Chironomus spp; mediante una prueba de ξ² se logró demostrar el orden en que ocurren estos elementos en la dicta. Los resultados que se desprenden de esta prueba, indican que los componentes anteriores guardan entre sí las siguientes proporciones: 3.6; 2.1; 1.9; 1.6 y 0.7. Sin embargo, la probabilidad de que estas proporciones se conserven, no resultó significativa (0.999 < P) La información analizada en este estudio indica que la actividad alimenticia de A. caerulescens, tiene lugar indiscriminadamente durante el día y la noche. Durante el día el número de individuos con estómagos conteniendo alimento fue de 115, en tanto que en la noche este número fue ligeramente mayor, 133. Sin embargo, la proporción entre estómagos sin contenido estomacal durante el día y la noche, mantuvo una proporción casi de 1:1. El análisis de esta información demostró que la actividad alimenticia de la especie anterior tiene lugar esencialmente en la noche (0.25 < p < 0-50). A esta prueba se le aplicó la corrección de Yates y se determinó la misma probabilidad. En apoyo a este planteamiento, al analizarse los datos porcentuales de estómagos vacíos y llenos en relación con la hora del día, se reconoció un incremento de la actividad alimenticia de A. caerulescens, desde las 18:00 hasta las 06:00 horas (Fig. 4). Cynosción xanthulus Jordan y Gilbert, 1882.
Espectro alimenticio/Régimen de PrecipitaciónSe reconocieron 22 grupos que sirven de alimento a esta especie; esta composición resulta influenciada por el regímen de sequía y precipitación. Los grupos alimenticios muestran una mayor diversidad durante la época de precipitación (Fig. 5A y Tabla 1). En la primera época de sequía (febrero a marzo de 1977), los grupos alimenticios reconocidos como predominantes fueron: Restos de Peces, Hidrozoa y Gobiidae-Clupeidae (Fig. 5A y Tabla 1). El primer componente destacó como uno de los elementos característicos durante el presente estudio. El segundo grupo alimenticio, aunque con un porcentaje considerable 50%, solo se presentó en el mes de marzo, por lo que su presencia se consideró accidental. Los grupos más notables en la segunda época de sequía (enero-abril de 1978), según su orden de importancia fueron: Restos de Peces, Restos de Crustacea y Achirus mazatlanus (Fig. 5A y Tabla 1). Durante éste periodo, de los 56 individuos de C. xanthulus capturados, solo en 28 se reconoció alimento en los estómagos (Fig. 5B). En la primera época de precipitación (octubre a diciembre de 1977), los elementos alimenticios determinados como sobresalientes fueron: Restos de Peces, Restos de Crustacea y Gobiidae (Fig. 5A y Tabla 1). Los elementos alimenticios registrados en la segunda época de precipitación Julio a noviembre de 1978) como predominantes fueron: Restos de Peces, Penaeus spp, y Gobiidae (Fig. 5A y Tabla 1). En esta temporada, de los 381 individuos de C. xanthulus capturados, solo en 206 se reconoció contenido estomacal (Fig. 5A). Dieta / Ciclo BiológicoSe obtuvieron 312 individuos juveniles 8.1 - 29.1 cm L. Std.) de los cuales solo 175 contenían alimento en sus estómagos. Sin embargo, de los 125 individuos adultos (29.1 - 45.6 cm L. Std.), en 59 se reconocieron integrantes alimenticios y 66 sin alimento. (Fig. 5B). En el caso de los juveniles se observó cierta tendencia a encontrar los estómagos con un volumen considerable de alimento; de cada 10 individuos, 6 se obtuvieron con estómago lleno. En los individuos adultos la relación entre el llenado y vaciado del estómago, no resultó tan evidente como en el caso anterior; sólo de cada 10 especímenes, aproximadamente 5 presentaron contenido en sus estómagos. Sin embargo, la relación entre la talla de C. xanthulus y la condición de llenado del estómago, demostró que ésta condición no depende de la talla de los individuos (Prueba de Contingencia 2x2, 0.10 < P < 0.25). Como se señalo anteriormente, la dieta de C. xanthulus incluyó 22 grupos alimenticios, los cuales aparecen en forma azarosa a través del año. Esto se logró establecer mediante la aplicación de la prueba de Kolmorov-Smirnov, la cual rechazó la hipótesis nula, en la que se asumió la distribución uniforme de la composición alimenticia (D = 0.4584* * observada, D0.05,256 = 0.00254 calculada). A pesar de no reconocerse una uniformidad en la ocurrencia de los grupos alimenticios, se logró distinguir por su predominancia 5 componentes básicos en la dieta de C. Xanthulus (Tabla 1): Restos de Peces, Penaeus spp, Gerridae, Gobiidae y Restos de Crustácea; mediante una prueba de ξ² se demostró el orden en que ocurren estos elementos en la dieta. Los resultados que desprenden de esta prueba indican que los componentes anteriores guardan entre sí las siguientes proporciones: 6.3; 1.3; 1.0; 0.8 y 0.4. Sin embargo, la probabilidad de que estas proporciones se conserven, no resultó significativa 0.75 < p < 0.90), 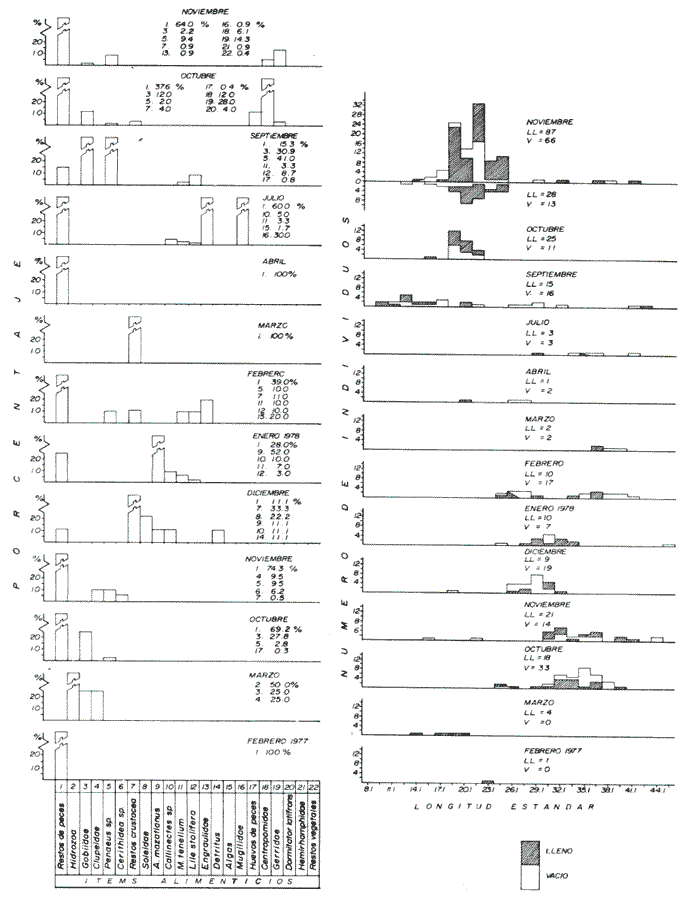 Figura 5. A.Espectro alimenticio de C. xanthulus durante febrero, 1977-noviembre, 1978. En el margen derecho del histograma se indica el porcentaje de frecuencia de los grupos alimenticios predominantes. B. Relación entre la composición de tallas y la condición de llenado del estómago. Excepto noviembre, 1978 en que los individuos se capturaron con la red 2, el resto de la población se obtuvo con la red 1. La información obtenida en este estudio, muestra que la actividad alimenticia de C. xanthulus, tiene lugar indiscriminadamente durante el día y la noche. Durante el día el número de individuos con estómagos llenos fue de 131, en tanto que en la noche este número fue ligeramente menor (103). Sin embargo, la proporción entre estómagos sin contenido estomacal durante el día y la noche, mantuvo una proporción casi de 1:1. El análisis de esta información demostró que la actividad alimenticia de la especie anterior, se intensifica durante la noche (0.25 < P < 0.50); mediante la corrección de Yates se determinó la misma probabilidad. Al analizarse los datos porcentuales de estómagos vacíos y llenos en relación con la hora del día, se evidenció un incremento de la actividad alimenticia de C. xanthulus, desde las 16:00 hasta las 04:00 horas (Fig. 4). Centropomus robalito Jordán y Gilbert, 1882
Espectro Alimenticio/Regimen de PrecipitaciónDurante el periodo de estudio se identificaron 21 grupos alimenticios en la dieta de esta especie, cuya composición resulta también afectada por el régimen de sequía y de precipitación; los componentes citados son más diversos durante la época de precipitación (Fig. 6A y Tabla 1). En la primera época de sequía (febrero a junio de 1977) los grupos esenciales en orden de importancia fueron: Chironomus sp, y Callinectes spp En la segunda época de sequía (enero a junio de 1978) se reconoció como alimentos alimenticios de importancia a los Restos de Peces, Detritus y Thyrinops crystallina (Fig. 7A y Tabla 1). Durante este tiempo de los 91 individuos capturados, solo en 43 de ellos se reconoció material alimenticio. (Fig. 6B). En la primera época de precipitación (noviembre y diciembre de 1977) los elementos alimenticios reconocidos como sobresalientes fueron: Restos de Peces,Restos de Crustácea y Macrobrachium tenellum. En la segunda época de precipitación (agosto a noviembre de 1978) los elementos alimenticios primordiales fueron: Penaeus spp, Restos de Peces y Restos de Crustacea (Fig. 6B). Dieta/Ciclo de VidaSe obtuvieron 91 individuos juveniles (6.6 - 14.1 cm L. Std.), de los cuales 45 individuos presentaron estómagos con alimento y 46 sin contenido estomacal En cambio de los 323 individuos adultos (15.6 - 47.1 cm L. Std.), en 111 se reconocieron integrantes alimenticios y 212 individuos sin alimento (Fig. 6B). En los juveniles se observó una tendencia marcada a encontrar los estómagos sin un volumen significativo de alimento; de cada 10 individuos, 5 se obtuvieron con estómago lleno. En los individuos adultos la relación entre el llenado y vaciado del estómago resultó más evidente que en el caso anterior; ya que de cada 10 ejemplares 3 presentaron contenido en sus estómagos. No obstante, al analizar la relación entre la talla de C. robalito y la condición de llenado, se demostró que tal condición no depende del tamaño del pez (Prueba de Contingencia 2x2, 0.005 < P < 0.001). Como se expresó anteriormente C. robalito incluye 21 grupos alimenticios, los cuales aparecen en forma azarosa a través del año, esto se logró establecer mediante la prueba de Kolmorov-Smirnov, la cual rechazó la hipótesis nula en la que se asumió la distribución uniforme de la composición alimenticia (D = 0.36971* observada, D0.05,175 = 0.06097 calculada. Si bien no se percibe una uniformidad en la ocurrencia de los grupos alimenticios, se logró discernir por su predominancia 6 componentes fundamentales en la dieta de C. robalito (Tabla 1): Penaeus spp, Restos de Peces, Restos de Crustacea, Isopoda, Callinectes spp, y Macrobrachium tenellum; a través de la aplicación de una prueba de ξ² se demostró la proporción en que ocurren estos elementos en la dieta. Los resultados que se desprenden de esta prueba indican que los componentes anteriores guardan entre si las siguientes proporciones: 3.9; 2.7; 1.5; 0.8; 0.7 y 0.4. La probabilidad de que estas proporciones se conserven no resultó significativa (0.25 < P < 0-50). La información analizada indicó que la actividad alimenticia de C. robalito, tiene lugar indistintamente durante el día y la noche. Durante el día el número de individuos con estómagos conteniendo alimento fue de 67, en tanto que en la noche este número fue ligeramente mayor, 89. Sin embargo, la proporción entre estómagos sin contenido estomacal durante el día y la noche, mantuvo una proporción casi de 1:1. El análisis de esta información demostró que la actividad de la especie anterior, se realiza esencialmente en la noche (0.025 < p < 0.05). A esta prueba se le aplicó la corrección de Yates y se determinó la misma probabilidad. En auxilio a este planteamiento, al analizarse los datos porcentuales de estómagos vacíos y llenos en relación con la hora del día, se reconoció un incremento de la actividad alimenticia de C. robalito, desde las 16:00 hasta las 24:00 horas (Fig. 4). 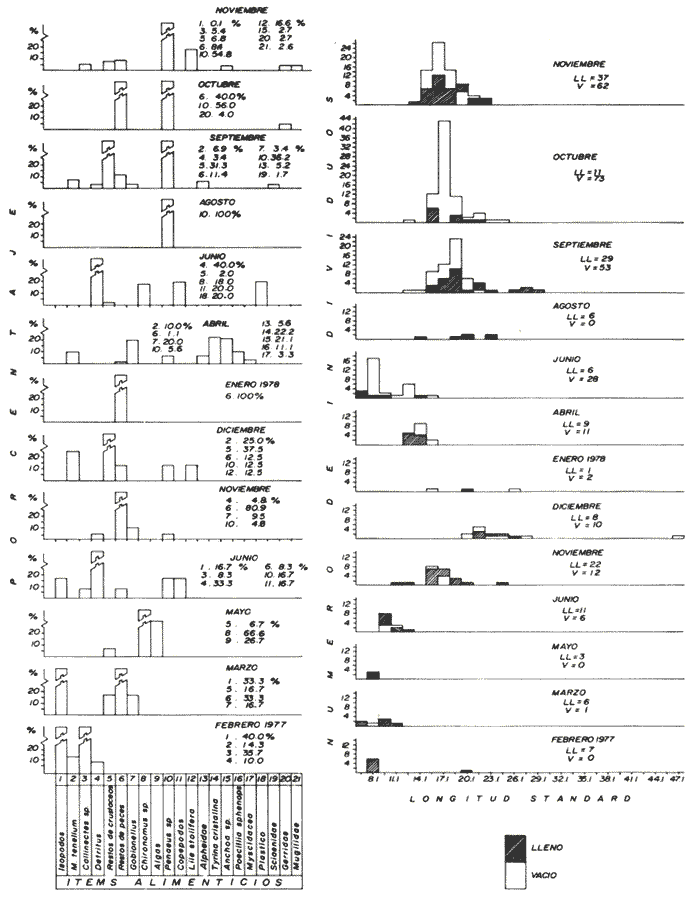 Figura 6. A. Espectro alimenticio de C. robalito durante febrero, 1977-noviembre, 1978. B Relación entre la composición de tallos y la condición de llenado del estómago Centropomus nigrescens Günther, 1864.
Espectro Alimenticio/Régimen de PrecipitaciónDurante el periodo de estudio se reconocieron 13 grupos alimenticios en la dieta de esta especie, cuya composición resultó influenciada por el régimen de sequía y precipitación. Al igual que en las especies anteriores dichos componentes alcanzan también una mayor diversidad durante la época de precipitación (Fig. 7A y Tabla 1). Los conjuntos alimenticios predominantes durante la primera época de sequía (mayo y junio de 1977) fueron los siguientes: Detritus, Mugilidae y larvas de Eucinostomus sp. En la segunda época de sequía (enero a junio de 1978) los elementos alimenticios reconocidos como sobresalientes fueron el Detritus, Restos de Peces y Copepoda (Fig. 7A y Tabla 1). Durante esta temporada de los 15 individuos capturados solo en 10 de ellos se reconoció material alimenticio (Fig. 7B). En la primera época de precipitación (octubre y noviembre de 1977), los grupos alimenticios en orden de importancia fueron los Restos de Peces, Penaeus spp, y Gobionellus sp. Durante la segunda época de precipitación (agosto a noviembre de 1978) los elementos alimenticios significativos fueron Panaeus spp, Restos de Peces y Restos de Crustacea (Fig. 7A y Tabla l). De los 142 individuos registrados en esta temporada, solo en 30 de ellos se reconoció contenido estomacal (Fig. 7B). Dieta/Ciclo biológicoSe obtuvieron 11 individuos juveniles (8.1 - 15.6 cm L. Std.), de los cuales 6 individuos presentaron estómagos con alimento y 5 sin contenido estomacal. En cambio de los 159 individuos adultos (17.1 - 56.1 cm L. Std.), en 43 se reconocieron integrantes alimenticios y 116 individuos sin alimento (Fig. 7B). En el caso de los juveniles se observó tendencia a registrar estómagos con un volumen considerable de alimento; de cada 10 individuos, 5 se obtuvieron con estómago lleno. En los individuos adultos la relación entre el llenado y vaciado del estómago resultó más evidente que en el caso anterior; ya que de cada 10 especímenes aproximadamente 3 presentaron contenido en sus estómagos. Sin embargo el análisis de la relación entre la talla de C. nigrescens y la condición de llenado del estómago, demostró que el llenado del estómago depende de la talla de los individuos (Prueba de Contingencia de 2x2, 0.05 < P < 0.10). Como ya se indicó, la dicta de C. nigrescens incluyó 13 grupos alimenticios, los cuales aparecen en forma azarosa a través del año. Esto se logró establecer mediante la aplicación de la prueba de Kolmorov-Smirnov, a través de la cual se rechazó la hipótesis nula en la que se asumió la distribución uniforme de la composición alimenticia (D = 0.3598** observada, D0.05,56 = 0.17823 calculada). A pesar de diferenciarse una uniformidad en la ocurrencia de los grupos alimenticios, se logró precisar por su predominancia a 6 componentes primordiales en la dieta de C. nigrescens (Tabla 1): Restos de Peces, Penaeus spp, Mugilidae, Detritus, Gobiidae y Lile stolifera; mediante una prueba de ξ² se pudo demostrar la proporción en que ocurren estos elementos en la dieta. Los resultados que se desprenden de esta prueba indicaron que los componentes anteriores guardan entre sí las siguientes proporciones: 4.1; 3.7; 0.9; 0.6; 0.4 y 0.2. Sin embargo la probabilidad de que estas proporciones se conserven, no resultó significativa (0.75 < p < 0.90). La información analizada en este estudio indicó que la actividad alimenticia de C. nigrescens, tiene lugar indiscriminadamente durante el día y la noche. Durante el día el número de individuos con estómagos conteniendo alimento fue de 27, en tanto que en la noche este número fue ligeramente menor (22). Sin embargo, la proporción entre estómagos sin contenido estomacal durante el día y la noche, mantuvo una proporción casi de 1:2. El exámen de esta información demostró que la actividad trófica de la especie anterior, se realiza esencialmente en el día (0.025 < p < 0.05). A esta prueba se aplicó la corrección de Yates y se determinó la misma probabilidad. Apoyando este planteamiento, el análisis de los datos porcentuales de estómagos vacíos y llenos, en relación con la hora del día, permitió reconocer un incremento de la actividad alimenticia de C. nigrescens, de las 10:00 atenuándose hacia las 24:00 horas (Fig. 4). 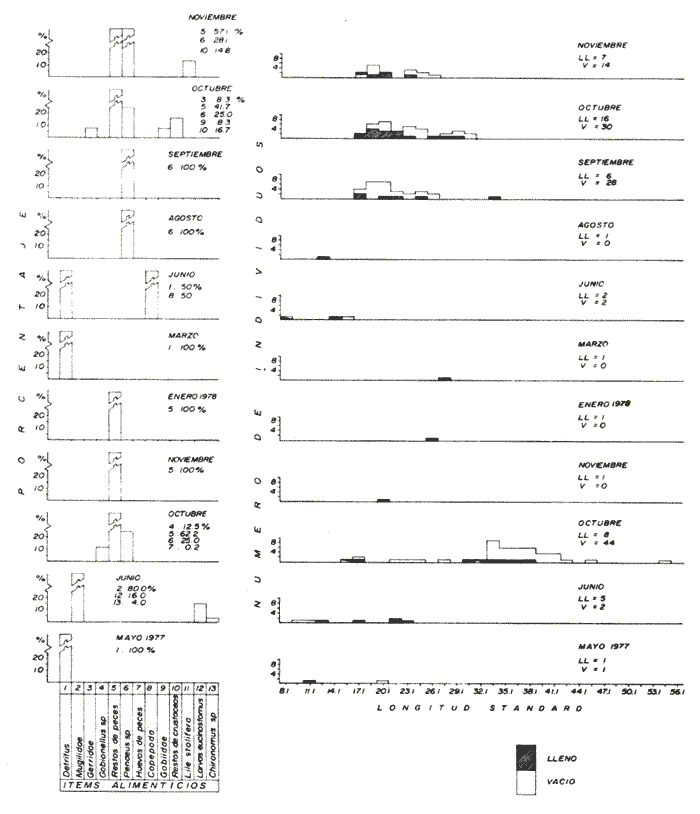 Figura 7. A. Espectro alimenticio de C. nigrescens durante mayo, 1977-noviembre, 1978. B. Relación entre la composición detalladas y la condición de llenado del estómago. Elops affinis Regan, 1909
Espectro alimenticio/Régimen de PrecipitaciónSe reconocieron 8 grupos alimenticios en la dieta de esta especie, su composición resultó influenciada por el régimen de sequía y de precipitación (Fig. 8A, Tabla 1). El grupo alimenticio reconocido como predominante durante la época de sequía Junio de 1978) fue el Detritus (Fig. 8A y Tabla 1). Durante este periodo solo se capturó un ejemplar en el cual se reconoció material alimenticio (Fig. 8B). En la primera época de precipitación (octubre y noviembre de 1977) el grupo alimenticio reconocido como sobresaliente fue el de Restos de Peces. En la segunda época de precipitación (agosto a noviembre de 1978) los elementos alimenticios predominantes registrados fueron el Detritus, Restos de Peces y Penaeus spp (Fig. 8A y Tabla l). De los 125 individuos registrados en esta temporada, sólo en 40 se reconoció contenido estomacal (Fig. 8B). Dieta/Ciclo BiológicoSe obtuvieron 72 individuos juveniles (15.6 - 26.1 cm L. Std.), de los cuales 23 individuos presentaron estómago con alimento y 49 sin contenido estomacal. En contraste, de los 54 individuos adultos (27.6 - 35.1 cm L.Std.), en 18 se reconocieron integrantes alimenticios y 36 individuos sin alimento (Fig. 8B). En el caso de los juveniles se observó marcada tendencia a encontrar estómagos sin volumen significativo de alimento; de cada 10 individuos, 3 se obtuvieron con estómago lleno. En los individuos adultos la relación entre el llenado y vaciado del estómago resultó tan evidente como en el caso anterior, ya que de cada 10 especímenes 3 presentan contenido en sus estómagos. Sin embargo, al aplicar una prueba de Contingencia de 2x2 para establecer la relación entre la talla de E. affinis y la condición de llenado del estómago, se demostró que el llenado del estómago depende de la talla de los individuos. La probabilidad de que esta relación establecida se conserve, no resultó significativa (0.975 < P < 0.99). La dieta de E. affinis incluyó 8 grupos alimenticios, los cuales aparecen en forma azarosa a través del año. Esto se logró establecer mediante la aplicación de la prueba de Kolmorov-Smirnov, a través de la cual se rechazó la hipótesis nula en la que se asumió la distribución uniforme de la composición alimenticia (D = 0.3869** observada, D0.05,42 = 0.20517 calculada). Si bien no se reconoció uniformidad en la ocurrencia de los grupos alimenticios, se logró diferenciar por su predominancia 5 componentes fundamentales en la dieta de E. affinis (Tabla 1): Penaeus ssp, Restos de Peces, Restos de Crustacea, Detritus y Macrobrachium tenellum; mediante una prueba de ξ² se demostró el orden en que ocurren estos elementos en la dieta. Los resultados que se desprenden de esta prueba indicaron que los componentes anteriores guardan entre sí las siguientes proporciones: 4.2; 3.7; 1.3; 0.5; y 0.2. Sin embargo la probabilidad de que estas proporciones se conserven, no resultó significativa (0.75 < P < 0.90). La información analizada indicó que la actividad alimenticia de E. affinis, tiene lugar indistintamente durante el día y la noche. Durante el día el número de individuos con estómago conteniendo alimento fue de 28, en tanto que en la noche este número fue menor (13). Sin embargo, la proporción entre estómagos sin contenido estomacal durante el día y la noche, mantuvo una proporción casi de 2:1. El análisis de esta información demostró que la actividad alimenticia de la especie anterior, se realiza esencialmente en el día (0.25 < p < 0.50). A esta prueba se le aplicó la corrección de Yates y se determinó la misma probabilidad. Al analizarse los datos porcentuales de estómagos vacíos y llenos en relación con la hora del día, se reconoció un incremento de la actividad alimenticia de E. affinis, desde las 10:00 hasta las 18:00 horas (Fig.4). Polydactylus approximans (Lay y Bennet, 1849)
Espectro alimenticio/Régimen de PrecipitaciónSe determinaron 13 grupos alimenticios en la dieta de esta especie, cuya composición resulto influenciada por el régimen de precipitación; dichos componentes fueron más diversos durante la temporada de lluvias (Fig. 9A y Tabla 1). En la época de sequía (enero de 1978) los grupos predominantes fueron: Macrobrachium tenellum, Restos de Crustacea y Lile stolifera (Fig. 13A y Tabla 1). Durante esta temporada se capturaron 4 especímenes en los cuales se reconoció material alimenticio (Fig. 9B). Durante la primera época de precipitación (diciembre de 1977) los grupos alimenticios primordiales fueron: Macrobrachium tenellum, Penaeus spp y Restos de Crustacea. En la segunda época de precipitación (septiembre y noviembre de 1978) los componentes alimenticios más significativos en orden de importancia fueron: Penaeus spp, Restos de Peces y Callinectes spp, (Fig. 9A y Tabla 1). De los 64 individuos registrados en esta temporada, solo en 48 se reconoció contenido estomacal (Fig. 9B). En los individuos adultos se observó tendencia a encontrar los estómagos con un volumen considerable de alimento; de cada 10 individuos, 8 se obtuvieron con estómago lleno. La ausencia de individuos de tallas menores evitó el establecer la relación entre el tamaño del pez y la condición de llenado estomacal. 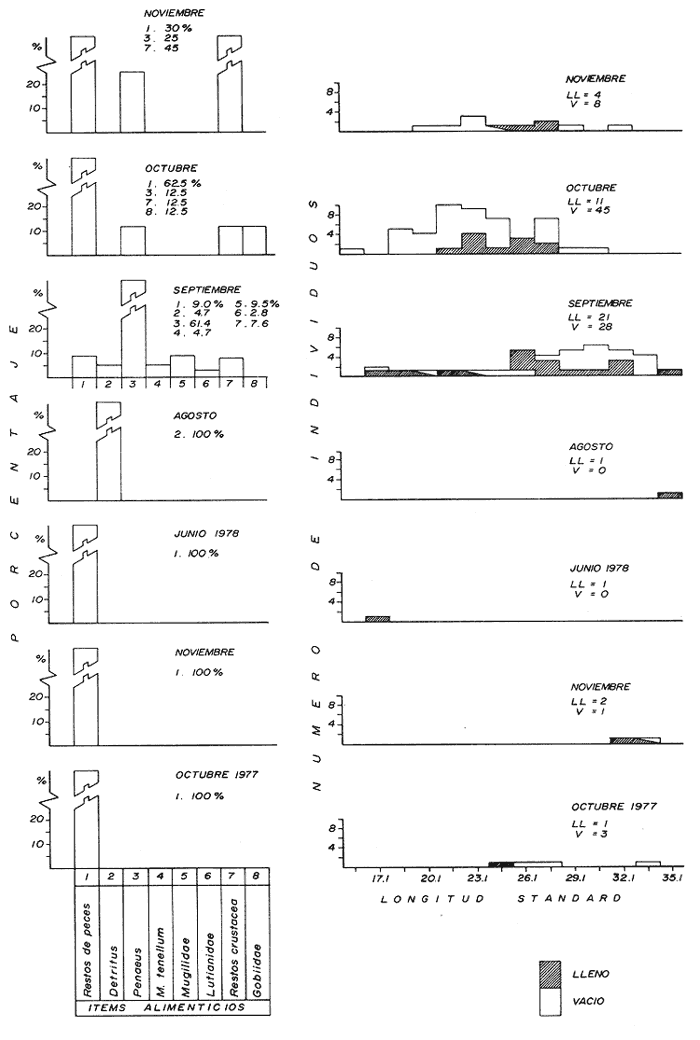 Figura 8. A. Espectro alimenticio de E.affinis durante octubre, 1977-noviembre, 1978. En el margen derecho del histograma se indica el porcentaje de frecuencia de los grupos alimenticios predominantes. B. Relación entre la composición de tallas y la condiciòn de llenado del estómago. 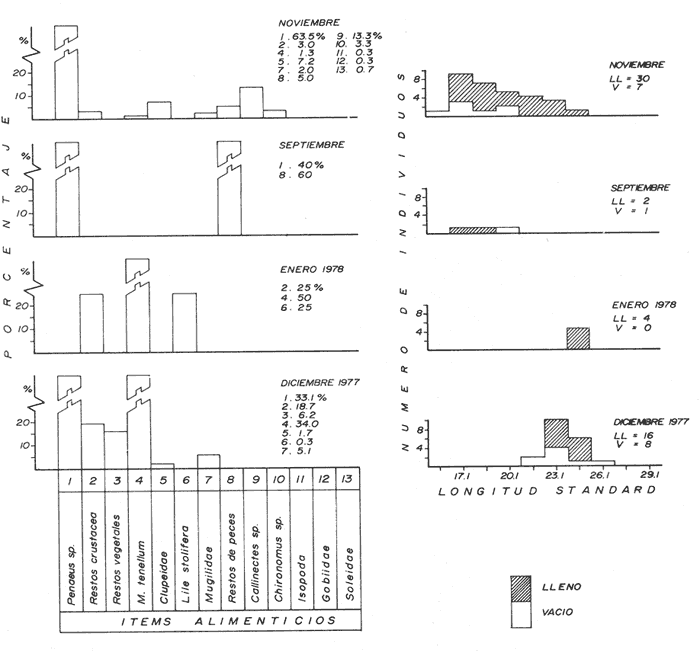 Figura 9. A. Espectro alimenticio de P. approximans durante diciembre, 1977-noviembre, 1978. B. Relación entre la composición de tallas y la condición de llenado del estómago. La dieta de P. approximans incluyó 13 elementos alimenticios, los cuales aparecen en forma azarosa a través del año. Esto se logró establecer mediante la aplicación de la prueba de Kolmorov-Smirnov, la cual rechazó la hipótesis nula en la que se asumió la distribución uniforme de la composición alimenticia (D=0.39003** observada, D0.05,71=0.15864 calculada). A pesar de no observarse una uniformidad en la ocurrencia de los grupos alimenticios, se reconoció por su predominancia a 6 componentes básicos en la dieta de P. approximans (Tabla 1): Penaeus spp, Clupeidae, Macrobrachitun tenelluni, Restos de Crustacea, Restos de Peces y Restos Vegetales; mediante una prueba de ξ² se determinó el orden en que ocurren estos elementos en la dieta. Los resultados de esta prueba indican que los componentes anteriores guardan entre sí las siguientes proporciones: 5.1; 1,7; 13; 0.8; 0.7 y 0.2. La probabilidad de que estas proporciones se conserven, no resultó significativa (0.75<P<0.90). La información referente a la actividad alimenticia de P. approximans, no se demostró estadísticamente debido al reducido número de ejemplares; sin embargo, al parecer esta especie se alimenta esencialmente en la noche. El análisis de los datos porcentuales de estómagos vacíos y llenos en relación con la hora del día, reveló un incremento en la actividad alimenticia de P. approximans, desde las 18:00 hasta las 24:00 horas (Fig. 4). DISCUSIÓNVarios autores, entre ellos Hynes (1950) y Man y Orr (1969), han establecido que cuando se discuten métodos de análisis de contenido estomacal de peces que involucran grupos alimenticios en la dieta, ésta deberá ser independiente del método empleado; cada método mostrará la medición de los grupos alimenticios importantes en la dieta de los peces de acuerdo a los atributos para que fue diseñado. Sin embargo, cuando son tomadas muestras pequeñas y la variación en categorías alimenticias es grande, métodos diferentes pueden producir resultados completamente distintos (Radforth, 1940). Por otra parte, Hyslop (1980) ha constatado que todos los métodos empleados para el análisis de estómagos tienen ventajas y desventajas. En ocasiones, la selección del método depende del tiempo disponible para su realización. En esta investigación, el procedimiento seleccionado de Frecuencia de Ocurrencia, tuvo la ventaja de ser sencillo y de llevarse a cabo en un tiempo razonable. No obstante, proporciona poca información sobre la cantidad relativa o volumen de cada alimento presente en el estómago. El método proporciona un espectro alimenticio cualitativo que se considera aceptable (Crisp, 1963; Fagade y Olaniyan, 1972, Blake, 1977). Johnson (1977) ha usado el Método de Frecuencia de Ocurrencia como un indicador de competencia interespecífica en dos o más depredadores. El método ha sido utilizado también para ilustrar cambios estacionales en la composición de la dieta (Frost, 1977). En el sistema lagunar Huizache-Caimanero, la aplicación del método referido permitió reconocer la incidencia trófica de A. caerulescens, C. xanthulus, C. robalilo, C. nigrescens, E. affinis y P. approximans sobre los habitantes macrobénticos entre los que destacan los camarones penéidos. El análisis del grado de competencia interespecifica entre estos tres componentes de la ictiofauna, deberá de considerar la amplitud del espectro alimenticio de cada especie; entre más diverso sea éste, mayor oportunidad de sobrevivencia tendrá el pez cuya población se adapte mejor a la disponibilidad de alimento en un sistema caracterizado por su contrastante régimen hidrológico a través del año. En el presente trabajo se establecieron nueve localidades de muestreo, con el propósito de establecer comparación con las investigaciones previamente efectuadas en el área de estudió (Carranza, 1969a; l969h; Carranza y Amezcua, 1971a; 1971b; Amezcua, 1977; Warburton, 1978 y 1979). Después de 9 meses, éstas se redujeron a Tapo Botadero y Tapo Agua Dulce, debido al número reducido de individuos obtenidos en las otras localidades. Estos dos últimos sitios representaron las áreas más importantes de pesca comercial de camarones y peces. En un principio se supuso que con la red agallera de abertura de 5 y 9 cm (Red. No.2) se lograría una captura abundante de ejemplares mayores de 40.0 cm de L. Std., sin embargo, solo se obtuvo el 15 % de los especímenes. Esto se justificó por exístir pocos individuos de esta talla, y porque este tipo de red construida de hilo de seda, es cortada fácilmente por las jaibas del género Callinectes spp. En contraste, la red agallera de 2,3 cm construida de hilo nylon (Red. No.1), capturó 80% de la muestra que incluyó peces hasta de 35 cm de L. Std. Con las atarrayas solo se obtuvo el 5% de la captura, pero compuesta por una mayor diversidad de tallas. La importancia de especímenes en poblaciones naturales a través de un ciclo de 24 horas está ampliamente demostrada, ya que proporciona una idea aceptable del tiempo de ocurrencia de los mayores periodos de alimentación (Darnell, 1958). Por otra parte, Darnell y Meierotto (1962) comprobaron en experimentos con el bagre Ictalurus melas, que en un tiempo de ingestión de 3 horas, el anfípodo Hyalella azteca aún es reconocible, aunque se nota la perdida de la placa dorsal y los apéndices empiezan a separarse. Después de 6 horas solo quedan algunos remanentes quitinosos en el estómago. No obstante que estos autores desarrollaron este trabajo en condiciones experimentales y con organismos de 4 a 6 m de L. Std., sus observaciones coinciden parcialmente con los resultados obtenidos en esta investación; ya que al retirar cada dos horas los ejemplares capturados, se logró obtener en la mayoría de los casos material estomacal identificable. La amplia distribución de los grupos alimenticios que consume A. caerulescens resultaron claramente influenciados por los periodos de sequía y precipitación. A pesar de que algunas especies de bagre tienen un amplio espectro alimenticio, estas presentan grupos dominantes, cuya presencia depende de la disponibilidad y accesibilidad del alimento, estación del año y la edad del pez (Thomas, 1966; Weatherley, 1972). El tratamiento estadístico aquí aplicado corroboró que los grupos alimenticios no se distribuyen uniformemente a través del año. Durante la época de sequía (enero - junio) el grupo alimenticio Callinectes spp, ocupó un lugar destacado. Esto se atribuyó a las altas densidades de juveniles de Callinectes arcuatus y Callinectes toxotes reportadas por (Paul, 1977) en el área de Huizache-Caimanero Salazar (1980) a este respecto señaló, que en los meses de febrero a mayo ocurre el mayor reclutamiento en el área de estudio. De el díptero Chironomus sp, no se tienen datos de la época de mayor abundancia, pero Thomas (1966) señala que este insecto es frecuente en áreas someras, ricas en sedimentos y vegetación enraizada. La alta presencia de Escamas de Peces, en esta época se debe a las escamas liberadas por los peces durante la captura comercial que se practica en este periodo del año (Salazar, 1980). La presencia constante de los Restos de Peces en la dicta de A. caerulescens obedece a la ocurrencia a través de todo el año de larvas y juveniles de peces pertenecientes a la familia Gobiidae (Alvarez et al., 1984); los miembros de esta familia han sido ampliamente reconocidos como forrajeros (Yañez y Díaz, 1977). Los peces son los más representativos de todo el espectro alimenticio de esta especie. Corbet (1961) indicó que los bagres pueden ser piscívoros en virtud de la amplitud de su boca y su relativa especialización béntica; este autor indica que en el bagre Clarias mossambicus se alimenta principalmente de ostrácodos e insectos hasta alcanzar los 3.0 cm. de LT, y los individuos mayores de esta talla se alimentan de peces. El Detritus es de importancia primordial dentro del Sistema. Edwards (1978) señaló que Callinectes spp, el camarón Penaeus spp, el langostino M. tenellum, los peces D. latifrons, Gobionellus sp, y Mugil curema incluyen en su dicta principalmente material de origen orgánico. La composición por talla de los individuos de A. caerulescens, incluyeron individuos en etapa de reproducción; es precisamente durante la época de sequía cuando tiene lugar el proceso de reproducción de esta especie en el sistema (Melchor, 1980). Por otra parte el predominio de estómagos vacíos en relación a los llenos debe de estar relacionado con las condiciones hidrológicas de la laguna durante la época de sequía el volumen de las lagunas disminuye considerablemente, con lo cual, se reduce también la cantidad de alimento disponible para los peces. En la época de precipitación (julio-diciembre), el predominio de los Restos de Crustacea, se debe a la diversidad de organismos de este tipo que ocurren en esta temporada; otra posibilidad puede ser que los bagres consuman directamente los restos de crustáceos desechados por otros depredadores. La alta presencia del grupo Callinectes spp, en esta época no es fácil de explicar, pues Paul (1977) reporta en los meses de noviembre a diciembre, predominancia de hembras ovígeras y los escasos juveniles que logró capturar con la red de arrastre, estuvieron mezclados con pastos de Ruppia sp, abundante en esta época. Thomas (1966) refiere que los bagres presentan cierta adaptación béntica como son tu barbas y la forma de "cuchara" de su boca, que en el caso de A. caerulescens podría facilitar la captura de Callinectes spp, aunque éstas estuvieran muy dispersas y su abundancia fuera reducida. Las poblaciones del camarón Penaeus spp, predominan en la época de precipitación. De acuerdo con Soto (1969) y Watkins (1980) el reclutamiento de los camarones peneidos comienza a mediados de julio y termina a mediados de septiembre. No obstante la alta disponibilidad de este alimento, éste no ocupa un lugar sobresaliente en la dieta de A. caerulescens, a excepción del mes de septiembre y parte de noviembre, cuando la producción de camarones juveniles alcanza su máximo. Esto viene a reforzar la opinión de que A. caerulescens no tienen una marcada preferencia por este alimento a pesar de su disponibilidad. No obstante la predominancia de las individuos adultos sobre los juveniles de A. caerulescens, al parecer las tallas, no influyen en la condición de llenado del estómago. La falta de correlación entro individuos con estómago lleno y vacío es probable que se deba a que se colectó mayor número de ejemplar" en la noche que en el día; de acuerdo con la prueba estadística aplicada a esta especie, se demostró que la actividad alimenticia se realiza preferentemente en la noche. El trabajo de Melchor (1980) demostró que el alimento preferencial de A. caerulescens son los peces, y en menor grado los camarones. Edwards (1978) anteriormente señaló que el alimento principal fué Penaeus SPP, los peces y Callinectes spp, ocupan una segunda y tercera categoría respectivamente. La diferencia entre los resultados de Melchor (1980) y los obtenidos en ésta investigación, es explicable ya que el estudio de Melchor (1980) abarcó solo el área de la Laguna de Caimanero. Los resultados de Edwards (1978) por otra parte, difieren con esta investigación debido a que la mayoría de las capturas de A. caerulescens las realizó este autor, en la época de precipitación, sobre todo en los meses en que el camarón es abundante. A. carulescens es residente del sistema Huizache-Caimanero y de acuerdo con la clasificación propuesta por Day et al. (1973), Sheridan et al. (1983), A. carulescens se comporta como una especie netamente carnívora. Según Yañez-Arancibia (1978), constituye un consumidor secundario en las lagunas costeras de Guerrero, cuyo espectro alimenticio es similar al descrito en este trabajo. Con el objeto de reducir la presión de depredación ejercida por la especie anterior sobre la población de peneidos, se sugiere su captura sobre todo en los meses de reproducción (junio y julio) por medio de redes agalleras, en los esteros y canales del Sistema Huizache-Caimanero; es en esta época según Watkins (1980), cuando ocurre la mayor inmigración de postlarvas. Por otra parte es importante la recomendación de Warburton (1978), quien sugiere el inicio de una pesquería comercial de A. caerulescens no obstante que localmente no es aceptable como alimento. La propuesta de Warburton (1978) resulta viable a mediano y largo plazo, estableciendo una empacadora con técnica especializada, donde se procesara en forma atractiva la pulpa le este pez como se hace en Alabama, EUA, con Ictalurus punctatus; esta especie ya se cultiva comercialmente Rosario, Sin., y las ventajas que tiene sobre A. carulescens es únicamente la blancura de su carne. En cuanto a los grupos alimenticios que ingiere C. xanthulus se determinó una variada distribución, los cuales son influenciados por el régimen de sequía y precipitación. De acuerdo con la temporada de sequía (enero-junio) el elemento alimenticio Restos de Peces, ocupó un lugar sobresaliente; la explicación coincide a lo discutido en A. caerulescens ya que C. xanthulus también tiene preferencia por organismos macrobénticos. El grupo alimenticio Hidrozoa, solo se registró en el mes de marzo por lo que se supone que su presencia en la laguna fue accidental. La importancia de los elementos alimenticios Gobiidae y Clupeidae ya ha sido discutida anteriormente; no obstante Warburton (1979) reconoció representantes de ambas familias en ambos periodos de sequía y precipitación. La alta presencia de los Restos de Crustacea, es probable que se deba a la gran densidad de individuos jóvenes de Callinectes spp, reportadas por Sala (1980); aunque por otra parte, Edwards (1978) señala que Macrobrachium sp y otros palemónidos completan su ciclo de vida dentro de la laguna. La constante ocurrencia del pez Achirus mazatlanus en la dieta de C. xanthulus, es consecuencia del reclutamiento contínuo de esta especie a través del año (Warburton, 1978). En la época de sequía se registraron las tallas mayores debido a que los individuos alcanzan su máximo desarrollo dentro de la laguna por causas de tipo trofeo. No se notó desproporción entre estómagos vacíos y llenos no obstante el reducido volumen de la laguna y la menor disponibilidad de alimento. Durante la época de precipitación (julio-diciembre), el predominio de los Restos de Peces es difícil de justificar, pues se asumió que en esta época, este alimento se presentaría en menor cantidad debido a la diversidad de otros grupos alimenticios; sin embargo, al parecer esto tiene relación con la talla de los especímenes de C. xanthulus. Moody (1950) registró para Cynoscion xanthulus en estudios realizados en Florida, EUA, que esta especie a partir de los 27.5 cm de L. Std., se alimenta principalmente de peces; esto coincide con la presente investigación ya que los individuos capturados en su mayoría sobrepasan dicha talla. La ocurrencia del elemento alimenticio Restos de Crustáceo se debe a la gran diversidad de organismos de este tipo que se registran en esta época. No obstante de aparecer en una posición secundaria el elemento alimenticio Penaeus spp, no deja de ser importante debido al creciente interés por conocer el efecto de los peces sobre este recurso (Minello et al., 1984). El integrante alimenticio Gobiidae apareció con una alta frecuencia durante la época de precipitación, lo cual coincide con las altas densidades reportadas por Amezcua (1977) para los integrantes de esta familia; según este autor los aportes fluviales favorecen la abundancia de este tipo de peces dado su origen dulciacuícola. No obstante la predominancia en C. xanthulus de individuos juveniles sobre los adultos, al parecer la talla no influyó en la condición de llenado del estómago. La desproporción entre especímenes con estómago lleno y vacío se debe a la mayor captura de organismos en la noche que en el día; la prueba estadística aplicada a esta especie, demostró que su actividad alimenticia se realiza esencialmente en la noche. De acuerdo con lo anterior, los grupos característicos en la dicta de C. xanthulus fueron: Restos de Peces, Penaeus spp, Gerridae, Gobiidae y Restos de Crustacea. Esto coincidió parcialmente con los trabajos realizados por Carranza (1969), Warburton (1979) y Yañez-Arancibia (1978), quienes señalan que esta especie es ampliamente carnívora. C. xanthulus es una especie residente estacional del Sistema Huizache-Caimanero y de acuerdo con la clasificación de Day et al. (1973), se puede considerar como un organismo netamente carnívoro. C. xanthulus por ser abundante durante la época de precipitación, ejerce un efecto depredador considerable sobre las poblaciones de Penaeus spp. Se sugiere establecer un control a principios del mes de agosto, cuando las tallas de C. xanthulus alcanzan los 23.0 cm de L. Std., usando para ello redes agalleras. Como la carne de la especie citada tiene aceptación en el mercado, su comercialización sería factible. Durante la época de sequía (enero-julio) dentro del espectro alimenticio de C. robalito el integrante alimenticio Isopoda, ocupó un lugar predominante; para explicar su presencia es necesario referirnos al sistema a Huizache-Caimanero como un ambiente sujeto a la desecación y cambios constantes de salinidad, soportando una comunidad de organismos que al cambiar las condiciones algunos emigran, otras se adaptan o tienen una generación efímera. Entre los que mejor se adaptan, figura un isópodo del genéro Sphaerom, que habita en aguas estuarinas, enterrado en el lodo en condiciones mínima de oxígeno disuelto; con respecto al díptero Chironomus sp, se desconoce la época de mayor abundancia, pero según Leadley (1977), éste organismo está adaptado para vivir en el lodo, rico en materia orgánica y con poco oxígeno disuelto, condiciones similares a las que presenta el área de estudio durante la época de sequía. Con relación al grupo alimenticio correspondiente a Callinectes spp, este ha sido identificado como parte de la dieta de ejemplares jóvenes de Centropomus spp (Chávez, 1963). Al parecer C. robalito ingiere el Detritus al depredar sobre otros organismos epibénticos. En relación a Thyri crystallina, este aterínido según lo señala Amezcua (1977), es un visitante ocasional que se presenta en la época de precipitación; sin embargo, en este estudio apareció en la época de sequía debido a la introducción de agua dulce al sistema a través de los caudales de los ríos Baluarte y Presidio. En la época de precipitación (julio-diciembre), el predominio de Penaeus spp es evidente, y a pesar de la disponibilidad de elementos alimenticios diversos, C. robalito tiene marcada selectividad por los camarones peneidos. Los Restos de Peces continuan ocupando un lugar relevante, de donde se desprende que este componente alimenticio es importante en la dieta de la especie citada. Los Restos de Crustacea, ocupan un lugar especial en la dieta de C. robalito, pues como se apuntó en casos anteriores, es probable que dentro de este grupo trófico se incluya Penaeus spp. El langostino Macrobrachium tenellum, constituye un componente trófico abundante en esta época; comunmente se le encuentra protegido entre la vegetación sumergida y es presa fácil de sus depredadores dada su lenta locomoción. A pesar de la predominancia de los individuos adultos sobre los juveniles, al parecer las tallas no influyen en el vacío o llenado del estómago. En la mayor parte de la captura predominaron los estómagos vacíos sobre los llenos debido probablemente, a que C. robalito al ser atrapado regurgita el alimento. Sin embargo, la prueba estadística aplicada a esta especie demostró que la actividad alimenticia se realiza esencialmente en la noche. Carranza y Amezcua (1971a) señalan como alimento preferencial de C. robalito a los peces, crustáceos y en menor grado insectos acuáticos. La diferencia entre los resultados de estos autores y los obtenidos en ésta investigación, se atribuyen a que su estudio no fue continuo y no abarcó los regímenes de estío y precipitación de casi dos años de estudio. C. robalito es residente estacional del Sistema Huizache-Caimanero y de acuerdo con la clasificación de Day et al. (1973), es una especie netamente carnívora. Por otra parte Yañez-Arancibia (1978) considera a C. robalito en las lagunas de costeras de Guerrero, como un consumidor de tercer orden que incorpora a su dieta peces, crustáceos, moluscos y accidentalmente detritus. La depredación de C. robalito sobre las poblaciones de los camarones peneidos, se registró en la mayor parte de la época de precipitación, intensificándose en los meses de agosto a noviembre, temporada en que se presenta la máxima abundancia de juveniles de Penaeus spp; en estos meses se debe capturar a C. robalito para reducir su acción depredadora, coincidiendo que en este periodo alcanzan tallas comerciales. Durante la épocas de sequía (enero - junio) el grupo alimenticio Detritus, sobresalió como predominante; anteriormente ya se ha destacado la importancia de este integrante trófico en la dicta de A. caerulescens, C. xanthulus y C. robalito. Yañez-Arancibia (1979), señala que C. nigrescens, ingiere, el Detritus de manera accidental, pero en éste casa el detritus se le encontró como único integrante del contenido estomacal, lo cual sugiere que en tu épocas de carencia de alimento, C. nigrescens depende casi exclusivamente de este material. Los Restos de Peces, como se ha discutido anteriormente en A. caerulescens, es uno de los elementos al parecer más accesible tanto en los periodos de estío como de precipitación. Con respecto a los Mugilidae los juveniles de M. cephalus y M. curema son característicos dentro del Sistema Huizache-Caimanero como resultado de su capacidad eurihalina y euriterma. A pesar de ser un recurso disponible, no se encontró en estómagos de A. caerulescens y en mínimo porcentaje en C. xantulus y C. robalito; la explicación tal vez se deba a que los mugilidos son de natación rápida y eluden con facilidad a sus depredadores. Las larvas de Eucinostomus sp después de los góbidos y cupleidos, es, uno de los elementos más abundantes en el sistema Huizache-Caimanero disponible en ambas épocas (Alvarez, 1984). El grupo trófico de los copépodos es abundante en la sequía, sobre todo en las áreas donde existe sedimento blando, rico en detritus. Yañez y Díaz (1977) registraron copépodos del género Cyclops sp principalmente en el mes de mayo como integrantes alimenticios en los atómagos de Dormitator latifrons. La captura de ejemplares en esta época fue escasa debido al reducido volumen de agua de la laguna. Además C. nigrescens no fue tan frecuente como otras especies. Predominaron los estómagos, llenos sobre los vacíos como resultado de la conducta alimenticia de esta especie. Fagade y Olaniyan (1972), señalan que los peces depredadores tienen hábitos alimenticios irregulares y tienden a tomar grandes cantidades mando sus presas están disponibles. En la época de precipitación (julio-diciembre), el predominio de los Restos de Peces en esta temporada se debe a la ocurrencia masiva de juveniles de peces al área de estudio, este componente se comporta como en el caso de las tres especies discutidas anteriormente. El camarón Penaeus spp, continúa ocupando un lugar especial en la dieta de C. nigrescens no obstante la variedad de otras posibles presas. La desproporción entre estómagos vacíos y llenos observada en C. nigrescens, se debe a que éste regurgita el alimento al ser atrapado. Otra razón es la señalada por Cervigón (1966), quien indica que los "Robalos" no son especies migratorias, por lo que su movimientos son muy limitados. El estudio realizado por Carranza (1969) demostró que el alimento más importante de C. nigrescens fueron los paces y en segundo término los camarones observándose cierta similitud con les resultados aquí expuestos. C. nigrescens es residente estacional del Sistema Huizache - Caimanero y de acuerdo con la clasificación propuesta por Day et al. (1973) es una especie netamente carnívora. Yánez-Arancibia (1978) cataloga a C. nigrescens en tu lagunas costeras de Guerrero como un pez consumidor de tercer orden alimentándose de peces, crustaceos insectos y detritus de manera accidental. La especie C. nigrescens se registró la mayor parte del año dentro del Sistema Huizache-Caimanero y la depredación sobre Penaeus spp, ocurrió principalmente en los meses de septiembre a noviembre; para reducir ésta acción depredadora, sería conveniente iniciar el control de C. nigrescens desde el mes de junio, pues es en este mes mando la especie alcanza tallas hasta de 23.0 cm L. Std.; por otra parte, este pez tiene una gran aceptación en el mercado regional debido a su excelente calidad. Durante la temporada de sequía (enero - junio) los Restos de Peces se destacaron como predominantes en la dieta de E. affinis. En estudios ictiológicos en Lagos Lagoon, Nigeria, Fagade y Olaniyan (1972) señalan que Elops lacena, una especie similar a E. affinis, se alimenta principalmente de peces juveniles. Durante la temporada da sequía la captura de la especie anterior fue muy escasa, ya que E. affinis penetra generalmente al sistema Huizache-Caimanero en la época de precipitación. Durante la época de precipitación (julio - diciembre), el elemento alimenticio sobresaliente en esta temporada estuvo representado por el camarón Penaeus spp. Al estudiar a Elops saurus, una especie característica del Golfo de México pero afín a E. affinis, Günther (1945) indica que de cada 5 ejemplares examinados, 3 contenían camarón del género Penaeus spp. Después del componente alimenticio anteriormente, los Restos de Peces Constituyen una parte importante en la dicta de E. affinis. En esta temporada predominaron los individuos jóvenes y se reconoció que la talla de E. affinis influye en la condición de llmado del estomago. La discrepancia entre especímena con estómago lleno y vacío se debe a que se capturaron mayor número de organismos en el día que en la noche; esto coincide con la prueba estadística aplicada a esta especie, la cual demostró que la actividad alimenticia se efectúa preferentemente durante el día. E. affinis es una especie marina que visita cíclicamente el Sistema Huizache- Caimanero en estadíos adultos y juveniles y de acuerdo con la clasificación de Day et al. (1973), se puede considerar como un organismo netamente carnívoro. Carranza (1969 y 1970) indicó que E. affinis es una especie carnívora que incluye en su dicta principalmente larvas de dípteros, peces y en menor grado los camarones, Sin embargo Yañez-Arancibia (1978) señala a E. affinis como un consumidor de segundo orden y dependiendo de la disponibilidad del alimento, consumidor de tercer orden incorporando a su dieta peces, crustáceos, insectos y en menor importancia vegetales y detritus. La presencia de E. affinis es frecuente en la época de precipitación por lo que ejerce una presión considerable sobre las poblaciones de los camarones peneidos. Se recomienda establecer un control previo a las primera inmigraciones de postlarvas de Penaeus spp al Sistema Huizache-Caimanero, la cual segun Watkins (1980), ocurre en junio y julio. Por otra parte, les pescadores no consumen el producto fresco de E. affinis, sino que lo distribuyen localmente después de procesarlo mediante salado y secado. Durante la temporada de sequía (enero-junio) el grupo alimenticio sobresaliente estuvo representado por Macrobrachium tenellum, el cual es abundante en ambas épocas de sequía y precipitación Edwards (1978). El clupeido Lile stolifera según Warburton (1978) es abundante en los meses de noviembre a marzo. Por otra parte Fagade y Olaniyan (1972) señalan que la especie P. quadrifilis, análoga a P. approximans, a partir de los 25.0 cm de LT es altamente piscívora. Durante este periodo la captura de P. approximans fue muy reducida, debido a que esta especie penetra al Sistema Huizache-Caimanero solo cuando las condiciones hidrológicas aun propicias Castro-Aguirre (1978) señala que P. approximans prefiere salinidades de 30 a 35 ‰; en la época de sequía Moore (1979) registró para el mes de enero, una salinidad de 28 ‰, con solo dos unidades de S ‰ fue suficiente para que el número de ejemplares se redujera considerablemente. En la temporada que comprendió la precipitación (julio-diciembre) el grupo Penaeus spp ocupó un lugar destacado. La captura de este crustáceo es mayor de septiembre a noviembre, decreciendo de diciembre a mediados de febrero; la época de mayor abundancia del camarón dentro del Sistema, coincide en parte con las condiciones hidrológicas favorables para que P. approximans ingrese a la laguna; en este lapso relativamente corto, depreda sobre los camarones de manera intensiva hasta que las condiciones hidrológicas le son adversas y regresa entonces al mar. Por otro lado, Cervigón (1966) da cuenta de un ejemplar de Polydactylus virginicus congénere de P. approximans, con una talla de 13.7 cm de L Std., cuyo estómago contenía 18 camarones juveniles. El palemónido Macrobrachium tenellum tambien ocupa un lugar sobresaliente en la dicta de P. approximans, al igual que en las otras especies de peces que conforman este estudio; éste grupo alimenticio está disponible casi en toda la época del año, incrementando su abundancia en la época de precipitación. En cuanto a las jaibas del género Callinectes spp, como se discutió con anterioridad, son abundantes en la época de sequía, aunque los juveniles de Callinectes spp son poco frecuente en ésta época, P. approximans lo incluye en su dieta. Los Restos de Peces, constituyeron una parte importante en la dieta de esta especie en la época de precipitación. En esta temporada no se registraron individuos jóvenes de P. approximnas. Amezcua (1977) señala que esta especie entra a las lagunas en estado adulto exclusivamente para alimentarse. Esta actividad como lo revela la información contenida en este trabajo, tiene lugar durante la noche y se incrementa ligeramente hacia el alba; es por ésta razón que la gran mayoría de los estómagos analizados contenían alimento. P. approximans es residente estacional del Sistema Huizache-Caimanero y de acuerdo con la clasificación de Day et al. (1973), se considera como un organismo netamente carnívoro; sin embargo, Yañez-Arancibia (1978) considera a P. approximans como un visitante excepcional en tu lagunas costeras de Guerrero y como un consumidor de segundo orden. El ingreso de P. approximans al Sistema Huizache-Caimanero ocurre en los meses de septiembre a noviembre, época en que es mayor la actividad depredadora sobre las poblaciones de Penaeus spp se sugiere por lo tanto como en los casos anteriores, un centro¡ de P. approximans previo a la zafra camaronera El aprovechamiento directo de esta especie es problemático, ya que al ser capturada despide un olor desagradable; unas alternativa de uso es su procesamiento como harina de pescado. Conclusiones1. El espectro trófico de las 6 especies estudiadas exhibió una diversidad considerable en composición la cual fluctuó entre 27 y 8 grupos alimenticios. Las especies A. caerulescens, C. xanthulus y C. robalito mantienen un espectro amplio a través del año, en tanto que la dieta de C. nigrescens, E. affinis y P. approximans es restringida a 13 u 8 componentes. 2. Los drásticos cambios ambientales observados en el Sistema lagunar Huizache-Caimanero ocasionados por el régimen de estío y precipitación ejercen un efecto directo sobre la dieta de las especies depredadoras. Durante la temporada de estiaje la superficie del sistema se reduce hasta un 80% de su área lo cual restringe la disponibilidad y accesibilidad de alimento, y a su vez reduce sensiblemente el espectro trófico de las especies. La temporada de precipitación produce el efecto opuesto, al incrementarse la disponibilidad y variedad de presas potenciales. 3. Las 6 especies estudiadas incluyen en su dieta elementos alimenticios cuya ocurrencia a través de sus fases de desarrollo como juveniles y adultos, es de tipo azarosa; la proporción en la que aparecen dichos elementos en el contenido analizado, mantienen una cierta tendencia, aunque esta no fue estadísticamente significativa. 4. En los 6 casos analizados, se reconocen como componentes tróficos predominantes los peces de las familias Gobiidae Clupeidae, Gerridae, Centropomidae, camarones peneidos, el detritus, los portúnidos del género Callinectes spp, y el palemónido Macrobrachium tenellum. El detritus y las jaibas Callinectes spp representan una fuente alimenticia importante durante la temporada de estiaje. A través de las distintas etapas de su ciclo de vida en el sistema lagunar, las especies investigadas desarrollan competencia teórica interespecífica, dado que su actividad alimenticia incide esencialmente sobre los componentes antes citados. Sin embargo, la proporción en la que estas aparecen en el contenido estomacal varió para cada especie debido a los mecanismos adaptativos (morfológicos/comportamiento) de cada especie para la mejor captación de alimento, lo cual permite la distribución de las fuentes alimenticias del sistema estuarino. 5. Las especies A. caerulescens, C. xanthulus y C. nigracens incluyen en su dieta a los camarones peneidos, pero su presión de depredación sobre este componente depende de su disponibilidad; las tres especies anteriores no son teóricamente selectivas. En contraste, C. robalito, E. affinis y P. approximans incorporan a su dieta como elemento esencial a los peneidos,independientemente de su disponibilidad. 6. Las diferencias observadas en la condición de llenado del estómago de las especies estudiadas varían según la composición por talla de las poblaciones capturadas; sin embargo, tal variación no resultó significativa en la mayoría de los casos analizados. 7. La actividad trófica de las especies A. caerulescens, C. xanthulus y C. robalito tiene lugar indistintamente durante el día como en la noche, aunque bajo condiciones de oscuridad esta actividad se incrementa notablemente. En el caso contrario se observa en C. nigrescens y E. affinis, cuya acción depredadora es preferentemente diurna; ambas especies requieren el reconocimiento visual de sus presas, entre las que destacan los camarones pencidos. La especie P. approximans, que ingresa al sistema lagunar por razones de carácter trófico en su fase adulta, ejerce una importante presión depredadora sobre los camarones peneidos durante los períodos de oscuridad la cual se incrementa ligeramente hacia el alba. 8. Se reconoce la necesidad de incrementar el esfuerzo pesquero sobre las 6 especies estudiadas con el propósito de atenuar su presión depredadora sobre las poblaciones de camarones peneidos explotados comercialmente en el sistema lagunar Huizache-Caimanero, a través del período de precipitación. El consumo directo o la industrialización de las especies en cuestión ofrece perspectivas de desarrollo regional. AgradecimientosLos autores agradecen a A. Ramírez Flores, G. Green, A. Ortega y A. Calderón, por sus indicaciones y revisión crítica del manuscrito. A B. Blake, Jefe del Proyecto Anglomexicano de colaboración auspiciado por la Overseas Development Administration y su grupo integrado por: A. Menz, N.H. Moore, J.L. Watkins y J. McD. Mair, por su ayuda y cooperación. A J. Alvarez Cadena, A. Toledano y A. Núñez por su participación en las colectas y procesado del material. El primer autor expresa su reconocimiento a A. Vázquez-Botello, responsable del Laboratorio de Contaminación Marina por las facilidades otorgadas para realizar este trabajo. LITERATURAÁLVAREZ GUILLÉN, H., A. YÁÑEZ-ARANCIBIA y A. L. LARA-DOMÍNGUEZ An. Inst. Cienc. Del Mar y Límnol. Ecología de la Boca del Carmen. Laguna de Términos. El hábitat y estructura de las comunidades de peces (sordas Golfo de México). Univ. Nal. Autón. México 1985. 107-44. 12 (1): ÁLVAREZ-CADENA, J. N., M. A. AQUINO, F. ALONSO, J.G. MILLAN y F. TORRES, An. Ints. Cienc. del Mar y Limnol. Composición y abundancia de las larvas de peces en el sistema lagunar Huizache-Caimanero. Parte I Agua Dulce 1978. Univ. Nal. Autón. México 1984. 163-180. (1): AMEZCUA, L. F., An. Centro Cien. del Mar y Limnol. Generalidades ictiológicas del sistema lagunar costero de Huizache-Caimanero, Sinaloa. México Univ. Nal. Autóm. México, 1977. 1-25. 4 (1): AYALA-CASTAÑARES, A., M. GUTIERREZ y V. M. MALPICA, Informe final de los estudios de Geología Marina en las regiones de Yavaros, Son., durante la primera etapa. 2o. Informe del Contrato de Estudios No. EI-69-93. Univ. Nal. Autón. México, Inst. Biol. Dpto. Cienc. del Mar y Limnol. y SRH: 1970. 3-190. ARENAS-FUENTES, V. Informe final de las investigaciones correspondientes a hidrología y productividad en los planes piloto Escuinapa y Yavaros. 2o. Informe del Contrato de estudios No.69-93. Univ. Nal. Autón. México, Inst. Biol. Depto. Carne. del Mar y Limnol. y SRH: 1970. 191-223. BLAKE, B. F., J. Fish. Biol, Food a feeding of the mormyrid fishes of Lake Kainji, Miseria, with special reference to seasonal variation and interspecific differences. 1977. 315-328. II: BLAKE, B. F., A. B. BOWERS y E. NAYLOR, Ecology and Penaeusfishery of Mexican Lagoons Overseas Development Administration. London. 1981. 58 p. BRUSCA, R. C., A handbook to the common intertidal invertebrata of the Gulf of California. University Arizona Press. Tucson, Arizona. 1973. 1-427 p BERDEGUE, J., Tesis Prof. Contribución al conocimiento de los peces de importancia comercial en la costa noroccidental de México. Esc. Nal. Ciencias Biológicas, IPN México 1954. 360 p. CASTRO AGUIRRE, J. L, Catálogo Sistemático de los Peces Marinos que penetran a las Aguas Continentales de México con Aspectos Zoogeográficos y Ecológicos. Serie Científica Dirección General Instituto Nacional de Pesca, 1978. 1-298. 19: CARRANZA, J., Reconocimiento preliminar de la fauna ictiológica en las áreas de los planes piloto Escuinapa y YáYaros y programa de las investigaciones, 1er. Informe del Contrato de Estudio No. E1-69-51. Univ. Nal. Autón. México. Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH. 1969a. 31 p. CARRANZA, J., Primer Informe sobre el estudio taxonómico de la fauna de peces de los planes piloto Escuinapa y Yávaros 2o. Informe de Contrato de Estudio No. El-69-51, Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH, 1969b. 39 p. CARRANZA, J., Informe final sobre la primera etapa del estudio de la fauna ictiológica y depredaciones del camarón en las lagunas y esteros de los planes piloto Escuinapa,Sin., y Yávaros,Son., Informe final Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH. 1970. 28 p. CARRANZA, J. y F. AMEZCUA L., Plan Nayarit, SRH Informe de avance de trabajo, incluyendo información ecológica básica datos de distribución de postlarvas e informe preliminar sobre la fauna ictiológíca. [2a. Parte del 2o. Informe de Contrato de Estudios No.] NAY-EST-7, Univ. Nal. Auton. México., Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH: 1971a. 58-80. CARRANZA, J. y F.AMEZCUA L, Plan Nayarit, SRH Resultados finales de hidrologia, plancton y fauna ictiológica en el sistema Teacapán-Agua Brava (octubre 1970- junio 1971) 2a. Parte Del Informe Final del Contrato de Estudios No. NAY-EST7 Univ. Nal. Autón. México Inst. Biol. Depto. Ciene. del Mar y Limnol. y SRH: 1971b. 88-115. CARRANZA, J. y F.AMEZCUA L, Informe final sobre la primera etapa del estudio de la fauna ictiológica y depredaciones del camarón en las lagunas y esteros de los planes piloto Escuinapa, Sin., y Yávaros, Son., Informe final Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH. 1970. 28 p. CABRERA, J., Informe sobre el programa de la biología del camarón, II Migración. 10o. Informe del Contrato de Estudio EI-68-115. Inst. Biol. Univ. Nal. Autón. México y SRH 1970. CHAPA, H., y R. SOTO, Lagunas Costeras, un Simposio Mem Simp. Int. Lagunas Costeras, Resultados preliminares del estudio ecológico y pesquero de las lagunas litorales del sur de Sinaloa. In UNAM-UNESCO, México, D.F. Noviembre 28-30 1969. 653-662 1967: . CHÁVEZ, H., Ciencia Méx. Contribución al conocimiento de la biología de los robalos, chucumite y constantino (centropomus spp) del estado de Veracruz (Pisces: Centropomidac). 1963 141-160. 22 (5): CERVIGÓN, F., Los peces Marinos de Venezuela . Estación de Investigaciones Marinas de Margarita Fundación La Salle de Ciencias Naturales Caracas, Monog. 1966. 1-951, 385 figs. 11 y 12: CRISP, D.T., Salm Trout Mag, A preliminary survery of brown trout (Salmo trutta L.) and bullheads (Cottus gobio L.) in high altitude becks. 1963. 45-59. 167: CORBET, P. S., Proc. Zool. Soc. Lond. The food of non-cichlid rishes in the Lake Victoria basin with remarks on their evolution and adaptation to lacustrine conditions. 1961 1-101. 136: DARNELL, R. M., Publ. Inst. Mar. Sci Univ. Texas Food habits of fishes and larger invertebrates of Lake Pontchartain, Louisiana, and estuarine community. 1958. 353-416. 5: DARNELL, R. M.y MEIEROTTO, R. R. . Tans. Am. Fish Soc., Determination of feeding chronology in fishes 1962. 313-320. 91: DAY, J. W., W. G. SMITH. P. WAGNER y W. STOWE, Communiti structure carbon budget of a salt marsh and shallow bay estuarine system in Louisiana. Publ. No.I LSU-56-72-04. Center for Wetiand Resources Louisiana State University, Baton Rouge, La. 1973. 79 p. EDWARDS, R, R, C., Estuar. Coast. Mar. Sci. Ecology of a costal lagoon complex in Mexico. 1978. 75-92. 6: FAGADE, S. 0. y OLANIYAN, C. I. 0., J. Fish. Biol. The food and feeding interrelationship of the fishes in the Lagos Lagoon, Nigeria. 1972. 205-225. 5: FROST, W. E., J. Fish. Biol. The food of Charr salvelinus willughbii (Gúnther)in Windemere. 1977. 531-547. GARCÍA-CUBAS, A., Informe sobre el avance de trabajo en el programa de estudio de los moluscos en los planes piloto Yavaros y Escuinapa, 2o. Informe del Contrato de Estudio No. El-69-9 Univ. Nal. Auton. México. Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH: 1970. 347-371. GARCÍA, E., Modificaciones al sistema de clasificación climática de Köppen. Instituto de Geografía, Univ. Nal. Autón. México, 1973. 246p. GÓMEZ-AGUIRRE, S., C. FLORES-COTO, S. LICEA-DURÁN, H. SANTOYO y A. MARTINE7, Informe final de la 2a. Etapa de los estudios del plancton en los planes piloto Escuinapa-Yávaros. 2o. Informe del Contrato de Estudio No. EI-69- 93. Univ. Nal. Autón. México. Inst. Biol. Depto. Cienc. del Mar y Limnol. y SRH: 1970. 234-286. GÓMEZ-AGUIRRE, S., S. LICEA-DURÁN y C. FLORES COTO, An. Centro Cienc. del Mar y Limnol. Ciclo anual del plancton en el sistema Huizache-Caimanero, México. (1969-1970). Univ. Nal. Autóm. México, 1974. 83-98. 1 (1): GONZALEZ, V. L. I, Tesis profesional, Aspectos biológicos y distribución de algunas especies de peces de la familia Ariidae de las lagunas litorales del noroeste de México. Fac.de Ciencias,Univ. Nal. Autón. México, 1972. 88 p., 33 figs. 30 tablas. GREENWOOD, P. H. D. E. ROSEN, S. H. WEITZMAN y G. S. MYER, Bull. Am. Mus. Nat. Hist. Phyletic studics of teleostean fishes, with a provisional classification of living forms 1966. 341-455. 131 (4): GÜNTER, G., Studies of marine fishes of Texas. Publ. Inst. Mar. Sci. Univ. Texas, 1945. 1-190. 1 (1): HYNES, H. D. N., J. Anim. Ecol., The food of freshwater sticklebacks (Gasterosteus aculcatus and Pygosteus pungitis) with a review of methods used in studies of the food of fishes. 1950. 36-58 19:. HYSLOP, P. J, J. Fish Biol., Stomach contents- a review of methods and their application. 1980. 411-429. 17: JOHNSON, F. H., J. Fish. Res. Ed. Can, Responses of Walleye (Stizotedion vitreum vitreum) and Yelleow Perch (Perca flavercens) populations to the removal of White Sucker (Catogomus commersoni) from a Minnesota lake 1966. 1977. 1633-1642. 34: JORDAN, D. S. y B. W. EVERMANN, Proc. U.S. Nat. Mus, The fishes collected by Lieut. Henry E. Nichols, U.S.N., in the Gulf of Califonia and on the west coast of Lower California, with description of four new spccies. 1896-1900. 273- 279. 4: LEADLEY, B. A., Ecology of Fresh Water . Harvard University Press 1977. 129 p. MANN, R. H. Y, y ORR, D. P, J. Fish. Biol. A preliminary study of the feeding relationship of fish in a hard-water and a softwater stream in Southern England. 1969. 31-44. 1: MENZ, A., Thesis Penacid shrimps in a Mexican lagoon Ph. D. University of Liverpool. 1976. 136 p. MENZ, A. y BLAKE, B.F., J. Exp. Mar. Biol. Ecol. Field and laboratory observationa of growth in Penaeus vannamei Boone from a Mexican costal lagoon system. 1980. 99-111. 48: MELCHOR, A. J. M., Tesis profesional, Estudio sobre la biología y ecología de los Arius caerulescens (Günther) y Arius liropus (Bristol) del estero de El Verde y Laguna de Caimanero. Sinaloa (Pisces: Ariidae). IPN, Centro Interdísciplinario de Ciencias Marinas (CICIMAR) La Paz, Baja Calif. Sur, 1980. 39 p. MINELLO, T. J., y P. J. ZIMMERMAN, Cynos nebolosus. Contributions in Marine Science, Selection for brown shrimp, Penaeus aztecus as prey by the spotted scatrout, 1984. 27. (en prensa) MOORE, N. H., Ph. D. thesis, The annual physical hydrographic cycle of a tropical logoon system of the Pacific Coast of México. University of Liverpool. 1979. 323 p. MOODY, W. D., Quart. Jour. Fla. Acad. Sci., A study of the natural history of the spotted trout, Cynoscion nebulosus, in the Cedar Key, Florida area. 1950. 147-171. 12 (3): NIKOLSKY, G.V., The Ecology of Fishes. Academic Press London, New York 1963. 352 p. ODUM, W. E., Sea Grant Program Tech. Bull, Patways of energy flow in a South Florida estuary. 1971. 1-162, 7: ODUM, W. E. y E. J. HEALD Bull. Mar. Sci. Trophic analises of an estuarine mangrove community. 1972. 671-738. 22 (3): ORTEGA, G. M., Informe de avance del estudio de la vegetación sumergida en los planes piloto Yávaros y Escuinapa. 2o. Inf. Contrato de Estudio No. EI-69-93. Univ. Nal. Autón. México. Inst. Biól. Depto. Cienc. del Mar y Limnol. y SRH: 1970. 287-346. ORTÍZ, F., La Pesca en México Fondo de Cultura Económica, Colección Testimonios del Fondo 1975. 1-63 (31) PAUL, R. K, G., Ph. thesis, Bionomics of Crabs of the genus Callinectes (Portunidae) in a lagoon complex on the Mexican Pacific coast. University of Liverpool, 1977. 136 p. PEREZ, I., Serie Div. Claves ilustradas para la identificación de los camarones comerciales de la America Latina. Inst. Nal. Inv. Biol. Pesq. SIC; 1970. 50 p. (3): RADFORTH, L, J. Animal. Ecol, The food of grayling Thymallus thymallus, flounder Platichthys roach Rutilus rutilus and gudgeon Gobio gobio with special reference to the Tweed watershed. 1940. 302-318. 9: SALAZAR, T. N., Tésis prof. Contribución al conocimiento de la biología y algunos aspectos pesqucros de dos especies de jaibas, Callinectes arcuatus 0rdway (1863) y Callinectes toxotes 0rdway (1863), de la Laguna de Caimancro, Sinaloa, México. CICIMAR, IPN La Paz, B.C. Sur, 1980. 79 p. SHERIDAN, P. F., y D. L. TRIMM, Fish. Bull., Summer foods of Texas coastal fishes relative to age and habit. 1985. 643-647. 81 (3): STARCK, W. A. y E R. SCHROEDER, Investigations on the Gray snapper Lutjanus griseus University of Miami Press Coral Gables, Fla. 1970. 1-221. SOTO, R., Tesis prof. Mecanismo hidrológico del sistema lagunar Huizache-Caimanero y su influencia sobre la producción camaronera. Esc. Sup. Ciencias Marinas Univ. Aut. B.C. 1969. 80 p. TAMAYO, J., Geografía General de México. Talleres Gráficos de la Nación. 1940. 1-628; 1-580. 1;,2: THOMAS, J. D., J. Zool. On the biology of the catfish Clarias senegalensis in a man-made lake in the Ghanaian savanna with particular reference to its feeding habits. 1966. 476.514. 148: WARBURTON, K., Estua. Coast. Mar. Sci. Community structure, abundance and diversityof the fish of a Mexican coastal lagoon. 1978. 497-519. 7: WARBURTON, K, J. Fish Biol., Growth and ecology of some important species of fish in a Mexican coastal lagoon system. 1979 449-464. 14: WATKINS, J. L, Ph. D. thesis, The inmigration of Postlarval Penacus shrimp into a lagoon system on the Pacific coast of México. University of Liverpool, Port Erin, Isle of Man. Great Britain. p. 1980. 1-200 WEATHERLEY, A. H., Growth and Ecology of Fish Populations. Academic Press. London. New York. 1972. 293 p. YÁÑEZ-ARANCIBIA A. y G. DIAZ-GONZALEZ, An. Centro Cienc. del Mar y Limnol. Ecología trofodinámica de Dorminator latifrons (Richardson). Univ. Nal. Autón. México 1977. 125-140. (1): 4 YÁÑEZ-ARANCIBIA, A., Centro Cienc. del Mar y Limnol. Taxonomía,ecologíca y estructura de las comunidades de peces en lagunas costeras con bocas efiffieras del Pacífico de México Publ. Esp. Univ. Nal. Autón. México 1978. 1-306. 2: ZAR, J. H., Biostatistical Analyisis. Prentice-Hall Inc. 1974 620 p.
|