|
CLADÓCEROS DEL EMBALSE RÍO TERCERO (PROVINCIA
DE CÓRDOBA, ARGENTINA)
Trabajo recibido el 26 de septiembre de 1985
y aceptado para su publicación el 26 de febrero de 1987.
Alicia H. Escalante
Carrera del Investigador (CONICET), Museo de La
Plata, Casilla de correos 55, Buenos Aires, Argentina. Contribución
Científica 275 del Instituto de Limnología "Raúl A.
Ringuelet" (CONICET-UNLP).
En el embalse Río Tercero se identificaron seis
especies de cladóceros planctónicos: 2 especies del género
Bosmina, 2 de Daphnia, 1
Ceriodaphnia y 1 Diaphanosonia.
Además, se registraron ocasionalmente una especie de
Moina y dos especies litorales pertenecientes a la familia
Chydoridae. Se estudió la distribución horizontal y vertical de
estas especies en cinco estaciones de muestreo, así como su
variación estacional durante el periodo comprendido entre julio de 1977
y mayo de 1981. B. huaronensis es la especie más
abundante y C. dubia la más frecuente entre los
cladóceros del embalse. La tendencia general de las especies es aumentar
sus poblaciones hacia la primavera y declinar durante el verano. B.
longirostris y C. dubia presentan registros altos
también en el otoño. En el caso de Diaphanosona
brachyurum, no suele estar limitada al verano, a diferencia de lo
indicado por Margalef (1983). En cuanto a la distribución vertical,
B. huaronensis es más abundante entre 1 y 7.5 m de
profundidad; B. longirostris a 1 m; C.
dubia y D. brachyurum entre 1 y 5 m.
Palabras clave: Cladóceros planctónicos,
distribución horizontal y vertical, embalse Río Tercero
The planktonic cladoceran fauna in the man-made lake
of Río Tercero comprises two species of Bosmina, two
of Daphnia, a Ceriodaphnia and a
Diaphanosoma. Besides, two littoral species of Chydoridae
and a Moina occur. Bosmina huaronensis
and Ceriodaphnia dubia are the most important species
related respectively to their abundance and frequency occurrence. B.
huaronensis and Diaphanosoma brachyurum are more
abundant at the sampling station A; Bosmina longirostris and
C. dubia at C; Daphnia laevis and
D. ambigua at B, showing some irregularity on their
horizontal spatial distribution. The usual tendency of these cladoceran species
is to increase their populations through spring and to decline during summer.
B. longirostris and C. dubia also present
high values in autumn. In relation to their vertical distribution, B.
longirostris at 1 m; C. dubia and D.
brachyurum between 1 and 5 m. depth.
Key words: Planktonic cladoceran fauna, horizontal and
vertical distributions, Río Tercero dam.
La información existente acerca de la tipología de los embalses y composición de sus comunidades bióticas está referida, por lo general, a ambientes artificiales de Africa y la Unión Soviética, donde se encuentran los mayores embalses del mundo, y menos frecuentemente a los de Checoslovaquia, España, América del Norte, Bulgaria, entre otros. En Argentina, el conocimiento de estos "lagos" es relativamente escaso, si bien los de la región central han sido objeto de no pocas investigaciones. Entre ellas, la realizada por Guarrera (1948) incluye el estudio del fitoplancton del embalse San Roque, Córdoba, y de los ríos y arroyos tributarios del mismo, con la caracterización físico-química del agua. Cordini (1949) proporciona un panorama general de los aspectos más importantes del embalse Río Tercero, al tratar su morfometría, térmica, composición química del agua, materiales del fondo y los organismos que allí viven y la comunidad del plancton. Señala los géneros Daphnia y Bosmina como cladóceros integrantes del zooplancton. Es probable, por lo que se conoce hoy día, que estos géneros hayan estado representados por más de una especie cada uno, y que la existencia de sólo dos géneros distintos de cladóceros se deba a que aún no se habían adaptado a este tipo de ambiente. Una década después, Boschi y Fuster de Plaza (1959) citan para el embalse Río Tercero a Daphnia spinulata, Diaphanosoma brachyurum, Bosmina obtusirostris y Ceriodaphnia dubia. Más recientemente, Paggi contribuye a clarificar la ubicación sistemática de los cladóceros mediante descripciones detalladas de varias especies (1973, 1977) y revisiones de las especies argentinas de los géneros Diaphanosoma y Bosmina (1978, 1979), la mayoría de las cuales se hallan presentes en el embalse Río Tercero. Bonetto et al. (1976) realizaron un detallado estudio de los caracteres limnológicos de 3 embalses de la provincia de Córdoba: el San Roque, Los Molinos y el del Río Tercero, incluyendo datos sobre su hidrología, geología, térmica, química, y composición de las comunidades. Con respecto a los cladóceros, estos autores indican la existencia de 8 especies para los 3 embalses, pero sólo 3 de ellas para el embalse Río Tercero. Mencionan además la familia Chydoridae, de hábitos no planctónicos, registrada circunstancialmente en el muestreo del macrobentos. Armengol (1978b), en su estudio sobre los crustáceos planctónicos de 104 embalses españoles, reconoce 64 especies, de las cuales más del 50% son consideradas características de habitats no planctónicos (litoral, bentónico, neustónico). En cuanto a las especies litorales en particular, el mismo autor (1980) señala que, en algunos casos, son frecuentes en el plancton aunque en bajo número. Explica su presencia, en gran parte, porque son arrastradas por los ríos o por una expansión temporal hacia aguas libres propiciada por la ausencia de especies genuinamente planctónicas. En su reciente tratado de limnología, Margalef (1983) dedica un capítulo al estudio de los embalses. Entre los cladóceros menciona la existencia de una Daphnia, a veces Ceriodaphnia, casi siempre Bosmina, y Diaphanosoma que suele estar limitada al verano. Concluye que si se compara este plancton con el lacustre, se observa en los embalses una mayor proporción de cladóceros filtradores de partículas muy pequeñas, como son los de los géneros arriba mencionados. En la presente contribución se identificaron 9 especies de cladóceros, 2 de ellas de presencia casi constante, 4 menos frecuentes, una especie registrada en una sola oportunidad y 2 especies litorales aparecidas ocasionalmente en las muestras de plancton. De las 6 especies planctónicas más frecuentes se estudió la distribución horizontal y vertical así como su variación estacional. ÁREA DE ESTUDIOEl embalse Río Tercero está situado a 32° 11' Lat. Sur y 64° 13'W, a una altura de 548 m sobre el nivel del mar, en el departamento de Calamuchita, Córdoba. Una descripción más detallada de este lago artificial, con características físicas, químicas y biológicas generales, puede verse en Boschi y Fuster de Plaza (1959) y en Bonetto et al. (1976). MATERIAL Y MÉTODOSEl muestreo abarcó un periodo de cuatro años, desde julio de 1977 a mayo de 1981 y se realizó en forma bimensual. Se establecieron 5 estaciones de muestreo: A (Confluencia), B (Camino), C (Garganta), D(Obelisco) y E (Dique), consideradas en conjunto representativas de todo el embalse, por abarcar lugares de confluencia de distintos ríos y arroyos en unos casos, por estar situadas en la parte más angosta del cuerpo de agua o cerca de la represa en otros (Fig. 1). Durante los dos primeros años las muestras de plancton fueron obtenidas por bombeo con un motor a explosión acoplado a una bomba centrífuga y un tubo de plástico de 25 m, marcado metro a metro. Se tomaron muestras a diferentes profundidades en cada una de las estaciones: a 0.05 m de la superficie, 1, 2, 5, 10, 15 y 20 m; estas dos últimas cuando la profundidad del embalse lo permitía ya que hubo ocasiones en que el nivel del espejo de agua no superó los 10 m. En la estación C se extrajeron además muestras a 7.5 m de profundidad. De cada profundidad se filtraron 50 litros de agua a través de una red de plancton de 35 micrones de abertura de malla. A partir de enero de 1979 hasta mayo de 1981 se tomaron en cada una de las estaciones muestras integradas de toda la columna de agua, desde el fondo hasta la superficie mediante un bombeador eléctrico sumergible. En ambos casos se fijaron las muestras con formol al 3% y se fraccionaron, previa homogenización, en cuatro partes iguales con el fraccionador Folsom. Se reconocieron 9 especies de Crustacea Cladocera pertenecientes a 7 géneros incluidos en 5 familias: Diaphanosoma brachyurum (Lievin, 1848) (Fam. Sididae); Daphnia laevis Éirge, 1878, D. ambigua Scourfield, 1947 y Cepiodaphnia dubia Richard, 1895 (Fam. Daphnidae); Moina micrura Kurz, 1874 (Fam. Moinidae); Bosmina huaronensis Delachaux, 1918 y B. longirostris (O.F. Müller, 1785) (Fam. Bosminidae); Alona pulchella var. canibouei (De Guerne y Richard, 1893) y Chydorus pubescens Sars, 1901 (Fam. Chydoridae). 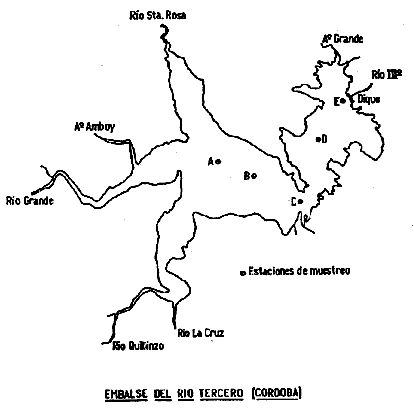 Figura 1. Ubicación de las estaciones de muestreo en el embalse Río Tercero. Con los datos de abundancia de estas especies, expresados en org. 10-3 m-2, se elaboraron curvas de distribución temporal para cada una de las 5 estaciones (distribución horizontal), graficadas en papel semilogarítmico (Figs. 2-7). Con los datos de recuento de las muestras obtenidas a distintas profundidades en la estación C (Garganta) durante los dos primeros ciclos anuales (julio 1977-mayo 1979) se elaboraron gráficos de distribución vertical (Figs. 8-12). Para realizar tales gráficos se consideró el dato integrado de la columna de agua como el 100% y se estableció en qué proporción se encuentra cada una de las especies a distintas profundidades. RESULTADOS Y DISCUSIÓN
Bosmina huaronensisDistribución horizontal: en la estación A (Confluencia), B. huaronensis presenta un comportamiento similar en los cuatro ciclos anuales estudiados. Declina hacia el verano y aumenta su población hacia el invierno. Presenta 3 registros máximos de número de individuos: el primero en septiembre de 1978 con 1,640 org. 10-3 m-2, el segundo en julio de 1980 con 1707 y el tercero en marzo de 1981 con 1168. Su mínimo registro ocurre en mayo de 1981 con 8.1 org. 10-3 m-2 (Fig. 2, A). En la estación B (Camino) dicha especie declina en noviembre de 1979 y enero de 1980, disminuyendo su número hacia el verano de los otros ciclos. Pueden señalarse 2 registros máximos: en septiembre de 1978 con 705 org. 10-3 m-2 coincidente con el primer registro indicado para la estación A y en marzo de 1981 con 624 en coincidencia con el tercer registro de la estación A. Sus registros mínimos se señalan en mayo de 1978 con 10 org. 10-3 m-2 y en mayo de 1981 con 13 org. 10-3 m-2. Este último coincide con el mínimo observado en la estación A (Fig. 2, B). En la estación C (Garganta), B. huaronensis declina en noviembre de 1979, enero de 1980 y mayo de 1981. Se observan 3 registros máximos; el primero en septiembre de 1978 con 1068.4 org. 10-3 m-2 al igual que en las estaciones A y B; el segundo en julio de 1977 con 896.6 org. 10-3 m-2 y el tercero en noviembre de 1980 con 828 org. 10-3 m-2. Los registros mínimos se señalan en enero de 1978 con 25.7 org. 10-3 m-2, mayo de 1979 con 24 julio de 1980 con 22.8 y enero de 1981 con 28 org. 10-3 m-2 (Fig. 2,C). En la estación D (Obelisco), esta especie declina en enero de 1978, enero de 1980 y mayo de 1981, disminuyendo hacia el verano de 1979. De acuerdo a los datos graficados en la curva de distribución temporal, expresados en org. 10-3 m-2 , B. huaronensis presenta 2 picos máximos: en julio de 1977 con 631.6 y en septiembre de 1980 con 744. El pico mínimo se observa en mayo de 1978 con 10 (Fig. 2, D). En la estación E (Dique), dicha especie declina sólo durante septiembre de 1979 y enero de 1980. Se observa un pico máximo en noviembre de 1978 con 576 org. 10-3 m-2. Un pico mínimo ocurre en enero de 1978 con 5 org. 10-3 m-2 (Fig. 2, E). De acuerdo a los datos de abundancia obtenidos desde julio de 1977 a mayo de 1981, B. huaronensis es más abundante en la estación A, siguiendo en orden decreciente, las estaciones C, D, B y E. Si se tiene en cuenta el promedio del no. de org. 10-3 m-2 de las 5 estaciones, el mayor número se observa en septiembre de 1978 con 794.7. Excepto enero de 1980 en que no se registró la presencia de esta especie en los recuentos realizados, el número menor se observa en noviembre de 1979 con 8. 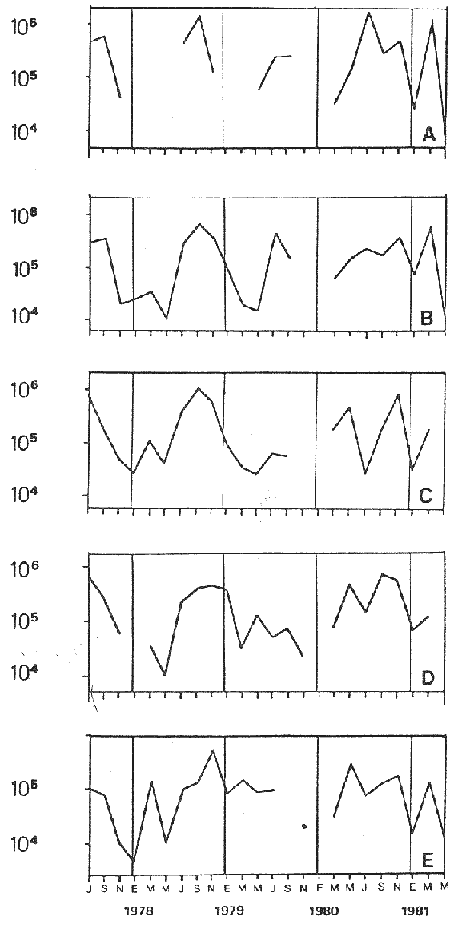 Figura 2. Distribución horizontal de Bosmina huaronensis. A = estación Confluencia; B = estación Camino; C = estación Garganta; D = estación Obelisco y E = estación Dique. Distribución vertical: los gráficos de distribución vertical permiten apreciar que durante el primer invierno y primavera, B. huaronensis es más abundante entre 1 m y 7.5 m de profundidad. Durante el primer verano, en cambio aparece en mayor número hacia los 20 m. En el otoño e invierno de 1978 vuelve a ser más abundante a 1 m. A comienzos de la primavera del segundo año aparece en mayor número a los 2 m mientras que hacia finales lo hace hacia los 10 m. Durante el segundo verano estudiado es más abundante a los 7.5 m y en el otoño a los 10 m. B. huaronensis no presenta un comportamiento similar en los dos ciclos anuales estudiados, en cuanto a su distribución vertical, siendo en general más abundante entre 1 y 7.5 m de la columna de agua (Fig. 8). Bosmina longirostris Distribución horizontal: en la estación A, B. longirostris presenta un comportamiento más o menos similar durante el periodo estudiado. Declina hacia el verano y aumenta su número hacia el invierno. Presenta 3 picos máximos de no. de org. 10-3 m-2: el primero en julio de 1978 con 1-50; el segundo en septiembre de 1978 con 205 y el tercero en julio de 1980 con 153. Su pico mínimo ocurre en marzo de 1981 con 8 (Fig. 3, A). En la estación B, esta especie declina hacia el verano de los cuatro cielos, estando presente sólo en julio y septiembre de los 4 años además de noviembre de 1977, enero y mayo de 1980 y marzo de 1981. Existen 2 picos de mayor no. de org. 10-3 m-2: en julio de 1978 con 100, coincidente con el primer pico ocurrido en la estación A y en septiembre de 1978 con 140 en coincidencia con el segundo pico señalado para la estación A. Pueden indicarse 2 picos mínimos: en enero de 1978 con 5 y marzo de 1981 al igual que el pico mínimo señalado en la estación A (Fig. 3, B). En la estación C, B. longirostris se comporta de manera algo diferente ya que recién declina en enero de 1979 para luego hacerlo en mayo y julio del mismo año; y en el verano e invierno del ciclo siguiente. Sin embargo, durante el primer ciclo anual de permanencia constante, sigue la misma tendencia demostrada en las anteriores estaciones. Es decir, disminuye su número hacia el verano y aumenta hacia el invierno y primavera. Se observan 2 registros máximos: en julio de 1977 con 317.4 org. 10-3 m-2 y en mayo de 1980 con 288. Ninguno de estos picos coincide con los señalados en las estaciones A y B. El pico mínimo está dado en noviembre de 1977 con 3 org. 10-3 m-2 (Fig. 3, C). En la estación D, el comportamiento de esta especie es similar al indicado para la estación C. No sólo declina durante el verano de los 4 ciclos anuales sino que también lo hace en julio de 1979 y 3 1980. Se observa el máximo registro del no. de org. 10-3 m-2 en mayo de 1980 con 360 que coincide con el aumento del número de individuos indicado para la estación C. El pico mínimo, al igual que en las estaciones A y B, ocurre en marzo de 1981 con 4 (Fig. 3, D). 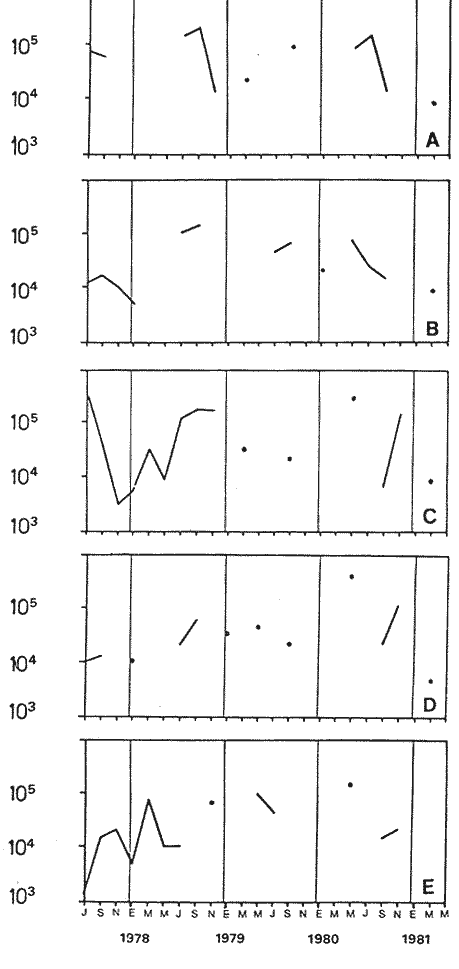 Figura 3. Distribución horizontal de Bosmina longirostris. En la estación E, repite de manera similar la distribución temporal de la estación C, señalada mediante las curvas semilogarítmicas. Su aparición es constante en los recuentos del primer ciclo anual. Declina por primera vez en septiembre de 1978 y luego lo hace en el verano de 1979, 1980, 1981 y en el invierno de 1980. El mayor número de individuos se observa en mayo de 1980 con 168 org. 10-3 m-2 en coincidencia con los picos máximos indicados para las estaciones C y D. El pico mínimo está dado en julio de 1977 con 1.6 (Fig. 3, E). Durante los 4 ciclos anuales muestreados, el mayor número de individuos de B. longirostris se observa en la estación C, siguiendo en orden decreciente la estación A, D, B y E. Tomando el promedio del no. de org. 10-3 m-2 de las 5 estaciones, el mayor número se observa en mayo de 1980, con 194.9. Salvo noviembre de 1979, marzo de 1980, enero y mayo de 1981, meses en los que B. longirostris no se registró en los recuentos del zooplancton del embalse, el número menor se observa en mayo de 1978 con 3.9. Distribución vertical: durante el invierno de 1977 y comienzos de la primavera, B. longirostris es más abundante entre 1 y 5 m de profundidad. En noviembre del mismo año aparece sólo a los 10 m. En el verano de 1978 también abunda a 1 m, declinando a los 5 m para reaparecer sólo a los 15 m. Por el contrario, a comienzos del otoño aparece en mayor número hacia los 20 m. Hacia fines del otoño y durante el invierno, esta especie vuelve a ser más abundante a 1 m. A principios de la primavera de 1978 vuelve a ser más abundante a los 2 m, al igual que en la primavera del primer cielo. A fines de la primavera abunda más en profundidad, hacia los 15 m. En enero de 1979 declina casi totalmente, estando presente sólo en bajo número en la estación D. A comienzos del otoño aparece en mayor número hacia los 15 m. Hacia fines de esta estación vuelve a ser más abundante a menor profundidad, desde la superficie (0.05 m del espejo de agua) hasta los 5 m para volver a aparecer a los 10 m (Fig. 9). Si bien B. longirostris no presenta un comportamiento muy similar en los dos ciclos anuales estudiados, pueden señalarse algunas semejanzas: 1) en los comienzos de la primavera de los dos cielos considerados dicha especie es más abundante a los 2 m; 2) a mediados de primavera se hace más abundante entre los 10 y 15 m; 3) hacia fines del verano es más abundante a mayor profundidad (entre los 15 y 20 m); 4) durante el otoño e invierno de los dos años muestreados vuelve a aparecer en mayor número a 1 m. Armengol (1980) establece que, dentro de las especies euplanctónicas, se encuentran las más comunes de los embalses, que en su mayoría se comportan como dominantes en ciertas épocas del año. Incluye dentro de este grupo algunas especies de Daphnia y Ceriodaphnia así como Bosmina longirostris. Esta última se encuentra en el plancton de lagos y embalses, así como en charcas y estanques, siendo común en lagos de tipo eutrófico (Hutchinson, 1967). Ha sido la especie de aparición más frecuente en los embalses españoles aunque no de forma constante ni muy abundante (Armengol, 1978a). Distribución horizontal de las especies del género Bosmina: si se comparan las curvas semilogarítmicas que grafican la distribución temporal de B. huaronensis y B. longirostris, puede señalarse que: 1.a) en la estación A, ambas especies declinan hacia el verano y aumentan su número hacia el invierno, excepto al comenzar el tercer ciclo anual (julio 1979) en que no se registra B. longirostris; 1.b) ambas especies presentan 3 picos máximos, coincidiendo el primero y segundo de huaronensis con el segundo y tercero de longirostris; 1.c) estos picos máximos coincidentes son muy diferentes en número, representando el primero y el segundo de longirostris sólo el 9 y el 12.5%, respectivamente de los señalados para huaronensis; 1.d) el mínimo registro para ambas especies se observa durante 1981 con igual no. de org. 10-3 m-2 , siendo en marzo para longirostris y en mayo para huaronensis; 2.a) en la estación B, B. longirostris declina hacia el verano y comienzos del otoño en los 4 ciclos estudiados mientras que huaronensis lo hace sólo en el verano de 1980, disminuyendo su número hacia el verano de los otros tres años; 2.b) las 2 especies presentan 2 registros máximos del no. de org. 10-3 m-2 correspondiendo en tiempo el primero de huaronensis con el segundo de longirostris; 2.c) el pico máximo registrado para longirostris representa el 20% del señalado para huaronensis; 2.d) ambas especies poseen 2 picos mínimos, el segundo de los cuales coincide, en cada una de las especies, con sus respectivos registros mínimos señalados para la estación A; 3,a) en la estación C, las 2 especies muestran un comportamiento similar a lo largo de los 4 cielos (especialmente desde julio de 1977 hasta noviembre de 1978) disminuyendo, hacia el verano y aumentando sus respectivas poblaciones durante el invierno y comienzos de la primavera; 3.b) difieren en que B. huaronensis declina sólo en el verano de 1980 y a fines del ciclo muestreado (mayo 1981) mientras que longirostris declina además en el verano de 1979 y 1981 y en el invierno de 1979 y 1980; 3.c) B. huaronensis y B. longirostris presentan un registro máximo coincidente en julio de 1977, representando el de longirostris el 35% del señalado para huaronensis; 3.d) mientras el registro mínimo de longirostris ocurre en noviembre de 1977, en la curva de distribución temporal de huaronensis se observan 4 registros mínimos, uno en cada uno de los ciclos anuales; 4.a) en la estación D, la frecuencia de aparición de longirostris en los recuentos es mucho menor que la observada en huaronensis. Esta ultima especie declina en enero de 1978 y de 1980 y en mayo de 1981. B. longirostris declina en noviembre de 1977, marzo, mayo y noviembre de 1978; marzo, julio y noviembre de 1979; enero, marzo y julio de 1980; enero y mayo de 1981; 4.b) los registros máximos del no. de org. 10-3 m-2 de huaronensis se observan en julio de 1977, en coincidencia con uno de los mayores registros señalados para la misma especie en la estación C, y en septiembre de 1980. B. longirostris presenta un registro máximo en mayo de 1980, coincidiendo con uno de los dos registros máximos indicados para esta especie en la estación C; 4.c) mientras que huaronensis presenta un pico mínimo en mayo de 1978, al igual que uno de los registros mínimos observados en la estación B, longirostris lo presenta en marzo de 1981, en coincidencia con los mínimos de las estaciones A y B; 5.a) en la estación E, B. huaronensis declina sólo en septiembre de 1979 y enero de 1980 mientras que longirostris lo hace también en septiembre de 1978, enero, marzo y noviembre de 1979, marzo y julio de 1980, enero, marzo y mayo de 1981; 5.b) en noviembre de 1978 se registra un pico máximo en la curva de huaronensis mientras que longirostris presenta el máximo registro en mayo de 1980, coincidente con uno de los dos picos máximos señalados para las estaciones D y C; 5.c) el registro mínimo en los recuentos de huaronensis se observa en enero de 1978 al igual que uno de los registrados en la estación C y el de longirostris en julio de 1977. Distribución vertical de las especies del género Bosmina: si se comparan las curvas de distribución vertical de B. longirostris con las de huaronensis, se observa que: 1) ambas especies presentan curvas bastante semejantes en los meses de julio y septiembre de 1977, marzo, julio, septiembre y noviembre de 1978; 2) en noviembre de 1977, B. longirostris aparece sólo a los 10 m mientras que huaronensis lo hace desde la superficie hasta los 15 m, siendo más abundante a 7.5 m; 3) en enero de 1978, B. huaronensis está presente a lo largo de toda la columna de agua mientras que longirostris aparece desde la superficie hasta los 5 m, siendo muy abundante a 1 m del espejo de agua; 4) en mayo del mismo año, las 2 especies son más abundantes a 1 m aunque en cantidades proporcionalmente diferentes; 5) en marzo y mayo de 1979, B. huaronensis se halló desde los 0.05 m hasta los 20 m. B. longirostris, por el contrario, se encontró sólo a determinadas profundidades. Durante el muestreo de marzo, B. longirostris aparece recién a 7.5 m y es más abundante a 15 m mientras que huaronensis lo es a 7.5 m. En el mes de mayo ocurre que longirostris es más abundante en superficie y huaronensis lo es a los 10 m. Ceriodaphnia dubiaDistribución horizontal: en la estación A, C. dubia tiene un comportamiento más o menos similar durante todo el periodo de estudio. Disminuye su población hacia el verano, llegando a declinar durante la estación estival de los dos primeros ciclos (1978 y 1979) y aumentando hacia el invierno y primavera. Las curvas semilogarítmicas de distribución temporal indican que existen 2 registros máximos en los recuentos: en noviembre de 1977 con 820 org. 10-3 m-2 y en marzo de 1978 con 854. Presenta un registro mínimo en marzo de 1981 con 24 (Fig. 4,A). 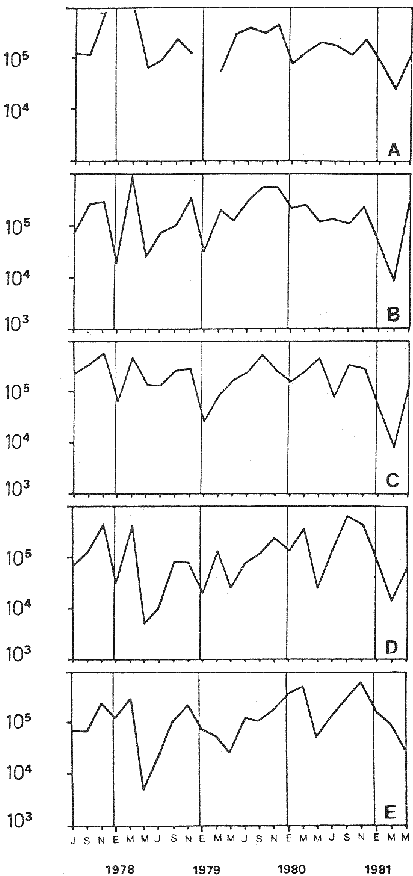 Figura 4. Distribución horizontal de Ceriodaphnia dubia. En la estación B, esta especie presenta una distribución temporal algo diferente. Si bien disminuye su población hacia el verano, como ocurre en la estación A, su aumento se observa inmediatamente antes y después de su disminución hacia finales de la primavera y en el otoño. Se señala un registro máximo en marzo de 1978 con 918 org. 10-3 m-2 , coincidente con el máximo registro observado en la estación A. El mínimo registro ocurre en marzo de 1981 con 8, también en coincidencia con el pico mínimo registrado en las curvas de la estación A (Fig. 4, B). En la estación C, dicha especie presenta un comportamiento similar al seguido en la estación B. C. dubia disminuye en enero de los 4 ciclos de muestreo y aumenta hacia la primavera y otoño. Se observan 2 picos máximos en las curvas: en noviembre de 1977 con 592,6 org.10-3 m-2, coincidente con el primer pico máximo señalado en la estación A y en septiembre de 1979 con 500. El registro mínimo se da, al igual que en las estaciones A y B, en marzo del 1981 con 8 org. 10-3 m-2 (Fig. 4, C). En la estación D, el comportamiento seguido por C. dubia es similar al observado en las estaciones B y C. La curva de distribución temporal se repite casi de igual manera en los 4 ciclos. Presenta un aumento marcado de la población hacia fines de la primavera, anterior a la disminución ocurrida en el verano. Luego del descenso estival, vuelve a aumentar el número de individuos abruptamente hacia comienzos del otoño para disminuir en forma rápida hacia fines de la misma estación. En uno de los ciclos estudiados (1978), esta disminución llega a ser tal que se observa en mayo un registro inferior a los señalados para el verano de los 4 ciclos. El registro máximo de C. dubia se observa en septiembre de 1980 con 680 org. 10-3 m-2. El registro mínimo, como se señala anteriormente, se observa en mayo de 1978 con 5 (Fig. 4, D). En la estación E, durante los 2 primeros ciclos y el último, esta especie sigue un comportamiento similar al indicado para la estación D. Se observa una disminución de la población hacia el verano y otra más marcada hacia finales del otoño. Por otro lado, se observa un aumento de la población hacia finales de la primavera, anterior al descenso estival. Por el contrario, en el tercer ciclo (julio 1979 julio 1980) no se observa disminución de la población durante el verano. Más aún, el número de individuos registrado en enero de 1980 es superior al indicado hacia fines de la primavera anterior. Se señalan 2 picos máximos: en marzo de 1980 con 510 org. 10-3 m-2 y en noviembre del mismo año con 596. El registro mínimo se observa en mayo de 1978 con 5, en coincidencia con el mínimo registro indicado en la estación D (Fig. 4, E). Desde julio de 1977 a mayo de 1981, C. dubia es más abundante en la estación C, luego en la B, A, D y E. Si se toma en cuenta el número promedio de organismos de las 5 estaciones, el mayor número se observa en marzo de 1978 con 586.5 org. 10-3 m-2-² y el menor en marzo de 1981 con 28. Distribución vertical: los gráficos indican que C. dubia no presenta el mismo comportamiento en los dos ciclos estudiados. No obstante, puede asimilarse la tendencia manifestada por dicha especie en los dos ciclos anuales a ser más abundante entre 1 y 5 m de profundidad durante el invierno y la primavera. No ocurre lo mismo hacia el verano ya que durante la primera estación estival, esta especie es más abundante a 20 m mientras que en la segunda abunda a 5 m. Hacia comienzos del primer otoño, partiendo de una escasa abundancia relativa en superficie, C. dubia alcanza su máxima abundancia a los 2 m y continúa a lo largo de toda la columna de agua con casi igual número de individuos. A principios del otoño siguiente, si bien esta especie abunda a los 10 m, presenta otro pico a los 2 m. Hacia fines del otoño, la distribución vertical de C. dubia se asemeja bastante en ambos ciclos. Es más abundante entre 1 y 2 m y sigue con una abundancia más o menos uniforme a lo largo de toda la columna de agua. Considerando los dos ciclos anuales completos, C. dubia es más abundante por lo general entre los 2 y 1 m de la superficie (Fig. 10). Si bien la ecología de C. dubia es poco conocida, se sabe que está adaptada a todo tipo de condiciones ambientales (Armengol, 1978a). Margalef (1953, fide Armengol, 1978a) señala que su presencia no está correlacionada con la existencia de vegetación acuática mientras que Olivier (1962) la encuentra preferentemente en lagunas con abundante vegetación. Diaphanosoma brachyurum Distribución horizontal: en la estación A, D. brachyurum se comporta de manera diferente durante los 4 ciclos estudiados. En los dos primeros años aparece sólo en la primavera, declinando hacia el verano, otoño e invierno. Por el contrario, se registra con mayor frecuencia en la segunda mitad del ciclo, declinando sólo hacia el otoño de 1980 y enero de 1981. Es en enero de 1980 cuando presenta el mayor registro con 76.5 org. 10-3 m-2. El registro mínimo se observa en marzo de 1981 con 4 (Fig. 5,A). En la estación B también presenta un comportamiento diferente en los 4 años investigados, siendo de aparición más frecuente en los recuentos correspondientes a los dos últimos años. Sin embargo, una característica similar observada en los 4 ciclos es la declinación de D. brachyurum durante el otoño. Asimismo, el segundo y tercer ciclos se asemejan también por la declinación de esta especie durante el invierno y comienzos de la primavera. No ocurre lo mismo durante el invierno y primavera del primer y cuarto ciclos. Por otro lado, declina durante el verano del segundo cielo (1978-1979). 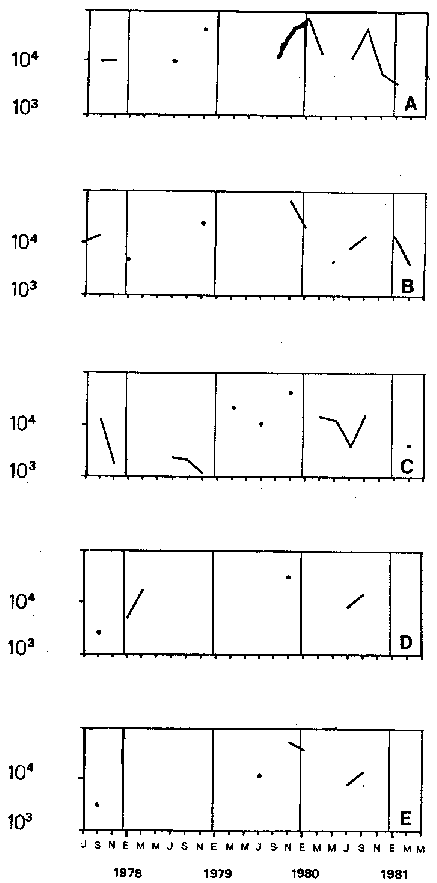 Figura 5. Distribución horizontal de Diaphanosoma brachyurum. El mayor registro se observa en noviembre de 1979 con 64 org. 10-3 m-2 poco antes del registro máximo señalado para la estación A. El mínimo registro se presenta en marzo del 1981 con 4, en coincidencia con el mínimo indicado en la estación A (Fig. 5, B). En la estación C, puede observarse que: a) D. brachyurum declina en enero y durante todo o parte del otoño de los 4 ciclos; b) durante el tercer ciclo (1979-1980) aumenta su población hacia la primavera, siendo en noviembre de 1979 cuando presenta el mayor número de individuos (41.2 org. 10-3 m-2), coincidiendo con el mayor registro observado en la estación B; c) el menor registro se señala en noviembre de 1978 con 1.2 org. 10-3 m-2; d) sólo en una oportunidad Gulio del primer ciclo) esta especie declina también en el invierno (Fig. 5, C). En la estación D, la aparición de esta especie es más esporádica, llegando a declinar a lo largo de todo el segundo ciclo (1978-1979). Durante el primer ciclo se registra a principios de la primavera y en el verano mientras que en el tercer y cuarto ciclos (1979-1980 y 1980-1981, respectivamente) aparece durante la primavera. El mayor registro del número de individuos se observa en noviembre de 1979 con 30 org. 10-3 m-2, al igual que en las estaciones B y C. El pico mínimo de la curva ocurre en septiembre de 1977 con 2.8 org. 10-3 m-2 (Fig. 5, D). En la estación E presenta una frecuencia de aparición similar a la indicada en la estación D, declinando al igual que en aquella, a lo largo de todo el segundo ciclo. Por otra parte, el comportamiento de esta especie durante el segundo ciclo es similar al observado en el primero y en el cuarto, ya que declina durante el verano y otoño. Por el contrario, en el tercer ciclo (1979-1980) aumenta su número hacia fines de primavera y comienzos del verano, presentando su máximo registro en noviembre de 1979 con 50 org. 10-3 m-2. Este registro coincide con los maximos señalados en las estaciones B, C y d. El menor registro se indica en septiembre de 1977 con 3.2 org. 10-3 m-2, en coincidencia con el mínimo registro de la estación D (Fig 5, E). Desde julio de 1977 a mayo de 1981, D. brachyurum es más abundante en la estación A. Siguen en orden decreciente, las estaciones B, C, E y D. Teniendo en cuenta el promedio del no. de org. 10-3 m-2 de las 5 estaciones, el número mayor se observa en noviembre de 1979 con 45.4 y el menor en noviembre de 1980 con 1.2. Distribución vertical: los gráficos de distribución vertical de la estación C señalan que D. brachyurum no presenta un comportamiento similar en los dos ciclos estudiados. Las diferencias existentes entre ambos ciclos son: 1) en el invierno del primer ciclo no se registra esta especie en toda la columna de agua mientras que en julio del segundo ciclo se registra a los 7.5 m y a los 15 m en igual cantidad; 2) mientras que en septiembre del primer ciclo es más abundante a 1 m de la superficie, en septiembre del segundo ciclo se registra sólo a 2 m; 3) en noviembre del primer ciclo aparece a 1 m de la columna de agua y abunda entre los 2 y 5 m, declinando a 7.5 m, en tanto que en el segundo ciclo aparece en superficie, abunda a 1 m y declina a los 7.5 m. La única característica común a ambos ciclos es la ausencia de esta especie durante los meses de enero y mayo (Fig. 11). Al igual que C. Dubia, D. brachyurum vive en todo tipo de ambientes acuáticos tanto pelágicos como heleoplanctónicos. En los embalses, su abundancia es bastante variable, dependiendo de las condiciones climáticas de la zona y de la coexistencia con otras especies con las que puede competir (Armengol, 1978a). Margalef (1953,fide Armengol, 1978a) destaca que en el plancton esta especie aparece en las capas superiores del epilimnion. Lewis (1979), en su análisis de la comunidad zooplanctónica de un lago tropical de las Islas Filipinas, menciona que Diaphanosoma es el único cladócero capaz de mantener abundancias relativamente elevadas a lo largo de todo el año, excepto durante el periodo de circulación y que es el dominante en términos de producción. Moina y Bosmina, en cambio, pasan por extensos períodos de extrema rareza aunque en ciertas épocas del año presentan picos de abundancia (Lewis, 1979). Daphnia laevisDistribución horizontal: en la estación A, esta especie presenta una frecuencia de aparición menor que las especies antes tratadas. En general, declina hacia el verano y otoño y aparece durante el invierno y primavera. Si bien no se comporta de igual manera en los 4 ciclos estudiados, pueden señalarse algunas semejanzas. D. laevis declina en septiembre del primer ciclo (1977-1978) y cuarto ciclo (1980-1981); tampoco se registra en los recuentos de enero, marzo y mayo del primero y segundo ciclos mientras que en el tercero y cuarto declina en enero y marzo pero aparece en mayo; además declina a fines de la primavera del segundo y cuarto ciclos. El pico máximo de la curva se registra en noviembre de 1979 con 150 org. 10-3 m-2 mientras que el mínimo se da en julio de 1977 con 0.6 (Fig. 6, A). 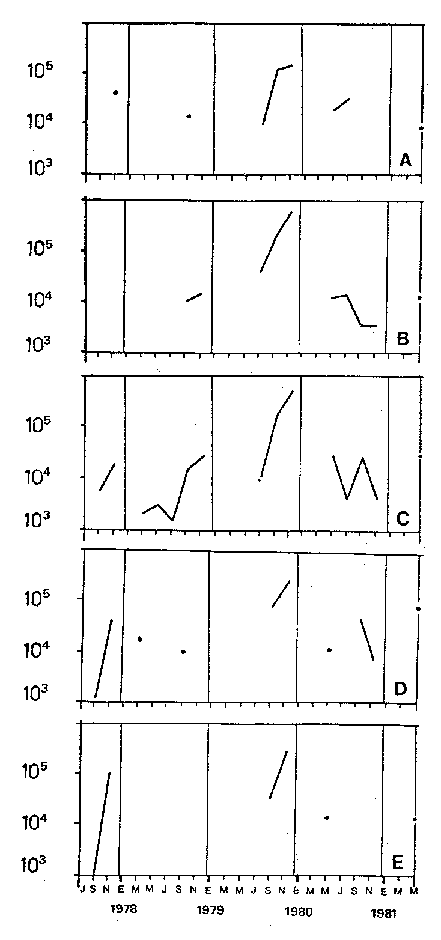 Figura 6. Distribución horizontal de Daphnia laevis. En la estación B, repite un comportamiento similar durante todo el período de muestreo, declinando hacia el verano, excepto durante el primer ciclo en que lo hace desde julio de 1977 a julio de 1978. El mayor registro se observa en noviembre de 1979 con 616 org. 10-3 m-2, en coincidencia con el mayor registro observado en la estación A. Los menores registros se señalan en septiembre de 1980 con 3.6 org. 10-3 m-2 y en noviembre de 1980 con 3.8 (Fig. 6, B). En la estación C tiene mayor frecuencia de aparición que en las otras estaciones. Por lo general, declina hacia el verano. En una sola oportunidad (julio 1977) declina también en el invierno. La población va en aumento hacia la primavera, siendo en noviembre de 1979 cuando se indica el mayor registro con 478.5 org. 10-3 m-2, coincidiendo con los registros máximos de las estaciones A y B. En julio de 1978 se señala el pico mínimo de la curva temporal con 1.5 org. 10-3 m-2 (Fig. 6, C). En las estaciones D y E, D. laevis aparece menos frecuentemente que en las estaciones A, B y C. En la estación D, se registra en los recuentos de septiembre y noviembre de cada uno de los ciclos, salvo durante el segundo ciclo (1978-1979) en que tampoco aparece en noviembre. Además de estos recuentos de primavera, sólo se ha registrado en marzo del primer ciclo y mayo del tercer y cuarto ciclos. El máximo registro se señala en noviembre de 1979 con 280 org. 10-3 m-2 y el mínimo en septiembre de 1977 con 1.3 (Fig. 6, D). En la estación E declina durante todo el segundo y cuarto ciclos y durante el verano del primer y tercer ciclos, apareciendo sólo en la primavera del primer y tercer ciclos y además en mayo del tercer y cuarto ciclos. El registro máximo, coincidiendo con las anteriores estaciones, se observa en noviembre de 1979 con 280 org. 10-3 m-2 y el mínimo en septiembre de 1977 con 1 (Fig. 6, E). Tanto en época como el no. de org. 10-3 m-2 de los registros máximo y mínimo son iguales para las estaciones D y E. D. laevis es más abundante en la estación B, luego en la C, D, E y A. Teniendo en cuenta el número de org. 10-3 m-2 promedio de las 5 estaciones, el mayor número se observa en noviembre de 1979 con 360.9 y el menor en julio de 1977 con 0.1. Daphnia ambiguaDistribución horizontal: en la estación A esta especie aparece sólo en la primavera de los 4 ciclos estudiados, presentando su máximo registro en septiembre de 1979 con 138 org. 10-3 m-2. En la curva de distribución temporal se señalan 2 registros mínimos: uno en septiembre de 1977 con 12.1 org. 10-3 m-2 y otro en noviembre de 1980 con 12 (Fig. 7, A). En la estación B tiene un comportamiento similar al indicado para la estación A aunque tampoco se registra en la primavera de 1978. Presenta su máximo registro en septiembre de 1979 con 162 org. 10-3 m-2 al igual que en la A y dos mínimos registros en septiembre y noviembre de 1980 con 3.6 y 3.8 respectivamente (Fig. 7, B). En la estación C, al igual que en la B, declina durante el verano, otoño, invierno y en la primavera de 1978. El pico máximo de la curva de distribución temporal se señala también en septiembre de 1979 con 50 org. 10-3 m-2 y el mínimo en julio de 1980 con 3.8 org. 10-3 m-2 (Fig.7, C). En las estaciones D y E, D. ambigua tiene aún una menor frecuencia de aparición. Sólo se registra su presencia en los recuentos de primavera de 1977 y 1980 en ambas estaciones de muestreo. Se observa un registro máximo de 20 org. 10-3 m-2 en noviembre de 1977 en las dos estaciones. Los mínimos valores de la curva están señalados en noviembre de 1980 con 4 org. 10-3 m-2 en la D y en septiembre de 1977 con 3.2 en la E (Fig. 7, D y E). D. ambigua es más abundante en la estación B, luego en las estaciones A, C, D y E. Teniendo en cuenta el número promedio de org. 10-3 m-2 de las 5 estaciones, el máximo registro se señala en septiembre de 1979 con 70 y el mínimo en septiembre de 1980 con 3. 1. Distribución horizontal de las especies del género Daphnia: de acuerdo a las curvas de distribución temporal de D. laevis y D. ambigua se concluye que: 1) D. laevis tiene una mayor frecuencia de aparición en las 5 estaciones de muestreo; 2) esta especie aparece más frecuentemente en la estación C en tanto que ambigua lo hace en la B; 3) D. laevis presenta sus registros máximos de no. de org. 10-3 m-2 en noviembre en las 5 estaciones, mes en el cual no se registra la presencia de D. ambigua. Es probable que esto se deba a problemas de competencia entre especies del mismo género; 4) también con respecto a la abundancia, D. laevis supera a ambigua. Distribución vertical de las especies del género Daphnia: Se consideraron ambas especies juntas en un gráfico. Si se comparan los dos ciclos anuales a lo largo de los cuales se estudió su distribución vertical en la estación C, pueden observarse algunas semejanzas: a) en septiembre de ambos ciclos las dos especies son más abundantes a 2 m, aunque en el primer ciclo aparecen hasta los 15 m de la columna de agua y en el segundo sólo hasta los 7.5 m; b) ninguna de las dos especies fue registrada durante el mes de enero de ambos ciclos; c) ambas especies aparecen sólo a 15 m en el muestreo de marzo del primer y segundo ciclos. Sin embargo, también pueden apreciarse algunas diferencias, tales como: a) ninguna de las 2 especies se registra en los recuentos de distribución vertical correspondientes a julio de 1977 mientras que sí aparecen a 7.5 y a 20 m en julio de 1978; b) en noviembre del primer ciclo las especies abundan a 5 m y declinan hacia los 15 m, en tanto que en el segundo son más abundantes a 1 m y llegan hasta los 20 m; e) D. laevis y D. ambigua presentan 2 picos de abundancia a 5 y 10 m durante el muestreo de mayo del primer ciclo, siendo mayor el de 10 m. Por el contrario, no se registran en mayo del segundo ciclo. Ramírez y Machado (1982) citan un sólo cladócero zooplanctónico del género Daphnia para la represa La Fe, 33 km al oriente de Medellín (Colombia). 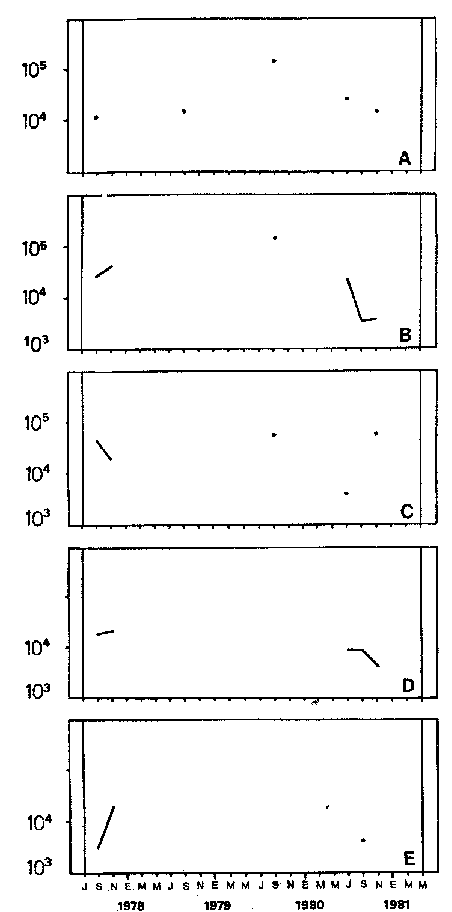 Figura. 7. Distribución horizontal de Daphnia ambigua. Otras especiesEn los datos de la columna integrada de agua de las 5 estaciones figura la aparición de Moina micrura en la estación B en el muestreo de marzo de 1979 con 20 org. 10-3 m-2 y de Alona pulchella var. cambouei (especie litoral y/o bentónica) en ocasión del muestreo de noviembre de 1979 en la estación E, también con 20 org. 10-3 m-2. M. micrura se registra también en la estación C, en las muestras de distribución vertical en enero y marzo de 1979. En enero esta especie aparece de manera discontinua a 1 m y a 15 m mientras que en marzo lo hace sólo en las muestras de superficie y de 2 m. Chydorus pubescens, otra especie litoral y/o bentónica, se registra en la mayoría de las estaciones durante los muestreos de julio, septiembre y noviembre de 1977 para volver a encontrarse sólo en septiembre de 1980 con 8 org. 10-3 m-2 en la estación E. Por otro lado, también se registra su presencia en las muestras de distribución vertical de la estación C en los 3 muestreos de 1977. En julio es más abundante a 5 m de profundidad, llegando hasta los 20 m, en tanto que en septiembre y noviembre abunda más hacia los 2 m y en superficie, alcanzando sólo una profundidad máxima de 5 m en septiembre y 15 m en noviembre. 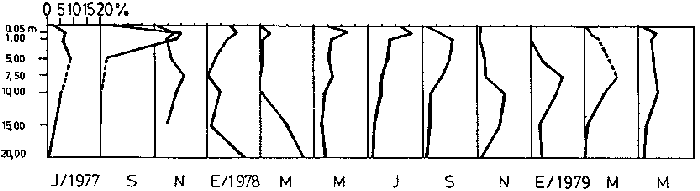 Figura 8. Distribución vertical de Bosmina huaronensis en la estación C. 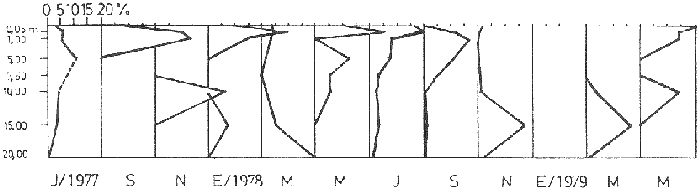 Figura 9. Distribución vertical de Bosmina longirostris en la estación C. 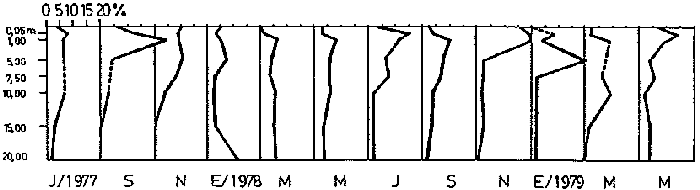 Figura 10. Distribución vertical de Ceridaphnia dubia en la estación C. 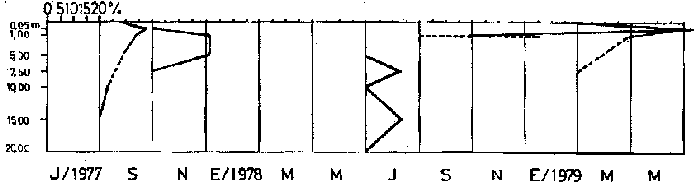 Figura 11. Distribución vertical de Diaphanosoma brachyurum en la estación C. 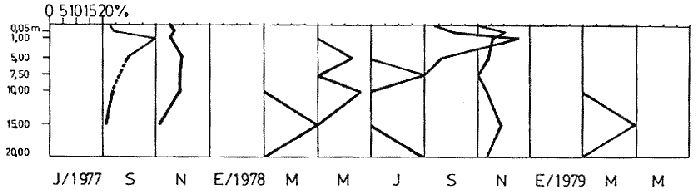 Figura 12. Distribución vertical de Diaphanosoma laevis y D. ambigua en la estación C. ConclusionesDel análisis de la distribución horizontal y vertical de las especies de cladóceros presentes en el embalse Río Tercero, así como de su variación estacional durante el período comprendido en la presente investigación pueden extraerse las siguientes conclusiones: 1) la especie más abundante en el embalse Río Tercero es Bosmina huaronensis y luego Ceriodaphnia dubia; 2) B. huaronensis y Diaphanosoma brachyurum son más abundantes en la estación A; Bosmina longirostris y C. dubia son en la estación C y Daphnia laevis y D. ambigua en la estación B; 3) La mayor parte de las especies tratadas se encuentran en menor número en la estación E; 4) C. dubia constituye la especie más frecuente, entre los cladóceros del embalse; 5) Por lo general, las especies del embalse declinan hacia el verano y aumentan sus poblaciones hacia la primavera. B. longirostris y C. dubia presentan altos registros también en el otoño; 6) en el caso de D. brachyurum, no suele estar limitada al verano, a diferencia de lo indicado por Margalef (1983); 7) en cuanto a la distribución vertical, B. huaronensis es más abundante entre 1 m y 7.5 m; B. longirostris a 1 m; C. Dubia y D. brachyurum entre 1 y 5 m; 8) tanto las dos especies de Bosmina como C. dubia se encuentran a mayor profundidad hacia el verano. AgradecimientosA Juan C. Paggi, la lectura crítica del manuscrito. LITERATURAARMENGOL, J., . Oecología aquatica, Los crustáceos del plancton de los embalses españoles 1978a. 3-96. 3: ARMENGOL, J., Verh. Intern. Verein. Limnol., Zooplankton crustaceans in Spanish reservoirs. 1978b. 1652-1656. 20: ARMENGOL, J. Oecología aquatica, Colonización de los embalses españoles por crustáceos planctónicos y evolución de la estructura de sus comunidades. 1980. 45-70. 4: BONETTO, A. A.; D. H. DI PERSIA, R. MAGLIANESI y M. C. CORIGLIANO, Ecosur, Caracteres limnológicos de algunos lagos eutróficos de embalse de la región central de Argentina. 1976. 47-120. 3 (5): BOSCI II, E. E. y M. L. FUSTER DE PLAZA, Publ. Dep. Inv. Pesq. Estudio biológico pesquero del pejerrey del embalse del Río Tercero (Basilichdys bonariensis). 1959. 61 p. 8: CORDINI, J. M., Minist. Agric. Public. Misc. Contribución al conocimiento limnológico del embalse del Río Tercero (Córdoba). Buenos Aires. 1949. 331 GUARRERA, S. A., Cs. Botánicas, El fitoplancton del Embalse San Roque (Provincia de Córdoba). Estudio preliminar. Rev. Inst. Nac. Invest. Cs. Nat. anexo al Ms. Arg. Cs. Nat. "Bernardino Rivadavia", 1948. 29-55. 1 (2): HUTCHINSON, G.E., A Treatise on Limnology. Part II: Introduction to lake biology and the limnoplankton. John Wiley and Sons Inc, New York. London, Sydney: 1967. 1116 p. LEWlS, W. M., Jr., Zooplankton Community Analysis. Studies on a Tropical System. Springer - Verlag. New York. Heidelberg. Berlin: 1979. 159 P. MARGALEF, R. Limnología. Ed. Omega: 1983. 1010 p. OLIVIER, S. R., Rev. Mus. La Plata (NS), Zoología. Los cladóceros argentinos. Con claves de las especies, notas biológicas y distribución geográfica. 1962. 173-269 7: PAGGI, J. C., Physis B, Contribución al conocimiento de la fauna de cladóceros dulceacuícolas argentinos. 1973. 105-114. 32 (84): PAGGI, J. C. Neotropica Aportes al conocimiento de la fauna argentina de cladóceros. I. Sobre Daphnia laevis Birge, 1878. 1977. 33-37. 23 (69): PAGGI, J. C., Acta Zool. Lilloana, Revisión de las especies argentinas del género Diaphanosoma Fischer (Crustacea Cladocera). 1978. 33 43-65. (1): PAGGI, J. C., Acta Zool. Lilloana, Revisión de las especies argentinas del género Bosmina Baird agrupadas en el subgénero Neobosmina Lieder (Crustacea: Cladocera). 1979. 137-162. 35 (1): RAMÍREZ, J. J., y T. MACHADO, Act. Biol. Univ. Antioquía Medellín - Colombia, Influencia de la precipitación y los ortofosfatos sobre el fitoplancton de la represa "La Fe". 1982. 321 11 (39):
|