|
POPULATION STRUCTURE OF THE DAB, Limanda limanda (LINNAEUS)
IN ISLE OF MAN WATERS, UK
Trabajo recibido el 4 de septiembre de 1985 y
aceptado para su publicación el 15 de febrero de 1988.
A. A. Ortega-Salas
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología.
Contribución 607 del Instituto de Ciencias del Mar y Limnología,
UNAM.
La estructura de la población del "dab",
Limanda limanda (L.) en las aguas de "Isle of Man" es
descrita en relación al peso y distribución de la frecuencia por
longitud, migración, mortalidad y una estimación de las
existencias. Los pesos individuales variaron de 1 g a 540 g y las longitudes de
30 mm a 360 mm. La longitud promedio de las hembras fue de 192 mm (skew: 0.6)
con un promedio de peso de 89 g; los machos alcanzaron una longitud promedio de
167 mm (skew: -0.7) con un peso promedio de 51 g. Hubo menos machos (40%) que
hembras (60%) en todas las edades y todas las capturas. Hubo una
inmigración a aguas someras (15-30 m) de mayo a septiembre, las hembras
migraron primero que los machos. La mortalidad total (z = M + F +
Migración) en dabs fue de 10% más alta en machos (z = 1.39) que
en hembras (z = 1.04). Varios métodos para estimar las existencias
fueron analizados, y una estimación final se obtuvo por el
análisis de la población virtual (VPA) dando aproximadamente
20,000 toneladas de "dabs" en el Mar de Irlanda en 1977.
The population structure of the dab, Limanda
limanda (L) in Isle of Man waters is described with reference to
weight and length frequency distribution, migration, mortality and stock
estimate. Individual weights varied from 1 g to 540 g and lengths from 30 mm to
360 mm. The average length of females was 192 mm (skew: 0.6) with an average
weight of 89 g; males average length being 167 mm (skew: -0.7) with an average
weight of 51 g. There were fewer males (40%) than females (60%) at all ages and
in all trawl catches. An immigration to shallow waters (15-30 m) occurred from
May to September, earlier in females then in males. Total mortality (z = M + F
+ migration) in dab was 10% higher in males (z = 139) than in females (z =
1.04). Several ways to get an stock estimate were analyzed, and a final stock
estimate by VPA gave approximately 20,000 tonnes of dab in the Irish Sea in
1977.
The dab is common in European and Icelandic waters and is used for human consumption. In the North Sea, dab is one of the six most important flatfish for consumption. It is also the most abundant flatfish on sandy banks 20 - 40 m depth (Bohl, 1959) and is very abundant around Iceland (Jonsson, 1966) and in the Baltic Sea (Pulsen, 1973). The European catch in 1977 was 13,998 tonnes (Nikolsaev, 1980), with annual catches in France of over 2,000 t and over 4,000 t in Netherlands and Denmark. In the summer dabs migrate inshore to reach the richer feeding grounds. An offshore migration follows as the temperature falls (Wheeler, 1969) so that in winter the fish seeks deep water but move shorewards in late winter for spawning (Jonsson, 1966). In the coastal area the average hauls per station are less productive in November and May than in August and September (Bohl, 1959). Total mortality of the North Sea Dab was calculated by Lee (1972) as z = 1.15 and natural mortality (M) as 0.3 for females and 0.5 for males. This study deals with the weight and length frequency distribution, migration, mortality and stock estimate of the population sampled. MATERIAL AND METHODSDabs were caught around the coast of the Isle of Man between 54° 00' and 54° 25' N, 4° 55' and 4° 10' W in depths from 4 - 40 in. A few additional samples were taken 20 miles E of the Isle of Man at a depth of 20 - 40 in and 3 miles SE of the Island between 30 - 50 in. Monthly samples were taken, weather permitting, from November 1976 to March 1979 by means of an otter trawl with wings of 70 mm mash size and cod end of 45 mm. Weskly samples of young fish were taken in Port Erin Bay at depth of 4 - 8 in whenever possible from April 1977 to May 1978, using a beam trawl with mesh size of 60 mm, exterior net, and 15 nun interior lining net. Records of each fish were maintained as a computer file. The results were processed in the ICL 1906A computer from the University of Liverpool and the CDC 7600 from the University of Manchester using the Statistical Package for the Social Sciences version 5 and 6, respectively. RESULTS
LENGTH AND WEIGHT FREQUENCY DISTRIBUTIONA total of 4,343 dabs sampled around the Isle of Man were examined. They are represented in figure I which shows their length and weight frequency distribution, 2,629 female dabs gave a mean weight of 88.8 g and a mean length of 192 mm with a skew of -0.7 which even though slightly negative could also be considered as a normal distribution. Similar features of weight and length frequency distribution are shown in figure 8 when male and female dabs were added together from otter, and beam trawl catches. Individual weights for females varied from 1 g to 540 g and lengths from 30 mm to 360 mm. Weight in males varied from 1 g to 200 g and lengths from 30 mm to 270 mm. The length and weight frequency distribution of male and female dab at each age-gp in years is shown in figure 2. MIGRATIONFish species normally occupy a particular environment mainly because of the availability of food but they may undertake migrations as conditions change throughout the year. These can be for reproductive purposes, for feeding or to avoid harsh environmental conditions in winter. Off the Cumberland coast, in the NE Irish Sea, Jones (1952) suggested that plaice and dab competed for food on the fine sand grounds. On the muddy sand, however, competition was less. In the same area but off Conway, Brander (1980) found that dab and palice (under 25 cm long) prefered to inhabit muddy sand and depth of not more than 40 m; also that dab were more numerous in March, although seasonal differences were not significant. Moller (1978) stated that in the southern North Sea there were few plaice in areas of sandy bottom were large number of dab were usually found. In the Isle of Man waters the same situation occurs. Furthermore Ortega Salas (1980) showed seasonal changes in the otolith accretion, in weight of gonads, liver, digestive tract, and also in the mean fish gutted weight and in the mean length of the dab throughout the year. All this suggest that there are seasonal variations of the behaviour of the dab, therefore if this is so, this indicates seasonal movements from one place to another. 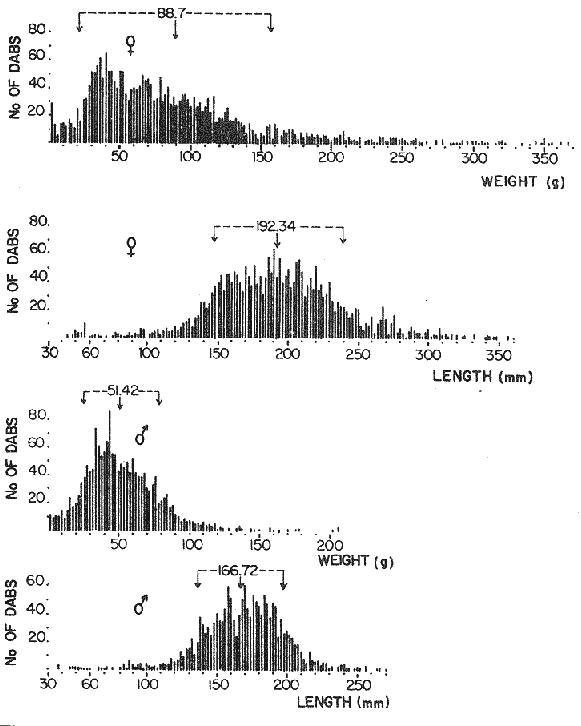 Figure 1. Weight and length of dabs caught in Isle of Man waters from November 1976-March 1979. Number of dabs at each 2 g weight intervals and at each 2 mm length intervals. Mean and standard deviation shown above each histogram. 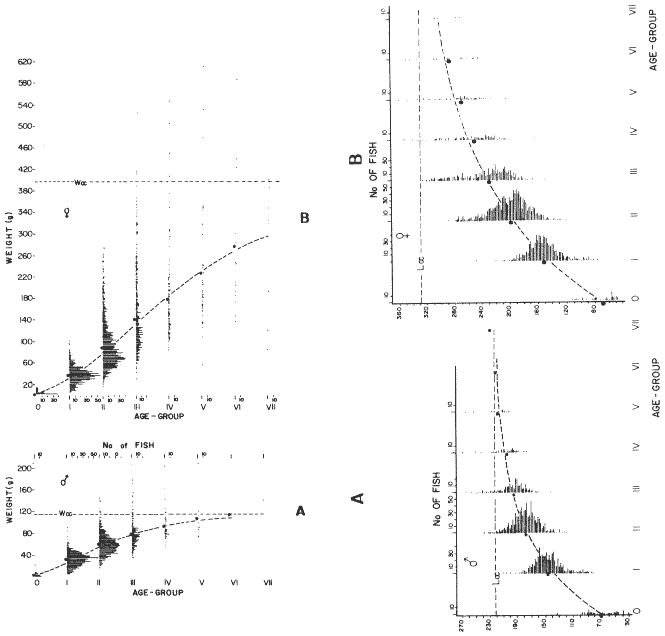 Figure 2. Length and weight frequency distribution at each age-group in years. Number of dabs at each 2 mm length intervals and at each 2 g weight intervals. Mean at each group represented by a dot and the theoretical von Bertalanffy's curve in dashed lines. 1-00 = asymptotic length; Woo = asymptotic weight. A = males; B = females. It is suggested here that the skew analysis could be use to detect emigration of dab from the sampled area. The length frequency distribution (Fig. 1) as was mentioned above gave a normal distribution for the whole population of male and female dab but the distribution of length at age showed a rather peculiar skew (Fig. 2). The length and weight frequency distribution of the fish at age in figure 2 shows that the length distribution of dab of agegp 1 is near normal with a slightly negative skew of -0.47 in females, and -0.44 in males. At age-gp the length frequency disstribution is also near normal, females 0.47 and males 0.01 but the length distribution of older dab showed a definitely positives skew (Table 1). The negative skew could be a result of mesh selection but the positive skew which represent anomalies in growth, could suggest two situations either fish have died or more of the larger than of the smaller fish have moved away falled to return. D. Harding's findings (personal communication) suggested that most of the dab moved to the Liverpool area to spawn around February and March (Fig. 3). 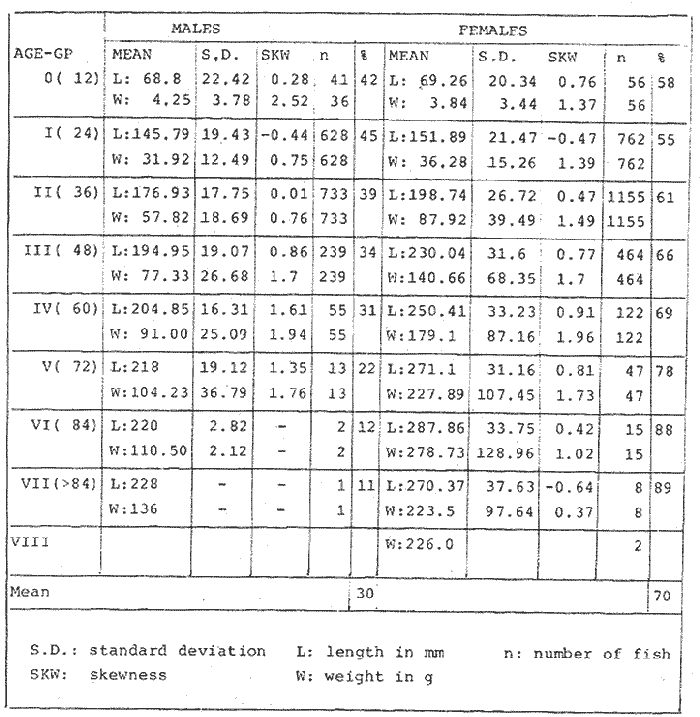 TABLE 1 SEVERAL STATISTICS AT EACH AGE-GROUP. Also the analysis of the population by catch per unit effort of the otter trawl around the Isle of Man showed that from month to month (Fig. 4) the catch was usually highest in June-August (summer) which coincides with high sea temperature and greater feeding activity (Ortega-Salas, 1979, 1980, 1981a, 1981c). Lower catches were in October/November to February/March when the temperature decreased. In Port Erin Bay the beam trawl catchs showed higher numbers of 0-gp dab from June and lower numbers from December to May (Fig. 5). This patter of catches throughout the year suggests that there is a seasonal movement of the dab into and out of the area sampled. A preponderance of females in the catches was also registered with a mean ratio of 1.5 (60%) females to 1 (40%) males (Fig. 6). Figure 7 shows that the variation in catches also applied to each age-gp, the oscillations in the graph will be due to natural sampling variability in trawl hauls or perhaps shows that dab move into and out of the area rather than remain as a resident population throughout the year. Recruitment offshore occurs at lengths and weights greater than 120 mm and 20 g respectively. There is a marked trend with age: the numbers tend to increase up to 2 year old and after 3 years decrease. The area sampled by trawl is not the total area of the distribution of the stock. It may be that dab younger than 2 years old were incompletely sampled either as result of mesh selection or because they had not been recruited into the area sampled. Figure 8 shows that the smaller and younger dab (0-gp) Were mostly caught in Port Erin Bay (nursery area), bigger and older fish were caught off shore. 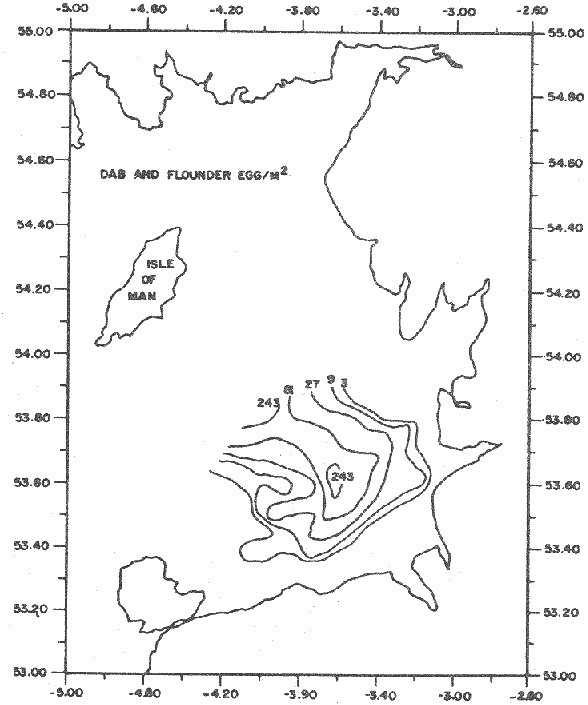 Figure 3. Dab and Flounder egg/m² sampled in NE Irish Sea march 1965. Data were kindly made available by D. Harding. 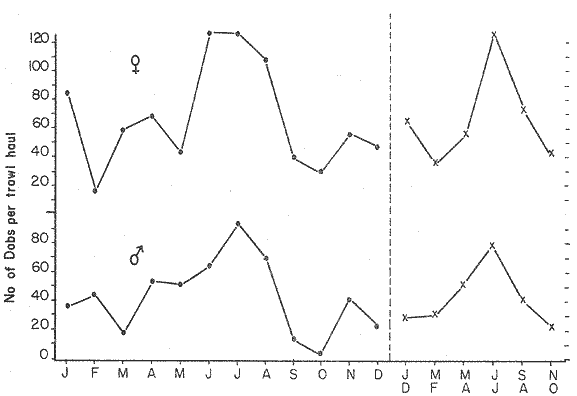 Figure 4. Number of dabs caught by otter trawl at each month of the year from November 1976 - March 1979. 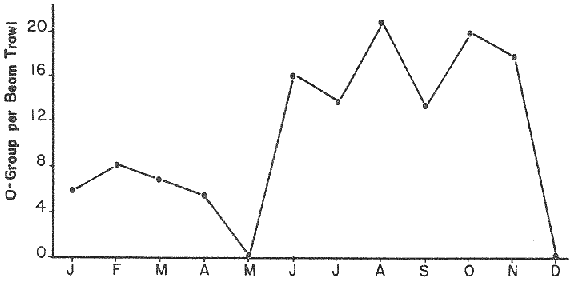 Figure 5. Number of 0-gp dabs per beam trawl caught in Port Erin Bay, June 1977 January 1979. 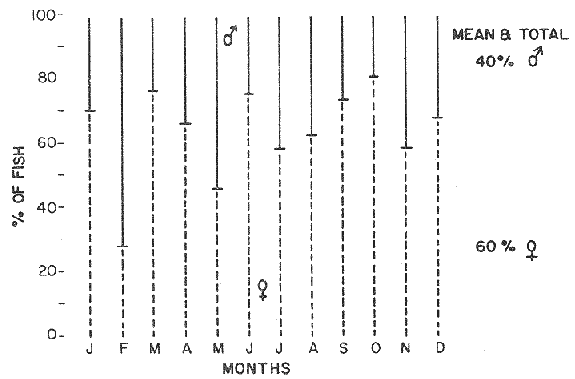 Figure 6. Percentage of male and female dabs caught monthly, November 1976 - March 1979. 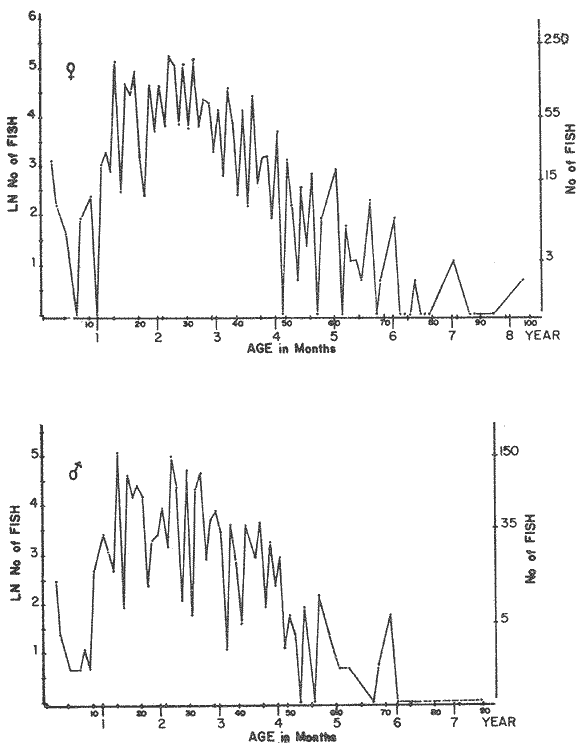 Figure 7. Number of fish for age at monthly intervals, A= females; B = males. In the North Sea for the female dab, Lee (1972) found a difference in the rate of decrease in growth (K) in the Central North Sea respect to west of Esbjerg and west of Helgoland as 0.22 and 0.46, respectively. And in the southern North Sea respect to Dutch coast and ET Buoys as 0.18 and 0.37, respectively. A similar situation has been suggested for plaice (Beverton and Holt, 1957) that there may be a differential migration of larger fish offshore. So, we may say that migration of fish from the area sampled will affect the calculation of total mortality if some of the fish tend to emigrate and fail to return. MORTALITYTotal mortality rate is one of the basic concepts of the fish population dynamics which enables an estimation to be made of the interrelation of the effects of natural morality (M), fishing mortality (F) and possible emigration of the species exploited. In this case the "catch curve" (Ricker, 1975) was used to determine total mortality (z) from the period sampled, assuming the recruitment is constant. Its analysis shows the number of dabs at each age-gp for both sexes in Table 1. The highest number was at age-gp 2 (11) with 1,155 females and 733 males. At age-gp 8 the numbers decrease to 2 females and t age-gp 7 to 1 male. The number at each age-gp were used to construct a catch curve which is shown in figure 9 as the number of fish transformed to natural logs (LN) against age in years. The ascending left limb and dome of the catch curve represents age-gps which were incompletely captured and were taken less frequently in relation to their abundance than the fish older. It would appear that dabs were fully recruited at two years old. 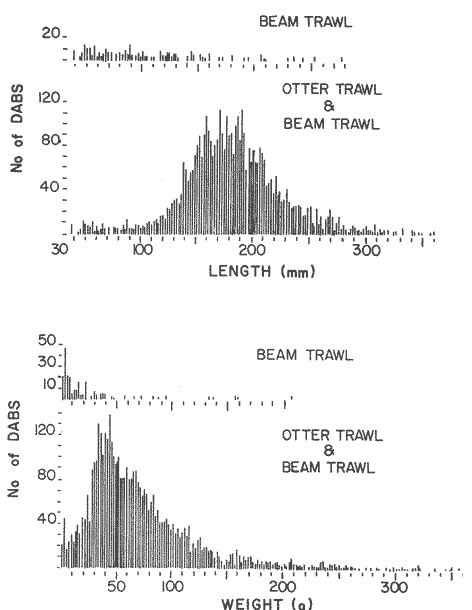 Figure 8. Length and weight frequency distribution of dab caught in Isle of Man waters, November 1976 March 1979. The slope of the straight right hand portion of the curve was -1.39 for males and -1.04 for females. This represents in total mortality (z) a loss per year from all causes of 75% for males and 65% for females. Thus the calculated value of z was from age-gp 2 onwards by Ricker's (1975) method as follows: Instantaneous total mortality rate z = F + M + migration: Total mortality rate: A = i - e-z; Survival rate S = e-z; where: F: fishing mortality; M: natural mortality; e: base of natural logs. The regression line gave an slope (-) which was z (+): Females: n = 7, r = -0.997, b = - 1.04, a = 9.195, z = 1.04, S = 35%/year, A = 65%/year; Males: n = 7, r = -0.995, b = -1.392, a = 8.091, z = 1.392, S = 25%/year, A =75%/year There is little information on the contribution of natural and fishing mortality coefficients towards z. However an approximation to M might be obtained as a comparative basis. The ratio of M/K for pleuronectids is close to unity (Beverton and Holt, 1959) which on this basis (OrtegaSalas, 1981a, 1981b) K was estimated for males K = 0.6 and for females K = 03. Then F (+ migration) automatically becomes 0.79 and 0.74 respectively. Fishing mortality depends on the gear employed and only operates on fish above certain sizes. In figure 9 the extremes on the left hand size of the histograms shows the effect of the beam trawl which was adapted to operate with smaller mesh size in Port Erin Bay. 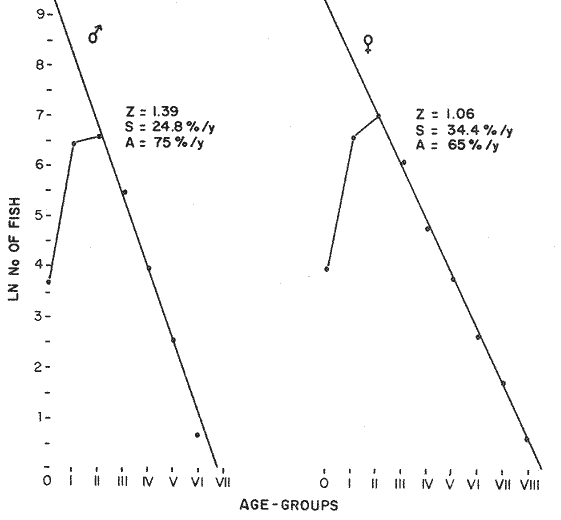 Figure 9. Catch curve. Ln = number of fish at each age - group. Calculated regression line fitted. STOCK ESTIMATEIn order to be able to assess any fishery, determination of the stock size is essential to be able to examine the evidence for either natural changes in the stock and/or changes by fishing effort. It is believed that the fishery on dabs might be much more important for future exploitation. The estimation of the actual catch of dab is essential in order to get a good estimate of the stock figure, but the dab is a fish which is not specifically trawled for because its commercial demand is lower than other flatfish like plaice and sole. Discards of the smaller dab must be very high, as it is taken incidentally, when trawlers are fishing for other species. The reported landings are very different from the total catches. One possible method for stock determination on dab is to relate its number and/or weight to landings of plaice or sole which are well documented. Then the ratio of numbers and/or weight per fishing effort of dab to plaice or sole would be a useful clue to reestimate the total catches. Using the reported landings and the estimate of rejection from England and Wales (MAFF-Lowestoft laboratory statistics) and raising them to the total official catches reported in "Bulletine Startistique" gives a better estimate of the total catch of dab. Van Leeuwen (1975-1978) has reported the results of ground fish surveys carried out in the east of the ground fish surveys carried out in the east of the Irish Sea using a beam trawl with 45 mm size. He reported the average length distribution in numbers per 100 h fishing of dab, plaice and sole caught in the surveys. Then the average of weightlength relationship between males and females of plaice and sole taken from ICES report (1976) for the Irish Sea and dab gave the following results (Table 2) using the equation: PLAICE: Wg=0.017117758 Lcm2.1538; SOLE: Wg= (0.00284412 Lcm3.3553) x 1.25; DAB Wg = -0.00000503 Lcm3.1632. These functional equations were applied to those data of Van Leeuwen as follows: If at length of 20.5 cm were found 3245 fish then 205 mm3.1632 x 0.00000503=103.3 g hence 103.3 g x 3245 = 335,208.5 g. Then the sum of all calculated weights for each length is shown in Table 2. In two years the dab biomass per 100 h fishing is greater than for the other two species, and this biomass was found to be the same. For the four year period dab was more abundant in numbers than plaice and sole. If we plot weight against numbers, there are approximately 15 thousand dab per 1 tonne but plaice or sole will give 5 thousand fish per 1 tonne. Quarterly surveys covering fishing grounds in NE of the Isle of Man, in Liverpool Bay and off Conway have been carried out by the fisheries laboratory since 1975 using a mean mesh size trawl of 79 mm. Survey data were kindly made available by K. Brander and F. Wallace. The mean ratio of dab; plaice was between 2 or 3 to 1 but for hauls more than 12 m from the coast of the Isle of Man in depth of 10-20 fm (which is an usual depth for commercial fisheries) the ratio was 5.6:1 (Table 3) which is roughly half that found by Lee (1972) (13: 1 dab: plaice) in the middle and south of the North Sea. Some data from the "Silver Star" survey carried out in June of 1980 off the east coast of the Isle of Man and Conway were used to calculate an estimate of weight at length using the weight-length relationship mentioned above of the length frequency distribution Table 4 and also an intent to relate the official commercial landings for plaice (which is over 24 cm) and or dabs (which is 15 cm and over). Since we do not know what the landed length disstribution is, it is assumed that the lowest commercially attractive size of dab is 20 cm. Smaller fish are considered to be discarded. 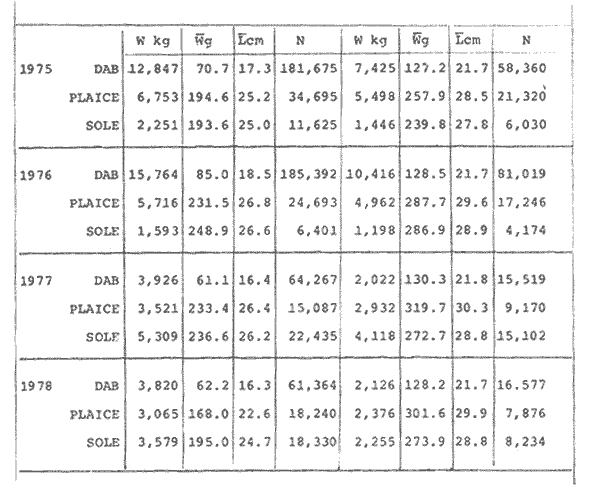 TABLE 2 SUM OF CALCULATED WEIGHT PER LENGTH DISIRIBUTION USING van LEEUWEN'S DATA (1975-1978). Wkg = weight in kg; Wg = mean weight in g; Lcm = mean length in cm; N = number of fish. commercial size = dab 19 cm; plaice and soIe = 24 cm. 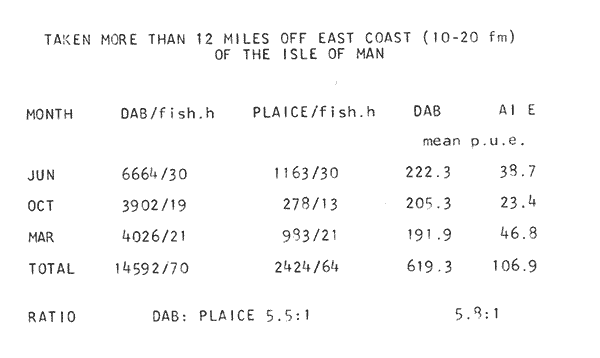 TABLE 3 RATIO DAB:PLAICE CATCHES. Thus, Van Leeuwen (1975-1978) data showed that the biomass of dab was greater than the total of plaice, or that of sole in 1975 and 1976; whereas in 1977 and 1978 it was similar. Also at the east of the Irish Sea K, Brander and P Wallace's data from a commercial vessel showed that dab was more abundant than plaice (2 to 3 : 1) and the frequency distribution by length showed a similar mean weight of landing size plaice to dab (Table 4). So the reported catches for plaice should not be very different than those of dabs. We might use the catch figures of plaice reported in 1977 with more confidence to get an idea of the stock of dab. 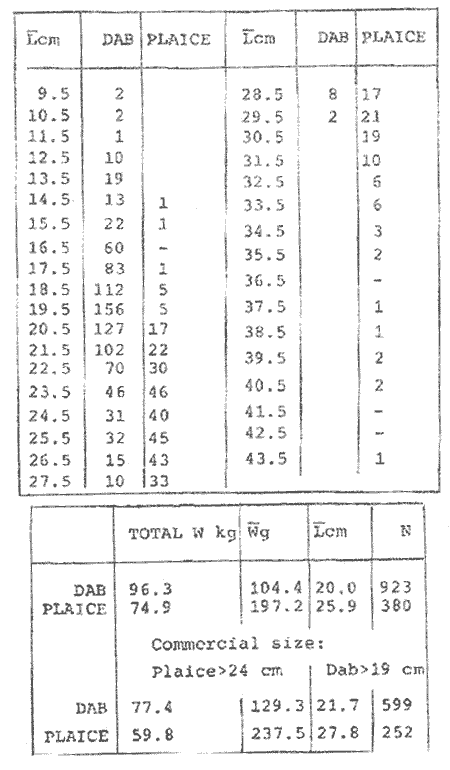 TABLE 4 NUMBER OF DAB AND PLAICE AT LENGTH AND THE CALCULATED WEIGHT. DATA WERE KINDLY MADE AVAILABLE BY K. BRANDER AND P. WALLACE. Eventhough we have good reasons to use plaice catches to estimate the stock of dabs, we should try another way in order to compare the estimate. Estimated figures of dabs caught by England and Wales with vessels of over 40 ft in the Irish Sea (Estimated statistics in Lowestoft) showed a figure of 524.68 t from which 471.51 were rejected and 53.17 were landed which means that approximately 90% were discarded from the catch. So in order to estimate the actual catch we may consider the following model: R´= Rejection/Estimated catch where: R'= proportional rejection; Est. catch = Rejection + Landings; then Landings = Est. catch (Est. catch x R'); Landings = Est. catch/(1-R'); Landings/(1-R') = Est. catch; R'= 0.89866; (1-R') 0.101338. International landings of dabs in the Irish Sea in 1977 = 327 t .•. Est. Intern. catch = 327/0.101338 = 3226 t. This figure is not very different from the plaice catch reported in "Bulletine Statistique" in 1977 (2.904 t) considering that most of this catch belonged to commercial size of plaice. Then if we split 3226 t into 27% for males and 73% for females which is the percentage weight as shown later (assumed to be my catch curve representative in the Irish Sea) and applying the formula: Pn= cxz/FxA where: Pn= population in numbers: c: estimated catch in t: z: total mortality coefficient; F. fishing mortality coefficient; A: 1 - e-z (actual total mortality rate). II and over + Less than II: Pn = 33,496,126.0; Pn = 49,060,439.7. TOTAL males and females Pn = 82.556.565.7. 40%: 33,022,626.3 x 51.28 g(W) = 1693.4 t 60% 49,533,939.4 x 88.81 g(W) = 4399.1 t TOTAL males and females = 6,092.5 t in the Irish Sea in 1977. It is known that commercial vessels fish on prawns and shrimps in the Irish Sea; they use smaller mesh size than 77 mm at the cod end. So discards of 0-gp flatfish should be enormous in those fisheries. Meyer-Waarden and Tiews (1965) reported that the German Shrimp fishery caught an average of 47 million young dabs per year in the North Sea. This kind of situation could introduce error in the calculated figure. Neverheless if fishing (F) is considered the same for all age-gps we may apply the following calculation: MALES: Pn = 17,185,266.6 x 1.39/0.79 x 0.75 = 40.316.490.4 FEMALES: Pn = 26,390,225.4 x 1.04/0.74 x 0.646 = 57,413.259.2 TOTAL males and females: 97,729,749.6 dabs 40%: 40,316,490.4 x 51.28 g = 2,067.4 t 60%: 57,413,259.2 x 88.81 g = 5,098.9 t TOTAL males and females = 7,166.3t in the Irish Sea in 1977. The estimated stock of dabs varied between 6 and 7 thousand t with a population of 82 to 98 million dabs as a minimum stock. This last figure in t compared with plaice of 7.143 t and sole of 6,905 t from virtual population analysis (VPA) in 1977 (ICES report, 1979) is approximately the same but if we apply the VPA formula (Gulland, 1965) to the catch number at age the results are showed in Table 5. In formula 1 and 2 M is fixed and F is also fixed for all or most of the ages. Then these coefficients were applied to the catch figure to get the stock. So the stock figure depend on fishing effort and mesh selection. On the other hand, using VPA by Gulland's method it also calculates the stock from catches but it is independent of fishing effort and mesh selectivity. It gives fishing mortality at age and a better estimate of the stock figure. 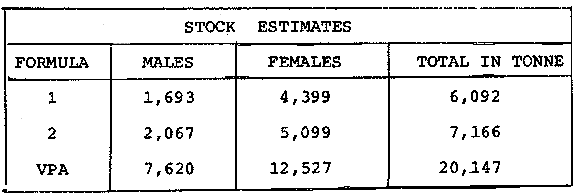 TABLE 5 STOCK ESTIMATES. This last figure might be high since z was high, probably because migration was involve but on the other hand we have used a M constant for all age-gps and it is probably that the age-gps 0 and 1 have higher M. DISCUSSIONThere was a marked seasonal variation in catches of dabs at all ages in the area sampled with a maximum in summer (June-August) and a minimum in October and February. This suggests that the area sampled was a feeding ground from which dabs moved away, probably, to deeper waters for overwintering and spawning. Most of the dabs caught in the sampling area around the Isle of Man varied between 11/2 and 4 years old. The 0-gp fish sampled were mostly from Port Erin Bay between June 1987 to January 1979. It was found that their number decreased from December to May. The 1-gp dab were scarce in the Bay and it is though that they had started to move into the fishing grounds. Edwards and Steel (1968) found a similar situation from Loch Ewe on the west of Scotland in June. The 0-gp lived just sublittoral in depths randing from low water neaps to 8 in with a maximum abundance at 4 m until September, when most of them moved out of the Bay. Mean weight and mean lengths at age varied throughout the year which in summer are higher. This is reflected by a rapid growth. In winter dabs decrease in weight and length (Ortega-Salas, 1980, 1981a, 1981b). Since it is not conceivable that fish became shorter in winter one must postulate a movement away from the sampled area of larger fish or assumed that the larger fish avoided the net but it seems unlikely that net avoidance should follows such a regular pattern. There is some evidence that suggests emigration of larger fish in the period DecemberMay: a) Number of dabs per trawl decreased from September/October to April/May and also in 0-gp from December to May. b) In figure 2, the length frequency curve at each age-gp showed that for 1-gp dabs which were still immature the curve was almost normal but from age-gp 2 onwards when fish had matured the distribution presents a positive skew which could be interpreted that most of the larger fish have emigrated, probably, for overwintering and spawning. Although they also might have died. Bohl (1959) explained the variation in catches of dabs in the North Sea as follows: as winter approaches the fish move away from the rapidly cooling coastal waters towards the open sea. They spend winter at some distance from the coast and after spawning they return to the summer warmed coastal waters. Within an extended spawning time the older and larger dabs spawn earlier than the younger and smaller one. In addition males stay in the spawning places longer than females. The older and larger females are first to remigrate to the coast than the younger and smaller females, lastly the male will follow. The later the remigration begins the less far the fish can advance towards the coast. Poulsen (1933) considered that there was an immigration into the Limfjord from the North Sea so the population built up from April to maximum numbers in September and fall to a low values in winter: dabs did not spawn in the fjord. Lee (1972) also showed that in winter dabs could emigrate from around 30 to 50 miles. In Isle of Man water it is assumed that most of the dabs migrate out of the shallow waters, probably, to deeper water with a possibility that the bigger ones move out of the Irish Sea. Most of the larger dabs emigrate by October/December, some of them remain in shallow waters, mainly the smaller ones, and probably spawn around there. Bal (1940) found eggs in Isle of Man waters near the coast from February to August. The catch curve (Fig. 9) shows a mortality rate in the area sampled of 75% per year for males compared with 65% per year for females. This loss could be explained in terms of death and/or emigration. Bohl (1959) and Poulsen (1933) reported that males disappeared from catches at a much earlier age than females; since a variety of sites and depths were sampled, it would seem that this reflects a higher mortality rates for females rather than a different migration pattern. Lee (1972) reported for the North Sea dab similar findings and calculated M as 0.49 for males and 0.31 for females. In the North Sea plaice Bannister (1977) postulated higher natural mortality and higher fishing mortality in males than in females, and the data of Bowers and Lee (1971) can be used to derive a catch curve for plaice take in about the same area as my dabs that shows similar features to the catch curve for dabs. So higher mortality in males may not be unusual in flatfish. Differences in natural mortality between male and female dabs produced an initial recruitment for males of 58% respect of the females of 42% when the VPA model was applied. This percentage should had been more similar, eventhough the total figure calculated by this method is considered acceptable. The biomass of dabs respect of the one of plaice, and that of sole was very similar in the Irish Sea, although the reported catches do not show so. A similar situation it seems to occur for the ICES regions Lee (1972) showed that in all these regions the annual total international landings of common dabs from 1908-1967 varied from 8.000 to 12,000 tonnes which compared with that of plaice landings is roughly 1/10. He also calculated the dab biomass as 0.5 million tonnes which is not very different from that of plaice of 0.7 million tonnes reported (ICES, 1977) for the period 1967-1971. Even more Poulsen (1937) has shown that the stock of dabs and plaice in Icelandic waters were not very different although there were some variations from year to year. In 1924 plaice was more abundant than dabs but in 1931 dabs were more. These findings confirm our thoughts that although dabs are very abundant and are trawled by catch in the ICES regions they are discard in high proportion because plaice and sole have higher commercial demand. Thus, to summarize this findings we could say the following; 1. The sex ration was 40% and 60% males and females, respectively, which has been assumed to be the pattern of the Irish Sea. 2. The length frequency distribution showed to be normal distribution but the distribution at age showed a positive skew gradient from age-gp2 onwards which could be either fish have died or have emigrated from the sampled area. 3. The catches per trawl haul showed immigration to the sampled area in summer and emigration from this area in winter. 4. Even the total mortality figure calculated was based in the catch curve obtained for the period sampled, the results were similar to those of Lee's (1972) for dabs in the North Sea. Also males showed higher mortality than females. 5. The results obtained were assumed to give the pattern of population of dabs in the Irish Sea from where the stock estimate was based. This results showed that the stock of dabs in weight is very similar to that of plaice and possibly of that of sole even dabs stock seems to be higher. AgradecimientosAcknowledgments are due to A. Burd, C. Bannister and M.S.M. Siddeek for their advise. For Laboratory facilities to Dept. of Marine Biology of the University of Liverpool, and MAFF-Fisheries Laboratory, Lowestof. And for the finantial support, Universidad Nacional Autónoma de México; Consejo Nacional de Ciencia y Tecnología; and the British Council. LITERATURABAL, D.V., Proc. Trans. Liverpool Biol. Soc. Observations on spawning periods and key to pelagic eggs of fishes in manx waters. 1940. 108. 74: BANNISTER, R. C. A., Fish Population Dynamics. North Sea Plaice. Gulland. J.A. Ed Wiley, Lon. 1977. 372 p. BEVERTON, J. H. and S. J. HOLT,. Fish Inv. London Ser. II 19, On the dynamic of exploited fish population. 1957. 533 p BEVERTON, R. J. H., and S. J. HOLT, Ciba Fdn. Colloq. Ageiong., A review of the life span and mortality rates of fish in nature, and their relation to growth and other physiological characteristics. 1959. 142-177. 5: BOHL, H., Ber. Dtch. Wiss. Komm. Meer. Die bioilogie der kliesche (Limanda limanda (L.)) in der Nordsee. 1959. 1-57. 15 (1): BOWERS, A.B. and J.W. LEE, T. J. Const. Int. Mer. The growth of plaice in Laxey Bay (isle of Man). 1971. 43-50 34 (1): BRANDER, K. Protection of life in Sea. Holg. Meer. Fisheries management and conservation in the Irish Sea. Kinne, O. and H.P. Bulnhelm Eds. 1980. 599-657. 33 (1-4): EDWARDS, R. R. and STELL, J.H., J. Explor. Mer. Biol Ecol. The ecology of 0-gp plaice and common dabs at loch ewe. I. Population and food. 1968. 215-238. (2): GULLAND, J. A., E. Estimation of mortality rates annex to arcic fisheries working group report, (meeting in Hamburg, January 1965). Doc. 3 (mimeo) ICES, C.M. 1965. 1965. GULLAND, J. A., Fish Manual of Methods for Fish Stock Assessment. Part 1. Population Analysis. FAO Man. Fish Soc. 1969. 154 p 4: I.C.E.S., Report of the North Sea Flatfish Working Group. Int. Counc. Explor. Sea. C.M. 1976/F: 4. 1976. I.C.E.S., Review of Some Fish Resources within the NEAFC Convention Area.(mimeo). Int. Counc. Expl. Sea C.M. 1977/ 1977. 37 P. IF: 5 ICES Report of the Irish Sea and Bristol Channel Working Group. (mimeo.) Int. Counc. Explor. Sea. C.M. 1979/ 1979. 99 p. G: 23 JONES, N.R., J. Anim. Ecol., The Bottom Fauna and the Food of Flatfish off the Cumberland Coast. 1952. 182-205. 21: JONSSON, G., Rit. Fish. Contribution to the Biology of the Dab (Limanda lilmanda (L.)) in Icelandic Waters. 1966. 3-36. (3): LEE, C.K.C., . Thesis. The Biology and Population Dyamics of the Common Dab, Limanda limanda (L.) in the North Sea. Ph. D. Univ. East Anglia, U.K. 1972. 105 p. LEEUWEN, VAN, Const. Int. Expl. Mer. Ann. Biol. A pre-recruit survey in the Irish Sea on Demersal Species Carried out by the Dutch R.V. " 1977-1980. 33-35. "Willem Beaukelsz", 26 May - 26 June 1975 and 8-28 May 1976. And by the Dutch Beamer K.W. 34 "Rose Marie" 25 April 14 May 1977 and April/May 1978. MEYERS-WAARDEN, P.F. and R. TIEWS, Bert. dt. Wiss. Kommn Meeres. Forshc. Der Belfang in den Fangen der Deutschen Garnelenfischeri in der Jahren 1954-1960. 1965. 13-78. 18 (1): MOLLER, C. J. Fisheries of the British and Northern European Seas. Penguin Books Ltd. 1978. 128 p. NIKOLAEV, Buff Stat. Pech. Mar. ED. Conseil International Pour L'Exploration de la Mer 1980 60. ORTEGA-SALAS, A. A., Cyclic Phenomena in Marine Plants and Animals. , Seasonal changes in dab (Limanda limanda) in Isle of Man waters. (Abstract). Naylor, E. and R.G. Harnoll Ed. Pergamon Press 1979. 477 P. ORTEGA-SALAS, A. A. J. Fish. Biol. Seasonal changes in the common dab, Limanda limanda (L.) in Isle of Man waters 1980. 75-82. 16 ORTEGA-SALAS, A. A., Ph. D. Thesis. Biology of the dab, Limanda limanda (L) in Isle of Man waters. Univ. Liverpool. U.K, 1981a. 98 p. ORTEGA-SALAS, A. A., An. Inst Cienc. del Mary Limnol. Age and growth of the dab (Limanda limanda (L.)) otoliths in Isle of Man waters. Univ. Nal Autón. México, 1981b. 69-78 14 (1): . ORTEGA-SALAS, A.A., . An. Inst. Cienc. del Mar y Limmol. Age and Growth of the dab, Limanda limanda (L.) in Isle of Man waters. Univ. Nal. Autón. México, 1981c. 15 (1). 1988 ORTEGA-SALAS, A.A., An. Inst. Cienc. del Mar y Limnol. Diet. food consumption and growth of the dab Limanda limanda (L) in Isle of Man waters. Univ. Nal. Autón. México, 1981d 15 (1). 1988. POULSEN, E.M., Rep. Dam Biol. Sta. The dab in the danish waters. Investigations on stock and growth. 1937. XLII. RICKER, W.E., Bull Fish. Res. Bd. Can. Computation and interpretation of biological statistics of fish population. 1975. 382 p. 191, WHEELER, A., The Fishes of the British Isles and North-West Europe. Macmillan London 1969. 113 p.
|