|
CRECIMIENTO Y MORTALIDAD DE LAS FASES ESTUARINAS
DEL CAMARÓN ROSADO Penaeus (Farfantepenaeus) duorarum BURKENROAD, 1939
EN LA LAGUNA DE TÉRMINOS, CAMPECHE, MÉXICO
Trabajo recibido
el 23 de mayo de 1986 y aceptado para su publicación el 19 de agosto de
1986
Fernando Álvarez N.
Adolfo Gracía G.
Luis A. Soto
Instituto de Ciencias Del Mar y Limnología,
UNAM, Apartado postal 70-305. México, D. F. 04510 Contribución
526 del Instituto de Ciencias del Mar y Limnología,
UNAM
Se
estiman las tasas de crecimiento y mortalidad de las fases estuarinas
dePenaeus duorarum(postlarvas epibentónicas y
juveniles: 6.2 a 80 mm de L.T.) durante un ciclo anual en dos localidades de la
Laguna de Términos previamente reconocidas como áreas de
establecimiento del camarón. Las tasas de crecimiento fluctuaron desde
0.58 mm/día en octubre hasta 1.12 mm/dia en agosto para la localidad del
Cayo y desde 0.82 mm/día en febrero y marzo hasta 1.13 mm/día en
agosto para la localidad de Chacahito. Los registros más altos se
presentaron durante el periodo de lluvias; sin embargo, las tasas de
crecimiento no fueron significativamente diferentes (P>0.05)durante el
periodo de estudio. La densidad de organismos fue el factor principal
determinante de las variaciones en las tasas decrecimiento responsable del 52
al 96% de la varianza. Las tasas de mortalidad diaria presentaron variaciones
desde -0.1145 a -0.1539 en las dos localidades y tampoco mostraron diferencias
significativas (P>0.05) en los periodos climáticos considerados
(estío y de mayor precipitación pluvial). La emigración
introduce un sesgo promedio de 25% a las estimaciones de mortalidad. Se propone
un mecanismo mixto regulador del crecimiento de tipo ambiental densodependiente
en el cual interactúan los factores asociados a la precipitación
pluvial con un periodo de retraso sobre la densidad de organismos y ésta
afecta a su vez al crecimiento idividual.
Palabras
clave:Crecimiento, mortalidad, fases estuarinas de camarón
rosado, variaciones estacionales, mecanismos reguladores del
crecimiento.
The
annual growth and mortality rates of estuarine stages (6.2 to 80 mm of total
length) of Penaeus duorarum were estimated from two selected
localities in Terminos lagoon, previously recognized as settlement arcas.
Growth rates exhibited ample fluctuations throughout the year on both
localities. At El Cayo site, the calculated daily growth ranged from 0.58 mm in
October to 1.12 mm in August, whereas at Chacahito site, the growth rates were
from 0.82 mm in February through March to 1.13 mm in August. Interesting
enough, maximum growth values were registered during the rainy season
(June-November), though no significant changes were observed (P>0.05). Such
a growth fluctuations were thought to be density dependent. This factor
explained 52 to 92% of the observed variance. Daily mortality rates calculated
for the above two sites fluctuated from -0.1145 to -0.1539, and did not display
significant differences (P>0.05) neither troughout the dry nor rainy
seasons. Shrimp emigration from the lagoon roughly accounted for 25% of the
estimated mortality. To explain growth variability in the stuarine stages a
combined regulatory mechanism is proposed which includes environmental and
density dependent factors. The precipitation regime promotes among other
things, an increase in shrimp density which in turn affects individual
growth.
Key words:Growth, mortality, pink
shrimp estuarine stages, seasonal variations, growth regulatory
mechanism.
El camarón rosado,Penaeus (Farfantepenaeus) duorarumBurkenroad, 1903, constituye un recurso pesquero valioso en el SO del Golfo de México, cuya producción representa apoximadamente el 60% de la captura de peneidos (23 x 10³ toneladas anuales peso entero) que se obtiene en el Banco de Campeche (Soto et al., 1982; Anónimo, 1982). A pesar de la importancia económica de esta especie, la información disponible sobre la biología y ecología de las diferentes fases de su ciclo de vida es escasa, en particular respecto a los estadios que habitan los sistemas estuarinos de esta región del Golfo de México. En el Banco de Campeche, la Laguna de Términos representa un área de crianza esencial para las fases estuarinas de P. duorarum. La inmigración de postarlvas planctónicas es continua a lo largo del año (Arenas-Mendienta y Yáñez-Martínez, 1981; Sánchez-Martínez, 1981; Gracia y Soto, 1986b), lo cual da como resultado la presencia de hasta 12 diferentes cohortes en la laguna sujetas a diferentes condiciones ambientales. Las respuestas de estas cohortes a las variaciones estacionales son importantes desde el punto de vista ecológico-pesquero, para comprender las fluctuaciones en la captura de altamar, ya que afectan directamente el nivel de reclutamiento a la población adulta. La estimación de las tasas de crecimiento y mortalidad de los prereclutas (postlarvas epibentónicas y juveniles) deP. duorarum y sus variaciones estacionales en ambientes estuarinos constituyen aspectos relevantes dentro de la biología poblacional de este crustáceo. No obstante la mayoría de las estimaciones se han realizado sobre organismos adultos (Lindner, 1965; Kutkuhn, 1966; Berry, 1967; Shapiro, 1983; Arreguín Sánchez y Chávez, 1985, y poca atención se ha prestado al comportamiento estacional y anual de estos parámetros en fases estuarinas. El presente trabajo tiene como objeto principal la estimación de las tasas de crecimiento y mortalidad de las fases estuarinas de camarón rosado en la Laguna de Términos durante dos periodos climáticos contrastantes de acuerdo al régimen hidrológico (estío y precipitación). 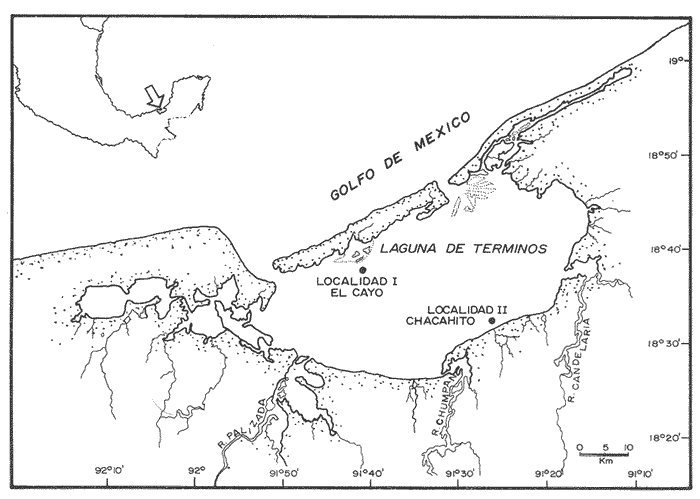 Figura 1. Situación de las localidades estudiadas en la laguna de Términos. ÁREA DE ESTUDIOLa Laguna de Términos está situada en el suroeste del Golfo de México y se comunica con la Bahía de Campeche por la Boca del Carmen al oeste y la Boca de Puerto Real al este. La laguna recibe el aporte de varios ríos entre los cuales los más importantes por su volumen de descarga son el Palizada, el Chumpán y el Candelaria. El clima de la laguna es cálido-húmedo AmW según la escala de García (1973). La precipitación pluvial en el área varía entre 1,100 a más de 2,000 mm anuales (Cruz-Orozco et al, 1979 a y b) con una temporada de lluvias de junio a septiembre. De octubre a marzo es común la presencia de vientos fríos provenientes del norte. El periodo de estío comprende de febrero a mayo. El desarrollo de este trabajo se llevó a cabo en dos localidades: El Cayo (localidad I) ubicada en los bajos del mismo nombre sobre el margen de la Isla del Carmen y Chacahito (localidad II) frente a la Laguna de Chacahito en el borde continental de la Laguna de Términos (Fig. l). La localidad I (El Cayo) se encuentra en un área de influencia marina cubierta de vegetación sumergida entre la cual predomina Thalassia testudinum. El sedimento es arenoso con un alto contenido de materia orgánica (Yáñez-Correa, 1963). La temperatura durante el periodo de etudio osciló entre 22.5°C y 32.3°C con un valor medio de 27.2°C. La salinidad presentó un intervalo de 13.4% (octubre) a 36.4% (marzo). La localidad II (Chacahito)está ubicada en una zona afectada por las descargas de los ríos que desembocan a la Laguna de Términos, y posee un sustráto cubierto por vegetación representada por Halodule wrightiiyThalassia testudinum. El sedimento presente es de tipo limo-arcilloso con arena fina (Yáñez-Correa, 1963). La temperatura fluctuó entre 23°C (enero) y 30.2°C (septiembre)con un valor promedio de 27.6'°. Los registros de salinidad variaron entre 10 a 28.5% Descripciones más detalladas de la Laguna de Términos y las localidades estudiadas se encuentran en Yáñez Correa (1963), Phleger y Ayala-Castañares (1971) y Yáñez-Arancibiaet al, (1983) entre otros. MATERIAL Y MÉTODOSEl periodo de estudio comprendió del 28 de julio de 1982 al 14 de mayo de 1983. Las colectas en cada localidad se tomaron en intervalos de 7 días con el fin de examinar las tasas de crecimiento de cada cohorte con mayor precisión. Las muestras se obtuvieron con una red de barra tipo Renfro (Renfro, 1962) de 1.8 m de largo por 0.7 m de boca y una malla de 0.8 mm. La red fue operada manualmente en transectos de 50 m de longitud orientados perpendicularmente a la línea de costa. El área cubierta por la red se estimó en 90 m². El material colectado se fijó inmediatamente en formol al 4% y posteriormente se separaron los camarones en el laboratorio. Los individuos se identificaron mediante las claves taxonómicas propuestas por Williams (1959), Cook (1966), Ringo y Zamora (1908) y Pérez Farfante (1970). Las mediciones de longitud total (L.T.) se obtuvieron con un microscopio estereoscópico graduado y un vernier (0.05 mm de precisión). Cuando el número de organismos colectados excedió a 200 se obtuvieron mediciones de sólo 150 individuos. TRATAMIENTO DE DATOSLos datos fueron agrupados en clases de 2 mm para la localidad 1 en virtud de la predominancia de organismos entre 6 y 30 mm de L.T. Para la localidad 11 se registró un intervalo de 8 a 80 mm de L.T. por lo cual se seleccionaron clases de 4 mm para reducir la dispersión de los datos. Las distribuciones de frecuencia de tallas de P. duorarumse analizaron mediante el método de Battacharya (1967) para la resolución de una distribución en sus componentes gausianos. Para la aplicación de este método se asumió que los individuos capturados representaron a la población de camarón rosado que se establece temporalmente en el área estudiada y de esta forma los muestreos sucesivos estiman la composición de las diferentes cohortes en cada localidad. La estimación de las tasas de crecimiento a partir de un análisis de progresión modal se realizó con las modas obtenidas por el procedimiento de Battachayra (1967). La mortalidad diaria de P. duorarum se estimó a través del ajuste de curvas exponenciales negativas (Nt = Noe-zt) a la distribución de frecuencia de tallas de los dos periodos climáticos considerados en este estudio (estío, de enero a marzo y de mayor precipitación pluvial de agosto a diciembre). La distribución de frecuencias de cada temporada se dividió en clases de edad de acuerdo a las estimaciones de crecimiento promedio de ambos sexos correspondiente, dado que no existe un crecimiento diferente por sexo (Gracia y Soto, 1986a). Este procedimiento incluye las variaciones promedio por temporada. El intervalo de tallas y edad empleado para realizar las estimaciones de mortalidad corresponden a un lapso corto del ciclo de vida de camarón; de tal forma, el sesgo introducido por la acumulación de organismos de mayor edad en grupos de tallas grandes fue mínima. La emigración de los camarones es un factor que puede introducir sesgos de mayor magnitud en las estimaciones de mortalidad. 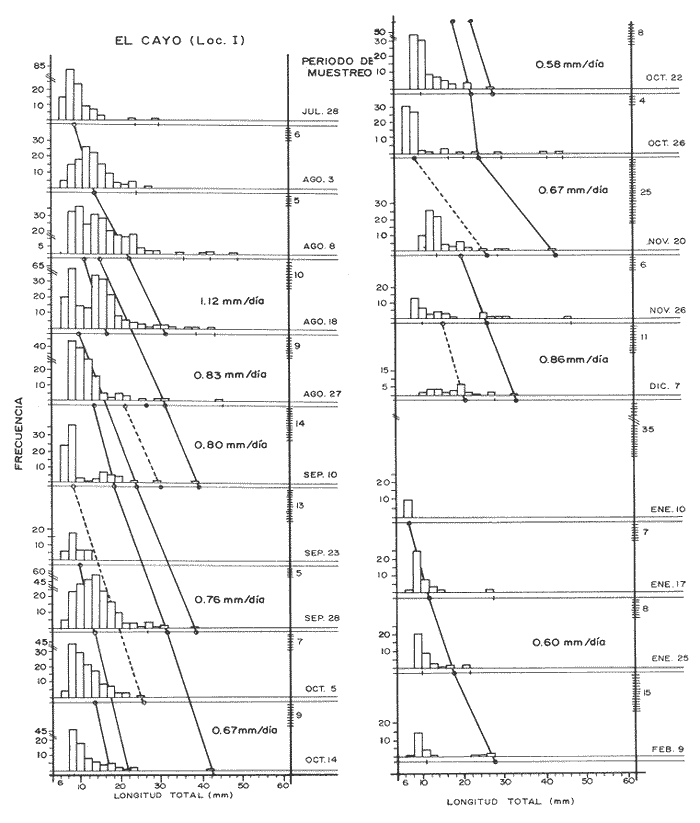 Figura 2. Estructura de la población y tasas de crecimiento de P. duorarum en el Cayo (Loc. I). RESULTADOSLa captura de camarones estuvo compuesta principalmente porPenaeus duorarum, el cual representó el 89.4% del total. En tanto que otros individuos identificados como P. setiferus, Xiphopenaeus kroyeriySicyonia sp constituyeron el 10.6% restante. El intervalo de tallas de P. duorarumcomprendió a los estadios de postlarvas y juveniles pequeños desde 6.2 a 80 mm de L.T. 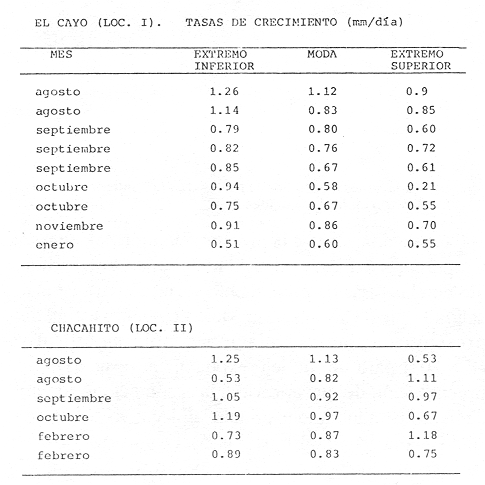 TABLA 1 T ASAS DE CRECIMIENTO DE P. Duorarum EN LAS DOS LOCALIDADES. CRECIMIENTO MODAL Y DE LOS EXTREMOS DE CADA DISTRIBUCIÓN GAUSIANA CARACTERÍSTICAS DE LA POBLACIÓNDurante este estudio se registraron fluctuaciones en la densidad de organismos en las localidades El Cayo y Chacahito. Se detectaron tres valores máximos en los siguientes periodos: verano (finales de agosto y principios de septiembre), otoño (octubre) e invierno (enero). Los dos valores de densidad más altos registrados (6.0 ind/m² en la localidad I y 1.0 ind/m² en la localidad II). Coincidieron con las salinidades más bajas (12%) durante el muestreo. La composición de tallas de P. duorarummostró diferencias entre las dos localidades. En la localidad I el promedio de tallas de longitud total varió entre 10 y 20 mm y se observó una presencia continua de individuos de tallas cercanas a 10 mm, lo cual indica un reclutamiento constante a esta área. En la localidad II los organismos presentaron una talla promedio entre 20 y 40 mm. En esta área también se colectaron organismos pequeños (10 mm, L.T.); sin embargo, la moda de reclutamiento se registró a los 28 mm de L.T. Los análisis de varianza de una vía aplicados a las tallas obtenidas, indicaron diferencias signficativas (P>.05) entre las muestras. A partir de este resultado se infiere que las fases estuarinas de camarón rosado se establecen transitoriamente y la composición de tallas de la población se modifica gradualmente en el tiempo. En el caso contrario la composición de tallas similar, sugeriría una corta permanencia de los individuos en el área y las estimaciones de mortalidad y crecimiento serían imprecisas. CRECIMIENTOLas curvas de crecimiento estimadas a partir del análisis de progresión modal basado en frecuencias de longitud de las fases estuarinas de P. duorarum se ajustaron a una relación lineal simple en todos los casos. Este modelo resultó el más adecuado para el patrón de crecimiento en tales etapas como lo comprueban los altos coeficientes de correlación (>0.9) de los ajustes practicados. En la localidad I las tasas de crecimiento fluctuaron desde 0.58 mm/día en octubre hasta 1.12 mm/día en agosto (Fig. 2). En la localidad II se analizaron seis cohortes cuyas tasas de crecimiento variaron desde 0.82 mm/día en los meses de febrero y marzo a 1.13 mm/día en agosto (Fig. 3). Adicionalmente se estimaron las tasas de crecimiento de los grupos ubicados en los extremos de cada una de las distribuciones gausianas, cuyas modas fueron empleadas para calcular las tasas de crecimiento promedio. Los valores obtenidos (Tabla 1) muestran que en 9 de los 15 casos, el extremo izquierdo de la distribución tiene una tasa de crecimiento más alta. Estos resultados sugieren que los organismos de mayor talla presentan una ligera disminución en las tasas de crecimiento dentro de la misma cohorte. Sin embargo, es conveniente señalar que los puntos empleados para estas estimaciones pueden estar sobrepuestos con componentes de las distribuciones contiguas. 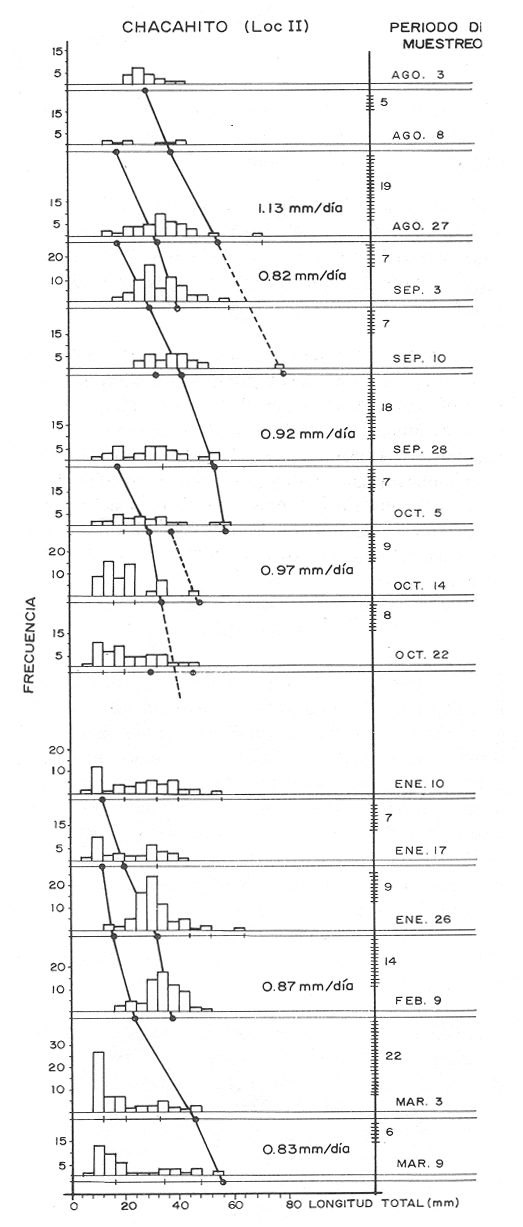 Figura 3. Estructura de la población y tasas de crecimiento de P.duorarum en el Chacahito (Loc. II). A pesar de que las tasas de crecimiento tienden a disminuir hacia finales del año (Fig. 4), no se registró una diferencia significativa (P>0.05) entre las pendientes calculas en cada localidad cuando se les examinó mediante un análisis de covarianza para comparación múltiple de pendientes. 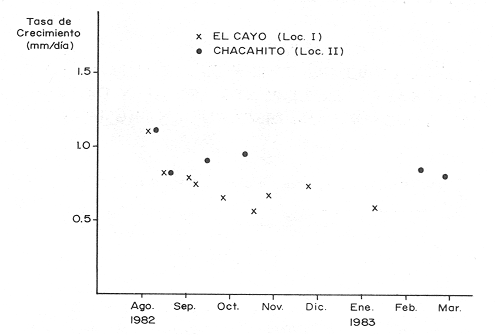 Figura 4. Variaciones mensuales de las tasas de crecimiento de P.duorarum en las localidades I y II. Las relaciones entre la precipitación pluvial mensual, temperatura del agua y densidad de organismos promedio con las tasas de crecimiento variaron para cada localidad. En el Cayo las tasas de crecimiento estimadas, presentaron una correlación simple significativa (P>0.05) para un modelo exponencial positivo con la precipitación del mes previo a las estimaciones de crecimiento y un modelo exponencial negativo con la densidad medía de organismos (Fig. 5). La correlación entre las tasas de crecimiento y la temperatura no fue sígnifícatíva (P>0.05). Al efectuarse un análisis de correlaciones parciales se comprobó que, entre las variables consideradas, la densidad afecta en forma considerable las tasas de crecimiento, ya que explica 52% de su varianza. La precipitación pluvial cuando se emplean los valores con un mes de retraso a las estimaciones de crecimiento justifica un mayor porcentaje (64%) de la varianza del crecimiento. En ambos casos las correlaciones son signficativas (P>0.05) (Tabla 2). En la localidad de Chacahito, de las correlaciones simples obtenidas, solo la efectuada con la densidad fue significativa (P>0.05) y se ajustó a un modelo líneal negativo (Fig. 5). De acuerdo al análisis de correlaciones parciales la densidad es responsable del 96% de la varianza en las tasas de crecimiento. La correlación más alta con la precipitación pluvial se presenta en un modelo potencial con los valores del mes real (en curso) y es menor que aquella obtenida entre la temperatura y el crecimiento; sin embargo, en ninguno de los dos casos la correlación es significativa (P>0.05). Por otro lado, la temperatura y la precipitación plulvial muestran una correlación lineal positiva significativa (P<0.05) (Tabla 2). 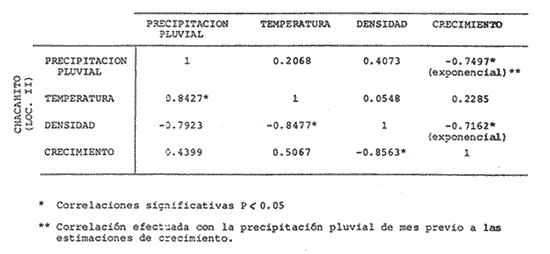 TABLA 2 CORRELACIONES SIMPLES DE LAS TASAS DE CRECIMIENTO DE P. DUORARUM Y LAS VARIABLES CONSIDERADAS EN LAS DOS LOCALIDADES 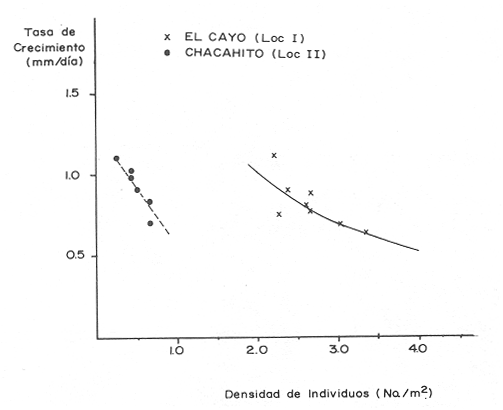 Figura 5. Relación entre las tasas de crecimiento y la densidad de individuos de P. duorarum. MORTALIDADLa estimación de la mortalidad diaria a través de los ajustes de la curva exponencial negativa dio como resultado una tasa de -0.1213 correspondiente al periodo de precipitación pluvial y de -0.1539 para el periodo de estío en la localidad I. La primera curva llega a la fase asintótica cuando han transcurrido 32 días y los organismos miden 32 mm de L.T.; la curva correspondiente al periodo de estío presenta la asíntota cerca de los 23 días cuando los organismos han alcanzado una talla de 22 mm de L.T. (Fig. 6). En la localidad II las tasas estimadas fueron: -0.1145 y -0.1125 para los periodos de precipitación y estío, respectivamente. En esta localidad las curvas se inician con clases de 24 a 28 mm de L.T. Durante el periodo de precipitación los organismos permanecen alrededor de 40 días y alcanzan una talla de 68 mm de L.T. En el estiaje la residencia es también de 40 días, pero los camarones sólo crecen hasta tallas de 60 mm de L.T. promedio (Fig. 7). Debido a que los cálculos de mortalidad diaria están sobreestimados por el efecto de la emigración, se elaboró la figura 8 a partir de un número inicial de 1,000 individuos y la mortalidad promedio anual en las dos localidades con el fin de estimar la magnitud del sesgo. La primera curva de la localidad I comprende desde la talla inicial de reclutamiento al área hasta 20 días de residencia que corresponde a una talla de 28 mm de L.T. 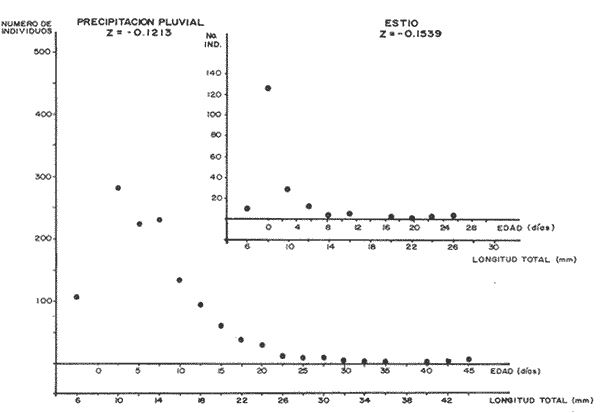 Figura 6 Curvas de mortalidad de P.duorarum durantedos periodos climaticos(mayor precipitación pluvial y estió )en Cayo(localidad 1) 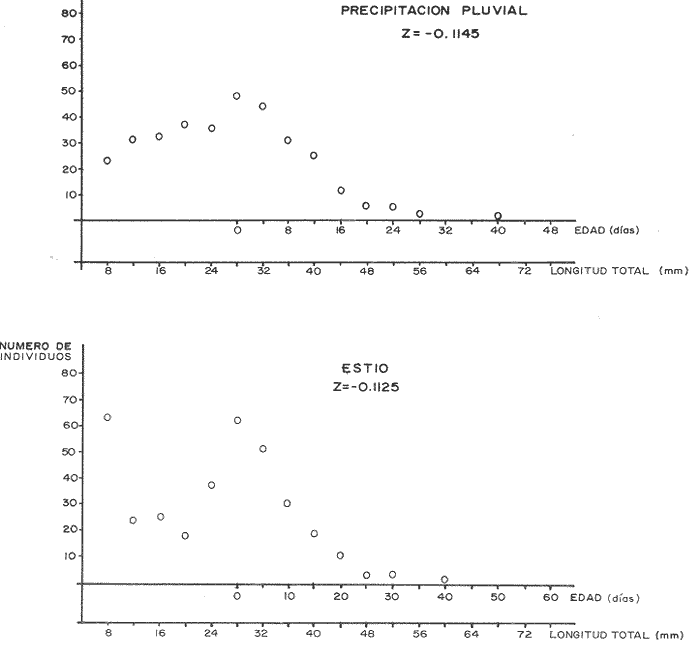 Figura 7. Curvas de mortalidad de P. duodarum durante dos periodos climáticos (mayo precipitación pluvial y estío) en la (localidad II). y además a la talla de reclutamiento a la localidad II. La segunda curva se inicia en la talla de 28 mm de L.T. y termina después que han transcurrido 45 días y los organismos tienen una talla de 44 mm de L.T. (Fig. 8). Estos ajustes dan valores de Z de -0.0958 y -0.0716, respectivamente. La comparación entre las dos curvas de mortalidad entre los días 20 y 25, cuando ocurre la mayor parte de la emigración en la localidad I, sugiere que aproximadamente el 40% de los organismos ausentes del área se deben a causas migratorias, lo cual representa cerca del 25% de la pendiente de mortalidad. No obstante la imposibilidad de asegurar que los individuos de la localidad I se desplacen a la localidad II ayudados por la circulación de marea, la estimación anterior es una aproximación de la magnitud del sesgo en las estimaciones de mortalidad de las fases estuarinas de camarón rosado en la Laguna de Términos debido a movimientos migratorios locales. Por otro lado, la disminución de la diferencia entre las dos curvas y por consecuencia del porcentaje de individuos que emigran de cada localidad conforme se incrementa el tiempo, sugiere que el proceso de emigración presenta máximos a determinada talla (localidad I, 26 mm de L.T.; localidad II, 44 mm de L.T.), después de lo cual disminuye paulatinamente. 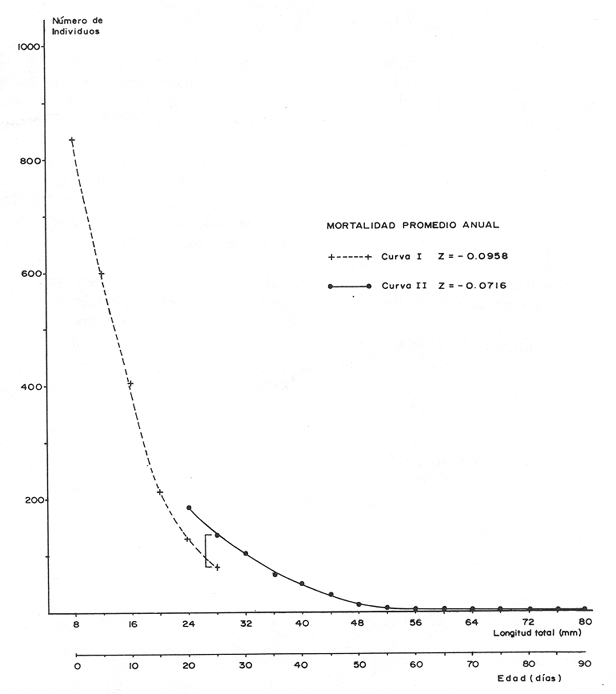 Figura 8. Comparación de las curvas de mortalidad promedio anual de P.duorarum en las dos localidades. La edad estimada está basada en un crecimiento promedico de 0.80 mm/día. DISCUSIÓN
CRECIMIENTOLas tasas de crecimiento deP. duorarum estimadas durante un ciclo anual coinciden con los intervalos de tasas de crecimiento registrados en la fase juvenil de las especies de peneidos comercialmente más importantes del Golfo de México. Entre estas estimaciones figuran las estimadas paraP. aztecus que varían entre 0.5 y 3.3 mm/día de L.T. (Cook y Lindner, 1970; Ford y St. Amant, 1971; Jacob, 1971) y paraP.setiferus entre 0.6 y 2.2 mm/día de L.T. (Williams, 1955; Gunter, 1956; Loesch, 1965; Klima, 1974). El crecimiento de P. duorarum en su etapa de postlarva y juvenil es similar a los valores señalados por Gracia y Soto (1986a) (0.83 mm/día) y Aguilar Sierra (1985) (0.76-1.2 mm/día) en juveniles de P. setiferus obtenidos a partir de intervalos de tallas mayores al empleado para el camarón rosado en este trabajo. La comparación entre el crecimiento de las dos especies sugiere una tasa de crecimiento menor para el camarón rosado en la Laguna de Términos donde coexiste con el camarón blanco, ya que generalmente la tasa de crecimiento es mayor en individuos pequeños y disminuye cuando aumenta la talla. Esta tendencia en la disminución de las tasas de crecimiento al incrementar al tamaño del organísmo se observó en los individuos ubicados en el extremo superior de las distribuciones de tallas deP. duorarum en este trabajo. En la Tabla 3 se muestran diferentes estimaciones de crecimiento de P. duorarum proporcionadas por varios autores, entre los cuales sólo en la tasa señalada por Galois (1973) para P. duorarum notialis se emplea un intervalo de tallas similar al de este estudio. La tasa de crecimiento estimada se aproxima al valor máximo calculado para el camarón rosado en la Laguna de Términos. El resto de las estimaciones provienen de un intervalo de tallas mayor y sus valores se acercan al obtenido en este trabajo; sin embargo, se debe tomar en cuenta que los valores de crecimiento señalados pueden cambiar en función de variaciones estacionales. Con respecto a las tasas de crecimiento calculadas a partir de las ecuaciones de von Bertalanff`y de adultos de P. duorarum en el Banco de Campeche (Shapiro 1983; Arreguín-Sánchez y Chávez, 1985), éstas son mas altas que el valor máximo estimado en la Laguna de Términos. Este resultado sugiere que la extrapolación del modelo de von Bertalanffy obtenido en la fase marina hacia los primeros estadios puede conducir a sobreestimaciones del crecimiento de estas etapas. Por otro lado, como señala Sainsbury (1980) puede ser el resultado del efecto de la variabilidad individual lo cual también puede originar sobreestimaciones de la longitud media de cada cohorte y mas aún cuando se hacen extrapolaciones. Independientemente de la causa, esta diferencia en las estimaciones plantea interrogantes acerca del efecto que puedan tener en los parámetros de la curva de crecimiento y en especial sobre modelos de producción que emplean estos parámetros. Además, reafirma la importancia de considerar las tasas de crecimiento en la fase estuarina, así como sus variaciones estacionales para la estimación correcta de parámetros y análisis de la población adulta. 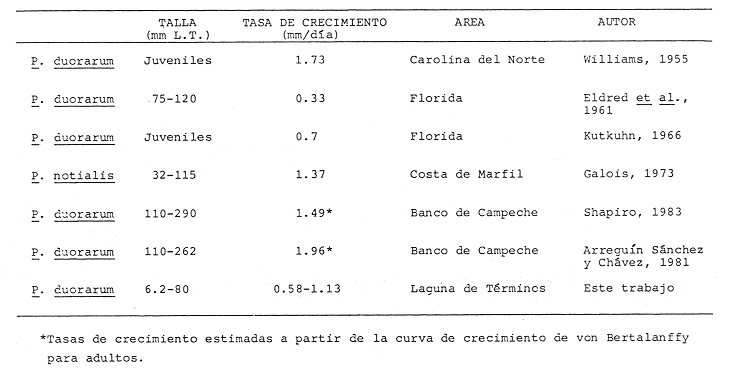 TABLA 3 COMPARACIÓN DE LAS TASAS DE CRECIMIENTO DE CAMARÓN ROSADO REGISTRADAS POR DISTINTOS AUTORES Variaciones estacionales en la tasa de crecimiento de peneidos han sido señaladas por diferentes autores (Edwards, 1977; Staples, 1980; Klima, 1981), cuyos valores máximos y mínimos están asociados a los periodos de verano e invierno como respuesta a la temperatura del agua. En las fases estuarinas deP. duorarum se observa una tendencia similar con tasas promedio más bajas en invierno, asociadas a valores mínimos de precipitación (8 mm); sin embargo, las variaciones en las tasas durante el ciclo anual no son estadisticamente diferentes. Cabe hacer notar que los tres autores anteriores no establecen pruebas estadísticas para verificar tales diferencias y se limitan a señalar las fluctuaciones. En el caso de P. duorarum las diferencias no significativas observadas pueden estar relacionadas con la variabilidad en las tasas de crecimiento que presentan las cohortes mensuales; esto sucede en la localidad I, donde el camarón rosado registra las tasas más altas y más bajas en el periodo de lluvias. La temperatura, como factor importante en los cambios de las tasas de crecimiento de peneidos, ha sido señalada en diferentes lugares con cambios estacionales marcados (Ford y St. Amant, 1971; Phares, 1980; Klima, 1981). En la Laguna de Términos este parámetro tiene un intervalo reducido de variación (25-30°C), que incluso se encuentra por arriba de la temperatura óptima de crecimiento (25°C) para postlarvas obtenida en condiciones experimentales (Zein-Eldin y Griffith, 1967; Costello y Allen, 1970). La temperatura no representa un factor limítante sobre el crecimiento, lo cual se demuestra por la baja correlación entre ambos parámetros (Tabla 2). Además, la relativamente baja estacionalidad del área de estudio no propicia la diferencia entre las características de las distintas cohortes a lo largo del año. Las correlaciones simples y parciales en las localidades I y II señalan a la densidad media de organismos en este ambiente tropical como la variable más importante para determinar las tasas de crecimiento de camarón rosado; esto sugiere un mecanismo de competencia a traves del cual, el factor limitante para el crecimiento puede ser la disponibilidad de alimento. La relación entre la densidad y el crecimiento presenta una forma exponencial negativa típica entre estos parámetros, la cual ha sido registrada en general para diferentes organismos (Weatherley, 1972) y en peneidos para P. vannamei(Edwards, 1977). La razón de una relación lineal negativa en la localidad II puede deberse a que en esta área el intervalo de densidades medias es pequeño y no se alcanza a definir la curva en su totalidad. La influencia de la precipitación pluvial sobre la producción de las poblaciones de camarones ha sido abordada por varios autores en diferentes especies (Ruello, 1973; Castro Aguirre, 1976; Castro Ortiz y Sánchez Rojas, 1976; Christmas y Eltzod, 1977; Vanceet al., 1985). En la Laguna de Términos, Gracia y Soto (inédito) señalan una correlación positiva entre la densidad de juveniles de P. duorarum y la precipitación mensual, la cual puede estar relacionada con el mayor aporte de nutrientes a la laguna por la descarga de los ríos. La correlación negativa entre el crecimiento y los volúmenes de precipitación pluvial del mes previo en la localidad I que de acuerdo a las correlaciones parciales explica la mayor parte de la varianza (62%); sugiere un mecanismo regulador mixto del crecimiento deP. duorarum, en el cual intervienen factores de tipo ambiental, sucedidos por la acción de factores densodependientes. De esta forma, primero aumenta la capacidad de carga del área en función del alimento disponible, y después actúa un efecto densodependiente sobre el crecimiento individual promedio. La productividad en fitoplancton y manglares según Day et al. (1982) en la Laguna de Términos durante la época de lluvias se ve afectada positivamente, mientras que la producción en pastos marinos disminuye en las áreas de mayor influencia marina situadas al Este de la Laguna. No obstante, los valores de producción cercanos a la localidad I reportados por estos autores son similares en ambos periodos y como ellos mismos mencionan el fitoplancton acarreado a las zonas de pastos marinos sirve como una fuente importante de nutrientes. Estos resultados apoyan la existencia del mecanismo regulador mixto propuesto aún cuando la correlación entre la precipitación y la densidad de P. duorarum en este trabajo no alcanza niveles significativos. En la localidad II el mecanismo anterior no se ajusta enteramente, ya que la precipitación pluvial no afecta el crecimiento con un periodo de retraso a través de la densidad; sino que el número de individuos se ve influenciado negativamente por la temperatura del mismo periodo, la cual a su vez está directamente correlacionada con la precipitación pluvial. La densidad de organismos en esta área presenta variaciones en un pequeño intervalo cuyos máximos están asociados a la temporada de estío, lo cual sugiere que además del alimento disponible estén involucrados otros factores en esta etapa estuarina del camarón; sin embargo, la salinidad cuyos incrementos estarían asociados al estío, tampoco actúa sobre la densidad de organismos en esta localidad (Alvarez, 1984). El efecto negativo de la temperatura y precipitación pluvial sobre la densidad media de organismos y la mayor productividad de la Laguna de Términos durante la temporada de mayor precipitación pluvial ocasionan que las tasas de crecimiento más altas se presenten en este periodo. Las tasas que alcanzan P. duorarum en esta localidad son semejantes a los valores más altos obtenidos para la localidad I en un intervalo de tallas menor. La diferencia entre los mecanismos por medio de los cuales la densidad afecta el crecimiento en las dos localidades estudiadas, refleja la respuesta de P. duorarum a dos hábitats distintos, cuya definición es esencial para comprender el comportamiento de camarón rosado en el ambiente lagunar estuarino. MORTALIDADLas tasas de mortalidad natural estimadas para las dos temporadas no difieren estadísticamente (P>.0.05) en las dos localidades, sólo en la localidad I la tasa calculada es mayor durante el periodo de estío sin alcanzar valores significativos. Esta diferencia puede estar relacionada con el corto período de residencia (23 días) de las postlarvas en la temporada de menor precipitación, lo cual sugiere una emigración temprana que tiene como consecuencia una sobreestimación de la mortalidad. El alimento disponible influenciado indirectamente por la precipitación pluvial, podría ser una de las causas de este movimiento local hacia áreas de pastos marinos con mayor productividad, como la parte noreste de la laguna señalada por Dayet al.(1982). Los resultados de esta comparación entre las tasas de mortalidad, al igual que lo que sucede en el crecimiento, confirman un comportamiento diferente del camarón rosado en las dos localidades. Por otro lado, es conveniente hacer notar que las tasas de mortalidad comparadas corresponden a las estimaciones promedio por temporada y no reflejan el comportamiento particular de las cohortes, ya que suavizan las variaciones de la mortalidad en cada una de ellas. El análisis fino de las variaciones de mortalidad con respecto a las tasas de crecimiento y los factores ambientales sería interesante desde el punto de vista ecológico y pesquero; sin embargo, se necesita información más detallada para llevarlo a cabo. Por lo pronto, las estimaciones efectuadas muestran los niveles de mortalidad deP. duorarum en la Laguna de Términos, que sólo son comparables a las tasas semanales (-0.76 a -1.51) proporcionadas por Kutkuhn (1966) para juveniles más grandes de camarón rosado en el área de Dry Tortugas, Florida. Al incorporar la estimación promedio anual de la emigración entre las localidades, la mortalidad natural alcanza valores entre 36 y 52%. Edwards (1977) en juveniles de P. vannameiseñala un valor similar (41%) de mortalidad total, mientras que Gracia y Soto (1986a) mencionan que la mortalidad natural de juveniles deP. setiferus en el área de la Laguna de Términos representa 20% semanal. Estas diferencias entre los estados de desarrollo se hacen más notables cuando se comparan los efectos de la mortalidad natural entre la fase estuarina y marina de P. duorarum en la misma región. Las tasas de mortalidad natural enP. duorarum cambian desde los altos valores mensuales (-3.375 a -44.617) calculados en este trabajo hasta la M registrada (-0.27 y -0.3771) para los adultos sujetos a explotación comercial. De acuerdo al tiempo medio de residencia (6-10 semanas) deP. duorarum en la Laguna de Términos (Gracia y Soto, inédito; Alvarez, 1984) las estimaciones indican una mortalidad total del 97% a 99% entre la fase de postlarva y subadulto; con una mortalidad media semanal de 45%. No obstante, esta cifra es posible que esté sobreestimada, ya que de acuerdo a la tendencia que presenta la curva de mortalidad en estos organismos y el efecto de emigración no estimados en la segunda localidad, la mortalidad debe disminuir en juveniles de tallas (80 mm de L.T.) no considerados en este estudio. ConclusionesLos registros más altos (1.12-1.13 mm/día) de las tasas de crecimiento deP. duorarum se presentaron durante el periodo de mayor precipitación pluvial. Sin embargo, las tasas estimadas durante un ciclo anual no mostraron diferencias significativas (P>0.05). La densidad media es el factor más importante que afecta las tasas de crecimiento del camarón rosado, responsable de 52 a 96% de la varianza de éstas. La relación entre la densidad y las tasas de crecimiento presenta una forma exponencial negativa. Las tasas de crecimiento deP. duorarum están reguladas mediante la interacción de la precipitación pluvial con un periodo de retraso sobre la densidad media de organismos y el efecto de factores dependientes sobre el crecimiento individual. Las tasas diarias de mortalidad natural de camarón rosado estimadas (-0.1145 a -0.15239) para dos periodos climáticos (estío y mayor precipitación pluvial) no difieren estadísticamente (P > 0.05). Los movimientos migratorios de P. duorarumen la fase estuarina introducen un sesgo aproximado de 25% en las estimaciones de mortalidad. AgradecimientosEste trabajo se realizó con el apoyo del Consejo Nacional de Ciencia y Tecnología y el Instituto de Ciencias del Mar y Limnología a través del proyecto PCMANAL-790334. Se agradece la colaboración de los participantes de dicho proyecto y el personal de la estación "El Carmen" en el trabajo de campo. Asimismo a las autoridades del Instituto de Ciencias del Mar y Limnología y la Estación "El Carmen" por su apoyo durante la realización de este estudio. LITERATURAAGUILAR SIERRA, V.A., Tesis Prof. Fac. Ciencias,Camarones peneidos de la Laguna de Términos, Campeche: Composición, Distribución y Parámetros Poblacionales.Univ. Nal. Autón. de México.1985.62 p. ALVAREZ, F., Tesis Prof. Fac. de Ciencias,Aspectos Poblacionales de las Postlarvas epibétnicas de Penaeus Farfantepenaeus duorarum, Burkenroad, 1939 en la Laguna de Términos, Campeche.Univ. Nal. Autón. México.1984.60 p. ANÓNIMO,Anuario Estadístico de Pesca de 1981.Dirección General de Informática y Estadística, Secretaría de Pesca.1982.796 p. ARENAS MENDIETA, M. R. y A. YÁÑEZ MARTÍNEZ,Tesis Prof.Fac. Ciencias Patrón anual de inmigración de postlarvas de camarón (Crustacea: Decapoda: Penacidae), en la Boca de Puerto Real, Laguna de Términos Campeche. Univ. Nal. Autón. México.1981.92 p. ARREGUÍN SÁNCHEZ, P. y E. A. CHÁVEZ, Estado del Conocimiento de las Pesquerías de camarón del Golfo de México. (en prensa).Investigaciones Marinas del CICIMAR,1985.2(2) BATTACHARYA, C.G., Biometrics, A simple method of resolutionn of a distribution into gaussian componentes.1967.115-135.23(1): BERRY, R. J., Ph. D. Thesis. Dynamics of the Tortugas pink shrimp population.University of Rhode Island.1967.160p. CASTRO AGUIRRE, J.L., Mem. Simposio sobre Biología y Dinámica Poblacional de Camarones. Guaymas, Son., México, Efecto de la temperatura y la precipitación pluvial sobre la producción camaronera.In: Castro Aguirre J.L. (Ed.).1976.74-88.1:8 al 13 de agosto de 1976, CASTRO ORTIZ, J.L. y M.A. SÁNCHEZ ROJAS, Mem. Simposio sobre Biología y Dinámica Poblacional de Camarones. Guaymas, Son. México Notas preliminares del comportamiento y dinámica poblacional de Penaeus stylirostris Stimpson, 1971, en los sistemas lagunarios del centro de Sinaloa. In: Castro Aguirre J.L (Ed.).1976.213-254.11:8 al 13 de agosto de 1976. COOK, H. L, Fish. Bull., A generic key to the protozoean, mysis and postlarval stages of the littoral penacidae of the northwestem gulf of Mexico.1966.437-447.65(2): COOK, H. L, y M. J. LINDNER, Synopsis of biological data on the brown shrimp, Penaeus aztecus Ives, 1891.FAO Fish. Rep.,1970.1471-97.57(4): COSTELLO, T. J. y D. M. ALLEN, Synopsis of biological data on the brown shrimp, Penaeus duorarum Burkenroad. FAO Fish. Rep.,1970.1499-537.57(4): CRUZ-OROZCO, R-, F. LEY LOU, A. MACHADO NAVARRO Y V. M. ALBA CORNEJO, An. Inst. de Geofísica, Resumen de las condiciones meteorológicas registradas en las estación de Investigaciones Marinas "El Carmen" durante el año 1979.Univ. Nal. Auton. México,1979a.39-52.25: CHRISTMAS J. Y. y D. J. ETZOLD,Coast Research Laboratory Technical Report Series The shrimp fishery of the Gulf of Mexico. United States: A regional management plan. Gulf.1977.128p.(2): DAY, J. W., R. H. DAY, M. T. BARREIRO, F. LEY LOU y C.J. MADDEN,Coastal Lagoon Oceanol. Acta Primary Production in the Laguna de Términos, a tropical estuary in the Southern Gulf of Mexico. p. 269-280. Vol. Spec., In: Lasserre P.yH. Postma (Eds.).1982.462 p.5(4): EDWARDS, R. R. C., Field Experiments in Growth and Mortality of Penaeus vannamei in a Mexican Coastal Lagoon Complex.Estuarine and Coastal Marine Science,1977.107-121.5: FORD, T.B. y L.S. ST. AMANT, Management guidelines for prediction of brown shrimps, Penaeus aztecus, production in Louisiana.Proc. Gulf and Caribb. Fisheries Institute,1971.149-161,23: GALOIS,R.Marguages de P. duorarum on Cote d'Ivoire RE-sultats preliminaires: taux de recapture, migrations, croissance.Doc. Scient. Centre Rech. Oceanogr. Abdijan,1973.29-48.4(3): GARCÍA, S., Modificaciones al Sistema de Clasificación Climatica de Köpen, para adaptarlo a las condiciones de la República Mexicana.Inst. Geofísica, Univ. Nal. Autón. México.1973.246p. GRACIA A. y L. A. SOTO, Estimación del Tamaño de la Población, Crecimiento y Mortalidad de los juveniles de P. setiferus (Linnaeus, 1767) mediante marcado-recaptura en la Laguna de Chacahito, Camp. México. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México,1986a.217-230.13(3): GRACIA, A. y L A. SOTO,IOC/FAO Workshop on Recruitment in Tropical Coastal Demersal Communities. Condiciones de reclutamiento de las poblaciones de camarones peneidos en un sistema lagunarmarino tropical: Laguna de Términos-Banco de Campeche: In: Yáñez-Arancibia, A. y D. Pauly (Eds.). IOC Workshop Report No. 44:1986b.257-265. GRACIA, A. y L A. SOTO. II Annual fluctuations and production of the shrimp populations (Inédito).Studies of penaeid shrimp in Terminos Lagoon, Campeche, Mexico. GUNTER, G., Principies of shrimp fishery management.Proc. Gulf and Caribb. Fish. Inst.,1956.99-106.8: JACOB, W. J., M.S. Thesis, Observation on the distríbution, growth, survival and biomass of juvenile and subadult Penaeus aztecus in Southern Lousiana.Louisiana State University.1971.168. KLIMA, E.F., A white shrimp Mark Recapture study.Trans. Am. Fish. Soc.,1974.107-113.103(1): KLIMA, E.F., Kuwait Bulletin of Marine Science,The National Marine Fisheries Service shrimp research program in the Gulf of Mexico. Proceedings of the International Shrimp Releasing, Marking and Recruitment Workshop, In: Farmer A.S.A. (Ed.).SalmiyaState of Kuwait.1981.185-207.(2):25-29 November 1978, KUTKUHN, J.H., U.S. Fish and Wildlife Service Fishery Bulletin, Dynamics of a penaeid shrimp population and management implication.1966.313-338.65: LINDNER, M.J., What we know about shrimp size and the Tortugas Fishery.Proc. Gulf. Caribb. Fish. Inst.,1965.18-25.18: LOESCH. H.C., Distributions and growth of penacid shrimp in Mobile Bay Alabama. Publs. Inst. Mar. Sci. Univ. Tex .1965.41-58.10: PÉREZ-FARFANTE, I.,Diagnostic characters of juveniles of the shrimp Penaeus aztecus, p. duorarum and P. brasiliensis (Crustacea, Decapoda, Penaeidae).US Fish. Wildl. Serv. Spec. Sci. Rep. (fish),1970.599. PHARES, P. L, Temperture associated growth of white shrimp in Louisiana. NOAA Technical Memorandum, NMFS-SEFC -56.1980.19p. PHLEGER, F. By A. AYALA-CASTAÑARES, Bull. Am. Ass. Petrol. Geol., Processes and history of Terminos Lagoon, Mexico.1971.2130-2140.55(2): RENFRO, W. C., Small beam net for sampling postlarval shrimp.US Fish. Wildl. Selv. Circ.,1962.86-87.161: RINGO, R.D. y G. ZAMORA, Bull. Mar. Sci., A penaeid postlarval Character of taxonomic value.1968.471-476.18(2): RUELLO, N.V., Marine Biology, The Influence of Rainfall on the Distributions and Abundance of the School Prawn Metapenaeus macleayi in the Hunter River Region (Australia).1973.221-228.23: SAINSBURY, K.J., Effect on Individual variability on the von Bertalanffy Growth Equation. Can. J. Fish. Aquat. Sci.,1980.241-247.37: SÁNCHEZ MARTÍNZ A., Tesis Prof. Fac. de CienciasComportamiento anual de las postlarvas epibénticas de camarones pencidos en el sector oriental de la Laguna de Términos. Univ. Nal. Autón. México.1981.97 p. SHAPIRO, B., M. Sc. thesis, Populations-Dynamics of the pink shrimp (Penaeus duorarum Burkenroad) based upon the fishery of the Campeche Banks, México.Univ. of Miami:1983.50 p. SOTO, L A., M.C. RODRÍGUEZ DE LA CRUZ, R. RODRÍGUEZ, A. GRACIA, F. J. SOTO, F. ARREGUÍN y E. CHÁVEZ, Reunión Nacional sobre la Investigación Científico-Pesquera. Cocoyoc, MorelosAnálisis del conocimiento Científico-Tecnológico Nacional de las Pesquerías de Camarón en el Golfo de México.Sec. de Pesca, Instituto Nacional de la Pesca. Univ. Autón. Metropolitana.1982.26-029 mayo de 1982. STAPLES, DJ., Ecology of the Juvenile and Adolescent Banana Prawns, Penaeus merguiensis, in a mangrove Estuary and Adjacent Off-shore Area of the Gulf of Carpentaria. II* Emigration, Population Structure and Growth of juveniles. Ausst. J. Mar. Freshwater Res.,1980.653-65.31: VANCE, DJ., D.J. STAPLES y J.D. KERR . Factors affectingyear to year variation in the catch of banana prawns Penaeus merguiensis) in the Gulf of Carpentaria,Australia. J. Cons. Int. Explor. Mer.,83-97.42: WEATHERLEY, A.N., Growth and Ecology of Fish Populations.Academic Press.London New York .1972.239 p. WILLIAMS, A.B., Bull. Mar. Sci. Gulf and Caribb.,Contributions to the life histories of commercial shrimp (Penaeidae) in North Carolina.1955.116-146.5: WILLIAMS, A.B., Bull. Mar. Sci. Gulf Carib., Spotted and brown shrimp postlarvae (Penaeus) in the North Carolina.,1959.281-290.9(3): YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. CHAVANCE y D. FLORES-HERNÁNDEZAn. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Environmental behavior of Terminos Lagoon Ecological System, Campeche, México.1983.137-176.10(1): YÁÑEZ CORREA, A., Bol. Inst. Geol.Batimetría, salinidad, temperatura y distribución de los sedimentos recientes en la Laguna de Términos, Campeche, México.Univ. Nal. Autón. México,1963.147.67(1): ZEIN-ELDIN, Z.P. y G.W. GRIFFTTH, Biol. Bull., The effect of temperature upon the growth of laboratory held postlarval Penaeus aztecus.1967.186-196.131(1):
|