|
A PHISICAL APPROACH TO THE POSTLARVAL Penaeus
IMMIGRATION MECHANISM IN A MEXICAN COASTAL LAGOON (CRUSTACEA: DECAPODA:
PENAEIDAE)
Trabajo recibido el 10 de enero de 1986 y aceptado para su
publicación el 21 de mayo de
1986
J. A. Calderón-Pérez
Instituto de Ciencias del Mar
y Limnología, Estación Mazatlán. Apdo postal 811,
Mazatlán 8200, Sinaloa,
México.
C. R. Poli
Universidad Federal de Santa Catarina, Brasil. Centro
de Ciencias Agrarias, Departamento de Acuicultura. Caixa Postal 476, Santa
Catarina 88041, Brasil. Contribución 522 del Instituto de Ciencias del
Mar y Limnología,
UNAM.
Se
realizó un estudio para explicar el mecanismo de migración de las
postlarvas dePenaeus . Se llevó a cabo un programa
mensual de muestreos de 24 horas en la boca del Río Baluarte, Sinaloa,
México, que es un punto de acceso permanente al sistema lagunar
Huizache-Caimanero. Se hicieron muestreos adicionales en el mar, en el
área cercana, en tres zonas particulares. Se midió la velocidad y
dirección de la corriente litoral y se siguió el patrón
general de movimiento del agua cerca de la boca por medio de flotadores. La
duración y dirección del viento se obtuvo de una fuente
confiable. Las postlarvas de Penaeus y la biomasa de
zooplancton fueron más abundantes cerca de la zona de rompientes y
corriente litoral que en aguas más profundas. Se asumió que
existe una acumulación de postlarvas cerca de la boca del río en
el periodo de reflujo ya que se observó que la densidad de postlarvas de
tres de las cuatro especies fué más alta durante el primer tercio
del periodo de flujo que en los otros dos tercios. La migración de
postlarvas de Penaeus hacia la costa se explica en
términos de dirección del viento y corrientes de deriva. Se
discuten alternativas conductuales para la
migración
In
order to explain the postlarvalPenaeus migration mechanism a
study was undertaken. A 24 hr monthly sampling program was carried out at the
Baluarte River mouth, Sinaloa, México, which is a permanent access point
to the Huizache-Caimanero lagoon complex. Additional sampling at sea, in a
nearby arca, was made at three distinct zones. Longshore current speed and
direction was measured and general water motion pattern near the mouth was
followed by means of floats. Wind duration and its direction was obtained from
a reliable source.Penaeus postlarvae and zooplankton
biomass were more abundant near the breaker zone and in the longshore current
than offshore. An accumulation of postlarvae near the river mouth at ebb tide
was assumed since the postlarval density of three out of four species was
higher during the first third of the flood tidal period than in the other two
thirds. The postlarvalPenaeus migration onshore is explained
in terms of wind direction and drift currents. Behavioural alternatives for
migration are
discussed.
Shrimp production in the Huizache-Caimanero lagoon complex, Sinaloa, México depends on two main factors: successful postlarval immigration and abundant rainfall (Chapa Saldaña and Soto López, 1969; Edwards, 1978). Penaeus postlarvae are know to inhabit coastal water bodies such as lagoon and channels(esteros) in which they find suitable environmental conditions lo carry through an important part of their life cycle up to the pre-adult stage (Cardenas Figueroa, 1950; Pearson, 1939). The spawning grounds are located at a considerable distance from these nursery areas (Rodríguez de la Cruz, 1976) thus a migration mechanism which explains the presence of shrimp in such water bodies must be involved. There is not agreement as to the nature of this mechanism, whether it is intrinsinc or extrinsic. Some authors favour the former, based on the behavioural responses of postlarvae, under laboratory condition; to some of the physical factors, which could act upon the larvae in their natural environment to orientate their movements to wards the source of such stimulus. Low salinity water has been frequently suggested as an important cue for postlarval immigration (Edwards, 1978; Huges, 1969; Keiser and Aldrich, 1973; Linder and Anderson, 1956; Mair, 1979b, 1980; Mair et al., 1982; Young and Carpenter, 1977). Changes in pressure has also been mentioned as a possible triggering factor for vertical migration (Mairet al., 1982). Similarly, postlarval immigration has been suggested to be motivated by a temperature gradient between estuarine and marine waters; i. e. when the temperature is higher in the coastal lagoon than in the sea (Macías-Regalado, in press). Finally, water with lagoon taste ,to which postlarvae orientate under experimental conditions, has been mentioned also as a cue for immigration (Mair, 1980; Mair et al.,1982). Several authors, on the other hand, have observed that larval migration inPenaeus from the spawning grounds to the nursery areas is the result of the water movements such as ocean currents (Jones et al., 1970; Kennedy and Barber, 1981; Racek, 1959), wind drift (Racek, 1959) and tides (Penn, 1975). Once inside the channels, most authors agree that flood tide is the main agent for postlarval immigration (Calderón-Pérez, 1977; Hughes, 1969; Macías Regaladoet al., 1982; Mair, 197%, 1980; Noor, 1976; Poli and Calderón-Pérez, 1987; Racek, 1959; Rossler and Rehrer, 1971; Watkins, 1980). The purpose of this paper is to explain the onshore larval transport from the spawning grounds and the eventual entry to the system, based on observations of wind direction, longshore current and tidal period, associated to changes of postlarval density in space and time. STUDY AREAThe Huizache-Caimanero lagoon complex in Sinaloa, México has been described extensively by several authors such as Ayala-Castañareset al.,1970; Edwards, 1977, 1978; Lankford, 1977, Macías-Regalado and Calderón- Pérez, 1979,1980; Mairet al., 1982; Menz and Bowers, 1980; Soto López, 1969; Warburton, 1978; Watkins, 1980. The study area is located in the Baluarte River mouth at the Southern end of the system (Fig. 1). 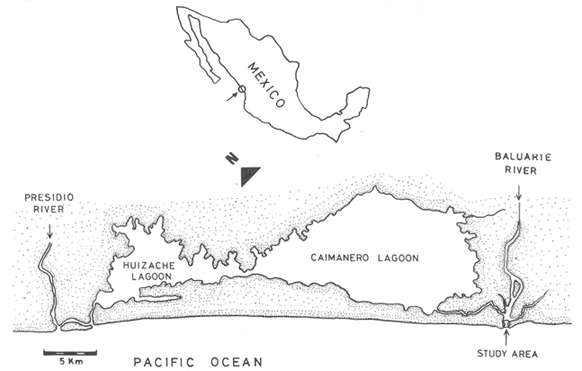 Figure 1. A diagram of the Huizache-Caimanero lagoon complex showing the study area. MATERIAL AND METHODSA series of 24 hr monthly samples was carried out from January 1981 to January 1982 during the full moon flood tides. Sampling was made at intervals ranging from 1 to 2 hours depending of the duration of the effective flood, i. e. flood duration inside the Huizache-Caimanero lagoon complex decreases proportionally to the Baluarte river discharge (Poli and Calderón-Pérez, 1987). Postlarval Penaeus was collected at surface and midwater by means of two conical plankton nets attached to a frame held at the bow of a 4 m long boat (Fig. 2A, B), propelled by a 15 HP outboard motor. Bottom. samples (Fig. 2C) were taken by means of a Colman-Segrove net (1952). Dimensions are: mouth, 0.24 x 0.48 m; length, 1.47 m; mesh 500u. All nets were equipped with a calibrated General Oceanics flow-meter. 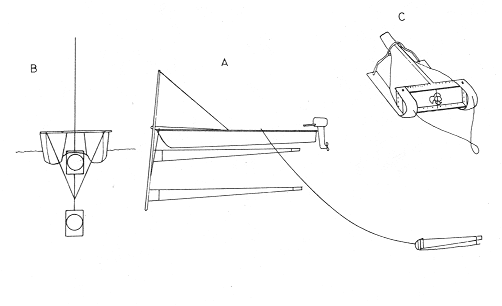 Figure 2. A schematic diagram of the plankton sampling method A lateral view of the three plankton nets towed simultaneously. B, frontal view of the surface and midwater plankton nets. C, Colman-Segrove net. Additional sampling was carried out on the coast between Mazatlán and the Presídio river mouth in September 1982 in three distinct zones: on the longshore current (L.C.), at few meters short of the breaker zone (N.B.) and offshore, at the 10 m isobath (0). The location of the 30 stations is indicated in figure 3. The sampling method was identical for stations 11 to 30 i.e. towing the plankton nets described above for 5 minutes at a speed of 2 knots (1m/sec) on a 5 m long boat propelled by a 70 HP outboard motor. Stations 1 to 10 were sampled on foot, towing the Colman-Segrove net by hand against the longshore current. 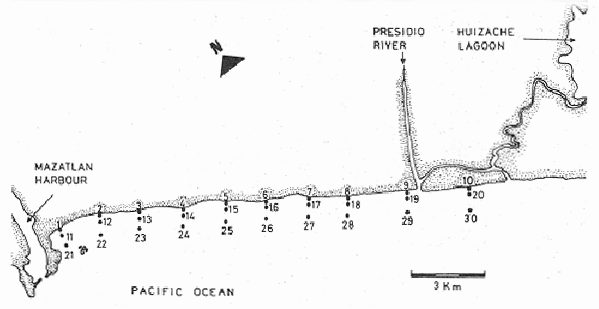 Figure 3. Diagram of the area between Mazatlan an the Caimanero-Huizache lagoon complex showing the plankton sampling sites. Stations 110 were made in the longshore current (zone LC); Stations 11-20, near the breaker zone (NB) and Stations 21-30 offshore, at the 10 m isobath (0). Penaeus postlarvae were identified to specific level by means of keys developed by Mair (197%) and Cabrera Jiménez (1983). The coastal water motion pattern near the Baluarte river mouth at ebb and flood tides was followed by means of oranges used as drifters, which were released at different sites. Each orange had six axes with plastic tags on the tips to ease detection. The adventage of orange over conventional floats is that the former have a slightly negative bouyancy, thus direct wind effect is minimal. Longshore current speed was calculated measuring the time lapse of the passage of drifters between a 50 m distance. Wind speed and its direction were obtained from the Meteorological Service of Mazatlán International Airport. Salinity and temperature data from the area of study for the same period are given by Poli and Calderón-Pérez (1987). RESULTS
WINDThe distribution of the times at which the wind started and ended blowing is shown in Table 1. These figures are percentages of the original data provided by the Meteorological Service of Mazatlán International Airport from May to November 1981. 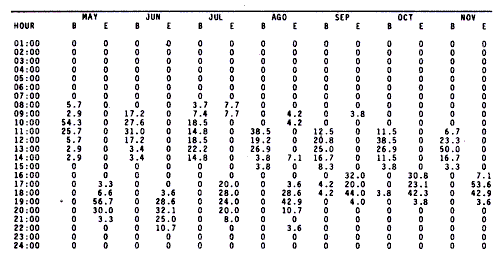 TABLE 1 PREVALENCE RATE (%) OF TIME AT WITCH THE WIND The prevalent starting time was between 10:00 and 13:00 hr while the prevalent ending time was between 18:00 and 21:00 hr. Troughout the whole period of study, wind speed was negligible between 00:00 and 07:00 hr. Total wind duration, in hours, and its direction is represented schematically for the period January 1981 to January 1982 in figure 4, only wind speed of 2.5 to 6.1 m/sec (5 to 12 knots) were considered. 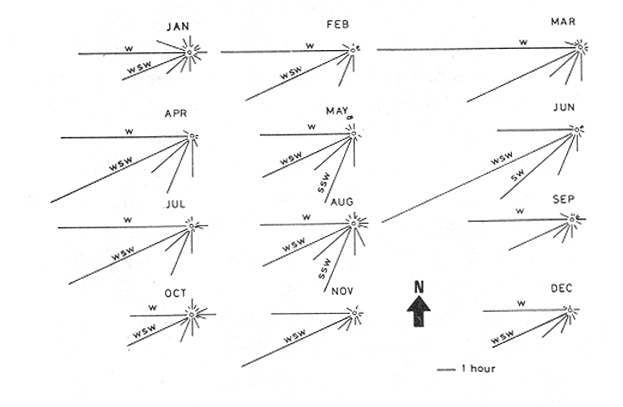 Figure 4. Schematic representation of monthly wind direction prevalence and total duration throughout the year 1981. Only speeds of 2-5 to 6.1 m/sec. (S to 12 Knots) were considered. LONGSHORE CURRENTSpeed values and direction of the longshore current from 17 May 1981 to 9 January 1982 are shown in Table 2. During the period which included the months between December and June, the prevalent direction of the longshore current was SE whereas from July to November it was observed to be the opposite;i.e. NW. Speed values ranged from 0.05 to 0.09 m/sec. The path of all drifters from their release site is also, included in Table 2. It was generally observed that the longshore current speed slowed down during ebb tide on the Nortwestern side of the river mouth when its direction was SE whilst on the Southeastern side its speed was not affected. 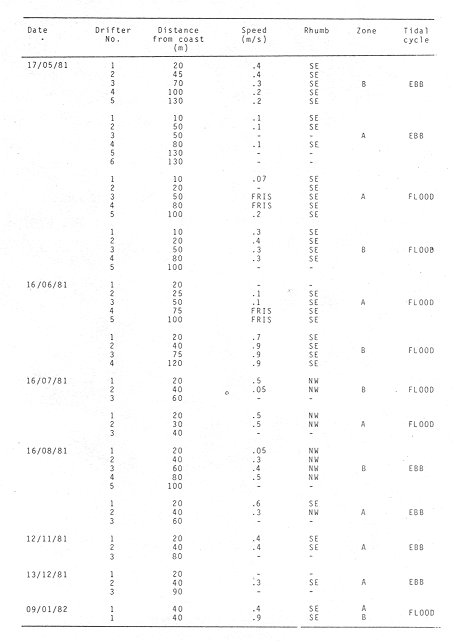 TABLE 2 RHUMB SPEED OF THE LONG SHORE CURRENT MEASURED BY MEANS OF DRIFTERS RELEASED AT DIFERENT DISTANCES FROM THE COAST. ZONE A IS LOCATED ON THE SOUTH EASTERN SIDE OF BALUARTE RIVER MOUTH; ZONE B ON THE NORTH WESTERN SIDE. FRIS = FLOATER RECOVERED INSIDE THE SYSTEM A schematic view of the water motion pattern near the Baluarte river mouth at both flood and ebb tides when the longshore current direction was SE is show in figures 5A and B. At ebb tide (Fig. 5A), floats released on the NW side of the mouth moved on a circular pattern (floats 2, 3 and 4) or went: ashore (float 1) whereas those released on the SE side moved on that direction and eventually went: ashore. A different picture was observed at flood tide on the NW side (Fig. 5B); floats 4 and 5 crossed the river mouth into the system, floats 2 and 3 probably did the same but got lost. The only float which went ashore was number 1. On the SE side, the path of all floats was roughly the same as the observed at ebb tide. 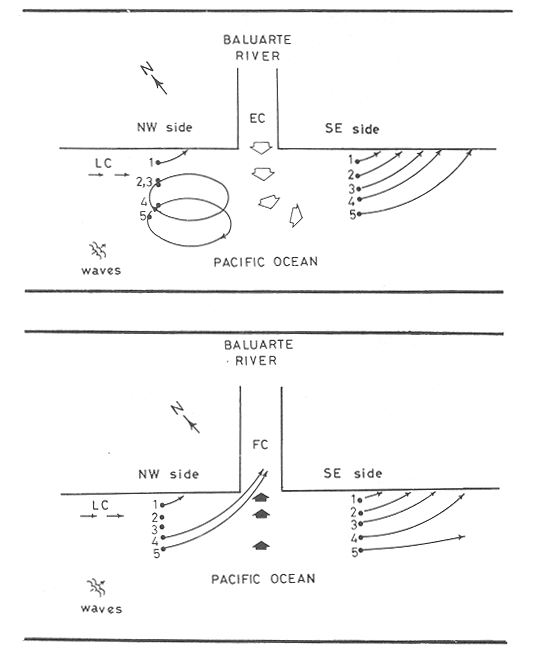 Figure 5. Observed trajectories of floats released at different sites on both sides of the Baluarte river mouth. A, at ebb tide; B, at flood tide. LC, longshore current; EC, ebb current; FC, flood current. POSTLARVAL DENSITY AT SEAPostlarval density values at sea in the three zones indicated in figure 3 are displayed in Table 3 where an index of zooplankton abundance is also included. These results suggest that the trend of both postlarval density and zooplankton biomass is to increase towards the shore line. To test if the differences observed between the three zones considered were significant, an analysis of variance was performed on the loge. postlarval density + 1 values. These results are shown on the upper half of Table 4. Since the test yielded a significant result,i.e. U1 = U2 = U3, a Student-Newman-Keuls test was applied in order to find out the significance of the difference among the means. The results which are displayed on the bottom half of Table 4 show that the postlarval density mean value of the near-breaker zone is significantly higher than that found offshore and although is higher than the mean value found in the longshore current the difference was non-significant. The latter was also higher than the mean value found offshore but again, the difference was non-significant in spite of the fact that the former was 10.5 times bigger than the latter. The explanation for this result lies in the fact that the standard error is very high. 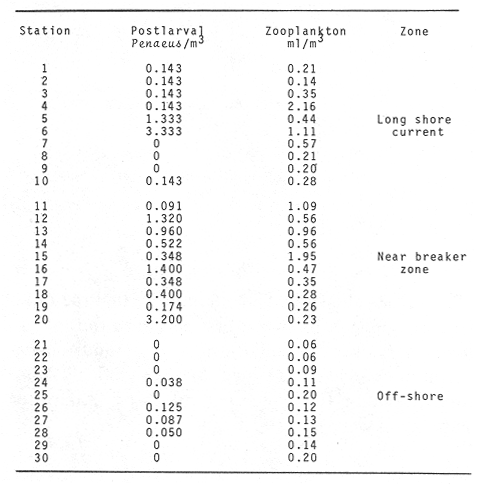 TABLE 3 POSTLARVAL PENAEUS DENSITY AND ZOOPLANKTON BIOMASS FOUNDD AT SEA, IN THE STATION SHOW IN FIGURE 3 POSTLARVAL DENSITY VALUES AT THE BALUARTE RIVER MOUTHIn order to test the hypothesis thatPenaeuspostlarvae accumulate near the river mouth as a result of the longshore current transport being interrupted by the tidal ebb current, all postlarval density data for each species were grouped into three intervals of flood duration. The over-all means are set out in Table 5 in which it is shown that the highest values for all species were observed during the first. third of tidal flood and the lowest during the third, except forP. vannamei. Since the variance values were extremely high and heterogeneous a Mann-Whitney test was applied to the raw data in order to find out whether the differences were significant. The significance of the resulting U values, which are not available in the critical values table for ni and/or n2 greater than 20, was determined by computing the test statistic Z which was compared to the critical value t0.05 (2) = 1.960 (Zar, 1974).  TABLE 4 RESULTS FROM THE ANALYSIS OF VARIANCE AND SNK TEST APPLIED TO THE POSTLARVAL DENSITY DATA SET OUT IN TABLE 3. ALL DENSITY + 1 FIGURES WERE TRANSFORMED TO LN VALUES. LC = LONG SHORE CURRENT, NB = BREAKER, O, OFF SHORE. The results shown in Table 6 indicate that the difference is significant between first and third thirds inP. vannamei, P. stylirostris and P. californiensis and non-significant in P. brevirostris which show a significant difference between first and second third. All other comparisons were non- significant. 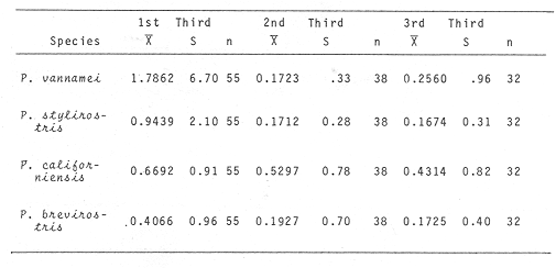 TABLE 5 MEAN POSTLARVAL DENSITY (X) AND STANDARD DEVIATION (S) OF THE FOUR SPECIES OF PENAEUS FOR EACH THIRD OF THE FULL MOON TIDAL FLOOD DURING THE PERIOD BETWEN JANUARY 1981 AND JANUARY 1982. n = NUMBER OF SAMPLES 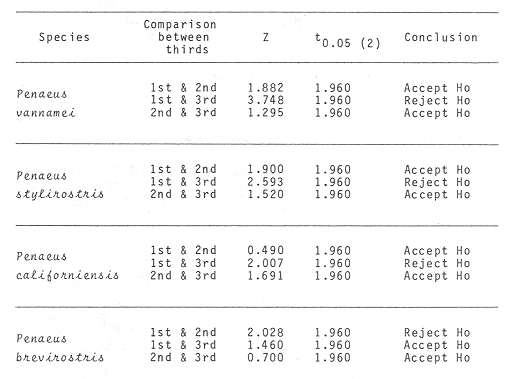 TABLE 6 RESULTS FROM THE MANN-WHITNEY TEST PERFORMED ON THE FULL MOON PERIOD POSTLARVAL DENSITY DATA FOR EACH SPECIES OF PENAEUS GROUPED INTO THREE INTERVALS OF TIDAL FLOOD DURATION FROM JANUARY 1981 TO JANUARY 1982. DISCUSSIONIt is generally accepted that penaeid larvae are weak swimmers and their movements are negligible when compared to the velocity of ocean currents, wind drift and tides (Penn, 1975). On the other hand, Aldrichet al. (1968) estimated a swimming speed of 4.8 km/day for postarvalPenaeus aztecusand P. setiferusin static waters. Based on this figures, Mair (1979b) suggested that it is possible for postlarvalPenaeus in the sea to reach the Baluarte river mouth from the spawning grounds in a short period of time if swimming was orientated to a certain stimulus of which the river mouth would be the source. This elaboration could account for the gathering of postlarvae in the vecinity of the river mouth if water motion were not involved and even so, it fails to explain the eventual immigration to the lagoon itself for the stimulus would not come from that source but from the river. Nevertheless,Penaeuspostlarvae do select waters of low salinity and are able to distinguishestero water (Mair, 197%, 1980; Mairet al.,1982). Salinity has frequently been mentioned as the most likely stimulus to orientate postlarvae to coastal lagoons or towards the shore (Edwards, 1978; Hughes, 1969; Keiser and Aldrich, 1973; Young and Carpenter, 1977). This stimulus-orientated migration mechanism could explain the large postlarval densities found at the access points of the system in July, August, September and October (Calderón-Pérez, 1977; Mair et al., 1982; Poli and Calderón-Pérez, 1987) since the river influence in the sea extends considerably as the rainy season progresses. Unfortunately, this mechanism is inadequate to explain the immigration detected during the dry season when the river discharge is at its lowest level, especially in June (Poli and Calderón-Pérez, 1987). Thus if the migration mechanism is not stimulus-orientated, an alternative explanation must be sought. Wind drift and tides could account for the larval transport onshore if the vertical migration pattern makes possible the coincidence of the larvae being at the surface while the top water layers travel towards the coast. Vertical diurnal migration, common to many decapod larvae, has been observed in Penaeus duorarum(Joneset al., 1970; Temple and Fischer, 1965);P, plebejus (Racek, 1959) and inferred indirectly inP. latisculatus (Penn, 1975). Although it has not been demostrated in the Penaeus specie on the Mexican Pacific Ocean, its like lihood is suggested by Mair et al. (1982) and Watkins (1980), who found that postlarvae are more active during the night. In general, throughout the afternoon and early night the wind always blew onshore, from the west and the southwest; hence considering the 45° deviation to the right of the wind-induced Ekman currents in relation to the wind direction (Stevenson, 1970; Sverdrup et al., 1942), the larval transport onshore would be facilitated in the latter case and slight1 y hindered in the former. Offshore current at the surface runs parallel to the shore; i. e. southerly from. January through April and northerly from. August to December; in June its direction is variable (Stevenson, 1970). Throughout the period of study the wind blew mostly from the southeast, thus is would be expected that postlarvae moved predominantly onshore. If the wind blows from the northwest, it would he expected that postlarvae at the surface should be carried offshore in order to comply with this hypothesis. Recently, Calderón-Pérez et al. (in preparation) sampled under the conditions described above, producing precisely those results: more postlarvae at the surface were found offshore; as far as 18 km from the coast, than onshore. Combining the effects of wind drift and tidal flood, postlarval transport onshore could be explained in those terms, albeit, the vertical migration mechanism must be coherent with those factors and that has yet to be proved. Discernment of tides through changes in hydrostatic pressure seems unlikely in the open sea (Mair et al., 1982; Penn, 1975), thus, changes in light intensity must be responsible for the larval vertical migration. As a results of this transport onshore, postlarvalPenaeus densities were higher just off the breaker zone and inside the longshore current zone, as shown in the previous section, and although the highest values were not found in the latter, a considerable amount of postlarvae were found within. Mair (1980) observed more postlarvalPenaeus onshore than offshore. The results of the comparison between intervals of flood lapse, suggest that, throughout the tidal ebb, postlarvae are being accumulated near the river mouth. This accumulation process could be explained as follow: Postlarvae reaching beyond the breaker zone will be transported by the longshore current until a rip current or an obstacle interrupts the flow; such an obstacle could be the hydrodynamic barrier caused by the tidal ebb current. Since the surface water was observed to describe a loop-like motion pattern on the side of the river mouth where the longshore current was moving from, a similar path for postlarvae seems logical. At the start of the flood tide all those postlarvae would pass through the mouth into the system, in a similar fashion to that of the floats released at different sites in the vecinity of the river mouth. Postlarval accumulation during ebb tide has also been observed in Penaeus plebejus (Young and Carpenter, 1977) at the Nerang river mouth and in the Mexican Pacific Penaeus spp at the Presídio river mouth (Calderón-Pérezet al., in preparation). The physical approach to the postlarval horizontal migration mechanism does not invalid the behavioural alternative since their response to changes in environmental factors has been observed under experimental conditions, as seen earlier. Furthermore, if postlarval accumulation and immigration were merely passive phenomena, other penaeid larvae which are also present in some areas where larvalPenaeus occur (Calderón-Pérezet al., in preparation) those larvae should be found inside the estuarine system. Penaeid larvae other thanPenaeus has never been reported inside the Caimanero-Huizache lagoon complex. Thus, it is possible, that the migration process and eventual entry into the lagoons is due to the concourse of both physical and behavioural factors, each having alternating relevant roles, at different stages of the whole process. LITERATURAAYALA-CASTAÑARES, A., M. GUTIÉRREZ and V.MALPICA In: Informe Final de los Trabajos Contratados en los Planes Piloto Escuinapa-Yavaros.Informe final de los estudios de geología marina en la regiones de Yavaros, Son., Huizache y Caimanero, Sin. y Agiabampo, Sin., durante la primera etapa.Inst. Biol. Univ. Nal. Autón. México y Sría. Rec. Hidraul.1970.3-190. ALDRICIH, D. V., C. E. WOOD and Y- N. BAXTER Bull. Mar. Sci. An ecological interpretation of low temperature responses in Penaeus aztecus and P. setiferus postlarvae.1968.61-71.18 CABRERA JIMENEZ, J. A. CrustaceanaCharacters of taxonomic value of the postlarvae of the shrimp Penaeus (Farfantepenaeus) brevirostris Kingsley (Decapoda: Natantia) of the Gulf of California, México.1983.292-300.44(3): CALDERÓN-PÉREZ, J. A. Tesis Prof. Biología. Efecto de algunos factores físicos sobre la inmigración de postlarvas de Penaeus en el Estero de Agua Dulce del sistema lagunar de Huizache-Caimanero, Sinaloa.Fac. Ciencias. Univ. Nal. Autón. México.1977.127 p. CARDENAS FIGUEROA, M.Tesis Prof. Contribución al conocimiento de la biología de los peneidos del noroeste de México. Esc. Nal. Cienc. Biol. Inst. Pol. Nal.1950.76 p. CHAPA-SALDAÑA, II. and R. SOTO LÓPEZ Lagunas Costeras un Simposio. Mem. Simp. Inter. Lagunas Costeras.Relación de algunos factores ecológicos en la producción camaronera de las lagunas litorales del sur de Sinaloa. In: Ayala-Castañares, A. y F. B Phleger (Eds.)UNAM-UNESCO:D.F.México1969.653-662.28-30 Nov. 1967. COLMAN, J. S. and F. SEGROVE J. Anim. Ecol.The tidal plankton over stoupe Beck Sands, Robin Hood's Bay (Yorshire, North Riding)1955.445-462.24(2) EDWARDS, R.R.C. Estuar. Coastal Mar. Sci. Field experiments on growth and mortality of Penaeus vannamei in a Mexican coastal lagoon complex.1977.107-121.5(1): EDWARDS, R.R.C. Oceanogr. Mar. Biol. Ann. Rev.The fishery and fisheries biology of penaeid shrimp on the Pacific coast of México.1978.145-180.16 HUGHES, D. A. Fish. Rep. On the mechanisms underlying tide-associated movements of Penaeus duorarum Burkenroad.FAO1969.867-874.57(3): JONES, A. C., D.E. DIMITRION, J-J. EWALD and J.H. TWEEDY Bull. Mar. Ser. Distribution of early development stages of pink shrimp Penaeus duorarum in Florida waters.1970634-661.20 KEISER, R- K. and D. V. ALDRICH Contr. Mar. Sci. A gradient apparatus for the study of salinity preference of small benthic and free swimming organisms.1973.153-162.17: KENNEDY, F. S. and D. G. BARBER J. Crustacean Biol.Spawning and recruitment of pink shrimp Penaeus duorarum off Eastern Florida.1981.474-485.1(4) LANKFORD, R. R. Estuarine Processes. II.Coastal lagoons of México - Their origin and clasiffication. In: Wiley, M. (Ed.)Academic Press.New York1977.182-215. LINDNER, M. J. and W. W. ANDERSON U.S. Wildl. Serv. Fish. Bull. Growth, migrations, spawning and size distribution of shrimp Penaeus setiferus.1956.555-645.56 MACÍAS-REGALADO, E. An. Inst. Cienc. del Mar y Limnol.(en prensa).La migración de postlarvas del género Penaeus del mar a una laguna costera en el oceáno Pacífico (Crustacea: Decapoda: Penaeidae).Univ. Nal. Autón. México. MACÍAS-REGALADO, E. and J. A. CALDERÓN. PÉREZ An. Centro Cienc. del Mar y Limnol. Talla de inmigración de postlarvas en el sistema lagunar de Huizache-Caimanero, Sinaloa, México (Crustacea: Decapoda: Penaeus) Univ. Nal. Autón. México1979.99-106.6(2): MACÍAS-REGALADO, E. and J. A. CALDERÓN-PÉREZ An. Centro Cienc. del Mar y Limnol. Influencia de los "tapos" (artes de pesca fijas) en la migración de postlarvas al sistema lagunar Huizache-Caimanero, Sinaloa, México. (Crustacea: Decápoda: Penaeus).Univ. Nal. Autón. México1980.39-507(1): MACÍAS-REGALADO, E., H. FERNÁNDEZ PÉREZ and A. CALDERÓN-PÉREZ An. Inst. Cienc. del Mar y Limnol.Variación diurna de la densidad de postlarvas de camarón en la boca del sistema lagunar Huizache-Caimanero, Sinaloa, México. (Crustacea: Decápoda: Penaeidae). Univ. Nal. Autón. México1982.381-386.9(1) MAIR, J. Mc D. J. Zool. Lond. The identification of postlarvae of four species of Penaeus (Crustacea: Decapoda) from the Pacific coast of México1979a.347-351.188: MAIR, J. Mc D. Ph. D. Thesis.Identification and behaviour of postlarval penaeid shrimp from West México.Univ. Liverpool1979b93p. MAIR, J. Mc D. J. Exp. Mar. Biol. Ecol. Salinity and water type preferences of four species of postlarval shrimp (Penaeus) from West México.1980.69-82.45 MAIR, J. Mc D., J. L. WATKINS and D. I. WILLIAMSON Oceanologica Acta Factors affecting the immigration of postlarval penaeid shrimp into a Mexican lagoon system1982.339-345.Vol. Spec., 5 MENZ, A. and A. B. BOWERS Estuar. Coastal Mar. Sci.Bionomics of Penaeus vannamei Boone and Penaeus stylirostris Stimpson in a lagoon on the Mexican Pacific coast.1980.685-69710: NOOR, S. H. Bull. Shrimp Cult. Res. Center JeparaThe occurence and abundance of shrimp fry at Jepara in 1975 and 1976.1976.158-168.2: PEARSON, J. C. Bull. Bur. Fish. Wash. The early life histories of some American Penaeid. Chiefly the commercial shrimp Penaeus setiferus (Linn.).1939.1-73.49(30) PENN, J. W. Aust. J. Mar. Freshwater Res. The influence of tidal cycles on the distributional pathway of Penaeus latisulcatus Kishinouge in Shark Bay, Western Australia.1975.93-102.26 POLI, C. R. and J. A. CALDERÓN-PÉREZ An. Inst. Cienc. del Mar y Limnol. Efecto de los cambios hidrológicos en la boca del Río Baluarte sobre la inmigración de postlarvas de Penaeus vannnamei Boone y P. stylirostris Stimpson al sistema lagunar Huizache-Caimanero Sinaloa, México. (Crustacea: Decápoda: Penaeidae). Univ. Nal. Autón. México1987.29-44.14(1) RACEK, A. A. Res. Bull. St. Fish. N.S.W. Prawn investigations in Eastern Australia.19591-57.6 RODRÍGUEZ de la CRUZ, M. C. Mem. Simp. Biología y Dinámica Poblacional de Camarones. Distribución de los estados larvales y postlarvales de los géneros de la familia Penaeidae en la parte central y norte del Golfo de California, México.INPGuaymas, Son.México1976.317-350.1:8-13 Agosto 1976. ROESSLER, M. A. and R. G. REHRER Bull. Mar. Sci.Relation of catches of postlarval pink shrimp in Everglades National Park, Florida, to commercial catches on the Tortugas grounds.1971.791-805.24: SOTO LÓPEZ R. Tesis Prof. Mecanismo hidrológico del sistema de lagunas costeras Huizache-Caimanero y su influencia sobre la producción camaronera.Univ. Autón. B.C.1969.75 p. STEVENSON, M. R. Inter-Amer. Trop. Tuna Comm. Bull. On the physical and biological oceanography near the entrance of the Gulf of California, October 1966 August 1961.1970.389-504.14(3): SVERDRUP, H. U., M. W. JOHNSON and R. H. FLEMMING The Oceans. Their Physics, Chemistry and General Biology.Prentice-HallEnglewood cliffs.1942.1087 p. TEMPLE, R. F. and C. C. FISCHER Publs. Inst. Mar. Sci.Vertical distribution of planktonic stages of penaeid shrimp. Univ. Texas.1965.59-67.10: WARBURTON, K Estuar. Coastal Mar. Sci.Community structure, abundance and diversity of fish in a Mexican coastal lagoon system.1978.497-319.7 WATKINS, J. L. Ph. D. Thesis. The immigration of postlarval penaeid shrimp into a lagoon system on the Pacific coast of México.Univ. Liverpool.1980.99 p. YOUNG, P. C. and S. M. CARPENTER Aust. J. Mar. Freshwater Res. Recruitment of postlarval penaeid prawns to nursery areas in Moreton Bay, Gueensland1977.745-793.28: ZAR, J. II. Biostatistical Analysis.Prentice-Hall Inc.New JerseyUSA.1974.620 p.
|