|
DISTRIBUCIÓN ESPACIAL DE LA FLORA Y
VEGETACIÓN ACUÁTICA Y PALUSTRE DEL ESTERO MARGA-MARGA EN CHILE
CENTRAL
Trabajo recibido el 26 de agosto de 1985 y aceptado para su
publicación el 22 de abril de
1986
B. Palma
C. San Martín
M. Rosales
L. Zúñiga
Instituto de Biología
Universidad Católica de Valparaiso, casilla 4059, Valparaiso,
Chile.
C. Ramírez
Instituto de Botánica, Universidad
Austral de Chile, casilla 567, Valdivia,
Chile
Se
estudiaron los patrones de distribución de la flora y vegetación
acuática y palustre a lo largo del curso del estero Marga-Marga, en la
provincia de Valparaíso, Quinta Región de Chile. Esta
distribución espacial se relacionó con las variables
abióticas ambientales. El estudio se realizó en la temporada
verano-otoño de 1981. Se determinaron 41 especies de cormófitos
acuáticos, siendoElodea
potamogetoneHydrocotyle ranunculoides las
más abundantes. La distribución de las especies permitió
establecer dos grupos vegetacionales: uno en la cabecera del estero y el otro,
en los sectores medio e inferior, los que están determinados
principalmente por la naturaleza del sustrato y las variables
físico-químicas del agua, especialmente salinidad y
eutroficación.
The
distribution patterns of the flora and aquatic as well as marshy vegetation
along the Marga-Marga stream in the province of Valparaíso, v
Región, Chile were studied during the 1981 summer-autumn season. This
spatial distribution was related to the environmental abiotic variables. Forty
one aquatic cormophytes were found, being Elodea
potamogetonandHydrocotyle ranunculoides the most
abundant. The distribution of the species separated two vegetational groups:
one at the source of the stream an the other in the middle and lower areas of
the course. These groups are mainly defined by the nature of the substrate and
by the physicochemical variables, specially salinity and eutrophication of the
water.
En la región de Valparaíso, desde la Cordillera de la Costa, nacen una serie de pequeños ríos, de corto recorrido, que se conocen localmente comoesteros (Palma et al.,1978). En sus riberas se han desarrollado ciudades que vierten sus desechos en ellos, contribuyendo a la eutroficación de sus aguas (Sáizet al.,1980). Uno de estos cursos limnéticos, el estero Marga-Marga, recibe además, acción antrópica directa en su parte superior como consecuencia de la búsqueda de oro y en la parte intermedia, por el uso para fines turísticos-recreacionales. En su curso inferior está sometido a la influencia de las aguas salinas del Océano Pacífico. La vegetación acuática y palustre de estos ambientes lóticos es muy rica en especies y en formas de vida (Ramírezet al., 1979). Sin embargo, ella está sujeta a grandes fluctuaciones estacionales debido a los cambios físicos que experimentan los cauces (Palmaet al.,1978) y a los cambios químicos provocados por la eutroficación del agua (Campos, 1979). Efectivamente, durante el invierno y primavera, el aumento del caudal primero por lluvias y luego por derretimiento de las nieves, es muy acentuado, provocando ocasionalmente, grandes crecidas e inundaciones, de consecuencias catastróficas para la población. Por el contrario, en verano y otoño, el caudal permanece estable permitiendo el desarrollo de una abundante vegetación acuática y palustre. Los cambios estacionales en la vegetación hidrófila de estos ambientes han sido descritos por Ramírezet al.(1985). El presente estudio analiza la distribución espacial de la vegetación acuática y palustre a lo largo del curso del estero Marga-Marga (Valparaíso, Chile), en la época de estabilidad estival y sus relaciones con los factores abióticos ambientales. MATERIAL Y MÉTODOSEl estero Marga-Marga se ubica en la provincia de Valparaíso, Quinta Región de la actual división político-administrativa de Chile (Fig. l). Se origina en la zona de Colliguay (Quilpué), por la confluencia de una serie de cursos de agua que surgen de los macizos de la cordillera costera (Jaffuel y Pirion, 1921) y después de un recorrido cercano a los 30 km recibe, en su tramo final, la descarga de aguas servidas urbanas e industriales, provenientes de las ciudades de Peñablanca, Villa Alemana y Quilpué. Su caudal experimenta grandes fluctuaciones estacionales y en invierno, por efecto de las lluvias corre torrentoso, arrastrándo la vegetación que lo coloniza. El muestreo se realizó mensualmente entre diciembre y abril de 1981 (verano-otoño), época que corresponde a la mayor estabilidad del cauce y al apogeo de la vegetación hidrófila. Por su corto trayecto no es posible sectorizarlo de acuerdo al esquema propuesto por Illies (1961), ya que prácticamente todo su curso corresponde a un rithrón con vegetación acuática y palustre, concentrada en remansos. El diseño de muestreo consideré la accesibilidad, y las zonas de posibles variaciones en las características ambientales, estableciéndose en total 8 estaciones (Fig. l). En cada una se consideró para la evaluación biológica, un área de 100 m de largo, dividida en 25 cuadrantes de 4 m por todo lo ancho del cauce, muestreándose al azar el 50 % de ellos, mientras que, para la ambiental se midió la temperatura superficial del agua, se determinó la salinidad mediante un salinómetro YSI 33 SCT, el pH con potenciómetro portatil ORION 201 y la velocidad de la corriente mediante el método descrito por Gessner (1950), considerando el promedio de varias mediciones. Para la clasificación de las aguas de acuerdo a su salinidad se aplicó el sistema de Venecia (Arenas, 1971). El sedimento fue analizado tanto en su composición granulométrica por cernido en seco utilizando tamices (Reinnarth, 1978; Bertrán, 1984), como en su contenido de materia orgánica, por consumo de permanganato (Arenas et al.,1980). 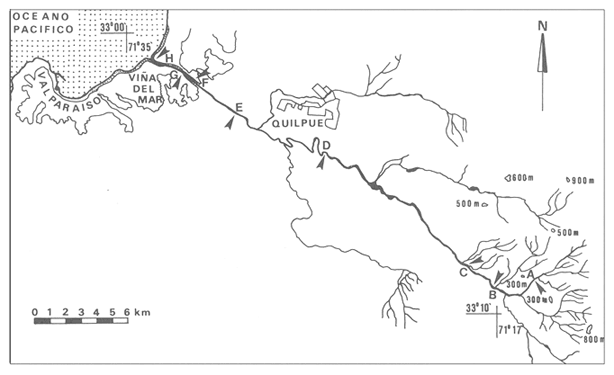 Figura 1. Curso del estero Marga-Marga en Valparaíso, Chile. Las puntas de flecha señalan los lugares de muestreo: A = Los Colihues, B = Los Perales, C = La Retuca, D = Poza larga, E = El Olivar, F = Puentes Miraflores y Lusitania, G = Puentes Cancha y Mercado, H Puente Casino - Barra. Para la vegetación se utilizó metodología no extractiva, identificando en terreno las especies y recolectando sólo aquellas que presentaban dificultades en su identificación. La taxonomía se determinó de acuerdo a los trabajos de Ramírez et al. (1976, 1979, 1980, 1982) y las formas de vida según el esquema propuesto por Ramírez y Stegmaier (1982). La diversidad de géneros se calculó según el índice de Shanon-Weaver (1964). La frecuencia absoluta de las especies presentes en cada estación se resume en una tabla de presencia/ausencia. Las estimaciones del grado de similitud taxonómica y taxocenótica se realizaron utilizando los índices de Cole y de correlación por rango de Spearman (Gounot, 1969). El primero considera solo la presencia/ausencia y el segundo toma en cuenta la abundancia relativa de cada especie. RESULTADOS Y DISCUSIÓNAl analizar las mediciones obtenidas de las variables ambientales, se observa que aún cuando el pH desciende hacia la desembocadura, se conserva levemente alcalino en todo el cauce. Por el contrario, la temperatura se incrementa en esta misma dirección (Tabla l). Sin embargo, la salinidad, en un comienzo baja y constante, va aumentando, hasta tener su máximo en el último lugar de muestreo. Según este parámetro las tres primeras estaciones son de aguas limnéticas, las cuatro siguientes, mixooligohalinas y la última mixomesohalina. La velocidad de la corriente es fluctuante debido a la conformación del cauce del estero. Los valores bajos se asocian con caudal escaso y la formación de remansos ribereños. Los mayores de determinaron en lugares donde el estero ha sido canalizado por el hombre, en la ciudad de Viña del Mar. 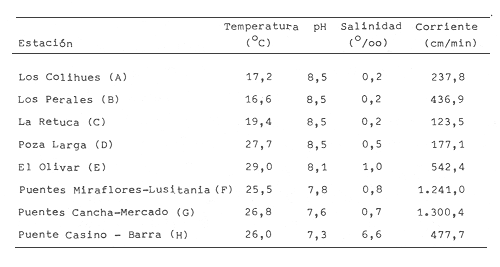 TABLA 1 VALORES PROMEDIO DE LAS VARIABLES ABIÓTICAS DEL AGUA EN LAS DIFERENTES ESTACIONES DE MUESTREO EN EL ESTERO MARGA-MARGA La figura 2 muestra los resultados del análisis granulométrico del sedimento en la ribera y en el centro del cauce, en las 8 estaciones. En general se observa un dominio de la fracción arenosa (sobre grava y limo) tanto en la ribera, como en el centro del estero, la que alcanza su máxima expresión en el límite con la barra de arena de la desembocadura, donde se forma una playa (playa Casino, estación H). La proporcionalidad de la grava varía, manteniendo la misma tendencia tanto en la orilla como en el centro, excepto en la estación F, donde probablemente la remoción de tierra para habilitar terrenos planos, causa la alteración. 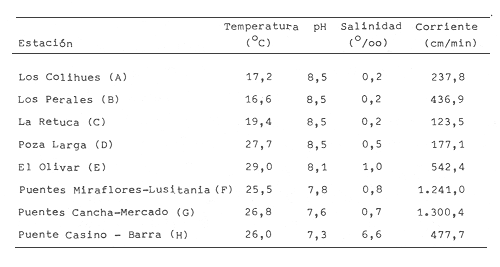 Figura 2. Granulometría del sustrato en la ribera (arriba) y en el centro (abajo) del cauce en los lugares muestreados. 1 = Grava (> de 5 mm), 2 = Arena (S mm a 0.08 mm ). Los valores se expresan en porcentajes. La materia orgánica del sedimento es baja, aunque superior en los dos extremos del transecto analizado. En el primero es consecuencia del pastoreo de animales que se desplazan libremente y en el segundo, por empozamiento y posterior acumulación de los desechos urbanos e industriales que trae el estero (Fig. 3). 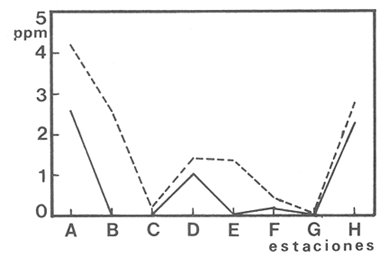 Figura 3. Contenido de materia orgánica en el sedimento del centro (línea cortada) y la ribera (línea continua). Se colectó un total de 41 especies vegetales en los lugares de muestreo (Tabla 2). De éstas, la mayoria (30.73 %) son helófitos emergidos. El resto (27 %) corresponde a plantas acuáticas propiamente. Entre ellas hay 5 sumergidas:Elodea potamogeton, Myriophyllum aquaticum, Potamogeton berteroanus, Ranunculus aquatilis y Zannichellia palustris.Otras cuatro, presentan hojas natantes que flotan sobre la superficie del agua:Hydrocotyle ranunculoides, H. volckmannii, Callitriche stagnalis y Jussiaea repens. Esta última con abundantes neumatóforos. Finalmente, se encontraron dos especies de plantas acuáticas flotantes libres, de pequeño tamaño:Azolla filiculoides y Lemna gibba. Al estudiar la diversidad de géneros de las plantas acuáticas y palustres en el estero Marga-Marga se obtuvo un valor de 5.19 bits/género, lo que concuerda con trabajos previos (Palma, 1971). La distribución de las especies en los lugares de muestreo fue muy heteorgénea (Tabla 3) la mayor frecuencia la presentaron Rumex obtusifolius y Paspalum distichum, que estuvieron presentes en todos los lugares prospectados. Veronica anagallis-aquaticayPolygonum mite solo faltaron en un lugar.Eleocharis bonariensis , Jussiaea repens, Rorippa nasturtium-aquaticum, Hydrocotyle renunculoides, Cyperus altemifolius y Lemna gibbaaparecen en 6 de los sectores investigados. En el resto de las especies las frecuencias disminuyen paulatinamente (Fig. 4). La figura 5 muestra en su parte superior el gradiente de salinidad del agua y en la inferior, la distribución del número de especies y de las formas de vida en los diferentes sectores prospectados. El mayor número de especies se constató en la tercera estación (C), lugar que muestra estabilidad tanto en la naturaleza del sustrato, como también en la corriente, lo que facilita la colonización de él por la flora hidrófila. Posteriormente, hay un rápido descenso hasta la quinta estación (E), para finalmente, en el lugar H encontrar el menor número de especies. La salinidad por un lado y la contaminación por otro, pueden ser causantes de este drástico descenso en la cantidad de plantas acuáticas y palustres que experimenta el estero. La curva del número total de helófitos palustres emergidos sigue un curso similar a la del total de las especies, aunque siempre es más baja. Esto indica que en todos los lugares la vegetación del estero está determinada por esta forma de vida. Las plantas natantes están presentes a lo largo del estero, desde la estación A a la G, faltándo únicamente en el último lugar. Las plantas sumergidas muestran una distribución irregular, que se puede relacionar con los cambios de profundidad, corriente y contaminación del cauce. Esta forma de vida es la mejor indicadora de condiciones abióticas en los ambientes lóticos (Kohler, 1975). Las dos plantas flotantes libres se presentaron en remansos y riberas de todos los lugares, a partir de la segunda estación. A pesar de las cantidad de especies colectadas, solo 6 de ellas (14.6 %) presentaron altos valores de cubrimiento (Fig. 6). Elodea potamogeton alcanzó alta cobertura en el único lugar que se encontró (C). Hydrocotyle ranunculoides se hizo muy abundante en los lugares aledaños a los centros urbanos, confirmando su capacidad para resistir la contaminación. Lemna gibba que presenta alta cobertura en el primer tramo del estero (B y C), desaparece en D, aumentando paulatinamente en el tramo inferior, para declinar en la estación H. Esto concuerda con Pietsch (1972) quien estableció que la frecuencia y abundancia de las Lemnaceae está relacionada con aguas duras, mientras que en aguas ácidas, pobres en carbonatos, son escasas. Sin embargo, hay que tener presente que estas especies flotantes libres, están también limitadas por la corriente, factor muy variable en el estero trabajado. Paspalum distichum yJussiaea repens presentaron su mayor desarrollo en la cuarta estación (D), disminuyendo gradualmente hacia ambos extremos. Dicha estación (Poza Larga) está caracterizada por la presencia de pozas y remansos de profundidad variable. 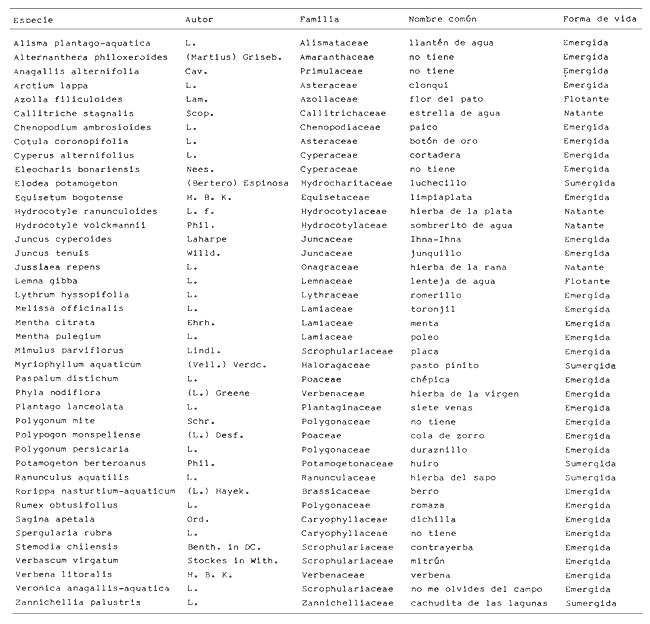 TABLA 2 CARCTERIZACIÓN SISTEMÁTICA, NOMENCLATURAL Y ECOLÓGICA DE LAS ESPECIES DE PLANTAS ACUÁTICAS Y Al comparar la similitud florística entre los distintos sectores, considerando sólo la presencia o ausencia de especies se observa que ellas muestran escasa afinidad entre sí (Fig. 7a). Los grupos formados lo hacen con muy baja similitud. Los sectores D y H presentaron la mayor afinidad, la que se basa en una gran cantidad de ausencias comunes y presencias de especies propias consideradas como raras (Tabla 3). Los lugares E, F y G forman también una unidad taxonómica, basada en presencias comunes y que coincide con aguas de rango mixooligohalino. En estos sectores ocurren en forma constanteArctium lappa y Cotula coronopifolia.Un último conglomerado lo forman las estaciones B y C, ubicadas en la cabeza del estero y las que se unen tambien al primer lugar (A), aunque con una similitud muy baja. Como especies características de esta zona pueden considerarse: Cyperus alternifolius, Eleocharis bonariensis, Rorippa nasturtium-aquaticumyVeronica anagallis-aquatica. Al comparar la similitud vegetacional, es decir, ponderando ahora la abundancia de las especies usando los valores de cubrimiento, se obtiene una mayor similitud entre los lugares, lo que indica que las especies importantes tienen una distribución espacial más amplia (Fig. 7b). En este caso los sectores A, B y C, ubicados en la parte superior del estero forman un conglomerado, separado del resto, al igual que el integrado por E, F y G. Estos dos últimos sectores son los más parecidos entre sí, por la dominancia en ellos de Hydrocotyle ranunculoidesyLemna gibba, especies que indican alto grado de eutroficación. Los lugares D y H permanecen aislados, lo que señala condiciones muy particulares. Estos resultados no permiten establecer un claro gradiente distribucional de las especies a lo largo del curso del estero, presentándose características vegetacionales relativamente diferentes en cada uno de los sectores muestreados, lo que señala una distribución discontínua. 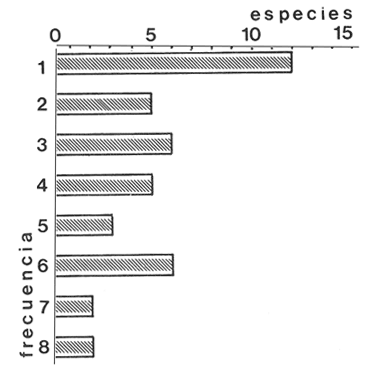 Figura 4. Histograma de frecuencia de las especies acuáticas y palustres en las estaciones muestreadas. 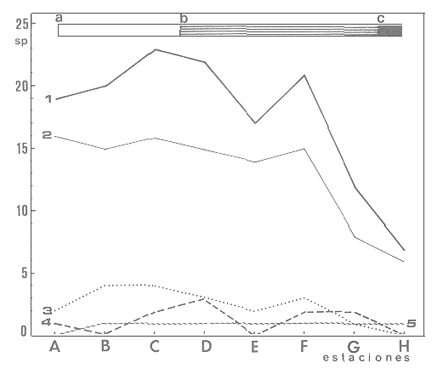 Figura 5. Número total de especies (1), especies emergentes (2), natantes (3), sumergidas (4) y flotantes libres (5) en los diferentes lugares de muestreo. La barra superior indica salinidad M agua: a = limnética, b = mixooligohalinas y c = mixomesohalinas. 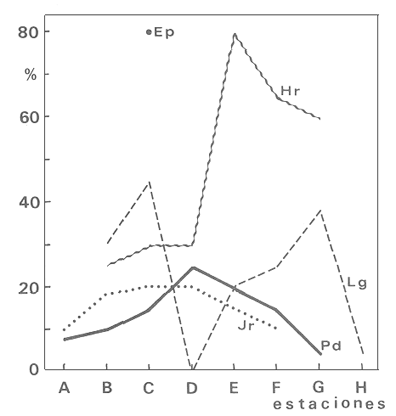 Figura 6. Porcentaje de cobertura alcanzado por las principales especies en los diferentes lugares de muestreo. Ep = Elodea potamogeton, Hr = Hydrocotyle ranunculoides, Lg = Lemma gibba, Jr = Jussiaea repens y Pd = Paspalum distichum.  TABLA 3 PRESENCIA Y FRECUENCIA DE PLANTAS ACUÁTICAS Y PALUSTRES EN LAS DISTINTAS ESTACIONES DE MUESTREO EN EL ESTERO MARGA-MARGA 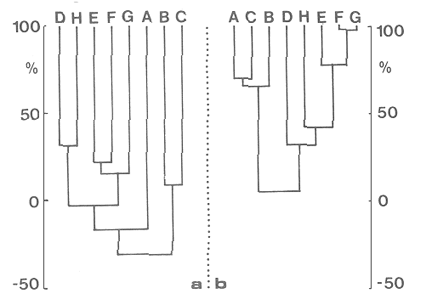 Figura 7. Dendograma de similitud florística (a) y vegetacional (b) entre los diferentes lugares de muestreo indicados en la Fig. Los valores obtenidos en las mediciones de variables ambientales denota variaciones en el espacio, que resultan relativamente coincidentes con las agrupaciones vegetacionales obtenidas fundamentalmente a través del índice de Cole. Los sectores de cabecera (A, B y C) con mayor vegetación, presentan una mayor cantidad de limo y materia orgánica, asimismo sus valores de pH son más altos y las temperaturas medias, la velocidad de la corriente y la salinidad, más bajos. El segundo grupo de sectores (E, F y G), se caracterizan ambientalmente por un bajo contenido de materia orgánica y de limo y un pH más cercano al neutro, siendo en cambio, los valores medios de temperatura, salinidad y velocidad de la corriente, mayores que en el caso anterior, a lo cual se asocia una fuerte contaminación, lo que lleva a una disminución de la vegetación. Los sectores D y H muestran aleatoriedad de condiciones ambientales, por el alto grado de acción antrópica. En el primero existe una constante remoción del terreno por la existencia de lavaderos de oro y en el segundo, se acumulan los desechos urbanos e industriales, además de recibir una fuerte influencia de las aguas saladas del océano Pacífico. La influencia de las condiciones ambientales sobre la vegetación acuática y palustre se puede analizar por el comportamiento de algunas especies individuales.Elodea potamogeton e Hydrocotyle volckmannii ocurren sólo donde los valores de las variables abióticas permiten inferir ausencia de contaminación. En cambio,Hydrocotyle ranunculoides , si bien se presenta en casi todos los sectores estudiados, domina ampliamente en el sector E, industrializado y en el cual aumentan los desechos de tipo urbano e industrial, lo cual corrobora los resultados obtenidos anteriormente por Palma et al. (1978) y por Sáiz et al. (1980). Por otra parte,Cotula coronopifolia y Zannichellia palustris , son especies que se detectan en aguas salobres, coincidiendo con los requerimientos señalados por Ramírezet al. (1976), para estas dos especies. Finalmente, como especie que requiere y soporta salinidad se presentó en el último sector estudiado (H), Alternanthera philoxeroides. Las crecidas periodicas de estos esteros no permiten que la vegetación ejerza su función estabilizadora de sedimentos (Ramírez y Añazco, 1982) y actúe como barrera protectora contra la erosión. Sería importante investigar la acción de esta vegetación acuática sobre la oxigenación y purificación biológica de las aguas, ya que su cultivo podría ser una alternativa para la depuración de los esteros en la zona central de Chile, como se hace en otros lugares (Kohler, 1971, 1973). Este mismo autor distingue dos grupos de hidrófitos sumergidos: los que son inhibidos y desaparecen ante la eutroficación y los que son estimulados por ella (Kohler, 1975). Estos últimos pueden servir como indicadores de contaminación. En el estero estudiado tienen este carácterPotamogeton berteroanus, Ranunculus aquatilisyZannichellia palustris (Kohler, 1975). CONCLUSIONESEl espectro taxonómico de los cormófitos acuáticos y palustres que crecen en el estero Marga-Marga, comprende dos especies de Pteridófitos y 39 plantas superiores. La vegetación estudiada presenta, en general, baja cobertura, siendo Elodea potamogetoneHydrocotyle ranunculoides , las especies más abundantes en los sectores donde aparecen. La composición florística, la abundancia de las especies y algunas variables ambientales controladas, permiten establecer dos grupos de sectores vegetacionales diferentes. Aquellos de cabecera (A, B y C) y los sectores medios e inferiores (E, F y G), correspondientes a aguas más eutróficas. Los sectores D y H presentan aleatoriedad en la composición florística, al ser los de mayor intervención, con un mayor número de especies exclusivas y con una baja cobertura vegetacional. Presentándose en el último lugar condiciones de mayor salinidad. AgradecimientosA las Direcciones de Investigación de las Universidades Católica de Valparaíso y Austral de Chile por el financiamiento a través de los Proyectos 122.729/82 y RSM-80-30, respectivamente y a D. Contreras por su ayuda técnica. LITERATURAARENAS, J.,Distribución de Elminius kingii Gray (Cirr.) en el estuario del Río Valdivia. Beiträge zur Neotropischen Fauna,1971.200-206.6(3): ARENAS, J., P. DOMÍNGUEZ y C. VARELA, Bol. Inf. limnológico, Métodos químicos empleados en estudios limnológicos en Chile.1980.13-63.4: BERTRAN, C., Studies on Neotropical Fauna and Environmental,Macroinfauna intermareal en un estuario del sur de Chile (estuario del Río Lingue, Valdivia).1984.33-46.18(19): BRAUN-BLANQUET, J.,Pflanzensoziologie-Grundzüge der Vegetationskunde.. Springer, Wien.1964.865 p CAMPOS, H., Bol. Inf. Limnológico, El recurso de agua dulce en Chile.1979.6-17.3: GESSNER, F., Die ökologische Bedeutung der Strömungsges-chwindikieit fliessender Gewässer und ihre Messung auf kleinstem Raum.Archiv. für Hydrob.,1950.159-165.43: GOUNOT, M., Masson et Cie., Méthodes d'etude Quantitative de la Végetation.París.1969.204 p. ILLIES, J., Hydrobiol., Versuch einer allgemeinen biozönotischen Gleiderung der Flissgewässer. Int. Revue ges.1961.205-213.46(2): JAFFUEL, F. y A. PIRION, Rev. Chilena de Historia Natural, Plantas fanerógamas del Valle de Marga-Marga.1921.350-405.25: KOHLER, A., Zur Ökologie submerser Gefässmakrophyten in Fliessgewä-ssern. Ber. Dt. Bot. Ges.,1971.713-720.84(11): KOHLER, A., Wasserpflanzen und Gewasserverschmutzung. Der Biologie-unterricht,1973.4-13.9(2): KOHLER, A., Submerse Makrophyten und ihre Gesellschaften als Indikatoren der Gewässerbelastung. Beitr. naturk. Forsch. Südw. Dtl.,1975.149-159.34: PALMA, B., Tesis Depto. de Biología,Contribución al conocimiento de las fanerógamas acuáticas de la región de Valparaíso.Univ. Catól. Valparaíso.1971.54 p. PALMA, B., F. SÁIZ y C. PIZARRO, Anal. Mus. Hist. Nat. de Valparaíso, Variaciones de la taxocenósis de hidrófitas en el curso del estero Limache.1978.61-67.11: PIETSCH, W., Ausgewählte Beispiele für Indikatoreingens-chaften höherer Wasserpflanzen.Arch. Naturschutzu. Landschaftsforsch,1972.121-151.12: RAMÍREZ, C. y N. AÑAZCO, Variaciones estacionales en el desarrollo de Scirpus californicus, Thypa angustifolia y Phragmites communis en pantanos valdivianos, Chile. Agro. Sur,1982.111-123.10(2): RAMÍREZ, C. y E. STEGMAIER, Medio Ambiente, Formas de vida en hidrófitos chilenos.1982.43-54.6(1): RAMÍREZ, C., B. PALMA y C. SAN MARTIN, Variaciones estacionales en la flora y vegetación hidrófila del estero Limache en Chile Central. Phyton. (en prensa).1985. RAMÍREZ C., M. ROMERO y M. RIVEROS, Pub. Ocas. Mus. Nal. Hist. Nat.Lista de cormófitos acuáticos de la región valdiviana.Santiago,1976.3-12.22: RAMÍREZ, C., M. ROMERO y M. RIVEROS, Aquatic BotanyHabit, habitat, origin and geographical distribution of Chilean vascular hydrophytes.1979.241-253.7: RAMÍREZ, C., M. ROMERO y M. RIVEROS, Bol. Mus. Nal. Hist. Nat.Lista de cormófitos palustres de la región valdiviana.Santiago,1980.153-177.37: RAMÍREZ, C., R. GODOY, D. CONTRERAS y E. STEGMAIER, Guía de Plantas Acuáticas y Palustres Valdivianas. Ed. Univ. Austral de Chile,Valdivia.1982.40 p. REINNARTH, G., Eine Methode zur ökologischen Untersuchung der Schlamm-Wasser-Kontaktzone. Verh. Ges. für Ökologie Kiel,1978.401-404.1: SÁIZ, F., P. DOMÍNGUEZ y B. PALMA, Anal. Mus. Hist. Nat. Valparaíso, Ingresos alóctonos, composición química del agua y taxocenósis de hidrófitas en el estero Limache.1980.133-143.13: SHANNON, C. y W. WEAVER,The Mathematical Theory of Communication.Univ. Illinois Press,USA.1964.125 p.
|