|
AGE AND GROWTH OF THE DAB Limanda
limanda (LINNAEUS) OTOLITHS IN ISLE OF MAN WATERS
Trabajo recibido el 4 de
septiembre de 1985 y aceptado para su publicación el 30 de abril de
1986
A.A. Ortega - Salas
Univercidad Nacional Autónoma de México. Instituto
de Ciencias del Mar y Limnología, Ciudad Universitaria, 04510
México, D.F. Contribución 519 de Instituto de Ciencias del Mar y
Limnología, UNAM.
La determinación de la edad
estuvo basada en zonas de verano (mayo - octubre) y anillos de invierno
(noviembre - abril) detectados. En el mes de mayo fue el cumpleaños. El
modelo de von Bertalanffy fue apropiado para estimar tanto la edad de longitud
del otolito como de su ancho en el tiempo t, así que los
parámetros para la longitud del otolito son los siguientes: Hembras:
Loo=6.1197, K=O.39135, to=0.0178566; Machos: Loo=4.9721, K=O.54406,
to=0.222197; y para lo ancho del otolito son los siguientes;: Hembras: Loo, =
3.87925, K = 0.410945, to = 0.02943; Machos: Loo = 3.10006, K = 0.671684, to =
0.27865; donde: Loo: longitud asintótica (mm), K: tasa de
disminución de crecimiento: to: tiempo en que se asume la longitud es
cero. La relación de la longitud y ancho del otolito respecto a la
longitud del pez estuvieron estrechamente relacionadas así que la
regresión funciona] de la longitud del pez al tiempo t - longitud del
otolito al tiempo t, y ancho al tiempo t puede ser escrita como sigue: Hembras:
F.L.t=32.6746 O.L.t 1,2445 Machos: F.L.t = 34.4724
O.L. t1.1944 y Hembras: F.L.t = 45.0917
O.W.t1.4349 Machos: F.L.t = 48.0052 O.W.t
1.3430donde: F.L.t: longitud del pez (mm) en el
tiempo t. O.L.t: longitud del otolito (m) en el tiempo t (obtenida por el
modelo de v.Bffy.) . O.W.t: ancho del otolito (mm) en el tiempo t (obtenido por
el modelo de v.Bffy.) Ecuaciones útiles para estimar la longitud del pez
en el tiempo t de la longitud del otolito en el tiempo t o lo ancho del otolito
en el tiempo t.
Age
determination was based on summer zones (May-October) and winter rings
(November-April detected in the otolith. Maywas assumed to be the birthmont.
The von Bertalanffy's model was appropiated to estimate either otolith length
or width at time t (age), so the parameters for otolith length are the
following: Females: Loo=6.1197, K=O.39135, to=0.0178566; Males: Loo=4.9721,
K=O..54406, to=0.2221970; and for otolith width are the following:; Females:
Loo=3.87925, K=O.410945, to=0.02943; Males: Loo = 3.10006, K = 0.671684, to =
0.27865; where: Loo: asymptotic length (mm), K: rate of decrease of growth;. to
= time assumed to be zero length. The otolith length, and width-fish length
relationship were close related, so the functional relationship fish length at
time t - otolith length at time t, and width at time t may be written as
follows: Females: F.L.t = 32.6746 O.L.t 1.22445
Males: F.L.t = 34.4724 O.L.t 1.1944 and Females:
F.L.t = 45.0917 O.W.t 1.43349Males: F.L.t = 48.0052
O.W.t 1.3430 , where: F.L.t: Fish length (mm) at
time t. O.L.t: Otolith length (mm) at time t. (obtained by the v.Bffy's) O.W.t:
Otolith width (mm) at time t. (obtained by the v.Bffy's.). Useful equations to
estimate fish length at time t from otolith length at time t or otolith width
at time t.
Age determination allows fish to be sorted out into agegps, therefore we are able to count and measure them at age. It is the possible not only to calculate their total mortality, or to calculate the average length and weight of the fish out also to register any other parameter such as stomach weight, liver weight, gonad weight at age (see Ortega-Salas, 1979, 1980, 198la, 1981b, 1981c, 198ld). In fisheries research it is common practice to age tem perate fish by any skeletal structures which show cyclical changes such as spines (Sullivan, 1978), scales (Hofstede, 1974), opercular bones (Fagade, 1954) or Otoliths (Bohl, 1959). The saccular otolith (sacculus) is the only one of three pairs which is important in fisheries research since they can tell us the age of the fish. Graham (1929) and recently Bagenal (1974) reviewed great detail the use of otoliths for age determination. Located at each side of the brain, the inner ear of a fish, its organ of hearing and balance, consists of fluid filled semicircular canals and chambers containing small otoliths (sacculus, utriculus and lagena) which are calcareous deposits. A change in the fish's position is recognized by the organ of equilibrium when the otolith moves. This stimulates reflex movements of fins, which correct the body position (Moller, 1978). After the fish has died, otoliths as calcareous deposits may be preserved for a long time buried into the soil. They can also he stored for long periods in the laboratory for further analysis. Otoliths may also be found during the digestion process in the digestive tract of the predators. All this suggest that otoliths are the remaining of the fish characteristics from which not only may have the information of the species' identification but also information of the age, fish length so weight deduced, and possible rate of fish growth. Therefore, if the rate of growth of the fish can be calculated using fish length at age, it is possible that the otolith length at age could also be calculated, and possible from this calculation to estimate fish growth. Therefore the aims of this paper are the following: 1. Determine age. 2. Calculate growth of the otolith. 3. Calculate the length at time t (age) from otoliths growth. MATERIAL AND METHODSDabs were caught around the coast of the Isle of Man between 54°00' and 54°25'N, 4°55' and 4°10'W in depths from 4 m - 40 m. A few additional samples were taken 20 miles E of the Island at a depth of 20 m - 40 m, and 3 miles SE of the Island between 30 m - 50 m. Monthly samples were taken, weather permitting, from November 1976 to March 1979 by means of an otter trawl with wings of 70 mm mesh size and cod end of 45 mm. Weekly samples of young fish were taken in Port Erin Bay at depths of 4 m- 8 m whenever possible from April 1977 to May 1978, using a beam trawl with mesh size of 60 mm, exterior net, and 15 mm interior lining net. A total of 4,297 dabs from all sources were examined. Otoliths were removed (Williams and Bedford, 1974), rubbed clean between finger and thumb and stored dry. They were subsequently examined in 95% alcohol against a background under low power magnification and the width and length of the otolith (Fig. 1) measured to the neartest 0. 01 mm, using a calibrated eye piece. The average of the measurements of both otoliths either length or width was taken as the final result. The number of opaque zones and hyaline rings were recorded. Records of each fish were maintained as a computer file. The results were processed in the ICI, 1906A Computer from the University of Liverpool and the CDC 7600 from the University of Manchester using the Statistical Package for the Social Sciences version 5 and 6, repectively; and Ortega-Salas (198la, 1981b) Basic program to calculate growth. Scattergram's program from the SPSS system calculated the straight regression line by the least square method, the statistical results are shown below each figure. The scattergrams represented individual data points by mean of an asterisk (*); more than one, in the same position, were marked by the number of fish to a maximum of 9, but all the data were used for the calculations. Scales differed on a given pair of scattergrams for males and females, since the feature of the computer program employed an automatic scale for the data provided. 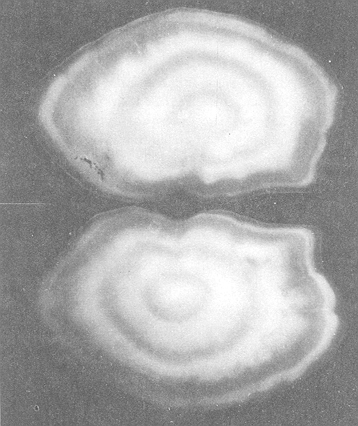 Figure 1. Otoliths of male dab viewed by reflected light against black background showing nucleus, tree summer zones (opaque: white) and four winter rings ( hyaline: dark). Assuming the fish as hatched in May, ege was read as 48 months (4 years old) Fish caught on 31st May 1977. O.L. = 4.72 mm; O.W. = 3.00 mm; F.L.= 203 mm; F.W. = 75 g; Digestive tract whit food = 4.85g; Gonad weight= 0.29g. RESULTS
AGE DETERMINATIONIn the North Sea Dab, Bohl (1959) stated that in otoliths of dabs most of the edges were hyaline between NovemberApril, and between May-October most of the edges were opaque. Also in the North Sea Dab Lee (1972) found a higher percentage of opaque edges from April to September than to the rest of the months which were more hyaline. In Isle of Man waters personal observations Ortega-Salas (1979,1980,198la) showed that in summer (May-October) most of the edges were opaque (white) and in winter (November-April) were hyaline (black) which suggested that opaque zones were mainly deposited in most otoliths between May-October (summer's zones) and the hyaline rings between November-April (winter's rings). Otoliths were, realively, easy to read. A May hatched date was assumed for all dabs and the age was recorded in months which was deduced from the number of opaque (summer) zones and hyaline (winter) rings in the otoliths at month of capture (Fig. l). OTHOLITH GROWTHIn figure 1 can be seen that the otolith growth has been recorded by mean of four zones (including nucleus) which represent four summers and four hyaline rings which represent four winters. The inner opaque zones which represent summer's growth are wider than the subsequent zones and the opaque zones are wider than the hyaline rings. This suggests that the otolith growth is faster in younger fish than older ones and that the growth is greater in summer than in winter. Measurements of the otolith by length and width (Fig. 1) at month of capture and age deduced gave as a result the scattergrams (Figs. 2 and 3) which show the behavior of the population sampled in the same way as fish length does (Ortega-Salas, 198la, 1981b). Most of the population is rePresented between age 2 and 4, younger fish were scarcely caught, probably, because the effect of mesh selectivity and/or they have not recruited into the area sampled. Fish older than 4 years, probably, have died. Eventhough the scattergram show a clear curvilinear relationship from younger to older fish at age. Mean otolith lengths and mean otolith widths were plotted separately for age at two monthly intervals (Figs.4 and 5). The overall curvilinear relationship is clearer. The figures also show at each age-gp that otoliths growth faster from June to October (summer) and slower from November to May (winter). An apparent reduction of the mean measurements (length or width) of the otolith in winter which also is obvious in the scattergrams is, possibly, atributed either to death of the larger fish or their migration away from the sampled area. 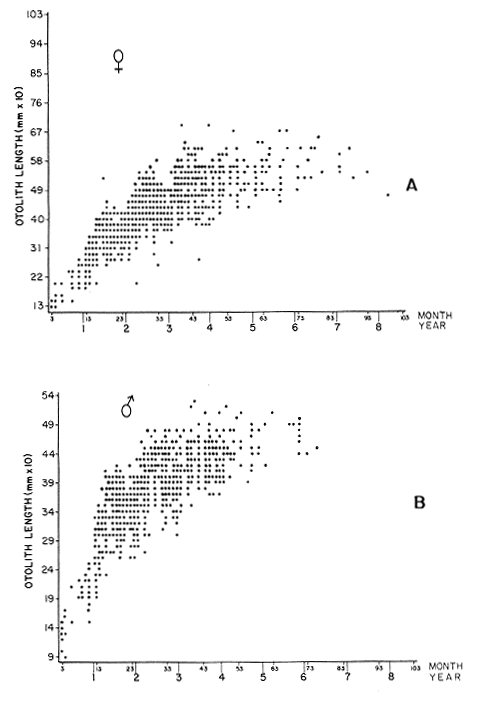 Figure 2. Otolith length for age montly intervals. A) Females B) Males Thus, it can be seen from the graphs (Figs. 4 and 5) that the earlier surnrner's growth periods of greater than the subsequent ones. This was also reflected in the earlier formation of opaque zones (Fig. 1) which were assumed to be formed in the otolith from May to October. 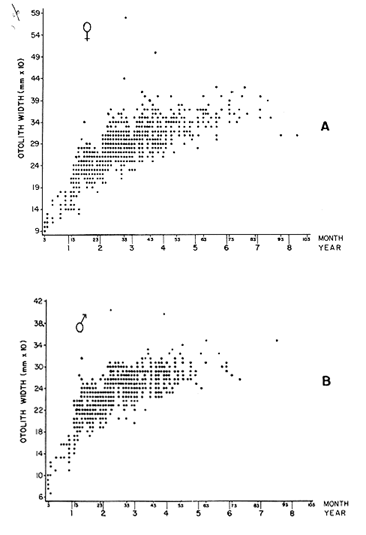 Figure 3. Otolith width for age at monthly intervals. A) Females B) Males It is well known in fisheries literature that scales or otolith dimensions are closely related to fish length, so it would be interesting to see whether the growth of otoliths could be described by the same type of curve as the growth of the fish does. The von Bertalanffy curve (von Bertalanffy, 1951) which is suitable for describing growth of fish (i.e. Berverton and Holt, 1957) could be very useful to calculate the otoliths rate of growth. Then, the von Bertalanffy's equation: 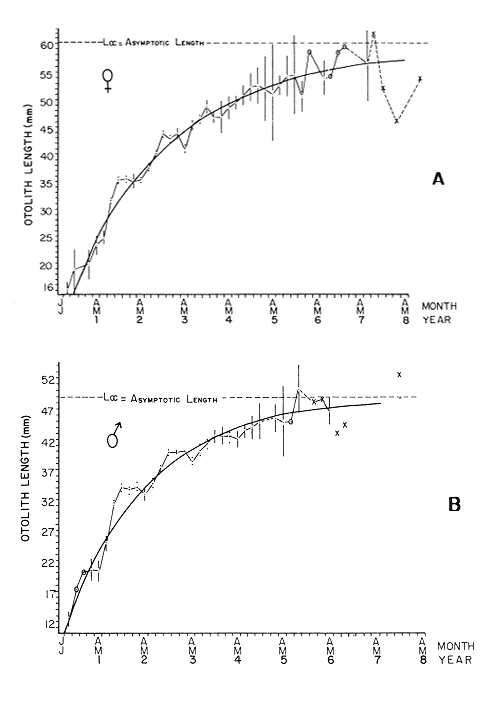 Figure 4. Mean otolith length for age at two monthly intervals with 0.95 C.I. A fitted von Bertalanffy curve is shown. A) Females: La=6.1197, K=O.39135, to=0.0178566. B) Males La = 4.9721, K = 0.54406, to = 0.222197 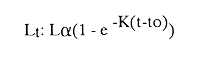 where: Lt: Length (mm) at time t. Lα: Asymptotic or maximum length. e: Base of natural logs. K: Rate of decrease in growth. t: Time in years. to: Time at which the otolith would theoretically have zero length. was used to devise a computer program (Ortega-Salas, 198la) to calculate the parameters of the curve by holts's method (Gulland, 1969).
The mean otolith lengths of each age-gp (at Oct/Nov) were calculated from the mean values at 2 monthly intervals at each age-gp. So it was in units of 12 months. 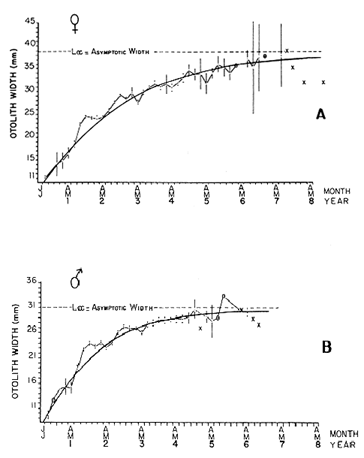 Figure 5. Mean otolith length for age at two monthly intervals with 0.95 C.I A fitted von Bertalanffy curve is shown. A) Females: La= 3.8725, K= 0.410945, to= 0.02943, B) MAles : L= 3.10006, K= 0.671684, to= 0.27865 The program was initiated with pooled data for 0-gp fish in August and September to April and May, then the designated t: 1 corresponded to an otolith from 3 months old to 12 months in May, t: 2 corresponded to an otolith from 13 months old up to 24 months in May and so on. This smoothing technique consists in stabilizing the length at age (Ortega-Salas, 198la, 1981b). Then the parameters of the equation found are the following: 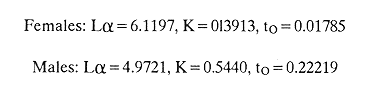 The theoretical curve fits reasonable well for mean values of otolith length at two monthly intervals (Fig. 4) but within the limitations of widespread individual values showed in the scattergrams (Fig. 2). That widespread situation is not very different that fish length show, comparatively (Ortega-Salas, 198la, 1981b). Otolith width forage was also calculated as it was done above. Then the parameters of the equation found are the following: 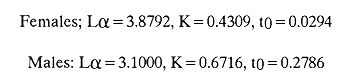 Comparing the fitted curves with observed mean lengths, and widths at age 1.5 and 2.5 years there is a wide difference. This occurs at the time of the year when there is an apparent change from rapid growth to slower growth, however, the theoretical curve fits the whole range of data reasonable well. These results suggest a possible use of summer zones and winter rings measurements of the otoliths in order to backcalculate the otolith rate of growth as individual. ESTIMATE OF FISH GROWTH FROM OTOLITH GROWTHSo far the age of the fish has been determined and also the rate of the otolith growth, now in order to estimate the fish growth from otolith growth we, have to find which is the relationship between otoliths and fish, therefore individual otoliths were measured by length and by width and plotted, separately, against the corresponding length of the fish whose results are shown in figures 6 and 7, respectively. The correlation was high. Also a transformation to natural logarithsm (LN) using the same parameters in the same relationship gave the scattergrams (Figs. 8 and 9), respectively. The correlation was also high. In the former cases the correlation coefficient was between 0.91 and 0.94, in the latter cases it was between 0.94 and 0.96 which suggest that the transformation LN/LN relationship was, slightly, better. An example in figure 6 is shown. So the fitted LN/IN relationship was plotted in its geometric form by the following equations: (Figs. 10 and 11). 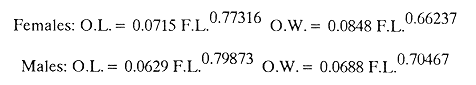 Then what is important is to be able to estimate the fish lenght from the otholith measurements, so we may use, firstly, Ricker's (1973) method to get the functional regression line which calculates the mean regression line from the dispersion of the correlation points, and secondly, we shall be able to interchange with the function of the axes "X" and "Y" each other. In the former case, a new slope (v) and a new Y-axis intercept (A) would be calculated from the original form: LNY = a + b (INX) where: R: Correlation coefficient. b. Slope. Y: Mean of Yaxis. A; Y-axis intercept. X: Mean of X axis. from where t b/R = v; LN Y - ((v) LN X) = A, therefore the functional regression line will be as follows: 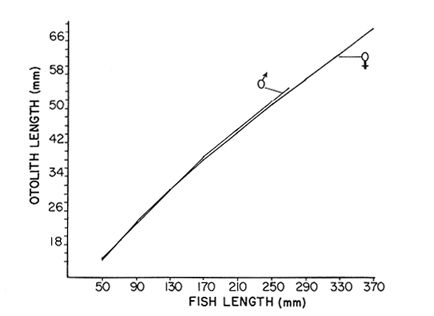 Figure 6. Otolith length - fish length relationship, A) Females, B) Males showing the theoretical straigth line and the LN/LN regression line (dashed line) 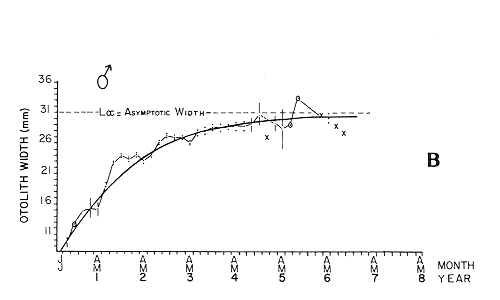 Figure 7. Otolith width-fish length relationship, A Females, B Males LNY = A + v(LNX) Secondly, in order to intrchange the axes' functions, instead of being Y = f(X) would be X = f (Y) as follows: vi = l/v and Ai = LN X - «vi) LN Y) therefore the functional regresion line would be written as follows: 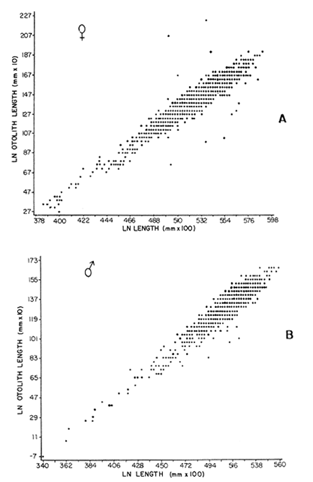 Figure 8. LN Otolith length - LN fish length relationship, A) Females, B) Males LNX = A1 +v1(LNY) which for practical purposes might be represented as figure 12 A and B. An example of the calculation for the functional otolith length-fish length relationship would be as follows: 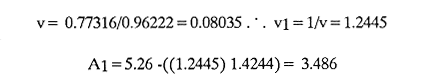 which for practical purposes might be written as follows (lenghts in mm): Females: Fish length = 1.244 (LN Otolith length) + 3.486. Thus, the functional regression lines may be written as shown in figure 12 A and B but because we intend to estimate fish length at age from otoliths, we just calculate the otolith length or otolith width at age as shown above and apply the same equation as figure 12 A and B which may be written as follows: 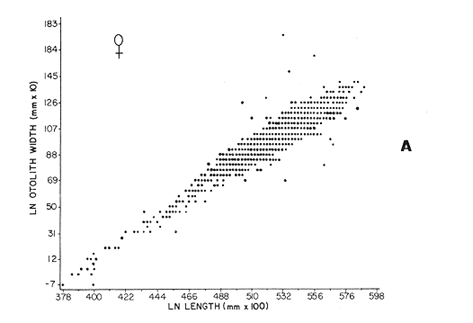 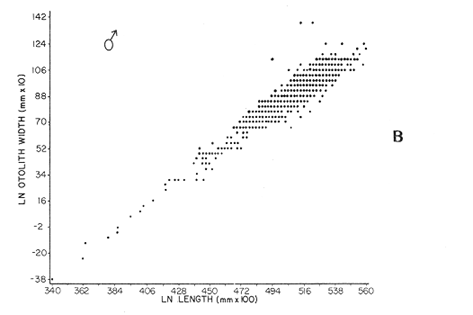 Figure 9. LN Otolith width - LN fishlength relationship, A) Females, B) Males 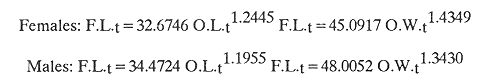 where F.L.t: Fish length (mm) at time t. O.L.t: Otolith length (mm) at time (calculated from von Bertalanffy's). O.W.t: Otolith width (mm) at time t (calculated from von Bertalanffy's). Comparing the fish length at age calculated in OrtegaSalas (198la, 1981b) with those fish lengths calculated from otolith length and those from otolith width we have the following results (Table 1): The results from table 1, show that in earlier stages using the otolith length gave better estimates for fish length but in older age-gps from age-gp III it seems to be better to use the otolith width. Eventhough the results showed that to estimate fish length from otolith measurements fits reasonable well. 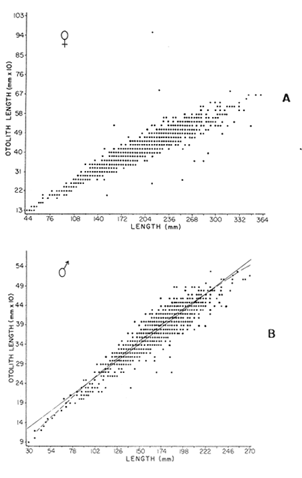 Figure 10. Otoligth - fish length relation given by the following equations: A) Females: O.L. = 0.0715 F.L. 0.77316, B)Males: O.L. =0.0629 F.L. 0.79873 Thus, we may conclude the following: 1. The age determination in months was considerd to be acceptable assumed May as the birthmonth: age in years was also accurate by using summer zones (May-October) and winter rings (November-April). 2. The von Bertalanffy's model was appropiate to estimate otolith length, and otolith width at age. Otolith growth was faster in summer than in winter. The relationship, otolith growth and fish growth was close related. 3. Using a combination of the von Bertalanffy's model and the relationship of the otolith to the fish ¡t should be possible to estimate fish length either from otolith length or otolith width. 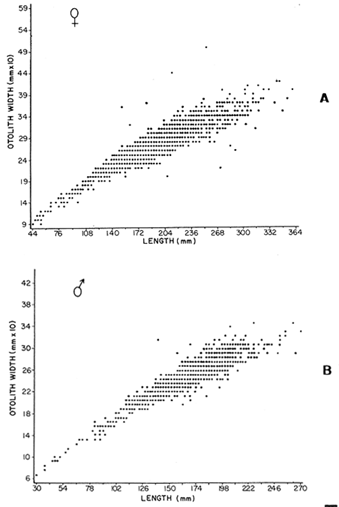 Figure 11. Otoligth width - fish length relation given by the following equations: A) Females: O.L. = 0.0848 F.L. 066237, B) Males : O.L. = 0.0688 F.L. 0.70467 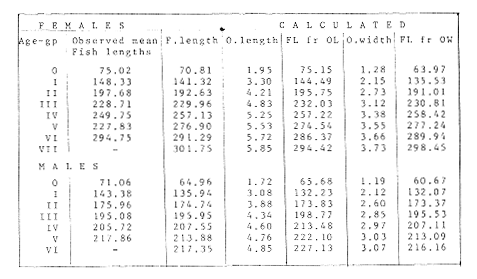 TABLE 1 ESTIMATES OF FISH LENGTH (FL) FROM ESTIMATES OF OTOLITH LENGTH (OL) AND FROM OTOLITH WIDTH (OW), AND COMPARED WITH ESTIMATES OF FISH LENGTH CALCULATED FROM OBSERVED MEAN FISH LENGTH AT EACH AGE - GP. THE UNITS ARE IN mm 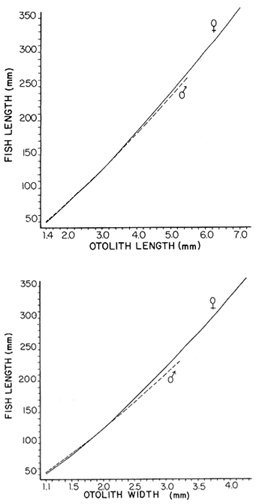 Fig. 12. Fishn length - Otolinth length relation given by the following funtional aquation: Females: f.l. = 32.6746 o.l. 1.2445 Males: F.L. = 45.0917 O.W. 1.4349 Males : F.L. = 48.0052 O.W. 1.3430 AgradecimientosAcknowledgments are du to A. Burd and T. Williams for their advise. For Laboratory facilities to Department of Marine Biology of the University of Liverpool, and MAFF-Fisheries Laboratory Lowestoft. And for the fi nantial support, Universidad Nacional Autónoma de México; Consejo Nacional de Ciencia y Tecnología and The British Council. LITERATURABAGENAL, T. B., (Ed.). The Ageing of Fish. Proc. Int. Symp. Unwin Bros. Ltd, 1974. 234 p. BERTALANFFY, L von, Theoretische Biologie-Zweiter Band: Stoffwechsel, Wachstum A. Francke A.G. Verlag, Bera. 1951. 418P. BEVERTON, R. J. and S. J. HOLT, On the Dynamics of Exploited Fish Population. Fish. Invest. London Ser. 1957. 533 p. 19, II BOHL, H., Die Biologic der Kliesche (Limanda limanda (L.) in der Nordsee. Ber. Dtsch. Wiss. Kemm. Meer., 1959. 1-57. 15: FAGADE, S. O., The Ageing of Fish. Proc. Int. Symp. Age determination in Tilapia melanostheron (Ruppell) in the Lagos Lagoon, Lagos, Nigeria. In: Bagenal. T.B. (Ed.). 1974. 71-76. GRAHAM, M., Fish Invest. London, Ser. Studies of Age-Determination in Fish, Part II A Survey of the Literature. 1929. 2, 11, 3, 50P GULLAND, J. A., Fish. Tech. Pap., Manual of Methods for Fish Stock Assessment, Part. 1. Fish Population Analysis. FAO 1969. 1-154. 40 (2): HOFSTEDE, A. E., The Ageing of Fish. Proc. Int. Symp. Studies on Growth, Ageing and Backcalculation of Roach. Rutilus rutilus (L.) and Dace, Leuciscus leuciscus (L). In: Bagenal, T.B. Ed. 1974. 137-147. LEE, C. Y, C., Thesis, The Boiology and Population Dynamics of the Common Dab, Limanda limanda (L) in the North Sea. Ph.D. Univ. East. Anglia, U.K 1972. 105 P. MOLLER, C. J., Penguin Books Ltd. Fishes of the British and Northern European Seas. 1978. 128 p. ORTEGA-SALAS, A. A., Cyclic Phenomena in Marine Plants and Animals. Pergamon Press Seasonal changes in Dab (Limanda limanda) in Isle of Man waters (abstract). In: Naylor, E. and R.G. Hartnoli (Ed.). 1979. 477 p. ORTEGA-SALAS, A. A., J. Fish. Biol., Seasonal changes in the common Dab, Limanda limando (L.) in Isle of Man waters. 1980. 7-2. 16: ORTEGA-SALAS, A. A., Thesis Biology of the Dab, Limanda limanda (L) in Isle of Man waters, Ph.D. Univ. Liverpool, U.K. 1981a. 98p. ORTEGA-SALAS, A. A., Age and growth of the Dab, Limanda limanda (L.) in Isle of Man waters 1981b. (in preparation). ORTEGA-SALAS, A. A., Diet, food consumption and growth of the Dab, Limanda limanda (L) in Isle of Man waters 1981c. (in preparation). ORTEGA-SALAS, A. A., Population structure of the Dab, Limanda limanda (L.) in Isle of Man waters 1981d. (in preparation). RICKER, W. E., J. Fish. Res. Bd. Can., Linear regressions in fisheries research. 1973. 409-434. 30: SULLIVAN, K J., N.Z.J. Mar. Fresh., Res., Age and growth of the Elephant Fish Collorhinchus milii (Elasmobranchii: Callorhyunchidae). 1978. 745-753. 11 (4): WILLIAMS, T. and B. C. BEDFORD, The Ageing of Fish Proc. Int. Symp. The use of otoliths for age determination. In: Bagenal, T.B. (Ed.). 1974. 114-123.
|