|
EFECTO DE LA MAREA EN EL PASO DE LAS
LARVAS DE ALGUNAS ESPECIES DE LA FAMILIA CLUPEIDAE (PISCES), EN LA BOCA DEL
CARMEN, LAGUNA DE TÉRMINOS, CAMPECHE, MÉXICO
Trabajo
recibido el 29 de agosto de 1985 y aceptado para su publicación el 21 de
mayo de 1986
César Flores Coto
Verónica Ducoing Chaho
Faustino Zavala García
Arturo Velarde Méndez
Sergio Méndez Velarde
Universidad Nacional
Autónoma de México. Instituto de Ciencias del Mar y
limnología. Contribución 518 del Instituto del Ciencias del Mar y
Limnología.
Se estudia el paso de las larvas de
la familia clupeidae a través de la Boca del Carmen en la Laguna de
Tériminos, Campeche, en relación a la marca y fisiografía
de la boca, estableciendose tres estaciones de muestreo en su parte más
angosta, ubicadas en su porción media (somera) y canales adyacentes. Se
muestreó a diferentes profundidades, con una periodicidad de tres horas
cubrien do un cielo de marca. Se realizaron ocho campañas entre abril de
1980 y enero de 1981. Las redes de plancton fueron de 50 cm de diámetro
en la boca y malla de 500 µm. A través de todas las
campañas fue aparente que la mayor entrada de aguas neríticas
ocurrió en la porción oriental, en tanto la salida de aguas
diluídas y dulces fue principalmente por la occidental. Se registraron
cuatro especies, Ophisthonerna oglinum, Harengula
jaguana, Brevoortia gunteri y una no determinada. O.
oglinum fue la especie más abundante particularmente al
principio (enero) y fin (abril) de la época de secas; sus larvas
penetran a la laguna acarreadas por la marea, principalmente por la zona
oriental que tuvo la influencia marina más fuerte. H.
jaguana fue la segunda especie mas abundante, ocurriendo
principalmente en primavera (abril-junio) y casi ausente en época de
lluvias y como la anterior, se mostró claramente sujeta a la marea en su
paso por la boca. B. gunteri penetró a la laguna por
la porción occidental y se establece la idea que su migración se
rige principalmente por un gradiente salino y la marea tiene una influencia
secundaria. Se estima que O. oglinum y H.
jaguana, aunque en forma mínima desovan en la
laguna.
The migration of larvae of the
Clupeidae family through the Carmen Inlet in Terminos Lagoon, Campeche, is
studied in relation to the tide and inlet physiograr by. Three sampling
stations were established, one in the medium area (shallow) and two at the
adjacent channels of the narrowest part of the inlet. The samples were taken at
differents dephs every three hotirs covering a tidal cycle. Eight trips between
April 1980 and January 1981 were carry out. A plankton net was used with a 50
cm diameter mouth and 500 µm of mesh. In all the trips it was apparent
that the major inflow of neritie waters was in the eastern region, whereas the
outilow of diluted and freshwaters occured at the westem region. Four species
were registred, three of them identified as Ophistfionerna
oglinum, Harengula jaguGna and Brevoortia gunteri; the last one was
not identific. The most abundant species was O. oglinum,
specially at the beginning (January) and the end (April) of the dry season. The
larvae entered the lagoon carried by the tide mairily through the eastern
region, which had the strongest marine influence. H. jaguana was the second
more abundant species with a maximal ocurrence in spring (Apríl to
June), and it was scarce in the rainy season. As weil as O.
oglinum its migration through the inlet has a strong tidal
dependence. B. gunteri penetrated the lagoon through the western region so it
was established that the migration was mainly influence by saline gradient,
whereas the tide had a secondary role. It was estimated that O.
oglinum and H. jaguana spawried in the lagoon,
although in minimal quantity.
Las lagunas costeras están generalmente separadas del mar por una barrera arenosa y se comunican con el por una o varias aberturas o bocas relativa mente estrechas, a través de las cuales se mezclan las aguas lagunares y marinas, teniendo como causa principal a las corrientes de marea. Estos cuerpos de agua son usados temporal o permanentemente por organismos salobres, marinos o dulceacuícolas, como áreas de refugio, criaderos o de transito; entre otros un alto porcentaje de los peces costeros desovan en el mar y sus larvas migran o son transportadas a estos sistemas a través de sus bocas. Probablemente uno de los factores más importantes de la relación entre las larvas, las áreas costeras y su migración hacia las lagunas, sea la marea. El ictioplancton en las lagunas costeras presenta una serie de componentes dependientes de las condiciones hidrológicas del sistema, existiendo una diversidad de especies que representan una alta complejidad, la cual es necesario entender para explotar y conservar racionalmente (Flores Coto y Alvarez-Cadena, 1980). Siendo la Laguna de Términos una de las más extensas e importantes del país, resulta obvia la necesidad de conocer acerca de las bocas en el paso de las larvas; algunas investigaciones se han realizado en este sentido: Alvarez-Guillén (1983), Bravo-Nuñez y Yáñez-Arancibia (1979), Yáñez-Arancibia y Lara-Domínguez (1983), que versan sobre peces adultos, Arenas-Mendieta y Yáñez-Martinez (1981), que analizan el paso de postlarvas de camarón y el de Méndez-Velarde y Velarde-Méndez (1982) sobre las familias de ictioplancton en la Boca del Carmen; este último es el antecedente del presente trabajo y que llevo a escoger a la familia Clupeidae, tanto por su relativa abundancia, como por comprender especies de importancia económica (Houde, 1975). Los objetivos planteados son en primer término, conocer el efecto de la marea sobre el paso de larvas a través de la boca, pero también el papel de otros factores, como la salinidad, temperatura y profundidad, durante un ciclo anual. ÁREA DE ETSUDIOEL área de estudio fue la Boca del Carmen, situada en la porción noroeste de la Laguna de Términos, la cual se localiza entre los 18° 20'y 19° 00'N y los 91° 10'y 92° 00' O; al suroeste del Golfo de México (Fig. l). Características de la laguna en forma extensa, son referidas en los trabajos mencionados anteriormente, así como los de Yáñez-Correa (1963), Botello (1978), Amezcua Linares y Yáñez-Arancibia (1980) entre otros. La Boca del Carmen (Fig. 2) tiene aproximadamente 6.2 km en su parte más ancha y 3.5 km en la más angosta, se aprecia un delta interior poco desarrollado y uno mayor en la porción oceánica (Pheleger y Ayala-Castañares, 1971). Su batimetría es irregular encontrándose un canal proximo a la Isla del Carmen, con una profundidad de 10 a 12 m y otro en el extremo opuesto (Punta Zacatal) donde se registran profundidades hasta de 6 m, la parte central es somera con profundidades de 2 a 4 m. La velocidad de la corriente en el canal oriental va de 25 a 60 cm/seg en el flujo y más de 80 cm/seg en el reflujo (Phleger y Ayala-Castañares, 1971). El patrón de sedimentación y la topografía están determinados principalmente por las corrientes de marea (Yáñez-Correa, 1963), incluso por aquellas que proveniendo de la otra boca, Boca de Paso Real, generan una circulación general en la laguna que lleva a un flujo neto este - oeste penetrando el mayor volúmen de agua marina por esta boca y saliendo por la Boca del Carmen (Mancilla-Peraza y Vargas-Flores, 1980). MATERIAL Y MÉTODOSSe establecieron tres localidades de muestreo en la parte más angosta de la Boca del Carmen, la estación A junto a la Isla del Carmen, sobre el canal principal, la B en la parte central que es la más somera y la C frente a Punta Zacatal, con un canal menos profundo (Fig. 2). En la estación A se muestreo a tres profundidades: superficie (1 m), media agua (3-4 m) y fondo (6-7 m), en la estación B se muestreó entre 0.5 y 1.5 m y en la estación C en superficie (1 m) y fondo (3-4 m). Se realizaron ocho campañas cubriendo un ciclo anual. Las fechas de colecta fueron: 22 y 23 de abril 9 y 10 de junio, 10 y 11 de julio, 9 y 10 de agosto, 14 de septiembre, 15 y 16 de octubre, 7 de diciembre de 1980 y 18 de enero de 1981. Para realizar los muestreos se seleccionaron los días de máxima amplitud de marea (sicigia), por ser ésta el principal factor a considerar. A fin de cubrir un ciclo de marea y para el análisis de los resultados, la curva de marea fue dividida en 7 momentos; el momento 1 es el inicio del flujo, los momentos 2 y 3 corresponden a los de máxima velocidad de corriente para el flujo, el 4 es el que registra el máximo nivel de marea (estoa), 5 y 6 corresponden a los momentos de máxima velocidad de reflujo y el 7 es el que finaliza el ciclo (Fig. 3). 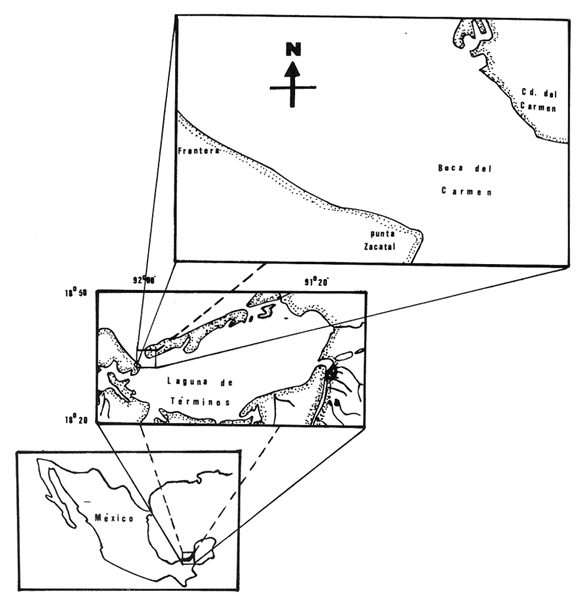 Figura 1. Localización del área de estudio Se determinaron la salinidad y la temperatura con un refractómetro y un termómetro de cubeta respectivamente, las muestras de media agua y fondo se colectaron con botella Van Dorn. Para el muestreo se utilizaron tres redes de plancton tipo trapecio de 0.5 in de boca, 1.5 m de largo y luz de malla de 500 micras, con un contador de flujo en la boca de cada red. Se hicieron arrastres horizontales y simultáneos a las diferentes profundidades. Estos muestreos se realizaron cada tres horas, con una duración de tres minutos. 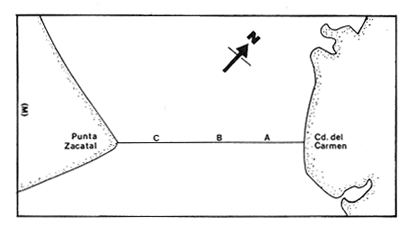 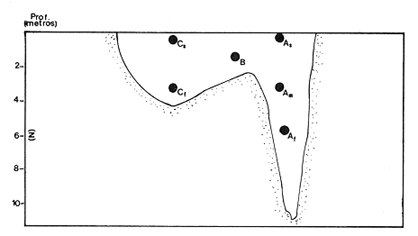 Figura 2. Localización de las estaciones de muestreo (M) y perfil batimétrico (N) con los niveles de muestreo El horario de muestreo se estableció de acuerdo al calendario de marea publicado por el Instituto de Geofísica de la UNAM (1979). 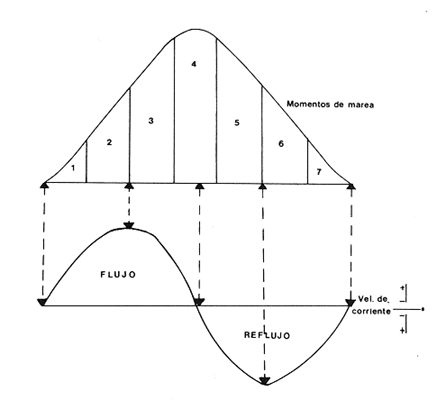 Firura 3. División de la curva de marea, atendiendo a las velocidades de corriente, que se generan en su ciclo Las muestras de plancton se fijaron inmediatamente después de la colecta, con formalina al 4 % neutralizada con borato de sodio. En el laboratorio las larvas de peces fueron extraidas en su totalidad e identificadas a nivel de familia, separando las de la familia Clupeídae en su ulterior identificación específica y medición. RESULTADOSLos ciclos salino y térmico registrados para cada estación y profundidad de muestreo, se muestran en la figura 4, cuyos valores correponden a los promedios del ciclo de marea muestreados. La salinidad tuvo sus valores mayores en los meses de primavera y los menores en el otoño como resultado de precipitación y descarga de grandes volúmenes de agua dulce en la laguna. La temperatura alcanza los mayores valores en junio y julio mostrando un ligero descenso en los siguientes meses y registrando los menores valores en diciembre y enero. Las diferencias entre las áreas oriental y occidental que se aprecian en la mencionada figura son más notorias en los valores individuales máximo y mínimo que ocurrieron en las distintas épocas climáticas que se presentan en la Tabla 1. De los valores de esta tabla cabe destacar la ocurrencia de mayores temperaturas y menores salinidades en la porción occidental (estación C) y viceversa en la oriental, por ser áreas de mayor influencia lagunar la primera y marina la segunda. La variación de la salinidad principalmente y en menor grado la de la temperatura, muestran que el condicionamiento hídrico por la marea no es homogéneo a través de la boca, así en la zona del canal oriental y siguiendo la curva de predicción de marea, durante el flujo las aguas oceánicas densas penetran por el fondo o cerca de él con las mayores velocidades, en tanto en la superfice las corrientes tienden a ser más débiles, con aguas menos salinas, mismas que durante el reflujo son las que salen con una mayor velocidad. Este esquema se observó durante casi todos los meses y fue menos evidente en aquellos de primavera, cuando las diferencias entre las aguas lagunares y costeras son mínimas. 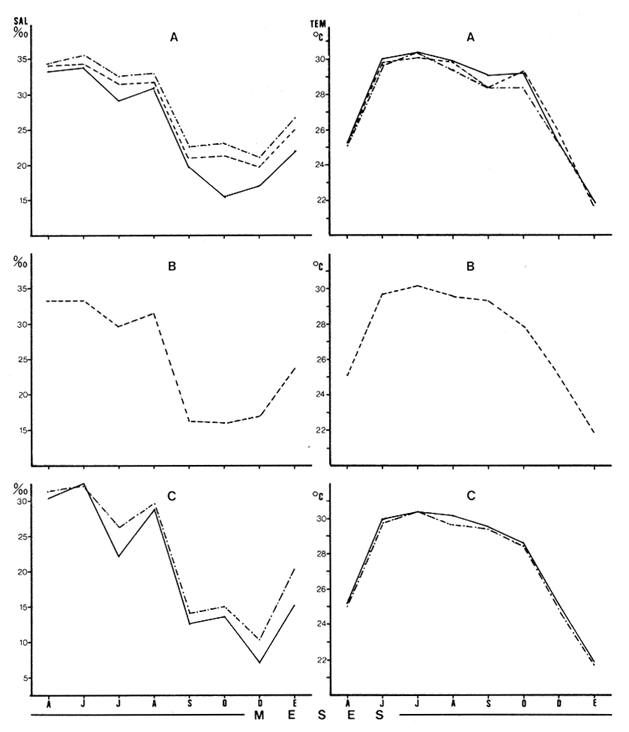 Figura 4. Salinidades y temperaturas promedio de cada ciclo de marea, para distintos meses muestrados; en las estacioes A, B y C. (superficie,____; media agua,----; fondo_._._._._). ICTIOPLACTONLa familia Clupeidae fue la cuarta más abundante durante las 8 campañas de muestreo, después de Engraulidae, Gobiidae Y Sciaenidae. Estuvo representada por: Opisthonema oglinum (799 larvas), Harengula jaguana (725 larvas) y Breyoodia gunteri (52 larvas), además de una serie de especímenes no determinados. 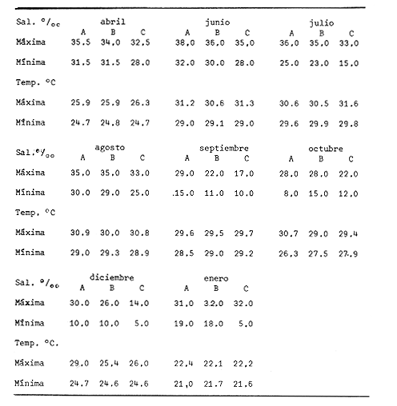 TABLA 1 SALINIDADES Y TEMPERATURAS DURANTE LOS CICLOS DE MAREA, DE ABRIL, 1980 A ENERO 1981, EN LAS ESTACIONES A, B, C. Opisthonema oglinum Esta es una especie de amplia distribución en el Atlantico, desde Maine, EU, hasta el sur de Brasil (Jones et al., 1978), cuyo habitat corresponde principalmente a las zonas costeras y someras en profundidades menores de 50 m y aguas de salinidades mayores de 27 %, no obstante llega a perietrar en aguas salobres (Houde, 1977a) como es el caso de la Laguna de Términos, donde larvas y adultos han sido colectados, aunque con bajos niveles de abundancia (Ferreira González y Acal-Sánchez, 1984; Flores Coto y Alvarez Cadena, 1980; Resendez, 1981; Yáñez-Arancibia et al.,1980). Fue la especie más abundante entre los clupeidos, capturándose 799 larvas a través del año. La mayor densidad larvaria ocurrió en dos periodos, el primero y más importante fue al final de la época de secas, con abril como el mes en el que presentó el máximo desove, decreciendo la abundancia notoriamente en los siguientes meses. El segundo periodo correspondió a enero, inicio de la época de secas. La presencia de esta especie durante la época de lluvias se restringe principalmente a los meses de julio y agosto, durante los siguientes meses (septiembre a diciembre), salvo por unos cuantos especímenes, prácticamente no aparece (Fig. 5, Tabla 2). 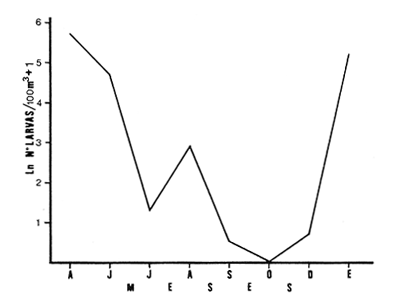 Figura 5 Ophisthonema oglinum. Densidad larvaria promedio de cada ciclo de marea, para los distintos meses muestrados. 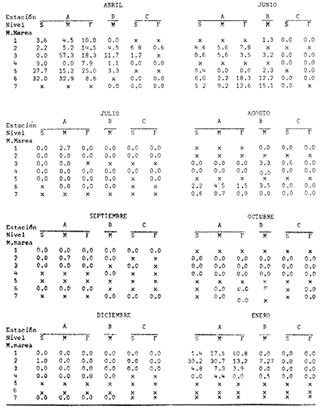 TABLA 2 DENSIDAD LARVARIA DE OPHISTHONEMA OGLINUM DE ABRIL 1980 A ENERO 1981 EN LAS ESTACIONES, NIVELES DE MUESTREO Y MOMENTOS DE MAREA. S = SUPERFICIE; M = MEDIA AGUA T F = FONDO. LOS VALORES SON NÚMEROS DE LARVAS POR 100 M3 Estos resultados corresponden cercanamente con los de Ferreira González y Acal-Sánchez (1984) para la propia laguna, aunque en su caso encuentran en enero la mayor abundancia larvaria, con un segundo máximo en abril. Asimismo, la especie fue también muy escasa durante la época de lluvias. En términos generales esta especie está sujeta a la marea en su paso por la Boca del Carmen, siendo los momentos de mayor velocidad de corriente (2-3 en el flujo y 5-6 en el reflujo), los que determinan donde ocurre la mayor densidad. Las diferencias de densidad larvaria en la columna de agua son también resultado del efecto de la marea, habiendose encontrado los valores mayores en el fondo y media-agua durante el flujo, en tanto durante el reflujo ocurrieron en superficie. En general las densidades altas se presentaron a media agua, lo que es atribuible a que por la propia topografía, las corrientes mas veloces se presentaron ahí y no en el fondo. Este esquema de distribución ligado a la marca no ocurrió en la zona occidental, no solo por la escasez de larvas, sino también porque ahí no se guarda estrictamente el patrón de predicción marea. Existe una diferencia notable del paso de las larvas por las distintas áreas de la boca, resultado de la propia topografía y por ende distintas velocidades de corriente durante los ciclos de marca y de factores tales como la salinidad y temperatura. En la figura 6 donde se muestra la densidad larvaria para sendas estaciones, se nota un gradiente muy marcado que va de la zona oriental (estación A) a la occidental (estación C). La época de lluvias (Julio - diciembre) fue como se mencionó, la de menor abundancia larvaria, aunque conservó el aspecto general de la distribución presentando las mayores densidades promedio de las zonas oriental y media de la boca (estaciones A y B), en los momentos de mayor velocidad de corriente. La época de secas presentó diferencias entre los ciclos de enero y abril que correspondieron al inicial y final respectivamente, del proceso de resalinización de la laguna (Fig. 7). Durante enero que fue una de las épocas de gran abundancia larvaria, éstas no se capturaron en la porción occidental de la boca y fueron muy escasas en la zona media; en la porción oriental en cambio (estación A), hubo gran densidad larvaria durante el flujo, aún en el momento uno que es de baja velocidad de corriente. Durante abril en cambio el balance entre la entrada y salida de larvas fue favorable a la salida (Tabla 3). 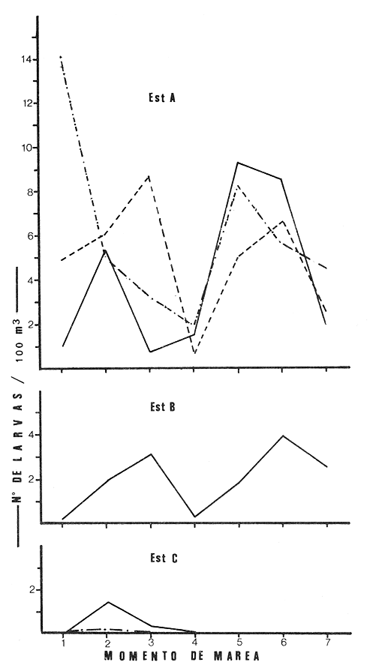 Figura 6. Ophithonema oginum. Dencidad larvaria promedio de las 8 campñas de muestreo, para los distintos momentos de marea en los diferentes niveles de estaciónes A, B y C de la Boca del Carmen (superficie___; medi agua---; fondo-.-.-.- Herengula jaguana Esta especie tiene amplia distribución en el Atlantico Oeste, desde Nueva Jersey hasta Santos, Brasil, inclu yendo el Golfo de México (Houde, 1977b). Hoese y Moore (1977) la refieren para la parte interna de la plataforma continental, al norte del golfo, como la más común entre los clupcidos, excepto donde es reemplazada por Brevoortia patronus. En la Sonda de Campeche es una de las especies más frecuentes y abundantes (Sánchez Gil et al., 1981). Ha sido registrada en la Laguna de Términos por Reséndez (1981).  TABLA 3 RELACIÓN DE TALLAS DE OPHISTHONEMA OGLINUM LOS VALORES SON NUMERO DE ORGANISMOS POR CLASE DE TALLA. EN: A = LIBRES; B = SUBRAYADOS Y C = ENTRE PARENTESIS. Sus hábitos son más bien costeros, incluyendo el desove, que según Houde (1977b) en el noroeste del golfo, está confinado a una banda de 85 km desde la costa hasta profundidades menores de 30 m, ocurriendo de enero a septiembre. Se obtuvieron un total de 725 larvas de H.jaguana. Su ciclo anual de abundancia muestra que su periodo de máximo desove ocurre en primavera (abril - junio), el resto del año fue sumamente escasa, llegando a estar ausente en septiembre y octubre (Fig. 8). Su presencia nuevamente en diciembre puede significar el inicio de un nuevo ciclo de penetración a la laguna correspondiendo con el de resalinización. Este ciclo concuerda con los resultados de Ferreira-González y Acal-Sánchez (1984), que indican la presencia de larvas principalmente en junio, siendo muy escasas el resto del año, lo que hace notorio un pequeño pico de abundancia en el mes de enero. Los resultados muestran que la marea es el factor principal que determina el paso de las larvas a través de la boca, existiendo una correspondencia entre los momentos de máxima velocidad de corriente y la mayor abundancia de larvas, lo que implica incluso, mayores densidades a media agua y fondo que en la superficie (Tabla 4). El balance entre la entrada y salida, se mostró fuertemente influenciado por las tallas de las larvas (Tabla 5). Cabe llamar la atención que este patrón se mantuvo para la estación occidental (C) solo durante abril y junio pues el resto de las campañas solo se capturaron 4 organismos y de gran talla, mayores de 19 mm. Las diferencias de abundancia fueron muy marcadas entre la zona oriental de la boca (estación A) donde ocurrió la mayor abundancia, y la parte media (estación B) y occidental (estación C); particularmente con ésta última donde se capturaron 23 especímenes (3 %) (Fig. 9). 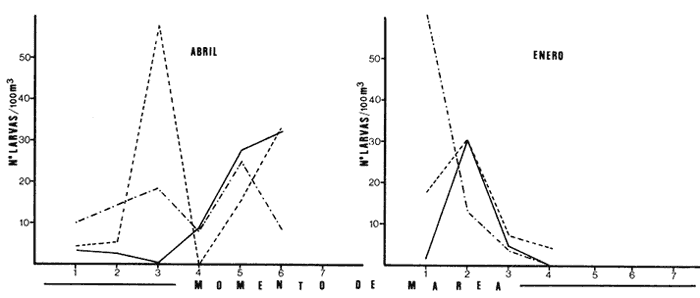 Figura 7. Ophistonema oglinum. Densidad para los distintos momentos de marea y niveles de muestreo en la estación A durante los meses de mayor abundancia. Superficie: media agua; fondo 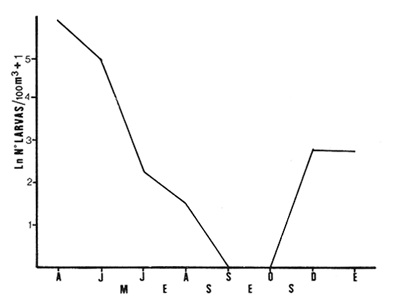 Figura 8. Harengula jaguana. Densidad larvaria de cada ciclo de marea para los meses muestreados 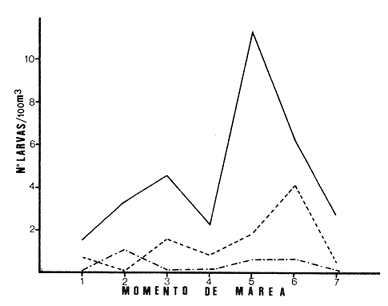 Figura 9. Harengula jaguana. Densidad larvaria promedio de las 8 campañas de muestreo en los distintos momentos de marea para las estaciones A (-), B (--)) y C(----). El patrón de distribución de un ciclo a otro, muestra diferencias atribuibles en primer término al estado o nivel salino de la laguna, así durante abril, el balance entre la entrada y salida de larvas es faverable a la entrada, en cambio en junio lo es la salida (Fig. 10). Los límites de salinidad y temperatura en que se capturó esta especie fueron de 15 a 38 %o y de 27.7 a 31°C respectivamente. Brevoortia gunteri La distribución de esta especie parece restringirse al Golfo de México, con su área de mayor abundancia entre la Bahía de Copano, Texas, EU y la Laguna Madre, Tamaulipas, México, habita en aguas costeras y puede penetrar en estuarios y lagunas costeras, como en el caso de la Laguna de Términos (Reintjes y Pacheco, 1966; Reséndez, 1981; Walls, 1975). Las larvas de esta especie fueron sumamente escasas y ocurrieron casi exclusivamente en enero, excepción hecha de algunos especímenes presentes en abril y junio. Se capturaron en momentos tanto de alta como de baja velocidad de corriente e incluso en estoa (Tabla 6). Su distribución a través de la boca mostró el mayor número de larvas en la porción occidental (estación C), que fue la que a través del año presentó las menores sa linidades; en tanto que en la porción oriental (estación A) la de mayor influencia marina estuvo prácticamente ausente. 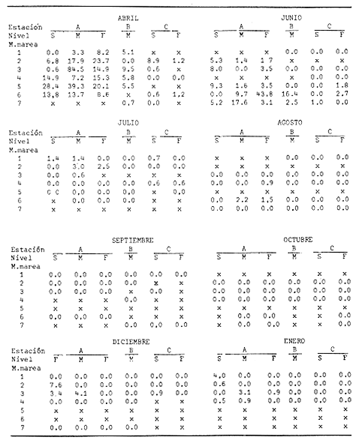 TABLA 4 DENSIDAD LARVARIA DE HARENGULA JAGUANA DE ABRIL 1980 A ENERO 1981, EN LAS DISTINTAS ESTACIONES DE MUESTREO Y MOMENTOS DE MAREA. LOS VALORES SON NÚMERO DE LARVAS POR 100M 3  TABLA 5 RELACIÓN DE TALLAS DE HARENGULA JAGUANA EN LOS DISTINTOS MOMENTOS DE MAREA Y NIVELES DE MUESTREO. LOS VALORES SON NUMERO DE ORGANISMOS POR CLASE DE TALLA. Ocurrieron en salinidades de 5.0 a 34 %o y temperaturas de 21.6 a 30.4°C. 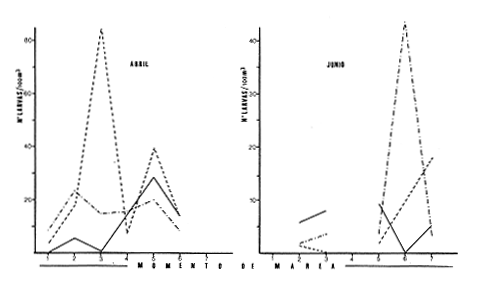 Figura 10. Harengula jaguana. Densidad larvaria para los distintos momentos de marea y niveles de muestreo en las estación A durante los meses de mayor abundancia. Superficie ___; media agua---; fondo-.-.-. 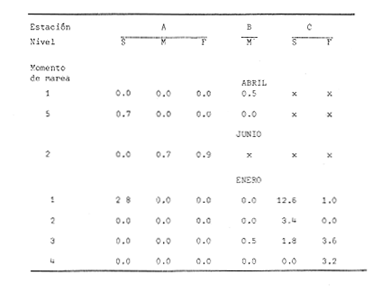 TABLA 6 DENSIDAD LARVARIA DE BREVOORTIA GUTERI DE ABRIL 1980 A ENERO 1981 EN LAS DISTINTAS ESTACIONES DE MUESTREO Y MOMENTOS DE MAREA DE LOS QUE SE ANOTAN SOLO LOS POSITIVOS, LOS VALORES DEL NÚMERO DE LARVAS POR M3 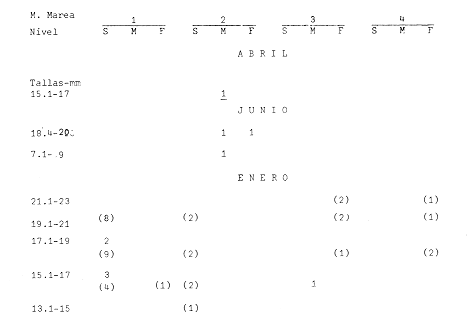 TABLA 7 RELACION DE TALLAS DE BREVOORTIA GUNTERI EN LOS DISTINTOS MOMENTOS DE MAREA Y NIVELES DE MUESTRE, LOS VALORES SON NÚMERO DE ORGANISMOS POR CLASE DE TALLA EN LA ESTACIÓN: A = LIBRES, B = SUBRAYADOS Y C = ENTRE PARENTESIS. Las tallas registradas variaron de 9 a 22 mm, pero el 95 % fueron mayores de 15 mm (Tabla 7). DISCUSIÓN
HIDROLOGÍALa Boca del Carmen se comporta como un típico estuario en el que las condiciones del medio ambiente son reguladas por la marea, con variaciones a veces muy grandes como sucede en el periodo de lluvias o pequeñas en el de secas, esquema particularmente evidente en el canal oriental (estación A). Las variaciones de salinidad en el canal occidental (estación C), seguramente también son resultado del efecto de la marea, no obstante no lo son de una manera clara para el patrón de predicción de marea que corresponde estrictamente a la porción oriental de la boca, donde se encuentra el mareógrafo. El no encontrar una estrecha correspondencia entre el flujo y el incremento de la salinidad y el reflujo con el decremento de la misma, es atribuible a una alteración del ciclo en ésta área y que sea una zona de mezcla más fuerte que el canal oriental, lo cual debe ocurrir por ser la porción por donde desaguan mayores cantidades de agua dulce, como lo muestran sus valores de salinidad siempre menores, comparados con el resto de la boca. La situación somera de la estación del centro de la boca (B), le lleva a tener un marco ambiental similar a la estación occidental (C) durante la época de lluvias y a la oriental (A) en la época de secas. Las corrientes en la estación media deben ser más débiles, pues su misma condición somera indica que los procesos abrasivos más fuertes se desarrollan en los canales laterales. ICTIOPLACTON
Ophisthonema oglinum Fue la especie más abundante. En términos generales se mostró sujeta a la marea en su paso por la Boca del Carmen, habiendo presentado sus mayores densidades en los momentos y niveles de mayor velocidad de corriente. La mayor abundancia en la zona oriental (estación A) parece consecuencia de ser el área donde se encuentra el canal principal de la boca y por tanto tener las mayores velocidades de corriente de marea, la mayor influencia de las aguas marinas, y las salinidades más altas. Contrariamente en la zona occidental la densidad larvaria fue sumamente baja, por mucho menor que la zona oriental e incluso que la parte media (estación B) que es una área somera, con bajas velocidades de corriente, tal escasez en la estación C es atribuible a que fue la zona que siempre tuvo las menores salinidades. Por lo anterior puede establecerse la idea de que la salinidad juega un papel preponderante en el paso de las larvas de O. oglinum a través de la Boca del Carmen. Parece obvio que deban existir diferencias de un ciclo a otro como resultado de la distribución de manchas o cardúrnenes de las larvas, de su propia capacidad de desplazamiento en la medida que crecen y de las condiciones ambientales que prevalezcan, particularmente entre las épocas de lluvias y secas. Por estas razones, la ausencia de esta especie durante la época de lluvias en la porción occidental de la boca y su escasez en la zona media y oriental, es atribuible a que durante ésta época, los grandes volúmenes de agua dulce descargados por el canal occidental e incluso por la parte media, hace que se restrinja principalmente a la porción oriental. Durante la época de secas (enero - abril) en que ocurrio la mayor abundancia particularmente en enero, la distribución larvaria fue similar a la época de lluvias, no habiéndose encontrado larvas en la porción occidental (estación C), muy escasas en la parte media y la mayor abundancia en la porción oriental (estación A), no obstante las causas son distintas y tal distribución debe ser consecuencia de que en este periodo inicial de resalinización cuando las aguas de la laguna son aun de baja salinidad, las larvas tienden hacia la laguna, alcanzándola a tallas mayores como puede inferirse del hecho que durante el flujo más del 28 % fueron mayores de 9 mm, capturadas en su mayor parte en momentos de baja velocidad de corriente; en abril en cambio más del 97 % de las larvas fueron menores de 9 mm, capturadas en su mayor parte en momentos de alta velocidad de corriente (Tabla 3). El que las larvas alcancen tallas cuyas capacidades natatorias les permitan movimientos independientes de las corrientes de marea, no obliga a que salgan de ellas, pero mientras no alcancen tales capacidades, dependerán totalmente de la marea en sus movimientos migratorios. Por otro lado, durante abril el balance de entrada y salida de larvas es favorable a la salida, lo que indica que la especie desova en la laguna al menos en áreas proximas al litoral interno de la Isla del Carmen, como pudo confirmarse por la presencia de huevos. Ferreira-González y Acal-Sánchez (1980), atendiendo a la distribución larvaria en la laguna, consideran que la especie desova en la zona costera y sus larvas penetran a la laguna principalmente por la Boca de Puerto Real, siendo arrastradas por las corrientes hacia la Boca del Carmen. Harengula jaguana Esta fue la segunda especie más abundante y como la anterior se vió en términos generales sujeta a la marea en su paso por la Boca del Carmen, lo que aunado a las condiciones fisiográficas de éstas producen que las mayores densidades larvales ocurrieran en la porción oriental, con gran escasez en la porción media y occidental, particularmente en ésta última, donde solo se capturó el 3 % del total de esta especie. Durante la época de mayor abundancia específicamente en abril, la mayor parte de las larvas presentes en el flujo, fueron menores de 11 mm, penetrando por la zona oriental; una cuantas mayores de 17 mm ocurrieron en la zona media y occidental ya en superficie o momentos de baja velocidad de corriente; esto es, notoriamente fuera del efecto de la marca (Tabla 5). Por otro lado, un alto porcentaje (25%) de aquellas presentes en el reflujo fueron de mayores de 11 mm, lo que muestra que hay un fuerte acarreo de pequeñas larvas hacia la laguna. Durante el mes de junio el 30 % de las larvas capturadas en el flujo fueron menores de 11 mm, en tanto en el reflujo fue de 95 % lo que parece mostrar que durante este mes en que la laguna alcanza su máxima salinidad la especie desova ahí y las pequeñas larvas quedan sujetas a la corriente. Este periodo de desove en la laguna ajusta bien a lo registrado por Ferreira González y Acal-Sánchez (1984) quienes señalan la mayor abundancia en junio y que fue muy contrastante con el resto del año, incluso con un pequeño pico de abundancia en enero. Durante la época de lluvias, etapa de desalinización de la laguna, las larvas prácticamente desaparecen y vuelven hasta el inicio de la etapa de resalinización, pero lo hacen a tallas relativamente grandes, pues excepto un par de ellas en enero, menores de 11 mm, las mas fueron mayores de 17 mm. Esto puede ser consecuencia de la presencia en la laguna de aguas oligo y mesohalinas y a que la condición estuarina se extiende bastante afuera de la boca y permite que especies estuarino-dependientes como H. jaguana no tengan que penetrar a la laguna y sólo lo hacen a tallas relativamente grandes que se guían por el gradiente salino y pueden soportar mejor salinidades bajas. En la revisión que hacen Reintjes y Pacheco (1966) sobre la relación de las especies del género Brevoortia y los estuarios, señalan que son costeras, desovando en el mar durante el invierno y sus larvas se mueven hacia los estuarios y lagunas costeras a los que penetran entre 12 y 34 mm, soportando una amplia variación de salinidad y temperatura (0 a 40 °C y 0 a 60 °/oo. Aunque no está incluida B. gunteri en la revisión, los resultados del presente trabajo ajustan cercanamente a tal esquema; así fue una especie que ocurrió principalmente en invierno y cuyas tallas fueron muy grandes variando de 9 a 22 mm, con el 95 % mayores de 15 mm. Estos resultados permiten establecer la idea que la migración de larvas de B. gunteri a la Laguna de Términos, se rige por un gradiente salino y que la marea no es factor primario que determine el paso de las larvas a través de la boca. La idea que puede verse reforzada por el hecho de que O. oglinum y H.Mjaguana que se mostraron claramente sujetas a la marea en su paso por la boca fueron mucho más abundantes en la porción oriental, ocurriendo sus mayores densidades en los momentos de mayor velocidad de corriente. Así mismo fueron muy escasas en la porción occidental, justo donde B. gunteri tuvo su mayor abundancia. Cabe señalar que Fore y Baxter (1972), refieren el paso de Bevortia thyranus en la Bahía de Galveston, Texas, como sujeta a la marea, con las mayores capturas durante el reflujo, de especímenes entre 19 y 23 mm, sin embargo, su metodología de campo y análisis no permiten una comparación estricta con el presente trabajo, pues por un lado emplearon una red arrastrada por el fondo y por otro no refieren diferencias entre las capturas de organismos durante la estoa y momentos de baja velocidad respecto a aquellas hechas en momentos de alta velocidad. ConclusionesLa Boca del Carmen se comporta como un estuario parcialmente mezclado, en el que el hecho más obvio es que las condiciones del medio son reguladas por la marea, con variaciones a veces muy grandes como sucede en el periodo de lluvias o mínimas en el de secas. Por otro lado, el condicionamiento hidrológico por la marea no es homogéneo a través de las distintas áreas de la boca, pues miestras en la porción oriental ajusta al patrón de predicción de marca, la occidental no lo hace, como consecuencia de ser la zona más proxima a desembocaduras de ríos y tener los mayores aportes de agua dulce. La parte media y somera de la boca tiene un comportamiento más similar a la parte oriental durante la época de secas y a la occidental durante las lluvias. La familia Clupeidae estuvo representada por las especies Opisthonema oglinum (799 larvas), Harengula jaguana (725 larvas), Brevoortia gunteri (52 larvas), además de una especie no identificada (157 larvas). Opisthonema oglinum y Harengula jaguana fueron especies que se mostraron claramente sujetas al efecto de la marea en su paso a través de la boca, por lo tanto tuvieron una mayor abundancia en la porción oriental, disminuyendo fuertemente hacia la occidental donde su presencia puede considerarse fortuita, o como consecuencia en algunos casos, de larvas de gran talla como lo fueron muchas de las capturadas en esta zona. El ciclo de abundancia de estas especies tiene una estrecha relación con el periodo de resalinización de la laguna, por lo que la máxima densidad ocurre al final de este periodo, siendo notable el decremento hacia la época de lluvias, llegando a estar ausentes hacia el final de este. El grueso de la población de estas especies desova en el área costera adyacente, pero los resultados evidencian que al menos una mínima parte lo hace en la laguna. Brevooplia gunteri fue una especie que penetró a la laguna por la porción occidental con tallas muy grandes, lo que permite establecer la idea que su migración se rige más por un gradiente salino que por la marea. AgradecimientosLos autores desean manifestar su agradecimiento a las autoridades del Instituto de Ciencias del Mar y Limnología cuyo financiamiento hizo posible la realización de este estudio. Especial mención a Rodolfo Cruz, cuyo apoyo durante el trabajo de campo fué fundamental. Asimismo a E. López por mecanografiar el manuscrito. LITERATURAÁLVAREZ-GUILLÉN, H., Tesis Prof. Biología, Ecología de la Boca del Carmen, Laguna de Términos. El habitat y análisis estructural de las comunidades de peces. Fac. Ciencias, Univ.Nal. Autón. México. 1983. 94 p. AMEZCUA LINARES, F. y A. YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol. Ecología de los sitemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces. Univ. Nal. Autón. México, 1980. 69-118. 7 (1): ANÓNIMO, Calendario gráfico de mareas. Inst. Geofís. Univ. Nal. Autón. México. Ciudad del Carmen, Campeche. 1980. ARENAS-MENDIETA, R. y A. YÁÑEZ-MARTINEZ, Tesis Prof. Biología, Patrón anual de inmigración de postlarvas de camarón (Crustacea: Decápoda: Penaeidae) en la Boca de Puerto Real, Laguna de Términos, Campeche. Fac. Ciencias, Univ. Nal. Autón. México. 1981. 93 p. BOTELLO, A. V., An. Inst. Cienc. del Mar y Limnol. Cuantificación de un derrame petrolero ocurrido en la Laguna de Términos, Campeche, México. Univ. Nal. Autón. México, 1980. 169-176. 7 (1): BRAVO-NUÑEZ, E. y A. YANEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol. Ecología de la Boca de Puerto Real, Laguna de Términos. I. Descripción del área y análisis estructural de las comunidades de peces. Univ. Nal. Autón. México, 1979. 125-182. 6 (1): FERREIRA-GONZÁLEZ, R. y D. E. ACAL-SÁNCHEZ, Tesis Prof. Biología, Estudio de la comunidad ictioplanctónica de la Laguna de Términos, Campeche. Esc. Nal. Est. Prof. Iztacala, Univ. Nal. Autón. México. 1984. 93 p. FLORES-COTO, C. Y J. ÁLVAREZ-CADENA, An. Centro Cienc. del Mar y Limnol. Estudios preliminares sobre abundancia y distribución del ictioplancton en la Laguna de Términos, Campeche. Univ. Nal. Autón. México, 1980. 67-78 7 (2): FORE, P. L. y K. N. BAXTER, Diel fluctuation in the catch of larva¡ Gulf Mcnhaden, Brevoortia patronus, at Galveston entrance, Texas. Trans. Amer. Fish. Soc., 1972. 729-732. 101 (4): HOESE, H. D. y R. H. MOORE, Fish of Gulf of Mexico (Texas, Louisiana and Adjacent Waters). Texas A and M Univ. Presss. 1977. 376 p. HOUDE, E. D., Proc. Gulf Carib. Fish. Abundance and potential for fisheries development of some sardine-like fishes in the Eastern Gulf of Mexico. Inst., Twenty-eighth. An. Session, October, 1975: 1975. 73-82 HOUDE, E. D., Fish. Bull., Abundance and potential yield of the Atlantic tread herring, Opisthonema oglinum, and aspects of its early life history in the Eastern Gulf of Mexico. 1977a. 493-512. 75: HOUDE, E. D., Fish. Bull., Abundance and potential yield of the scale sardine, llarengula jaguana, and aspects of its early fife history in the Eastem Gulf of Mexico. 1977b. 613-628. 75: JONES, W. P., F. D. MARTIN y J. D. HARDY, Power Plan Project, Office of Biological Service, Fish and Wildlife Serv. Development of Fishes of the Mid-Atlantic Bight. An Atlas of Egg, Larva¡ and Juvenile Stages. Acipenscridae Through Ictaluridae. USA Dept. Int. 1978. 300 p. Vol. 1. MANCILLA-PERAZA, M. y M. VARGAS-FLORES, An. Centro Cienc. del Mar y Limnol. Los primeros estudios sobre la circulación y el flujo neto de agua a través de la Laguna de Términos, Campeche. Univ. Nal. Autón. México, 1980. 1-12. 7 (2): MÉNDEZ-VELARDE, S. F. y A. VELERDE-MÉNDEZ, Tesis Prof. Biología, Estudio de ictioplancton en la Boca del Carmen, Laguna de Términos, Campeche. Fac. Ciencia. Univ. Nal. Autón. México. 1982. 77 p. PHLEGER, F. By A. AYALA-CASTAÑARES, Bull. Processes and history of Términos Lagoon, México. Am. Ass. Petrol. Geol., 1971. 2130-2140. 55 (2): REINTJES, J. W y A. L. PACHECO, The relation of Menhaden to estuaries. Am. Fish. Soc. Spec. Publ., 1966. 50-58. 3: RESENDEZ-MEDINA, A., I. Biótica, Estudio de los peces de la Laguna de Términos, Campeche, México. 1981. 239-291. 6 (3): SÁNCHEZ-GIL, P., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Diversidad, distribución y abundancia de las especies y poblaciones de peces demersales de la Sonda de Campeche. Univ. Nal. Autón. México, 1981. 209-239. 8 (1): WALLS, J. G., Fishes of Northern Gulf of Mexico. T.F.H. Pubis, New Jersey, USA 1975. 432 p. YANEZ-ARANCIBIA, A., F. AMEZCUA-LINARES y J. W. DAY, Jr., Estuarine Perspectives. Fish community structure and function in Términos Lagoon, a tropical estuary in the southern Gulf of México. In: V. Kennedy (Ed.). Academic Press Inc., NY: 1980. 465-482. YÁÑEZ-ARANCIBIA, A. y L. A. LARA-DOMINGUEZ, An. Inst. Cienc. del Mar y Limnol. Dinámica ambiental de la boca de Estero Pargo y estructura de sus comunidades de peces en cambios estacionales y cielos de 24 hrs, Laguna de Términos, sur del Golfo de México. Univ. Nal. Autón. México, 1983. 85-116. 10 (1): YÁÑEZ-CORREA, Batimetría, salinidad, temperatura y distribución de los sedimentos recientes de la Laguna de Términos, Campeche, México. Bol. Inst. Geol. Univ. Nal. Autón. México, 1963. 147. 67 (1):
|