|
ESTIMACIÓN DEL TAMAÑO DE LA
POBLACIÓN, CRECIMIENTO Y MORTALIDAD DE LOS JUVENILES DE PENAEUS
SETIFERUS (LINNAEAUS, 1767) MEDIANTE MARCADO RECAPTURA EN LA LAGUNA CHACAHITO,
CAMPECHE, MÉXICO
Trabajo recibido el 27 de mayo de 1985 y aceptado
para su publicación el 27 de agosto de 1985.
ADOLFO GRACIA G. y LUIS A. SOTO
Universidad Nacional Autónoma de México. Instituto
de Ciencias del Mar y Limnología. Contribución 464 del Instituto
de Ciencias del Mar y Limnología.
Se liberaron 2,716 individuos de
Penaeus setiferus(80—122 mm de longitud total)
marcados con dos colorantes vitales y cintas vinílicas en la Laguna de
Chacahito, Camp. Los organismos recuperados presentaron incrementos de longitud
total en un intervalo de 0.44 a 1.5 mm/día, con una tasa promedio de
0.83 mm/día para los dos sexos. No se encontró diferencia
significativa (P >0.05) en el crecimiento por sexo. La constante de
crecimiento (K) de la curva de von Bertalanffy estimada a partir de los
juveniles de ambos sexos fue 0. 1942. Se estimaron las tasas
instantáneas de mortalidad diaria total (Z= 0.0455-0.0481), por Pesca
(F=0.0167) y Natural (M= 0.0288-0.0314) mediante el análisis de
frecuencia de tallas y número de recapturas. La mortalidad natural
estimada es relativamente alta comparada con otros registros para P.
setiferus en el Golfo de México. El tamaño medio de la
población se estimó en 0.619 ± 0.298 x 106 ind. La
densidad poblacional promedio alcanzó un valor de 0.124 ± 0.06
ind./m² con una biomasa de 0.62 ± 0.3 g/ m² . La baja
densidad y la alta mortalidad de P. setiferus ocasionan su
baja produción, la cual podría incrementarse atenuando la
presión de los factores bióticos
(depredación-competencia).
Marked juvenile
(2,716) ofPenaeus setiferus (80-122 mm total length) were
released into Chacahito Lagoon, Camp. Growth in total length of shrimp
recovered was between 0.44 to 1.5 mm per day. Mean growth rate for both sexes
was 0.83 mm/day. No significant differences (P > 0. 05) were found between
male and female growth. von Bertalanffy's growth coefficient (K) was 0. 1942.
Number of recaptures and size frequency analysis were applied to estimate daily
instantaneous rates of Total mortality (Z = 0.0455-0.0481), Fishing mortality
(F = 0.0167) and Natural mortality (M = 0.0288-0.0314). Natural mortality was
higher than other values reported in the northern Gulf of Mexico. The estimated
population size averaged 0.619 ± 0.298 x 106 individuals. Population
density was 0.124 ± 0.06 ind./m² which represented 0.62 ±
0.3 g/m² of biomass. High mortality and low density of P.
setiferus cause low production in Chacahito Lagoon. Shrimp production
may be increased by controlling predation and competition in the
area.
El camarón blanco,Penaeus setiferus (Linnaeus, 1767) es la tercera especie de importancia comercial explotada en el Banco de Campeche, cuya captura representa aproximadamente 20% de la producción anual de dicha área (Soto et al., 1982). P. setiferus permanece parte de su ciclo de vida en el interior de lagunas costeras y estuarios, donde lleva a cabo su desarrollo de postlarva a juvenil, para después emigrar al ambiente marino e incorporarse a la población adulta. La Laguna de Términos situada frente al Banco de Campeche constituye un área de crianza y crecimiento para el camarón blanco. La inmigración de postlarvas planctónicas se realiza a través de las bocas de Puerto Real y del Carmen que comunican a la Laguna con el Golfo de México (Arenas- Mendieta y Yáñez-Martínez, 1981; Gracia y Soto, en prensa). Las fases juveniles de P. setiferus se concentran en la parte suroeste de la Laguna de Términos (Signoret, 1974; Paulino, 1979; Gracia y Soto, en prensa); en la cual se ven sujetos a una pesquería artesanal ilegal. Actualmente el conocimiento de las poblaciones de P. setiferus en los sistemas estuarinos del Golfo de México es limitado. Sin embargo, se requiere mayor información sobre los parámetros poblacionales de las fases tempranas del ciclo de vida de esta especie para el manejo adecuado del recurso. Al respecto, existe literatura sobre aspectos de inmigración de postlarvas planctónicas (Arenas- Mendieta y Yáñez-Martínez, 1981) y aspectos biológicos y pesqueros en la Laguna de Términos y las poblaciones adultas del Banco de Campeche (Signoret, 1974; Fuentes et al., 1976; Schultz-Ruíz y Chávez, 1976; Paulino, 1979; Soto, 1979; Sotoet al., 1981; Gracia y Soto, en prensa; Aguilar Sierra, 1985). El objetivo principal del presente estudio es el estimar el crecimiento individual, la mortalidad y el tamaño de la población de P. setiferus en un subsistema estuarino adyacente a la Laguna de Términos mediante el empleo de técnicas de marcado y recaptura. ÁREA DE ESTUDIOEl experimento de marca recaptura se realizó en la Laguna de Chacahito localizada en el margen continental de la Laguna de Términos a una distancia de 40 km de Ciudad del Carmen entre los 91º 24.6´-91°25.8' de longitud O y 18°30.7' – 18°31.8' de latitud N (Fig. 1). La Laguna de Chacahito tiene un área aproximada de 500 ha, una profundidad media de 1.20 m, el tipo de sedimento es limo arcilla y se comunica a la Laguna de Términos mediante un canal angosto (~ 10m) cuya profundidad máxima alcanza 1.4 m. El subsistema estuarino es oligohalino con temperatura entre 29° y 31°C en el período de lluvias. El nivel medio de la marea presenta una diferencia de 0.9 m con respecto a la Boca del Carmen. Tanto el canal, como la laguna están rodeados de vegetación, entre la cual destaca el manglar rojo Rhizophora mangle. La Laguna de Chacahito recibe la descarga de tres arroyos, además de la escurrentía en el margen sur, que se presenta en el periodo de lluvias. Durante el periodo de estudio una red fija experimental ("tapo") perteneciente al Centro de Investigaciones Pesqueras del Instituto Nacional de la Pesca en Cd. del Carmen se encontraba instalda en la parte norte de la Laguna de Chacahito sobre el canal de comunicación a la Laguna de Términos. MATERIAL Y MÉTODOS
CAPTURA Y MANEJO DE ORGANISMOSLas actividades de marcado de juveniles de P. setiferus se realizaron en el margen continental de la Laguna de Términos en las cercanías del canal de acceso a la Laguna de Chacahito, del 24 al 30 de agosto de 1983 (período de lluvias) (Fig. 1). Los organismos se mantuvieron en un sistema abierto de acuarios compuesto por dos tanques cilíndricos de fibra de vidrio (1.20m de diámetro y 0.80m de altura) y cajas de plástico, (0.70 x 0. 40m de base y 0. 35 m alto). El agua proveniente de la Laguna de Términos se mantuvo en circulación constante mediante una bomba de 1/ 4 H. P. alimentada por un generador portátil de 1,000W. Los juveniles deP. setiferus se capturaron con una atarraya en el "tapo" y se transportaron a los acuarios en contenedores de plástico con agua de la laguna. Con el fin de evitar mortalidad debido al manejo, el traslado a los acuarios se hizo rápidamente en cajas de plástico en las que se mantuvo una densidad de 50 individuos. 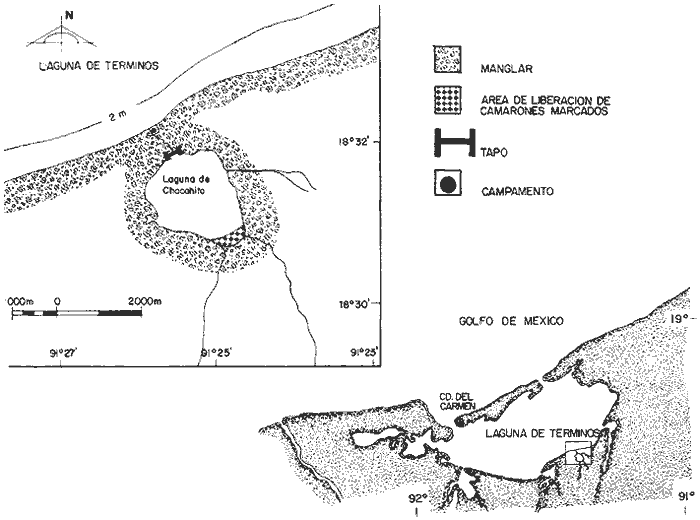 Fig. 1 Localización del área de estudio. MARCADO Y LIBERACIÓNLos camarones se transfirieron en pequeños grupos de los acuarios a charolas con 6 cm de agua dispuestas sobre la mesa de trabajo. Cada uno de los organismos fue identificado, medido en su longitud total (L.T.), sexado y posteriormente marcado y regresado a los acuarios de recuperación. Se emplearon dos métodos de marcado: tinción por inyección de colorantes vitales y cintas vinílicas numeradas. El primero de éstos no permite una identificación individual de los organismos, por lo cual se seleccionaron dos grupos de tamaño uniforme con base en las modas presentes en la población (Fig. 2). De esta forma los Juveniles en el intervalo 75-84mm de longitud total se tiñieron con azul Chicago ("Chicago blue") y aquellos del intervalo 95-104 mm de L. T. con verde rápido ("Fast green"). Las concentraciones de colorantes utilizadas para el azul Chicago y verde rápido fueron de 0.4 g/ 100 ml y 0.6 g/ 100 ml de agua destilada, respectivamente; ensayos realizados en laboratorio indican que estas concentraciones presentan índices bajos de mortalidad y mayor permanencia en los tejidos de los organismos (Araujo, 1984). La inyección del tinte (0.01—0.03 ml) se efectuó mediante una jeringa de pipeteo continuo de 1 ml a través de la membrana articular lateral entre el quinto y sexto segmento abdominal. El colorante se fija posteriormente en las branquias donde permanece por un período de varias semanas (Costello, 1964; Neal, 1969). El marcado con las cintas vinílicas numeradas se realizó según el método de Marullo et al. (1976). La cinta se insertó en individuos mayores de 85 mm de L.T. entre el primero y el segundo segmento abdominales con una aguja previamente sumergida en una mezcla de antibiótico y petrolato. Después de insertada la cinta, la aguja fue retirada y el camarón introducido en los acuarios de recuperación. 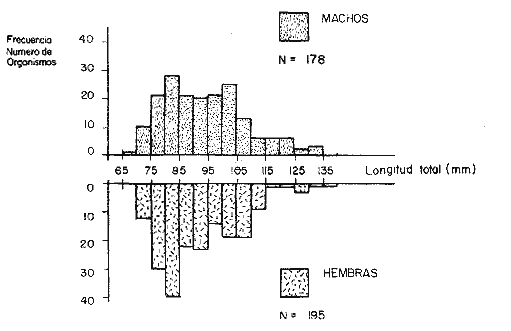 Fig. 2. Estructura de la población de juveniles de P. setiferus de la Laguna de Chacahito capturados en el "tapo". La liberación de los individuos marcados se efectuó en la parte sur de la Laguna de Chacahito, después de 4-6 hrs de observación de los organismos. Algunos autores han extendido este período hasta 24 horas (Klima, 1964; Berry, 1967; Klima, 1974); sin embargo, otros señalan periodos de recuperación hasta de 1 hora para otras especies (Lhome, 1973; Le Reste y Marcille, 1976 vide: García y Le Reste, 1981; Penn, 1975). En el caso de P. setiferus en la Laguna de Términos, Araujo (1984) indica que en 6 horas los camarones recobran su actividad normal. La recuperación de los organismos marcados se llevó a cabo mediante las capturas del arte fijo en el canal de acceso y muestreos con red de arrastre en el interior de la Laguna de Chacahito 15 y 30 días después del período de liberación. ANÁLISIS DE LOS DATOSLa información de las recapturas se procesó de acuerdo a los métodos de Gulland y Holt (1959), Garrod (1963) y Paulik (1983) para la estimación de tasas de crecimiento de recapturas en intervalos de tiempo desiguales. La tasa de mortalidad se estimó a través del análisis de la distribución de frecuencias y el número de marcas recapturadas. Para el tamaño de la población se utilizó el índice de Petersen modificado por Bailey (1951) y el método de mínimos cuadrados de Paloheimo (1963) para poblaciones explotadas comercialmente. Los intervalos de confianza al 95% de cada estimación se calcularon de acuerdo a las fórmulas correspondientes (Seber, 1973). Las capturas obtenidas en el "tapo" se registraron en peso por lo cual fue necesario estimar el número de individuos a partir de la estructura de la población y una regresión de longitud total–peso. RESULTADOS
COMPOSICIÓN POR TALLAS DE LA POBLACION MARCADALa figura 2 ilustra la composición por tallas de los juveniles capturados por el "tapo" en el momento de emigrar hacia la Laguna de Términos. En esta parte de la población sobresalen dos modas en los intervalos de clase 80 — 84.9 y 100—104.9 mm de L. T. para ambos sexos. Organismos menores de 65 mm de L.T. no fueron detectados en el "tapo". En los muestreos con red de arrastre se capturaron tallas pequeñas hasta de 35 mm de L.T.; sin embargo, el intervalo de clases de 35 a 70 mm de L.T. no constituyó más del 8% de los organismos obtenido en cada período de muestreo. Entre los factores responsables de este resultado se encuentran la selectividad de la red y el comportamiento de los estadios iniciales de juveniles. La proporción de sexos en la población sujeta a estudio fue de 1:1. TOTAL DE ORGANISMOS MARCADOS Y RECAPTURADOSSe liberaron 2,716 individuos en buenas condiciones de los cuales, 1,028 fueron marcados con azul Chicago, 756 con verde rápido y 932 con cintas vinílicas numeradas. La mortalidad durante el período de marcado osciló entre el 10 y 14% del total liberado (Tabla 1). El número de recapturas fue de 36 (1.3 % del total liberado) y se obtuvieron mediante el arte fijo. Los porcentajes de recaptura para cada tipo de marca, así como el período máximo de recaptura se muestran en la Tabla 1. Los porcentajes más altos corresponden a los organismos marcados con azul Chicago y los más bajos a aquellos con cintas vinilicas. La proporción de sexos en los organismos recuperados aumentó a 1.6 hembras por cada macho (22:14). 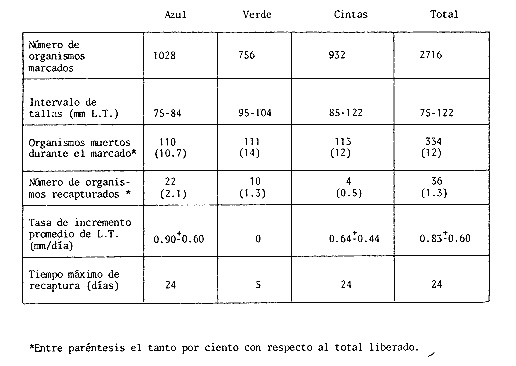 TABLA 1 NÚMERO DE ORGANISMOS MARCADOS Y RECAPTURADOS CRECIMIENTOLos organismos recapturados en períodos cortos (<5 días) no presentaron incrementos en la longitud, por lo cual el número utilizado para estimar las tasas de crecimiento se redujo a 16 individuos. Las tasas de incremento diarias obtenidas de estos organismos (12 marcas de azul Chicago y 4 cintas vinilicas) fluctuaron entre 0.44 y 1.5 mm de L.T. en períodos de 6 a 24 días, desde un intervalo de longitud inicial de 80-90 mm L.T. El promedio de estas tasas fue mayor para las marcas de azul Chicago (Tabla 1); es conveniente indicar, que la longitud inicial promedio de los organismos teñidos con azul Chicago fue mayor (80 mm de L.T.) que la de recapturas con cintas vinílicas (87 mm. L.T.). La proporción de sexos de los juveniles recapturados que mostró crecimiento se mantuvo 1: 1. El incremento diario promedio resultó mayor para los machos (1.01 mm/ día L.T.) en comparación con hembras (0.66 mm/ día L.T.). Esta diferencia no fue significativa (prueba det, P>0.05). La aplicación del método de Gulland y Holt (1959) a los datos de juveniles de ambos sexos permitió la estimación de los valores de K y Loo a partir de un intervalo corto de tallas (Fig. 3; Tabla 2). Para evitar el sesgo en el cálculo de crecimiento debido a las limitantes del método anterior, los datos fueron analizados mediante el método "forzado" de Gulland y Holt que considera los incrementos medios relacionados con la longitud asintótica (Pauly, 1983). En la figura 3, se ilustran las regresiones calculadas por los dos métodos para ambos sexos. En la Tabla 2 se presentan los valores de K para cada sexo, obtenidos mediante el método "forzado" de Gulland y Holt, con las longitudes asintóticas (Loo) de P. setiferus señadas par la Sonda de Campeche (Schultz-Ruiz y Chávez, 1976). Las estimaciones de —to. realizadas de acuerdo al método señalado por Gulland (1971) se muestran también en la Tabla 2. ESTIMACIÓN DE LAS TASAS INSTANTÁNEAS DE MORTALIDAD (Z, F, M)Los juveniles de P. setiferus distribuidos en la Laguna de Chacahito estan sujetos sólo a la mortalidad natural. La mortalidad por pesca se presenta en el momento de la emigración cuando son capturados por el "tapo". Por esta razón la estimación de la mortalidad en los juveniles de esta área está influenciada por dos factores relacionados: la emigración y la alta mortalidad por pesca debida ala eficiencia del "tapo". Estos factores son más marcados en los organismos de tallas grandes, pero puede afectar a los individuos de tallas superiores a 80 mm de L.T. (Fig. 2). 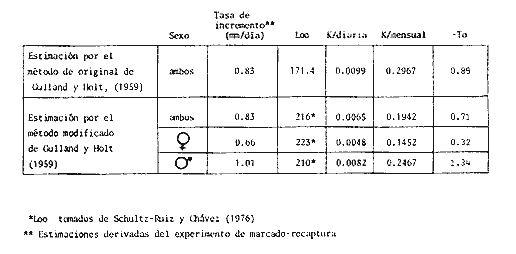 TABLA 2 ESTIMACIONES DI, LA TASA DE CRECIMIENTO DE P. SETIFERUS 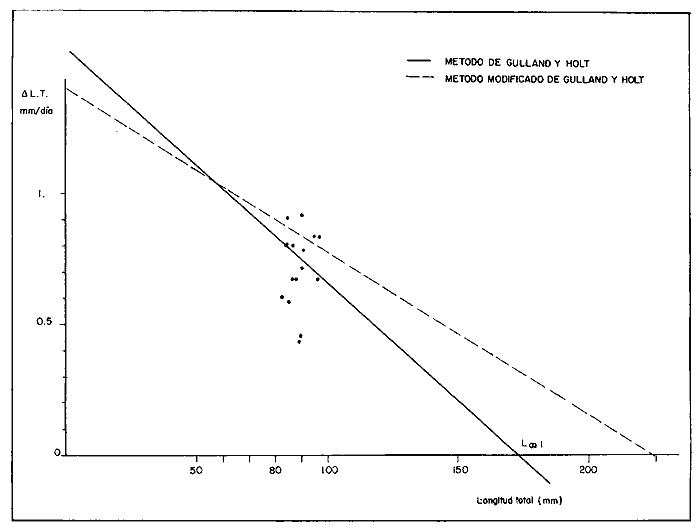 Fig. 3. Crecimiento de P. setiferus estimado por el método de Gulland y Holt (1959) normal y modificado. Para estimar la tasa de mortalidad de los juveniles a partir de la estructura de la población capturada en el "tapo" (Fig. 2) es necesario eliminar las tallas grandes donde la curva se hace asintótica y asumir que la muestra es un reflejo de la población residente dentro de la Laguna de Chacahito. Con base en estos dos ajustes, la curva de mortalidad para el intervalo de 80–120 mm de L.T. de individuos juveniles de camarón blanco presenta una tasa de mortalidad total diaria de —0.0455 e indica un valor semanal de —0.319 (Fig. 4). Las estimaciones de la mortalidad a partir de las recapturas y el número de días transcurridos después de la liberación. (Fig. 4) dan un valor de Z diario de —0.0481 y —0.337 semanal. (Tabla 3). Las tasas instantáneas de mortalidad por pesca (F) y de mortalidad natural se estimaron mediante la relación de Ricker (1958) E = (F/ Z)( 1 —e -Zt); donde Z es la tasa de mortalidad total obtenida y la tasa de explotación (E) representa la razón del número de marcas recapturadas y liberadas. Los resultados indican una mortalidad por pesca diaria de —0.0167 y una mortalidad natural de —0.0288 a —0.0314 de acuerdo a la estimación de Z que se utilice (Tabla 3). TAMAÑO DE LA POBLACIÓNLa estimación del tamaño de la población se realizó mediante el método de Petersen modificado por Bailey (1951), el cual evita el sesgo en las estimaciones según la siguiente ecuación: N = r (n + 1) ÷ (m + 1) donde N es el estimado del tamaño de la población, r es el número de individuos marcados, n es la captura o tamaño de la muestra y m el número de marcas recapturadas. La varianza y el error estándar de la población fueron estimadas mediante la fórmula correspondiente (Poole, 1974; Begon, 1979). 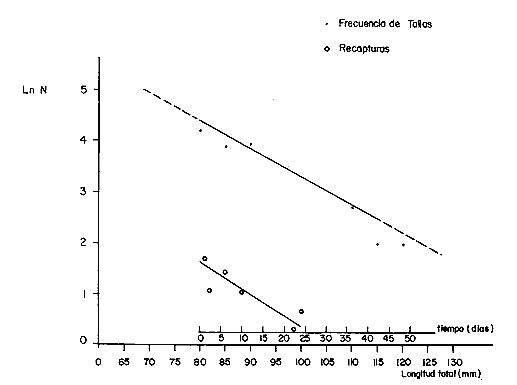 Fig. 4. Estimación de la mortalidad de P. setiferus mediante el logaritmo natural del número de organismos por clase de talla y el número de recapturas. Los supuestos y ajustes sobre los cuales se basó la aplicación de este método son los siguientes: 1) Los individuos marcados con tintes y cintas vinilicas se mezclan al azar con la población residente en la Laguna. 2) Los organismos con y sin marca presentan el mismo patrón de emigración hacia la Laguna de Términos y por lo tanto la misma vulnerabilidad al arte fijo ("tapo"). 3) El comportamiento y la mortalidad no se ven afectadas por la marca. Araujo (1984) señala que en condiciones de laboratorio la mortalidad por efecto de marcado es de 36% para azul Chicago, 5 % para verde rápido y 11 % para cintas vinílicas. Otros autores señalan cifras de mortalidad más altas (Costello y Allen, 1962; Howe y Hoyt, 1982); sin embargo, la estimación del número poblacional en este caso, se efectuó con reducción en el número de marcas liberadas de acuerdo a los valores mencionados por Araujo (1984). 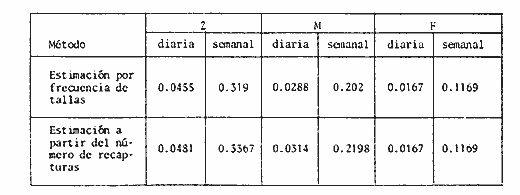 TABLA 3. TASAS DE MORTALIDAD INSTANTANEA DE P. SETIFERUS EN LA LAGUNA DE CHACAHITO 4) La mortalidad por depredación afecta de igual forma a los organismos con y sin marca. Algunos autores (Costello y Allen, 1962; Howe y Hoyt, 1982) señalan mayor vulnerabilidad a la depredación en individuos marcados con cintas. 5) El reclutamiento a la población durante el experimento de marca recaptura es nulo. Este supuesto está apoyado por la existencia de cohortes mensuales (Gracia y Soto, en prensa) cuya incorporación a la población estudiada no se presenta durante el período del experimento. 6) La emigración de organismos del área de estudio no altera la fracción r/ N, dado que los juveniles marcados y no marcados abandonan el sitio en la misma proporción. Debido a este supuesto y al señalado en el inciso 5, la estimación por el método de Petersen corresponde al tamaño inicial de la población (Ni.). El número total de los tres tipos de marcas en la población se redujo a 2205 después de incluir la mortalidad causada por el marcado de los organismos (inciso 3). Con este ajuste se estimó el tamaño de la población y error estándar mediante el método de Bailey (1951) para cada uno de los períodos en que se obtuvieron recapturas en el "tapo". Las variaciones de estos estimados se presentaron en un intervalo de 0.391 a 1.534 x 106 ind. y mostraron una tendencia a disminuir conforme se incrementó el período de recuperación. La media de los valores obtenidos indica una población inicial de 1.056 ± 0.535 x 106 ind. (5,298 ± 2,268 kg). Al incluirse la tasa de mortalidad total en juveniles para estimar el número efectivo de organismos marcados en cada período, la estimación media del tamaño de la población por el método de Bailey disminuye notablemente a 0.666 ± 0.584 x 106 ind. (3,341 ± 2,915 kg) (Tabla 4). La aplicación del método de mínimos cuadrados de Paloheimo (1963), para poblaciones comercialmente explotadas, donde la segunda muestra del método original de Petersen consiste en una serie de muestras que son removidas en forma permanente de la población, ofrece una estimación de 1.083 x 106 ind. El intervalo de confianza de 95% para este estimado se encuentra entre los siguientes límites 0.665 y 3.415 x 106 ind., de acuerdo a la fórmula señalada por Paloheimo para este caso (Tabla 4). 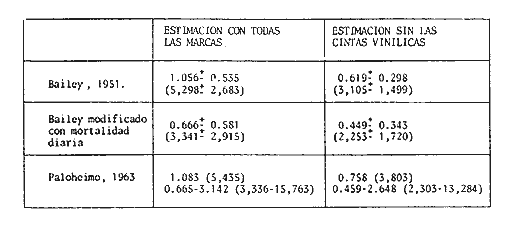 TABLA 4 TAMAÑO DE LA POBLACIÓN DE P. SETIFERUS EN LA LAGUNA DE CHACAHITO, CAMP. NÚMERO DE INDIVIDUOS X 10^6 Y BIOMASA EN KG ENTRE PARÉNTESIS. La probabilidad de que los individuos marcados con cintas vinílicas (inciso 4) estén sujetos a mayor depredación, puede introducir sesgo en la estimación del tamaño de la población. El aumento de la depredación sobre los organismos con cintas no se comprobó directamente en este estudio; sin embargo el porcentaje de recapturas para esta marca fue el menor de los tres tipos utilizados (Tabla 1). El análisis de estas recapturas mediante una tabla de contingencia mostró diferencias significativas (P < 0.00 1) entre la capturabilidad y/o tasas de supervivencia de las marcas; por lo cual se hicieron otras estimaciones, del tamaño de la población sin incluir los organismos liberados y recapturados con cintas vinílicas. Los resultados muestran una disminución sustancial del valor medio de la población en los distintos métodos empleados (Tabla 4). 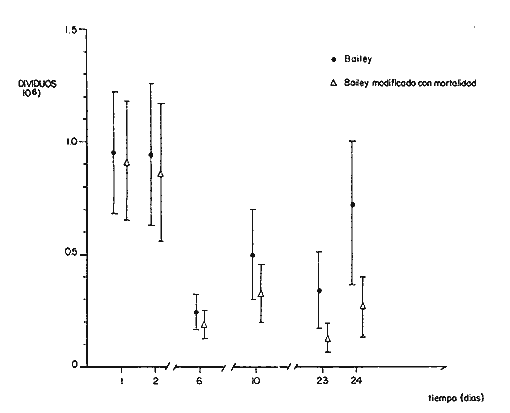 Fig. 5. Representación gráfica de las estimaciones del tamaño de la población de P. setiferus (± error estándar) en la Laguna de Chacahito por el método de Bailey (1951). Las estimaciones obtenidas para las dos modalidades del método de Bailey (1951) presentan variaciones para cada período de la recaptura y además diferencias en el tamaño de la población estimado. Cuando se incluye la mortalidad disminuye el valor de la población inicial estimado, pero los intervalos de confianza aumentan a más de 50% de esta población (Fig. 5). DISCUSIÓN Y CONCLUSIONES
CRECIMIENTOEntre los pocos antecedentes de estudios sobre el crecimiento de juveniles de P. setiferus en la Laguna de Términos, se halla el trabajo de Paulino (1979), donde el incremento máximo de longitud en el intervalo 70-90 mm de L.T. da un valor de 0.66 mm/ día, obtenido mediante análisis de la secuencia de modas. Este valor es inferior a la tasa media estimada mediante el experimento de marcado recaptura; sin embargo, si se realiza un análisis de modas por el método de Battacharya (1967) con la estructura de tallas de las capturas realizadas en el interior de la Laguna de Chacahito, el incremento medio es simílar (0.63 mm/ día). Esta diferencia resulta de la aplicación del método de análisis de la frecuencia de tallas que puede dar estimaciones sesgadas por la sobreposición de clases (Westhreim y Ricker, 1978; Bartoo y Parker, 1983) y la variabilidad de las tasas de crecimiento en una clase que no son detectadas por este método. En la estimación de crecimiento por el método de marcado se podría esperar un sesgo debido al efecto de los tintes y las cintas sobre el organismo. Araujo (1984) señala una disminución en el crecimiento de juveniles de camarón blanco en condiciones experimentales, de 64 % (significativa P = 0. 05) y 11 % (no significativa) para individuos marcados con azul Chicago y cintas vinilicas respectivamente. Esta disminución puede atribuirse al manejo a que se somete a los organismos en cautiverio (Penn, 1975; Menz y Blake 1980). La aproximación de las tasas obtenidas por los dos métodos empleados en este estudio (análisis de modas y marcado recaptura), sugiere que el efecto del tinte azul Chicago sobre los organismos en condiciones naturales no afectó la estimación de crecimiento. La tasa de crecimiento estimada por marcado-recaptura es menor a las indicadas por Aguilar Sierra (1985) en Laguna de Términos (1.0-1.2 mm/ día) durante el período de lluvias, cuando se presentan los valores más altos de crecimiento (Gracia y Soto, en prensa). Las causas responsables de esta diferencia pueden atribuirse al intervalo de tallas empleado en las estimaciones y a variaciones ambientales anuales, entre las cuales la densidad de organismos tiene un efecto considerable sobre el crecimiento (Edwards, 1977). Las tasas de crecimiento de verano estimadas para el camarón blanco en los estuarios del Norte del Golfo de México varían entre 0.6 y 2.2 mm/ día (Williams, 1955; Gunter, 1956; Loesch, 1965; kLIMA, 1974). Los valores estimados para la Laguna de Chacahito se encuentran dentro de dicho intervalo, pero su valor es bajo comparado con la media de crecimiento obtenido por Klima (1974) mediante técnicas de marcado (1.25 mm/ día). De acuerdo a la tendencia de variaciones geográficas en las tasas de crecimiento (Parrack, 1979) los juveniles de P. setiferus en esta área deberían presentar una tasa mayor. No obstante, la tasa es menor o similar si se consideran los valores proporcionados por Aguilar Sierra (1985). Este aspecto presenta interrogantes interesantes en cuanto al efecto de los parámetros ambientales; en especial la temperatura y la presión de los factores bióticos tales como la competencia y la depredación. La comparación de las tasas de crecimiento de machos y hembras señala que no existe diferencia significativa (P >0.05) en estado juvenil. Esto permite suponer que las tallas alcanzadas por los adultos (Schultz-Ruíz y Chávez, 1976) estén relacionadas con un crecimiento diferencial después del proceso de maduración. Este proceso está relacionado con la estrategia de crecimiento del organismo y su tratamiento requiere de mayor información sobre el comportamiento de muda (Incremento y Frecuencia) en función del proceso reproductivo (Hartnoll, 1983). Por otro lado, los parámetros de la ecuación de von Bertalariffy (Tabla 2) obtenidos mediante el método de Gulland y Holt (1959), se aproximan a los obtenidos por Paulino (1979); pero estos no reflejan el comportamiento adecuado de la curva total, ya que se basan en intervalos pequeños de crecimiento total del organismo, donde el modelo adecuado es de tipo lineal. Por lo anterior, el método "forzado" de Gulland y Holt (Pauly, 1983) ofrece estimaciones menos segadas del parámetro K, cuyo valor mensual para ambos sexos (Tabla 4) es cercano al señalado por Schultz-Ruíz y Chávez (1976) en adultos (0.2066). Las estimaciones del coeficiente para cada sexo muestran variación respecto a las de estos dos últimos autores (hembras: 0.1833; machos; 0.2264), debido a la amplitud del intervalo de tallas utilizado. MORTALIDADLas tasas de mortalidad obtenidas a partir del número de recapturas, fueron mayores que las estimaciones realizadas mediante el análisis de frecuencia de tallas (Tabla 3). Por lo general las estimaciones de mortalidad derivadas de información de marcado -recaptura tienden a producir sesgos en estos parámetros por las siguientes razones: a) aumento en la mortalidad después de la liberación por efectos del marcado, b) pérdidas de organismos marcados por emigración del área, c) reportes incompletos de los organismos recapturados. Como ya se ha mencionado en este trabajo la mortalidad por efectos de marcado puede considerarse mínima; sin embargo, la mortalidad por depredación es factible que se incremente en los organismos marcados con cinta (Costello y Allen, 1961; Blake y Menz, 1980; Howe y Hoyt, 1982). En el interior de la Laguna de Chacahito se observó la presencia de peces(Centropomus, Arius, Epinephelus), crustáceos (Callinectes similis y C. sapidus) y aves que pueden ejercer un efecto depredador sobre el camarón. Otro factor que puede afectar la estimación de las tasas de mortalidad es la emigración de los organismos marcados, sólo que ésta influye sobre los organismos con y sin marca y además sobre los dos métodos utilizados para calcular la mortalidad. Con respecto al reporte incompleto de marcas no se logró confirmar la eficiencia en la recuperación de individuos marcados. Aunque los tres incisos anteriores no se satisfacen enteramente, la diferencia entre las tasas de mortalidad de recapturas y de frecuencia de tallas no es grande. Esto sugiere que la posibilidad de depredación selectiva sobre organismos marcados con cintas vinilicas no afectó la estimación de la mortalidad. No obstante, que los valores de mortalidad puedan presentar sesgos, principalmente por causa de la emigración, resultan de utilidad como indicadores de la mortalidad por pesca. Un aspecto importante es la obtención de la mortalidad natural de P. setiferus,en esta área, sobre la cual no existe registro previo en la fase juvenil. Su estimación provee un punto de referencia para examinar M en función de diferentes condiciones ambientales en el sistema de la Laguna de Términos (estiaje, lluvias, tormentas). Con motivos de comparación con otras estimaciones de mortalidad deP. setiferus en el Golfo de México, se realizó la Tabla 5, en la cual se incluyen los estimados de este estudio (Tabla 3). Las tasas son muy variables; las obtenidas en este trabajo son más altas que la mayoría de las estimaciones y se ubican en el intervalo registrado para la Laguna de Términos (Paulino, 1979; Aguilar Sierra, 1985). 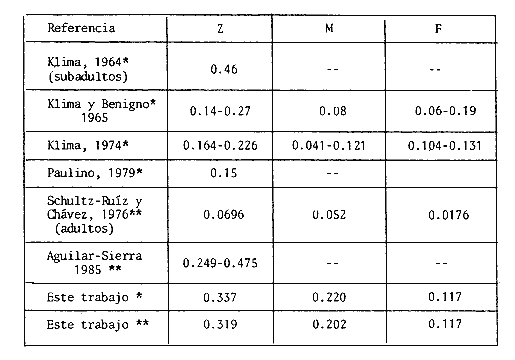 IMAGEN TABLA 5 TASAS INSTANTÁNEAS, DE MORTALIDAD SEMANAL (Z,M, F) DE P SETIFERUS OBTENIDAS MEDIANTE ESTUDIOS DE MARCADO Y RECAPTURA (*) Y ANÁLISIS DE FRECUENCIA DE TALLAS EN LA POBLACIÓN (**). (LOS VALORES SE AJUSTARON A UN PERIODO SEMANAL, INDEPENDIENTEMENTE DEL TIEMPO ORIGINAL). La notable diferencia entre los valores de mortalidad natural deP. setiferus en la Laguna de Chacahito con los del norte del Golfo de México y la consistencia de las estimaciones de M, obtenidas por los métodos empleados en este trabajo eliminan la posibilidad de que los cálculos esten influenciados por variaciones en los procedimientos de marcado y estimación utilizados. El origen de esta diferencia radica, más bien, en las presiones medio-ambientales a que estan sujetas las poblaciones de camarón blanco en cada localidad. Entre éstas, los factores bióticos (depredación) representan un papel preponderante en un ambiente tropical (Paine, 1966; Sanders, 1968) como la Laguna de Chacahito. Por otro lado, las estimaciones de mortalidad natural de este trabajo coinciden con el período de mayor crecimento en el año (Gracia y Soto, en prensa; Aguilar, 1985), que a su vez está relacionado con periodos de intensa actividad de los camarones y de mayor riesgo y exposición a mortalidad por causas naturales. Al respecto Amezcua Linares y Yáñez Arancibia (1980) señalan, para varios sistemas fluvio lagunares adyacentes a la Laguna de Términos, incrementos importantes en las poblaciones de peces al final del período de lluvias. Esto representa una mayor presión de depredación e influye sobre los valores de mortalidad natural que P. setiferus muestra en el período de precipitación pluvial. La mortalidad en camarones adultos de la Sonda de Campeche (Schultz-Ruíz y Chávez, 1976) disminuye en comparación con la fase juvenil, de acuerdo al comportamiento general de la curva de mortalidad que presentan estos organismos. Las variaciones en las estimaciones de mortalidad responden además de las condiciones medioambientales a las tasas de explotación a que está sujeta la población. En la Laguna de Términos se presenta una pesquería artesanal ilegal cuyas variaciones en el esfuerzo pesquero se desconocen debido a lo cual las tasas de mortalidad que existen se refieren a Z. Las estimaciones de mortalidad por pesca de este trabajo no pueden ser aplicadas a los organismos juveniles en la Laguna de Términos ya que se utilizan artes y esfuerzos de pesca distintos. Sin embargo, sería interesante comparar con los niveles de mortalidad por pesca a que se ven sometidos los camarones blancos durante el proceso de emigración hacia el mar; ya que existe la necesidad de obtener estimaciones precisas de los componentes de la mortalidad total en el período de permanencia en las lagunas para la aplicación correcta de modelos de producción. TAMAÑO DE LA POBLACIÓNEl número reducido de recapturas puede ocasionar sesgo en la estimación deN, debido a que la precisión del estimado depende directamente de m. De acuerdo a Robson y Regier (1964) el sesgo es estadísticamente despreciable en un nivel de 95 %, ya que el producto de rn es mayor que 4N en cada caso. El problema principal reside en la consistencia de las suposiciones del método y cuanto se desvían de la realidad. La discrepancia más importante es la diferencia en la capturabilidad y/o tasas de supervivencia de las marcas, probablemente debido a una depredación selectiva, lo que origina una sobreestimación de la población cuando se emplea el conjunto total de marcas. Debido a esto las estimaciones realizadas sin las cintas vinilicas presentan menor sesgo, aunque también son susceptibles de una sobre-estimación en particular por reportes incompletos de los organismos recapturados. No obstante que el riesgo de sobreestimación de N sea mayor que el de subestimación, el modelo empleado es válido y por lo tanto proporciona estimados preliminares confiables del tamaño de la población. Entre estas estimaciones, la más precisa es la obtenida por el método de Bailey (1951) sin modificar que se sobrepone con el intervalo de la estimación que incluye mortalidad diaria (Tabla 4). La diferencia entre la precisión de estos valores se debe a la disminución de organismos marcados en la población por mortalidad conforme aumenta el período de recaptura. Con respecto al estimado de Paloheimo (1963), este presenta un límite superior muy alto poco confiable. Las variaciones en los estimados deben considerarse para la afinación de N en estudios posteriores; pero para este trabajo la estimación simple de Bailey se acepta como la más adecuada en forma preliminar. Por otro lado, esta estimación media de la población inicial coincide con la media de los valores obtenidos para cada tipo de marca por separado (0.685 x 10 6 ind.). Las fluctuaciones en los estimados sugieren una disminución del tamaño de la población de camarones de la Laguna de Chacahito relacionada con la salida de un gran número de organismos en los primeros días de recaptura (Fig. 5). Con base en esta observación se podría esperar que las estimaciones de los primeros períodos reflejaran en mejor forma la magnitud de la población inicial; sin embargo, el lapso transcurrido entre la liberación y estas recapturas puede ser pequeño para que los individuos marcados se mezclen con la población. García (1973, 1974) señala una velocidad media de desplazamiento de P. notialis de 1 a 6 km diarios; la cual de presentarse en el camarón blanco permitiría a los organismos marcados incorporarse rápidamente a la población en proceso de emigración y daría validez a las estimaciones de los primeros períodos de recaptura. Por otro lado, la posibilidad de error en las estimaciones del método de Bailey se incrementa mientras más grande es el intervalo de recaptura debido a la salida de organismos de la laguna. Otro aspecto que influye sobre la estimación de la población es la variación en la actividad de los juveniles, ya que los estimados están basados en los organismos que emigran de la laguna, y como sucede en otros peneidos (García y Le Reste, 1981), este movimiento se presenta con mayor actividad en días cercanos (1, 2 y 23) a la luna llena (Fig. 5). La determinación correcta de estas variaciones requiere de mayor información, por lo pronto es conveniente considerar el tamaño medio de la población del promedio de las estimaciones y las variaciones como indicio de los niveles que puede alcanzar la población. La densidad de biomasa de la población media de juveniles en el intervalo de 80-122 mm de L.T. en la Laguna de Chacahito corresponde a 0. 124 ± 0. 06 ind. m - 2 y 0 - 62 ± 0.3 g m - 2, respectivamente. A pesar de que estos parámetros estimados pertenecen a la temporada de mayor producción (Gracia y Soto, en prensa), los valores son bajos comparados con la producción registrada en experimentos de cultivo y semicultivo de camarón blanco (4.8- 111 g m-²); (Rose et al.,1975; Kitakka, 1981) y con aquellos obtenidos de pesquerías lagunares (10. g peso de abdomen m-²) como el sistema Huizache-Caimanero en la costa del Pacífico (Edwards, 1977). No obstante, las estimaciones de biomasa se encuentran dentro del intervalo (0.6-3.0 g m-²) señalado para P.setiferus en sistemas estuarinos del norte del Golfo de México (Jacob, 1969; Loesch, 1976). Es conveniente resaltar que estas estimaciones corresponden a una población en condiciones naturales, cuya producción se ve afectada por la presencia de organismos depredadores y competidores representados principalmente por peces. La presion de estos factores sobre la población de juveniles de camarón blanco en este ambiente tropical, se refleja en la mortalidad natural relativamente alta que presentan. Con fines de semicultivo la producción de camarón en la laguna se puede incrementar mediante la aplicación de las siguientes consideraciones: a) Control de depredadores y competidores del camarón blanco. La extracción de estos miembros de la comunidad animal de las áreas de semicultivo resulta en un aumento de la supervivencia de los peneidos y en consecuencia de la producción. Rose et al. (1975) señalan que la producción de P. setiferus puede incrementarse de tres a cuatro veces en estanques sin depredadores. b) Aumento en la densidad de organismos.Esto se ve directamente influenciado por el inciso anterior; sin embargo, dado que la densidad estimada para la Laguna de Chacahito es baja, se puede introducir mayor número de postlarvas y/o juveniles pequeños al área sin que esto afecte la tasa de crecimiento de los organismos. Edwards (1977) indica para P. vannamei densidades de 2. 5 ind. m - ² sin disminución del crecimiento; pero para P. setiferus es necesario establecer la densidad máxima que puedan soportar las áreas escogidas para semicultivo sin alterar el proceso de crecimiento. Por último un aspecto importante es el efecto que puede tener el establecimiento de este tipo de semicultivos sobre la fase estuarina deP. setiferus y su repercusión en los niveles de reclutamiento de la población adulta. La conservación del recurso exige una evaluación cuidadosa de las cantidades de postlarvas y/o juveniles utilizables para semicultivo sin afectar el equilibrio de la población y además complementarla con información sobre el comportamiento de la población que haga compatible el desarrollo de la pesquería de altamar y semicultivos. AgradecimientosA Galo Escanero Figueroa jefe del Centro Regional de Investigaciones Pesqueras del Instituto Nacional de la Pesca en Ciudad del Carmen por las facilidades prestadas para el desarrollo del presente trabajo. Al personal que colaboró entusiastamente en las actividades de marcado en el campo. Este estudio se llevó a cabo con el apoyo del Consejo Nacional de Ciencia y Tecnología y el Instituto de Ciencias del Mar y Limnología, a través del proyecto PCMANAL-790334. LITERATURAAGUILAR SIERRA, V.A., Camarones peneidos de la Laguna de Términos, Campeche: Composición, Distribución y Parámetros Poblacionales. Tesis Prof.Fac. Ciencias, Univ. Nal. Autón. de México:1985. 62 p. AMEZCUA-LINARES, F. y A. YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limmol. Univ. Nal. Autón. México, Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces. 1980.69-118.(1): 7 ARAUJO LEAL, M. A., Efectos del marcado con cintas vinilicas y colorantes vitales sobre el crecimiento y mortalidad del camarón blanco Penaeus setiferus (Linneaus, 1767). Tesis Prof. Fac. Ciencias, Univ. Nal. Autón. de México: 1984. 52 p. ARENAS-MENDIETA, M. R. y A. YÁÑEZ MARTINEZ, Patrón anual de inmigración de postlarvas de camarón(Crustácea: Decapoda: Penaeidae), en la Boca de Puerto Real, Laguna de Términos Campeche. Tesis Prof. Fac. Ciencias Univ. Nal. Autón. de México1981 92 p. BAILEY, N. T. J., Biometrika On estimating the size of mobile populations from capture-recapture data. 1951.29-306. 38: BARTO, N. W. y K. R. PARKER, Fish. Bull.Stochastic agefrecuency estimation using the von Bertalariffy growth equation. 1983. 91-96.(1): 81 BATTACHARYA, C. G. Biametrics A simple method of resolution of a distribution into gaussian components. 1967.115-135. (1): 23 BEGON M., Investigating Animal Abundance. University Park Press EUA: 1979,97 p. BERRY, R. J., Dynamic of the Tortugas pink shrimp population. Ph. D. thesis. University of Rhode Island,1967.160p. COSTELLO, T. J., US. Fish. Wildl. Serv. Spec. Sci. Rep. Fish. Field Techniques for stainingrecapture experiments with commercial shrimp.1964. 13 p. (484): COSTELLO, T. J. y D. M. ALLEN, Proc. Gulf and Caribb. Fish. Inst. Survival of stained, tagged and unmarked shrimp in the presence of predators. 1962. 16-20.14: EDWARDS, R. R. C., Estuarine and Coastal Marine Science Field Experiments in growth and mortallty of Penaeus vannamei in a mexican coastal lagoon complex. 1977. 107-121. 5: FUENTES, D., R. G. CASTRO, L. SCHULTZ, R. PORTUGAL y M. OROPEZA, Mem. Simposio sobre Biología y Dinámica Poblacional de Camarones del 8 al 13 de agosto de 1976, Guaymas Son. México Pesquería de Camarón de altamar en el Golfo de México. S.I.C/ Subsecretaría de Pesca, Instituto Nacional de la Pesca. 1976.187-210. 1: GARCÍA, S. Doc. Scient. Centre Rech. Océanogr. Abdija Marquages de Penaeus duorarum en Cote D'Ivoire, resulttats preliminaires: Taux de recapture, Migrations et Croissance. 1973. 29-48.(3): Vol. IV GARCÍA, S. Doc. Scient. Centre Rech. Oceanogr. Abdijan Marquages de Penaeus duorarum motialis en Cote D'Ivoire: II-Migrations et Mortalities.1975.21-44.(1): 6 GARCÍA, S. y L. LE RESTE, FAO Fish. Tech. Pap. Life cycles, dynamics exploltation and management of coastal penaeid shrimp stocks. 1981. 215 p. (203): GARROD, D. F., Spec. Publ. Intern. Comm. Northw. Atlant. Fish. The application of a method for the estimation of growth parameters from tagging data at unequal time intervals. 1963. 258-261. 4: GRACIA, A. y L. A. SOTO. Studies of penaeid shrimp in Terminos Lagoon, Campeche, México. II. Annual fluctuations and production of the shrimp populations (en prensa). GULLAND, J. A., Manual de Métodos para la evaluación de las poblaciones de peces. Editorial Acribia, España1971: 160 p. GULLAND, J. A., y S. J. HOLT, J. Cons. Int. Explor. Mer., Estimation of growth parameters for data at unequal time intervals. 1959.47-49.(1): 25 GUNTER, G. Proc. Gulf and Carib. Fisch. Inst.,Principles of shrimp fishery management.1956.99-106. 8: HARTNOLL, R. G., Papers from the conference on the biology and evolution of Crustacea Australian Museum Memoir Strategies of crustacean growth. Lawry. J. K. (Ed). 1983.121-132. 18: HOWE, N. R., Trans. Am. Fish. Soc. Mortality of juvenile brown shrimp Penaeus aztecas Asociare with streamer tags. 1982.317-325. 111: JACOB, W. J. Observations on the distribution growth, survival and biomass of juvenile and subadult Penaeus aztecus in Southern Louisiana. M. S. Thesis,Lousiana State University: 1969.1-68. KITAKKA, J., Kuwait Bulletin of Marine Science. Large scale production of shrimp for releasing in Japan and in the United States and the results of the releasing programme at Panama city, Florida. Proceedings of the International Shrimp Releasing, Marking and Recruitment Workshop, Farmer A. S. D. Ed. Salmiya State of Kuwait. 1981. 149-163.(2) 25-29 November 1978, KLIMA, E. F. Proc. Gulf and Caribb. Fish. Inst.Mark-Recapture experiment with brown and white shrimp in the northern Gulf of México.1964. 52-64. 16: KLIMA, E. F., Trans. Am. Fish. Soc., A white shrimp Mark Recapture study. 1974. 107-113. (1):103 LE RESTE, L. y J. MARCILLE, Cah. ORSTOM Biologie de la crevette Penaeus indicus H. Milne Edwards à Madagascar: croissance, recrutement, migration, reproduction, mortalité. (Sér. Océanogr.)1976.109-127. (2): 14 LHOME, F., Biologie et dynamique de Penaeus duorarum notialis au Sénégal. Partie 1. Sélectivité. Soc. Sci. Cent. Rech. Oceanogr. Dakar Thiaroye, 1978. 30 p. (63): LOESCH, H. C., Publis. Inst. Mar. Sci. Univ. Tex.Distribution and growth of penaeid shrimp in Mobile Bay Alabama. 1965.41-58. 10: LOESCH, H. C., Gulf. Res. Rep. Shrimp population densities within Mobile Bay.1976.11-16.(2): 5 MARULLO, F., D.A. EMILIANI, C. W. CAILLOEUT y S. H. CLARK Trans. Am. Fish. Soc. A vinyl streamer tag for shrimp (Penaeus spp). 1976.658-663.(6): 105 MENZ, A. y B. F. BLAKE, J. exp. mar. Biol. Ecol. Experiments on the growth of Penaeus vannamei Boone. 1980.99-111 48: NEAL, R. A. FAO Fisheries, Report Methods of marking shrimp. In: Procceedings of the World Scientific Conference on the Biology and Culture of Shrimps and Prawns, Mexico City, Mexico1969. 1149-1165. (57)3: 12-21 june 1978. PAINE, R. T. Amer. Nat. Food web complexity and species diversity. 1966. 65-75100: PALOHEIMO, J. E., Estimation of catchabilities and population sizes of lobsters.J. Fish. Res. Board Canada1963.59-88. 20: PARRACK, M. L. Fish. Bull. Aspects of brown shrimp, Penaeus aztecus, growth in the northern Gulf of Mexico. 1979.827-836.(4): 76 PAULINO, J. M. Datos sobre las poblaciones de camarón blanco Penaeus setiferus Linnaeus en la Laguna de Términos, Campeche. Tesis Prof. Esc. Nal. Cienc. Biol. IPN 1979.65 p. PAULY, D., Doc. Tec. Pesca Algunos Métodos simples para la evaluación de recursos pesqueros tropicales. FAO1983. 49 p. (234): PENN, J. W., Aust. J. mar. Freshwat. Res.Tagging experiments with Western King Prawn Penaeus latisulcatus Kishinouye. I. Survival, Growth, and Reproduction of tagged prawns, 1975.197-211. 26: POOLE, R. W. An introduction to quantitative ecology. Mc Graw-Hill Kogakusha, LTD. Tokyo.1974. 532 p. RICKER, W. E., Bull. Fish. Res. Board Can. Handbook of computation for biological satitistics of fish populations. 1958. 119-300 p. ROBSON, D. S. y H. A. REGIER, Trans. Am. Fish. Soc. Sample size in Petersen mark recapture experiments.1964.215-226. 93: ROSE, D. C., A. H. HARRIS y B. WILSON, Trans. Am. Fish. Soc., Extensive culture of penaeid shrimp in Lousiana salt-marsh impoudments.1975.296-308. (2): 104 SENDERS, H. L. Amer. Nat. Marine benthic diversity a comparative study.1968.243-282. 102: SEBER, G. A. F., The estimation of animal abundance and related parameters. Griffin London1973. 506 p. SCHULTZ-RUIZ, L. E. y E. A. CHÁVEZ, Mém. Simposio sobre Biología y Dinámica Poblacional de Camarones del 8 al 13 de agosto de 1976, Guaymas, Son. México, Contribución al conocimiento de la biología pesquera del camarón blanco (Penaeus setiferus (L.) del Golfo de Campeche, México.In: S. I.C./ Subsecretaría de Pesca, Instituto Nacional de la Pesca. 1976.58-72. Vol. I: SIGNORET, M, A An. Inst. Biol. Univ. Nal. Autón. México Serv. Zoología Abundancia, tamaño y distribución de camarones (Crustácea, Penaeidae) de la laguna de Términos Campeche y su relación con algunos factores hidrológicos.1974.119-140.(1): 45 SOTO, L. A. Proc. Gulf. and Caribb Fish. Inst.Decapod crustacean shelf fana of the Campeche Bank: Fishery aspects and ecology. 1979.66-81. 32: SOTO, L. A., A. GRACIA y A. V. BOTELLO, Proc. Gulf and Caribb. Fish. Inst. Study of the penacid shrimp population in relation to petroleum hidrocarbons in Campeche Bank.1981. 81-100. 33: SOTO, L. A., M. C. RODRIGUEZ DE LA CRUZ, R. RODRÍGUEZ, A. GRACIA, F. J. SOTO, F. ARREGÍN y E. CHAVEZ, Reunión Nacional sobre la Investigación Científico Pesquera. Cocoyoc, Morelos 26-29 mayo de 1982. Análisis del conocimiento Científico-tecnológico Nacional de las Pesquerías de Camarón en el Golfo de México.Sec. de Pesca, Instituto Nacional de la Pesca. Univ. Autón. Metropolitana 1982. WESTHREIM, S. J. y W. E. RICKER, J. Fish. Res. Board Can., Bias in using an age-length key to estimate age frequency distributions.1978.184-189. 36: WILLIAMS, A. B., Bull. Mar Sci. Gulf and Caribb. Contributions to the life histories of comercial shrimp (Penaeidae) in North Carolina.1955.116-146. 5:
|