|
ESTUDIO PROSPECTIVO DE LA MACROFAUNA BÉNTICA ASOCIADA A LAS
PRADERAS DE THALASSIA TESTUDINUM EN LA LAGUNA DE TÉRMINOS, CAMPECHE,
MÉXICO
Trabajo recibido el 29 de abril de 1985 y aceptado para su
publicación el 27 de agosto de 1985.
VIVIANE SOLIS-WEISS y SUSANA CARREÑO
Universidad Nacional Autónoma de
México. Instituto de Ciencias del Mar y Limnología.
Contribución 463 del Instituto de Ciencias del Mar y Limnología,
UNAM.
En este trabajo se analizan
los primeros resultados de la prospección de comunidades
macrobénticas asociadas a las praderas de Thalassia
testudinum en la Laguna de Términos (Campeche). La fauna
asociada a este ecosistema resultó muy variada y abundante. Los grupos
dominantes fueron los Crustáceos (46%), los Anélidos
Poliquetos(39%) y los Moluscos (14,%). La exposición a la influencia
marina de las aguas que penetran del Golfo de México parece ser el
factor más importante en la abundancia de organismos moradores de las
praderas de Thalassia.
In this study,
the first results of the prospection of macrobenthic communities, associated to
Thalassia testudinum in Laguna de Términos (Campeche)
are analysed. The fauna collected was highly diversified and abundant. The
dominant groups were the Crustaceans (46%), Polychaetes (39%) and Mollusks
(14%). The exposition of the communities to the marine influence of the current
penetrating from the Gulf of México into the lagoon seemed to be the
major factor influencing the abundance of organisms associated to the sea
grasses.
Las lagunas costeras son áreas cuyo estudio representa gran interés para el hombre por el potencial de aprovechamiento que le otorga la riqueza de su fauna y accesibilidad tanto para la explotación como para recreo. En México, la Laguna de Términos es la laguna más grande del país (aproximadamente 2000 km² de aguas abiertas), y cuenta con considerables recursos pesqueros que son la tradicional actividad de sus habitantes. La localización de la Laguna, frente a la Sonda de Campeche que constituye la zona de pesquería camaronera más productiva del país, los bancos de ostión y almeja muy florecientes dentro de la laguna, en las zonas de Atasta y Pom y las diversas pesquerías de peces y crustáceos que alberga, la convierten en un área de interés para el estudio de su ecología y otros procesos biológicos. Además, al interés intrínseco que representa su estudio en condiciones naturales, se viene a agregar el interés que despierta la zona por el desarrollo industrial y poblacional a que se ha visto sometida a raíz de la explotación petrolera en estos últimos años. Tanto la misma explotación petrolera como la concentración urbana no han tenido aún impacto nocivo sobre el medio ambiente de la Laguna (Botello, 1980). Sin embargo, se puede preveer, quizás a corto plazo que la contaminación provocada por estas actividades afecte el equilibrio ecológico de la Laguna y por ende las pesquerías de la zona por la estrecha relación que existe entre la Sonda de Campeche y la Laguna de Términos. En la Laguna de Términos se distinguen diferentes habitats bénticos que son: manglares, praderas de pastos marinos; bancos de ostión y fondos sin vegetación. Algunas zonas estan bajo influencia marina directa como son las bocas del Carmen y de Puerto Real que comunican a la Laguna con el Golfo de México y otras bajo fuerte influencia dulceacuícola, como son las desembocaduras a la laguna de los ríos Chumpan, Palizada y del complejo de las lagunas de Atasta-Pom. Las praderas de pastos marinos, en especialThalassia testudinum, son zonas de alta productividad y aparentemente áreas de crianza y protección de especies comerciales (camarones y peces). Este complejo ecosistema ha sido estudiado intensamente en los últimos años tanto por su amplía representación en los mares y estuarios tropicales del mundo (Ostenfeld, 1914; McRoy, 1973; Moore, 1963; Thoraug y Roeddler, 1971; Jackson, 1973; Croset al., 1975: Young et al., 1976; Kikuchi y Péres, 1977; Heck, 1977, 1979; Heck y Wetstone 1977; Brook, 1978; Yáñez-Arancibia, 1981; Thayer y Engel, 1982), como por su importancia ecológica ya que alberga una fauna variada y abundante (Hoese y Jones, 1963). En México, los estudios de pastos marinos han sido sobre todo de tipo florístico (Lot-Helgueras, 1968; Hornelas, 1975) e ictiológicos (Vargas Maldonado et al., 1981). Es el objeto de este trabajo estudiar las comunidades bénticas moradoras de las praderas de Thalassia testudinum, es decir emprender el estudio faunístico de este ecosistema con el fin de a) conocer la composición de especies que ahí habitan, qué grupos filéticos están representados y cuales dominan y b) poder determinar mediante este estudio prospectivo las áreas de mayor interés para futuros estudios de monitoreo de impacto ambiental. Se escogió este ecosistema como base de estudio de impacto ambiental por dos causas principales: 1) la facilidad de localización visual y 2) la poca resistencia de este a la contaminación. En efecto, las praderas de Thalassiasolo prosperan en aguas claras y limpias (DenHartog y Helferich, 1977). En este caso, es de suma importancia detectar a tiempo las señales de deterioro ambiental por la estrecha relación ecológica que tiene la laguna con la Sonda de Campeche en el aspecto de pesquerías (Yañez-Arancibia, 1982). A este respecto, los animales bénticos, por su poca movilidad, revisten especial interés puesto que sufren los efectos sinergéticos de los factores ambientales prevaleciente en su medio y reflejan las condiciones existentes en un período de tiempo previo al muestreo y no una "instantánea" como en el caso de las determinaciones químicas. ANTECEDENTESLa importancia ecológica que reviste la Laguna de Términos se refleja en el hecho que existen hasta la fecha más de 50 publicaciones sobre esta región que versan sobre diferentes grupos taxonómicos o aspectos ecológicos, físicos, químicos y geológicos. Las revisiones más relevantes y recientes sobre el área y en las cuales se podrá encontrar bibliografía completa son dos: Yáñez-Arancibia y Day (1982) que dan una caracterización ecológica de la laguna y Yáñez-Arancibia et al. (1983) que analizan su comportamiento ambiental y funcionamiento. Para los grupos faunísticos que se consideran en este trabajo los antecedentes son mucho más escasos. Para los moluscos, el grupo mejor documentado, está el trabajo de García-Cubas (1963) sobre micromoluscos y, del mismo autor, una monografía de los moluscos de la Laguna de Términos (García-Cubas, 1981) que resulta de mucha utilidad para las determinaciones taxonómicas de estos. En cuanto a los anélidos poliquetos, solo se tiene el estudio de Marron-Aguilar (1976) y algunas descripciones de Rioja (1960) sobre el material colectado ahí. Sobre crustáceos se pueden consultar los estudios de Morales (1966), Signoret (1974), Sánchez (1981). Sobre equinodermos existe la revisión de Caso (1979). Los pastos marinos de la laguna solo han sido estudiados de manera prospectiva por Zarur (1961) y Hornelas (1975). ÁREA DE ESTUDIOLa laguna de Términos tiene 60 km x 25 km en su parte más ancha y está situada en la parte Sureste del Golfo de México entre los meridiados 91°15' y 92°00' de latitud Oeste y los paralelos 18°15' y 19°00' de longitud Norte. (Fig. 1). Se localiza en el límite de una zona de sedimentación de carbonato de calcio biogénico en la parte Este y una zona de deposición de detritus terrígenos originarios de los ríos y sus tributarios en la parte Oeste (Phleger y Ayala-Castañares, 1971). La distribución de los sedimentos recientes fue analizada por Yañez Correa (1963). Las praderas de Thalassia testudinum estan ampliamente distribuidas en la parte Sur de la Isla del Carmen y en la parte Este y Sureste de la laguna (Fig. 1). Se encuentran en las zonas de mayor influencia marina dentro de la laguna ya que Thalassia testudinum solo se desarrolla en salinidades comprendidas entre 24% o y 35% o (Zieman, 1973). Llegan a cubrir hasta una quinta parte de la laguna (Zarur, 1961) pero en la parte Sureste solo se presentan de manera esporádica (observación personal). En el presente estudio se eligieron 10 estaciones de muestreo representativas de las praderas más prósperas del área. Se encuentran indicadas en la figura 1. MATERIAL Y MÉTODOSSe utilizaron diferentes métodos de muestreo con el fin de definir el mejor para este tipo de substrato. En efecto, no existe ningún método convencional que se pueda aplicar al estudio del bentos en estos ecosistemas: las dragas de arrastre quedan atrapadas en los rizomas de la fanerógama y las dragas de tipo van Veen no cierran adecuadamente por los mismos rizomas, imposibilitando el muestreo por estas técnicas. 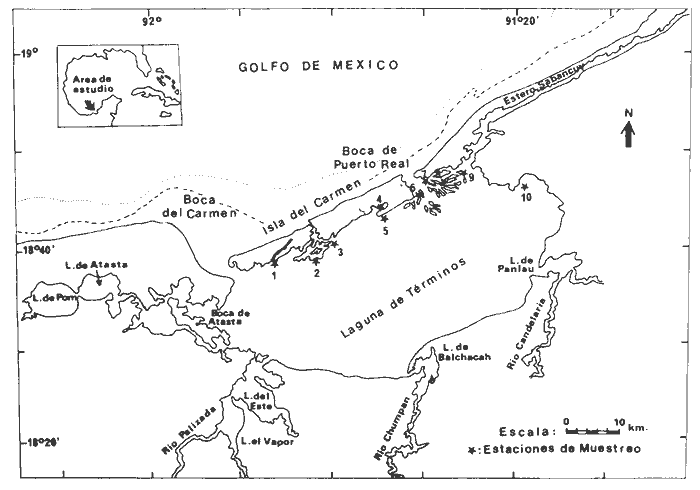 Fig. 1. Mapa de Localización del área de estudio y estaciones de muestreo. Inicialmente se utilizó un nucleador de PVC de 25 cm de diámetro y 1 m de altura según diseño utilizado en la Universidad de Texas en estos ambientes. Sin embargo, este procedimiento resulta poco práctico por necesitar de 4 ó 5 personas para el manejo del nucleador. Posteriormente se decidió utilizar un muestreo mixto que consiste en 1) recortar un área determinada por cuadrante (colectando así a la fauna sésil) y 2) efectuar un arrastre con red de tipo Colman-Sea Grove con lo cual se colecta a la fauna móvil sobre el sustrato o entre las hojas deThalassia. Esta red no penetra en el sedimento por lo cual se le llama comunmente "red de Patin". Tiene las características siguientes: 1.25 m de largo total; 0.32 m de altura; 0. 68 m de ancho; 0.57 m x 0.18 m de abertura de boca; 451 µmde abertura de malla; colector de PVC tipo plancton. Se arrastra 50 m. El cuadrante mide 0.30 m por lado. Se recolecta una área de 0.36 m² (correspondiente a 4 cuadrantes) por estación extrayendo también el sedimento hasta 20 cm de espesor. Se lava y tamiza la muestra in situ, fijándose con formol al 10%. En el laboratorio se separa a los organismos de las fracciones gruesas del substrato y de las hojas y rizomas de Thalassia y se fijan con alcohol 70º. Además, en cada estación se midió la temperatura, salinidad, profundidad, altura de la Thalassiay densidad de ésta por m². Se tomaron muestras de sedimentos que fueron analizadas granulométricamerite: en cada muestra se utilizaron las fórmulas de Folk (1965) al calcular los diferentes parámetros que caracterizan el sedimento en cada estación. 1) Mz (media) = ø 16 + ø 50 + ø 84) / 3 2) 1 (Coeficiente de sorteo) = ( ø 84 - ø 16) / 4 + (ø 95 - ø 5) / 6.6 3) SK1 (Skewness = (ø 16 + ø84 - 2 ø 50)/2 (ø 84 - ø 16) + (ø 5 + ø 95 - 2 ø 50) / 2 (ø 95 - ø 5) Los poliquetos fueron identificados con las claves de Fauvel (1923, 1927), Fauchald (1977), Hartman (1951, 1969) y Rioja (1960); los moluscos se identificaron gracias a los trabajos de García-Cubas (1981), Abbot (1974), Myra-Keen (1958) y Morris (1966) y los crustáceos con las claves de Williams (1965) y Fenner (1972). RESULTADOS Y DISCUSIÓNFueron recolectados e identificados para este trabajo 8,375 organismos pertenecientes a los tres grupos faunísticos dominantes de estas praderas deThalassia y que son: Poliqueros, Moluscos y Crustáceos. Se colectaron además esponjas, nemertinos, anélidos oligoquetos, sipuncúlidos, equinodermos y peces. Todos estos grupos fueron escasos en comparación con los tres grupos principales de manera que se anota su presencia sin considerarlos numéricamente. El caso de los peces es especial ya que abundan en estas praderas (Vargas Maldonado et al., 1981). Sin embargo, no se toman en cuenta en absoluto en este trabajo ya que el método de muestreo, no diseñado para captura de peces, sólo los recoge de manera accidental y por lo tanto no representativa. Se encuentran en el mismo caso los crustáceos grandes (como Callinectes sapidus). En la Tabla 1, se encuentran los resultados de los parámetros físico-químicos medidos. Las variaciones de temperatura y salinidad medidas en los diferentes meses de muestreo muestran congruencia con los resultados obtenidos en esta área por Botello (1978). Los resultados del análisis granulométrico se encuentran expresados en las figuras 2 a 10, por estación y los valores para cada estación de la media (Mz), el coeficiente de sorteo (1), Skewness (SK1,) y Curtosis (Kg) están expresados en la Tabla 2. De los tres grupos que se analizan, 3,328 organismos pertenecen a la Clase Polychaeta representada por 22 familias, 36 géneros y 41 especies; 1, 186 al Phylum Mollusca representado por 12 familias y 15 especies en la Clase Gasterópoda y 8 familias y 10 especies en la clase Bivalvia; 3,860 organismos pertanecen a la Clase Crustácea representada por 3 ordenes, 9 familias y 12 especies. 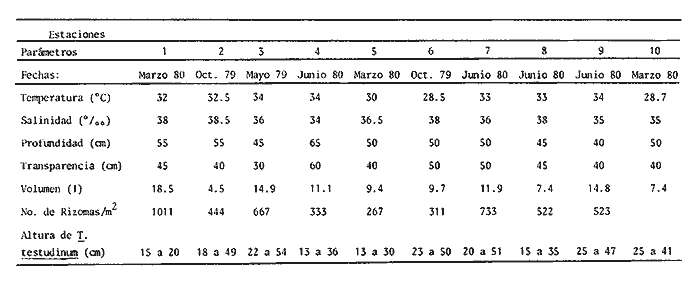 TABLA 1. PARÁMETROS FÍSICO-QUÍMICOS MEDIDOS EN LAS ESTACIONES DE MUESTREO Todas las especies colectadas están anotadas filogenéticamente en las Tablas 3, 4 y 5, indicándose también su abundancia y dominancia relativa en cada estación. CARACTERÍSTICAS FÍSICO-QUÍMICAS Y TEXTURALES, DEL ÁREA DE ESTUDIO.En este estudio, se consideran las plantas deThalassia como componentes estructurales del habitat. En efecto, la Thalassia no constituye una fuente de alimento para la mayoría de los organismos, excepto para la tortugaChelonia midas y el erizo Lutechinus variegatus (Kikuchi y Péres, 1977) especies que no se han colectado en esta área. Vargas Maldonado et al.(1981) reportan algunas especies de peces consumidores de 1º 2º orden, es decir que consumen vegetales en la totalidad o la mayoría de su dieta usual. Sin embargo, no se precisa ahí si se trata de la especieThalassia testudinum que es consumida directamente o si el pez ingiere la fanerógama como parte de las numerosas epífitas y epizoos de esta que sí consume y aprovecha. En este contexto, la función de la Thalassia es importante como substrato adicional para la implantación de especies animales sésiles y vegetales que sí participan directamente en las cadenas alimenticias del ecosistema (algas diversas, hidroideos o micromoluscos). La Thalassiaentra a la cadena alimenticia y se integra al flujo de energía en el ecosistema generalmente por la vía detrítica a través de la descomposición por microorganismos de las hojas muertas. Una discusión sobre estos procesos se puede encontrar en Yáñez-Arancibia (1981). 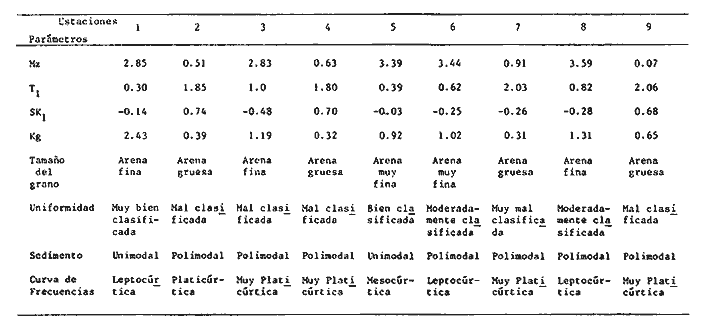 TABLA 2. COEFICIENTES DE TEXTURA SEDIMENTOLÓGICA 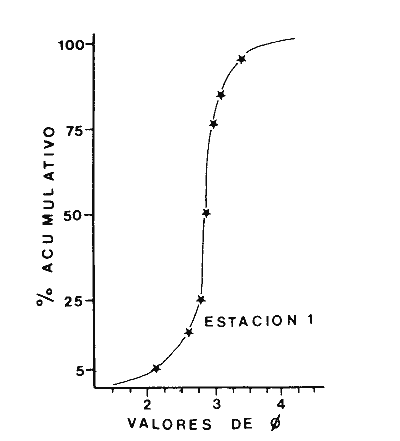 Fig. 2. Curva granulomética. Estación 1. 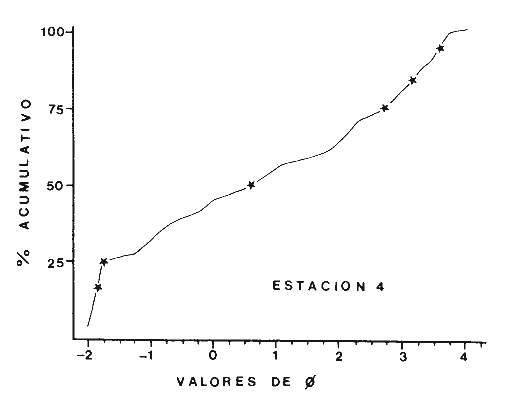 Fig. 3. Curva granulomética. Estación 2. 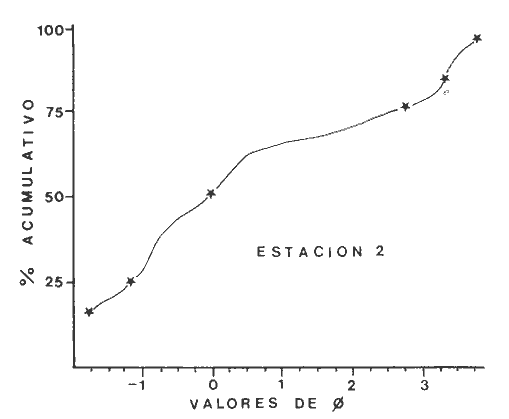 Fig. 4. Curva granulomética. Estación 3. 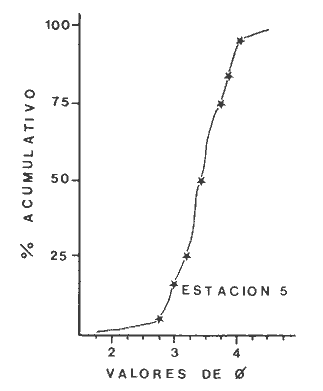 Fig. 5. Curva granulomética. Estación 4. 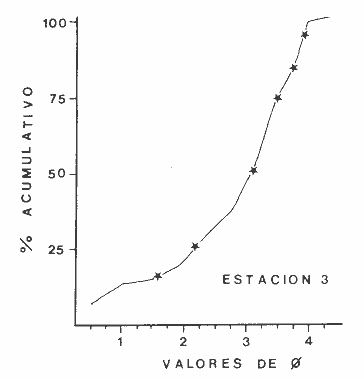 Fig. 6. Curva granulomética. Estación 5. 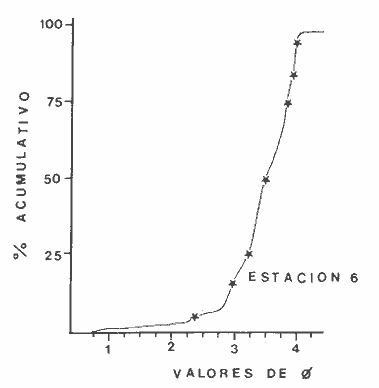 Fig. 7. Curva granulomética. Estación 6. 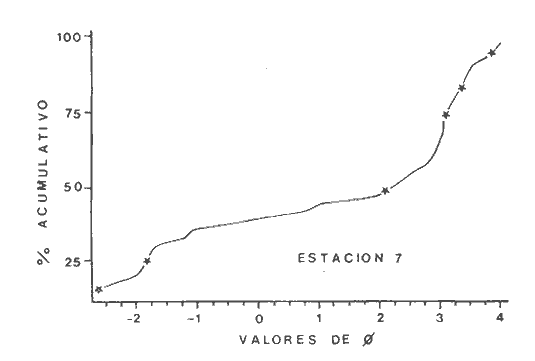 Fig. 8. Curva granulomética. Estación 7. 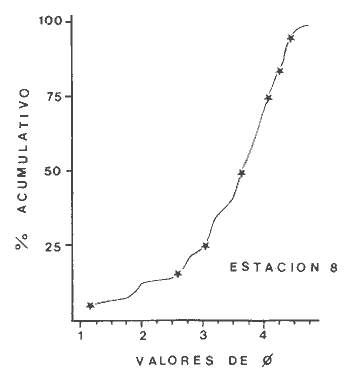 Fig. 9. Curva granulomética. Estación 8. 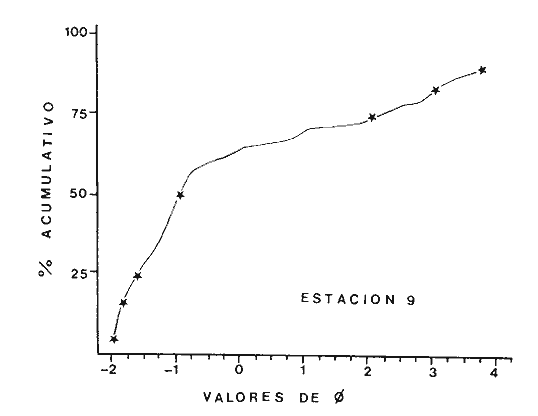 Fig. 10. Curva granulomética. Estación 9. Otra característica importante de esta planta para el presente estudio es que atenúa la corriente favoreciendo activamente la sedimentación y captación de materia orgánica (Ginsburg y Lownstan, 1976). Además, la densidad y altura de las hojas deThalassia proporcionan protección a las especies moradoras del ecosistema tanto de sus depredadores como de excesiva insolación. Tabla 5 Las diez estaciones muestran amplia variación en sus características en cuanto a la fisionomía de la Thalassia y al tipo de sedimento en que la crece (Figs. 2 a 10). Se observó que la altura de las hojas varía de 13-15 cm (estaciones 1 y 4) hasta 51-54 cm (estaciones 3 y 7) (Tabla 1). La densidad del follaje expresada como número de tallos por m² también muestra amplia variación ya que se encontraron desde 267 tallos deThalassia por m² (estación 5) hasta 1011 (estación 1). Estos valores representan el promedio de varias mediciones. La altura de las plantas varía con la época del año pero, independientemente de ésta, si una mayor cobertura de follaje por área (ya sea debida a la altura o a la densidad) confiere ciertas ventajas a la fauna, aunque sea de manera estacional, esto se reflejará en los resultados faunísticos que se analizarán más adelante. En la Tabla 2, se reportan los resultados del análisis granulométrico que se encuentran expresados gráficamente en las figuras 2 a 10. Se puede observar que en las estaciones 1 y 5 se presentan sedimentos similares de arena fina bien clasificada y sedimentos unimodal. Además de observó en estas dos estaciones gran compactación del sedimento. La estación 6 con características gráficas similares a las dos precedentes, presenta sin embargo un sedimento poco compactado y moderadamente clasificado. Las estaciones 2 y 4 muy parecidas entre sí, al contrario de las estaciones 1 y 5, presentan un sedimento de arena gruesa mal clasificada y polimodal cuya curva de frecuencias es platicúrtica. El tamaño del grano, así como la variación de tamaños del grano, expresada en el grado de clasificación y la compactación del sedimento son factores muy importantes para la fauna béntica que se llega a caracterizar inclusive por el tipo de sedimentos en que habita preferencialmente (Solís Weiss, 1982). La compactación del sedimento tiene efectos de tipo etológico ya que un sedimento muy compactado favorecerá a la epifauna y un sedimento poco compacto al contrario favorecerá a la infauna. Se observó además que en las estaciones 1 y 5 los rizomas forman una intrincada red debajo de la superficie del substrato, que dificulta el muestreo. En las estaciones 2, 4 y 6 al contrario, los rizomas son escasos y menos gruesos. En las demás estaciones se encuentran situaciones intermedias. Se mencionó anteriormente que la Thalassiafavorece la activa sedimentación de todo tipo de partículas, que pueden ser tanto orgánicas como inorgánicas. Al tamizar las muestras, se encontró en varias estaciones, gran abundancia de fragmentos de conchas o conchas enteras de moluscos incorporadas al sedimento. Estas constituyen microsubstratos duros aptos para ser utilizados por organismos característicos de estos substratos (como Sabélidos) que de otra manera no se podrían fijar en el substrato relativamente blando de arena y lodo presentes en todas las estaciones. En la Tabla 1 se puede observar que las estaciones que presentaron mayor cantidad de fragmentos de este tipo fueron las estaciones 2, 4, 5 y 7 y las que casi no tenían bioclastos incorporados al sedimento fueron las estaciones 3, 8, 9 y 10. Estas son apreciaciones cualitativas.
TABLA 3
TABLA 4 ANÁLISIS DE LA FAUNA COLECTADALa fauna colectada en estas praderas de Thalassia testudinum fue muy variada y abundante como se indicó más arriba, independientemente de la época del año. Sin embargo, las proporciones de individuos, es decir las dominancias relativas y, por ende, la composición faunística presentan amplio rango de variación entre las diferentes estaciones. En la Tabla 6, se pueden ver las dominancias relativas registradas para los tres grupos dominantes en las diez estaciones de muestreo. Esta Tabla, al tomar en cuenta únicamente los porcentajes representados por los Anélidos Poliquetos, Moluscos y Crustáceos da la proporción de cada estación permitiendo comparaciones directas. 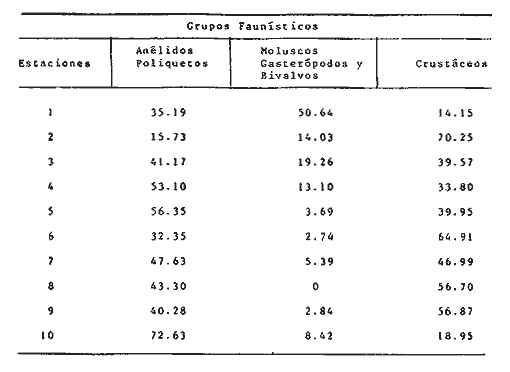 TABLA 6. DOMINANCIAS RELATIVAS (EXPRESADAS EN PORCENTAJE) DE POLIQUETOS MOLUSCOS Y CRUSTÁCEOS EN LAS ESTACIONES DE MUESTREO. Se observa en particular que los moluscos son el grupo proporcionalmente menos representado ya que solo dominan en la estación 1. Los Poliquetos dominan en cinco estaciones (3, 4, 5, 7 y 10) y los Crustáceos en seis estaciones (2, 3, 6,7, 8 y 9). Los Crustáceos además dominaron en números totales de organismos colectados (3861 individuos por 3328 de Poliquetos y 1186 de Moluscos). La diversidad de los organisos, es decir el número de especies encontradas por estación fue mayor en las estaciones 6 y 7 y menor en la estación 10 como se puede observar en las Tablas 3, 4 y 5: Se encontraron 21 especies de Poliquetos en la estación 7 y 19 en la estación 6 en comparación con 7 en las estaciones 10 y 2. Para el phylum Mollusca se recolectaron 8 especies en las estaciones 6 y 7, 0 en la estación 8 y 3 en las estaciones 9 y 10. En cuanto a los Crustáceos se colectaron 11 especies diferentes en la estación 6, 10 en la estación 7 y 4 solamente en la estación 10. A pesar de que la diversidad fue mayor en las estaciones 6 y 7, en números totales de organismos colectados estas dos estaciones difirieron ampliamente. En efecto, como se puede observar en la figura 11, las estaciones en que se colectó el mayor número de organismos fueron las estaciones 5 y 6, en cambio la estación 7 presenta pocos organismos en comparación. En efecto, en la estación 5 se colectó el 20.7% del total de individuos de las 10 estaciones y en la estación 6 el 22.6% del total, es decir que entre las dos estaciones se encuentra casi el 50% del total de la colecta de este estudio, correspondientes a 1732 y 1898 individuos respectivamente. En cambio, la estación 7 con 464 individuos en total, a pesar de poseer la más alta diversidad (39 especies) solo representa 5.5% de la colecta total. La estación 6 con el mayor número de individuos también posee la más alta diversidad después de la estación 7 ya que se colectaron ahí 38 especies diferentes pertenecientes a los tres grupos dominantes. Al comparar las características físicas y texturales de las estaciones 5 y 6 en que se encontró mayor número de organismos, se ve que son totalmente diferentes. Desde el punto de vista textural, la estación 5 posee un sedimento compuesto de arena muy fina, bien clasificada unimodal y bastante compacta. (Tabla 2). La estación 6 posee arena muy fina, moderadamente clasificada, polimodal y poco compacta. La densidad de Thalassia es la más baja de las diez estaciones en estas dos. En cambio en su altura las plantas sí presentan gran diferencia (13 a 30 m en la estación 5; 23 a 50 cm en la estación 6). Las diferencias básicas entre estas dos estaciones consisten en la compactación y la altura de laThalassia. Las estaciones en que se colectó el menor número de organismos en total fueron las estaciones 4 y 8 (145 y 194 individuos de los tres grupos respectivamente). Consultando las Tablas 1 y 2, se ve que estas estaciones difieren texturalmente (arena gruesa mal clasificada polimodal, muy poco compacta en la estación 4; arena fina moderadamente clasificada, polimodal y poco compacta en la estación 8). 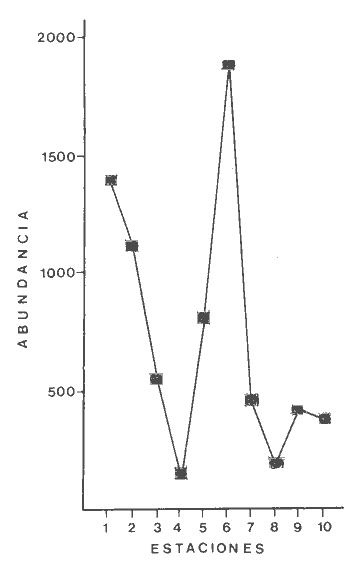 Fig. 11. Abundancia de los poliquetos, moluscos y crustáceos en las estaciones de Muestreo. Al comparar las estaciones 4 y 8 entre sí y con las estaciones 5 y 6 se ve que no destacan diferencias particulares a las cuales se pueda atribuir la gran disproporción en la cantidad de organismos colectados que son diez veces más abundantes en las estaciones 5 y 6 que en las estaciones 4 y 5. Al observar la localización de las estaciones (Fig. 1) se ve que la corriente marina preponderante que penetra del Golfo de México a la laguna pasa por las estaciones 6 y después 5. La estación 4 se encuentra fuera de esta corriente principal puesto que está en un estero y la estación 8, ubicada en la parte posterior de los bajos de la Bayoneta también se encuentra bajo la influencia más moderada de la corriente marina (Graham et al., 1981). Así, al no poderse relacionar ningun factor textural ni de densidad o altura de la Thalassia en las diferentes estaciones que explique la abundancia o escasez de la fauna asociada, se destaca el hecho que la abundancia de organismos parece estar en relación directa con su exposición a la corriente marina principal que penetra del Golfo. El caso de la estación 1 que presenta gran cantidad de organismos no constituye una excepción puesto que dicha abundancia se deriva únicamente de la gran cantidad de Neritina virgínea que constituye una característica local de esa estación y que se analiza más adelante. DISTRIBUCIÓN PARTICULAR DE LOS GRUPOS FAUNÍSTICOS
MoluscosComo se puede observar en estos resultados, el grupo de los moluscos fue el más pobremente representado en las praderas de Thalassia. Constituyen menos del 15% del total de la colecta y esto se debe únicamente a la gran abundancia del gasterópodo Neritina virgíneaque con 648 individuos colectados (únicamente en la estación 1) constituye 54.45% del total de los moluscos, de no ser por esta especie, los moluscos hubieran representado únicamente el 7% de la colecta total. El caso de N. virgínearepresenta un problema interesante ya que se colectó únicamente en la desembocadura de Estero Pargo, (localización de la estación 1), y ahí en grandes cantidades. En las otras estaciones de muestreo se colectaron algunos ejemplares de N. virgínea pero muertos, es decir que solo se recolectó la concha, en buen estado pero vacía. Este molusco es, según Andrews (1977) y García-Cubas (1981) una especie estuarina, eurihalina, comunmente encontrada en praderas de pastos marinos; es epifáunico, micrófago y se alimenta raspando la superficie de dichos pastos. No es posible, de momento, concluir sobre la causa probable de la abundancia de estos moluscos en esta localidad. Se necesitan estudios posteriores para esclarecer este problema. En la mayoría de las estaciones, las poblaciones de moluscos no alcanzan el 10% de la población de la estación, excepto en las estaciones 2, 3 y 4 en que alcanzan 14%, 19.6% y 13% respectivamente. El caso de la estación 1, es excepcional por la presencia deN. virgínea que se menciona en el párrafo anterior, ya que esta especie sola ocasiona que los moluscos alcancen 50.6 % de la población total. Además de N. virgínea, solo fueron colectadas otras 3 especies de Moluscos en muy pocas cantidades (Tabla 4). Las otras especies de moluscos colectados son, prácticamente todas eurihalinas y se alimentan de partículas en suspensión (Andrews, 1977 y García-Cubas, 1981). Esto es importante ya que los pastos marinos, al frenar la corriente favorecen este tipo de alimentación. PolíquetosLos poliquetos representan casi 40% de la colecta y son el grupo que representa mayor diversificación (comparar Tablas 3, 4 y 5). Esto no es de sorprender ya que los poliquetos representan generalmente el grupo más abundante y diversificado en la mayoría de los ambientes marinos bénticos, en especial de substrato blando. (Fauchald y Jumars, 1979). El poliqueto más abundantemente colectado fueSabella microphtalma,- se colectaron 942 individuos referibles a la especie (casi 30 % de la población; Tabla 3) y repartidos en las 10 estaciones de muestreo. Le sigue en abundancia la especie Capitella capitata que son 662 individuos forma prácticamente el 20% de la población de poliquetos en las 10 estaciones. Este poliqueto es interesante de mencionar ya que ha sido ampliamente estudiado por su comportamiento altamente oportunista (Reish, 1959; Grassle y Grassle, 1974) y por ser generalmente reconocido como indicador de condiciones de contaminación orgánica (Pearson y Rosenberg, 1978). En este caso fue colectado en 8 de las 10 estaciones siendo especialmente abundante en la estación 6 en la que constituye casi 1/ 4 parte de la población total de la estación (Tabla 4). Hay que hacer notar que el tamaño de los individuos colectados es muy pequeño (0.5 cm o menos). Otro poliqueto abundantemente colectado fue Neanthes succinea(330 individuos en total). La especie fue colectada en las 10 estaciones siendo más abundante en las estaciones 1 y 5 (Tabla 4). Este nereido es cosmopolita especialmente en bahías y estuarios ya que soporta condiciones de poca salinidad y considerables aportes de agua dulce a su medio (Hatman, 1951). CrustáceosLos crustáceos constituyeron el grupo dominante del muestreo puesto que se colectaron 3861 organismos correspondientes al 46.1 % del total de la fauna en las 10 estaciones. La gran cantidad de crustáceos colectados se debe a: 1) los anfípodos y 2) Hippolyte zostericola. (Tabla 5). 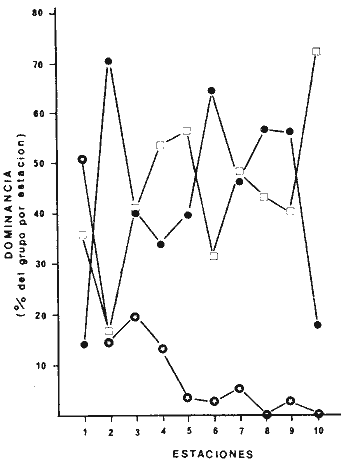 Fig. 12. Dominancia (%) de poliquetos, moluscos y crustáceos en las estaciones de muestreo. Anfipodos. Puesto que no existen claves satisfactorias para los anfipodos de nuestra región, se determinaron a nivel de orden a estos individuos por lo que no se pueden hacer comentarios muy extensos fuera de notar su enorme abundancia en especial en las estaciones 2 y 6. Hippolyyte zosterícola. Es un carideo típico de las praderas de pastos marinos como su nombre específico lo indica. La especie es eurihalina y su color verde similar al de las hojas de Thalassia la ayuda a pasar desapercibida en ella. Fue más abundante colectado en las estaciones 6 y 5 (500 y 464 individuos respectivamente). Tanto los anfípodos como H. zosterícola son especies nadadoras que se encuentran comúnmente entre las hojas de la Thalassia. Los peneidos Penaeus setiferus y Penaus duorarum se colectaron en estadíos de post-larva, juveniles y adultos. Estos son camarones de importancia comercial y se encontraron solo en las estaciones en que la influencia marina es más marcada (estaciones 5, 6, 7, 8 y 9), especialmente en la estación 6, la más "marina" de todas. Su número decrece proporcionalmente al alejarse, en el sentido de la corriente, de la Boca de Puerto Real (Tabla 5). La abundancia relativa de estas especies en las praderas confirma que son sitios preferenciales para criadero de camarones ya sea por la fuente de alimento que proporcionan a estos o por la protección contra sus depredadores. El pagúrido Pagurus annulipes fue especialmente abundante en la estación 2 (198 individuos), encontrándose solo de manera esporádica en las demás estaciones. La presencia de los anomuros depende directamente de la presencia previa en el medio de conchas de gasterópodos en las que se pueden introducir. En este caso se encontraba en conchas de Cerithiun eburneum y Modulus modulusque también fueron colectados vivos en esta estación. XifosurosLos xifosuros de la especieLimulus polyphemus no fueron cuantificados en las Tablas ya que los métodos de muestreo utilizados no permitían colectarlos de manera cuantitativa. Sin embargo es de mencionar su gran abundancia en la estación 5 y esporádicamente en la estación 7. El área de Punta Gorda (estación 5), es conocida por los pescadores como área de reproducción de las "cazuelas de mar" como se les llama vulgarmente. EquinodermosEs de notar la escasez de esto organismos en las praderas de pastos marinos de la Laguna de Términos. En efecto, en este caso solo se colectaron dos holoturoideos de la familia Cucumaridae y 3 ofiuros de la especie Amphioplus abditus,siendo que se reporta que usualmente en las praderas de Thalassia testudinum, el phylum está bien representado (Kikuchi y Pérez, 1977). Incluso, el equinoideo Lytechinus variegatus es conocido como uno de los pocos organismos que consume directamente Thalassia. Caso (1979) reporta para la laguna de Términos varios géneros colectados enThalassia en el área correspondiente a las estaciones 6, 5 y 3. La escasa representación del phylum es sorprendente. Es posible que esto se deba a los cambios rápidos de la salinidad, en especial cuando la salinidad es muy baja como de hecho ocurre en la laguna en las diferentes épocas del año (Yáñez- Arancibia, 1983). EsponjasSe encuentran en abundancia colonias de esponjas adheridas a los tallos de Thalassia únicamente en la estación 1. Es de notar la asociación de pequeños anélidos poliquetos de la familia Syllidae con estas esponjas. SipuncúlidosEstos pequeños animales fueron esporádicamente colectados en las estaciones 2, 4 y 6. Su número es reducido por lo que solo reporta su presencia. Nemertinos y oligoquetosFueron comunes en todas las estaciones de muestreo. Los nemertinos estaban invariablemente fracmentados. Los oligoquetos están representados por dos especies al menos, cuyo estudio y determinación están en proceso. PecesSon muy abundantes y variados pero no se tomaron en cuenta aquí porque el método de muestreo no permite representatividad de estos. Estudios detallados sobre este grupo en Thalassia testudinum del área se pueden encontrar en Vargas et al. (1981). ConclusionesLos resultados de este estudio prospectivo de la macrofauna béntica asociada a las praderas de Thalassia testudinum en la Laguna de Términos permiten poner en evidencia la gran diversidad y abudancia de esta fauna en diversas épocas del año. De los tres grupos dominantes en este medio: poliquetos, moluscos y crustáceos, este último fue el que resultó más abundante. Dentro de los crustáceos, las especies mejor representadas: afípodos eHippolyte zostericola, son nadadores entre las hojas deThalassia y no sésiles del bentos. Los poliquetos se encuentran asimismo bien representados y con mucho mayor diversidad que los otros dos grupos. Los moluscos resultaron ser el grupo faunístico con menor representatividad. Se destaca la enorme abundancia local (estación 1) del gasterópodoNeritina virginea. Por lo que aparece en los resultados, las estaciones con mayor influencia marina son las que gozan de mayor abundancia de organismos aunque no forzozamente de mayor diversidad. Es ahí donde se encontró también el mayor número de especies de interés comercial (Penaeus dourarumy P. setiferus). A este respecto se confirma la idea de las praderas de pastos marinos como áreas de crianza para este tipo de especies. La altura o densidad de la Thalassia en las zonas de muesteo parece de importancia secundaria en la distribución faunística. Sin embargo, son necesarios los resultados de estudios posteriores, estacionales, para confirmar cuales son los parámetros más importantes para dicha distribución. AgradecimientosLos autores desean expresar su agradecimiento a Rodolfo Cruz por todas las facilidades prestadas para el desarrollo de este estudio durante su periodo como jefe de la Estación "El Carmen" y por los análisis granulométricos efectuados en las estaciones de muestreo. Se agradece asimismo a los miembros del Laboratorio de Malacología en especial a Antonio García-Cubas y Felipe Flores por la ayuda prestada en la identificación de moluscos. Igualmente se agradece a los miembros del Laboratorio de Ecología de Bentos en especial a Luis Soto por la ayuda en la identificación de crustáceos a Alberto Sánchez por proporcionar algunos datos de salinidad y temperatura del área de estudio a Arturo Yáñez y Rocio Arenas por su ayuda en el campo. También agradecemos a Alejandro Yáñez-Arancibia por su ayuda y consejos en la revisión del trabajo y Alfredo Laguarda por el apoyo brindado durante su periodo como Director del Centro de Ciencias del Mar y Limnología. A José Andrés Reda por su ayuda en el trabajo de campo. LITERATURAABBOT, T.R., American Sea Shells. Van Nastrad Reinhold Co. 2a. Ed.New York, 1974.663p. ANDREWS, J., Shells and shores of Texas. Texas University Press. Austin y London1979. 365 p. ARENAS, R. y A. YÁÑEZ MARTÍNEZ, Patrón anual de inmigración de postlarvas de camarón en la boca de Puerto Real, Laguna de Términos, Campeche. Tesis Prof.Facultad de Ciencias. Univ. Nal. Autón. México,1981. 92 p. BOTELLO, A. V., An. Centro de Cienc. del Mar y Limnol. Variación de los parámetros hidrológicos en la Laguna de Términos, Campeche, Mexico. Univ. Nal. Autón. México1978.159-178.(1): 5 BOTELLO,A. V., An. Centro Cienc. del Mar y Limnol. Cuantificación de un derrame petrolero ocurrido en la Laguna de Términos, Campeche. Univ. Nal. Autón. México, 1980.169-176.(1): 7 BROOK, I., Bull. Mar. Sci., Comparative macrofaunal abundance in turtlegrass (Thalassia testudium) communities in South Florida Characterized by high blade density.1978.211-217. (1): 28 CASO, M.E., Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Publ. Esp. Los equinodermos (Asteroldes Ophuiroidea y Echinoidea) de la Laguna de Términos, Campeche. 1979. 1-186. 3: CROS, L.D., B. DEL ROSONO, G. CHONG y G. YEE, Simposio Latinoamericano sobre Oceanografía Biológica,Ecología de las praderas de Thalassia en la costa atlántica de Panamá. Venezuela.1975.35-49. 2: DEN HARTOG, C. Structure functión and classification in seagrasses communities Mc Roy, P. and C. Helfferich (Eds.) Inc. New York y Basel: 1977.89-121. FAUCHALD, K., The Polychaete Worms. Definitions and Keys to the orders Familles and Genera. Nat. Hist. Mus. of Los Angeles,1977. 188 p. FAUCHALD, K. y P. JUMARS, Oceanogr. Mar. Biol. Ann. Rev. The diet of worms: A study of Polychaete feeding guilds.1979.198-289. 17: FAUVEL, P. Polychetres Errantes Faune de France (5). le Chevalier (Ed.) Paris. 1923. 488 p. FAUVEL, P., Polycheles Sédentaires. Faune de France (16) Le Chevalier (Ed). Paris.1927. 494 p. FENNER, A. C. The shrimps of the Smithsonian Bredin Caribbean expeditions Smithsonian Inst. Press. Washington D. C.1972. No. 98. FOLK, R.L., Petrology of Sedimentary Rocks Hemphill's Ed. Texas.1965 159 p. GARCÍA-CUBAS, A.J., Bol. Inst. Geol. Univ. Nal. Autón. México,Sistemática y distribución de los micromoluscos de la Laguna de Términos, Campeche, Mexico.1963.671-55. GARCÍA-CUBAS. A., Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Publ. Esp. Moluscos de un sistema lagunar tropical en el sur del Golfo de México (Laguna de Términos, Campeche). 1981.1-182. 5: GINSBURG, R. M. y H. A. LOWNSTAN, Mem. Cong. Int. Geol. Mex., Influence of Marine bottom communities on the depositional environmente of sediments. 1958. 232 p. GRAHAM, D. S., J. P. DANIELS, J. M. HILL y J. W. DAY Jr. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, A preliminary model of the circulation of Términos lagoon, Campeche, Mexico.1981.51-62.(1): 8 GASSLE, J. F. y J. P. GASSLE, J. Mar. Res.Opportunistic life histories and genetic system in marine benthic polychaetes 1974,253-284. 32: HARTMAN, O., Publ. Inst. Mar. Sci. The littoral marine annelids of the Gulf on Mexico.Univ. Tex.1951,7-124. 2: HARTMAN, O. Atlas of the sedentary Polychaetes Annelids from California. 1969.827 p. HECK, K. L, Comparative species richness compositions and abundance of invertebrates in Caribbean seagras (Thalassia testudinum). Dept. of Biol. Sci. Fla. State Univ. Mar Biol., 1977.335-348. 41: HECK, K. L., J. of Biogreography, Some determinants of the composition and abundance of motile macroinvertebrate species in tropical and temperte turtlegrass (Thalassia testudiunum) meadows. 1979.183-200. 6: HECK, K. L. y G. S. WETSTONE, J. of biogeography,Habitat complexity and invertebrate species richness and abundance in tropical seagrass meadows 1977.135-142. 4: HOESE, H.D. y R.D. JONES, Publ. Inst. Mar. Sci.Seasonality of larger animals in a Texas turtle grass community. Univ. Texas: 1963.33-47. HORNELAS, Y., Comparación de la biomasa y de algunos aspectos morfometricos de la fanerógama Thalassia testudinum, Koening 1805 en tres diferentes áreas del Golfo de México. Tesis Profesional Fac. Ciencias, Univ. Autón. Méxíco1975. 80 p. JACKSON, J., Bull. Mar. Sci. The ecology of mollusks of Thalassia commuinities, Jamaica, West Indies. I. Distribution environmental physiology and ecology of shallow water species.1973.313-350.(2): 23 KIKUCHI, T. y J. M. PÉRÈS Seagrass Ecosystems Consumer ecoloty of Seagrass beds, In: McRoy, P. and C. HeIfferich (Eds.) M. Dekker Inc. New York.1977.89-121. McRoy, P., Seagrass Ecosystems. Proceedings of the International Seagrass Workshop Leiden, the Netherlands.1973. MARRÓN-AGUILAR, M. A., Estudio cuantitativo y sistemático de los poliquetos (Annelida: Polchaeta) bentónicos de la laguna de Términos, Campeche, Mexico. Tesis Doctoral, Fac. Ciencias. Univ. Nal. Autón. Mexico. 1976. 142 p. MOORE, D. R., Bull. Mar. Sci. Gulf and Carib.,Distribution of the Sea Grass Thalassia in the United State. 1963.329-362. 13: MORRIS, A. P., A Field Guide to Pacific Coast ShellsHoughton Mifilin Co.2nd. Ed.Boston 1966. 297 p. MYRA KEEN, A., Sea Shells of Tropical West America. Stanford Univ. Press Stanford, California.1958. 694 p. OSTENFEELD, C. H., Proc. Soc. Roy. Victoria, On the geographical distribution of the seagrasses.1914.179-190. 27: PEARSON, T. H. y R. ROSENBERG, Oceanogr. and Mar. Biology. An Annual review, Macrobenthic succession in relation to organic enrichment and pollution of the marine environmente. 1978. 229-311. 16: RESIH, D. J., Bull. So. Calif. Acad. Sci., An ecological study of the Polychaetous Annelids associated with fouling material in Los Angeles Harbor with special reference to pollution.1959.170-181.(3) 68 RIOJA, E., An. Inst. Biol. Méx. Estudios Anelidológicos XXIV. Adiciones a la fauna de Anélidos Poliquetos de las costas orientales de México.1960.289-316. 31: SÁNCHEZ, M. A. Comportamiento anual de las postlarvas espibénticas de camarones peneidos en el sector oriental de la Laguna de Términos, Campeche. Tesis Prof. Fac. de Ciencias, Univ. Nal. Autón. Mexico.1981. 97. p. SIGNORET, M., An Inst. Biol. Univ. Nal. Autón. México Ser. Zoología.Abundancia, tamaño y distribución de camarones (Crustacea-Peneidae) de la Laguna de Términos, Campeche, México y su relación con algunos parámetros hidrológicos.1976.119-114 (1): 45 SOLIS-WEISS, V., An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Aspectos ecológicos de la contaminación orgánica sobre el macrobentos de las cuencas de sedimentación en la había de Marsella (Francia). 1982. 19-44.(1): 9 THAYER, G. W. y D. W. ENGEL, J. Exp. Mar. Biol. Ecol. Evidence for short circuiting of the detritus cycle of seagrass beds by the green turtle Chelonía mydas,1982.173-183. 62: THORAUG, A. y M. A. ROESSLER, AquacultureSeagrass community dynamics in a subtropical estaurine lagoon. 1971.253-277. 12: VARGAS MALDONADO, I., A. YÁÑEZ-ARANCIBIA, A. y F. AMMEZCUA LINARES, An. Inst. de Cienc. del Mar y Limnol. Univ. Na. Autón. México. Ecología y estructura de las comunidades de peces en áreas de Rhizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos, sur del Golfo de México. 1981. 241-266.(1): 8 WILLIAMS, A. B., An. Fish. Bull.Marine decapod crustaceans of the Carolinas.1965.1-298. (1): 65 YÁÑEZ-ARANCIBIA, A., UNESCO technical papers in marine scienceEcology in the inlet of Puerto Real, Términos lagoon, México. 1981.191-220. 33: YÁÑEZ-ARANCIBIA, A. yJ. W. DAY Jr., Costal Lagoon. Oceanologica Acta. Ecological. Characterization of Términos lagoon a tropical lagoon estuarine system in the southern Gulf of Mexico. p. 431-440. Vol. Spec., In: Laserre P. y H. Postma (Eds.) 1982.462 p. (4): 5 YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. CHAVANCE y D. FLORES HERNÁNDEZ, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México,Evironmental behavior of Términos Lagoon ecological system Campeche, México. 1983.137-176.(1): 10 YÁÑEZ-CORREA, A., Bol. Inst. Geol. Univ. Nal. Autón. México, Batimetría, salinidad, temperatura y distribución de los sedimentos recientes en la laguna de Términos, Champeche, México.1963.1-47.(1): 67 YOUNG, D.K. M. BUZAS, y M. YOUNG. J. Mar. Res.Species densities of macrobenthos associated with seagrass: A field experimental study on prediation.1976.577-592.(4): 34 ZARUR, M.A., Estudio biológico preliminar de la Laguna de Términos, Campeche, México. Tesis Prof. Fac. Cienc. Univ. Nal. Autón. México1961. 64 p. ZIEMAN, J.C., Estuarine Research, Chemistry, Biology and the estuarine System. Quantitative and dynamic aspects of the ecology of turtle grass Thalassia testudinum. In: Cronin, L.E. (Ed.). Academic Press Inc. N. York:1975.541-563.
|