|
FAUNE MOBILE DES HERBIERS DE PHANÉROGAMES MARINES (HALODULE ET
THALASSIA) DE LA LAGUNA DE TÉRMINOS (MEXIQUE, CAMPECHE). II LES
GAMMARIENS (CRUSTACEA)
Trabajo recibido el 23 de abril de 1985 y aceptado
para su publicación el 9 de septiembre de 1985.
MICHEL LEDOYER
Station marine
d'Endoume, rue de la Batterie des Lions. 13007 Marseille.
En los pastos
de fanerógamas marinas de la Laguna de Términos, cuantitativamente
los anfipodos, constituyen generalmente el segundo grupo animal después
de los Caridea de la población móbil. Diecisiete especies
se encuentran en estas aguas sobre las cuales dos;Cymadus compta
y Gitanopsis laguna estan presentes durante la estación
estudiada en todas las muestras. Una especie Nasageneia yucatanensis
n. sp. parece vivir únicamente en la parte de la laguna más
sometida à la influencia marina. Poblaciones de anfipodos, parecen
ser también más importantes durante la noche entre el
follaje y muy parecidas de las que se encuentran en los pastos de
Zostera marina o deThalassia de las costas atlánticas
de los Estados Unidos. Les gammariens,
quantitativement et de façon générale, représentent
dans le secteur étudié et durant la période hivernale
le second groupe faunistique. 17 espèces ont été
recensées dans les deux types d'herbiers (Halodule etThalassia)
parmi lesquelles 2 (Cymadusa compta et Gitanopsís laguna)
ont une répartition quasiment constante. Une espèce, considérée
comme nouvelle, Nasageneia yucatanensis n. sp. paraît cantonnée
dans la partie la plus marine du secteur étudié. Chez
les amphipodes gloablement on observe à peu près le même
type de répartition que chez les Caridea. Un groupe limité
à 3 ou 4 espèces yit dans presque tous les biotope de
phanérogames prospectées. Un autre apparaît préférentielle
des herbiers d'Halodule des zones les plus dessalées.
Quelques espèces se limitent aux herbiers de Thalassia
ou aux zones les plus soumises aux influences marines. De façon
générale cette faune paraît très semblable
à celle qui est signalée des herbiers des côtes
Nord Est des U.S.A. Quant aux variations nycthémèrales
elles semblent très importantes, tout au moins en ce qui concerne
les abondances spécifiques.
Ce travail constitue
la suite de l'étude entreprise sur les Caridea (Crustacea, Decapoda)
capturés dans la même série de récoltes (Ledoyer,
1986). Ces récoltes furent réalisées en janvier 1983 grace
aux accords de coopération franco-mexicains avec financement conjoint du
CONACYT (Mexique)
Projet: "Mecanismos de Produccion en ecosistemas
lagunares costeros". Clave CONACYT: QCMABFR-001698.
Les données de méthodologie, les sites de prélèvements ayant été exposés ou définis dans le travail précédent, nous ne reviendrons pas sur ces points. Nous insisterons simplement sur le fait suivant: dans le cas de la faune mobile hôte des herbiers d'Halodule et deThalassia de la Lagune de Términos, les Gammariens, en abondance et à la suite des Caridea, constituent le groupe faunistique le plus important ce qui justifie leur étude après celle des Caridea. Dans le cas des herbiers d'Halodule les Gammariens représentent en moyenne, d'après nos données, près de 6% de la population et dans ce cas ils sont précédés par les Caridea (63%) et les jeunes Penaeidea (22%). Dans les herbiers de Thalassia alors que les Caridea représentent globalement 60% de la population, les Gammariens avoisinnent les 20% (26.56% à 17.39%) suivant que l'on prenne en compte prélèvements nocturnes ou non. Il paraît d'ailleurs presque certain que durant la période nocturne ils constituent, toujours en abondance, le groupe dominant. En ce qui concerne l'étude systématique proprement dite, cette faune de Gammariens apparaît relativement bien connue et très semblable à celle que l'on recontre dans les herbiers de Phanérogames des côtes nord est américaines. Dans ces prélèvements, nous avons reconnu 17 espèces. Les plus abondantes sont très largement représentées dans les herbiers nord est américains (Cymadusa compta, Elasmopus levis etc.). Quelques autres, parfois bien représentées, sont peu connues:Gitanopsis laguna, Melita planaterga, Cerapus benthophilus, Lembos unicornis. L'une, connue des côtes ouest africaines, est signalée pour la première fois: Gammaropsis togoensis.Enfin, une semble nouvelle pour la science: Nasageneia yucatanensis n. sp. Nous terminerons en indiquant qu'à coté de nos propres récoltes, des prélèvements, effectués par l'équipe d'Ecologia costera de l'Instituto de Ciencias del Mar y Limnologia de l'UNAM, ont été examinés lors de notre séjour à México. Nous n'avons pas observé, de façon formelle, d'autres espèces que celles dont il est question ici, mais, par contre, nous avons noté que certaines des espèces qui apparaissent mal représentées dans le cadre de ce travail pouvaient être, dans certaines stations et à certaines époques, beaucoup plus abondantes: Lembos unicornis, Cerapus benthophilus, Corophium divers Paraphoxus spinosus et Gammaropsis togoensis. ETUDE SYSTEMATIQUE
AMPELISCIDAE
Ampelisca vadorum Mills, 1963 Figure 1AMills, 1963: 972, fig. 1-3 Bousfield, 1973: 135, pl. 37 (1). Description: (femenino) de 11 mm, Estero Pargo, 16/ 08/ 1982. Al depassant le pédoncule de 1'A2 cette dernière étant approximativement égale à la moitié de la longueur du corps. Extrémité céphalique relativement large à deux paires de lentilles. Epimère 3 anguleux mais sans ornementation particulière. Processus dorsal de l'urosomite 1 abruptement tronqué. Patte 7 sans caractère remarquable, à article ischial nettement plus long que l'article méral. Uropodes trapus et peu épineux. Telson orné de une petite épine apicale à l'extrémité de chacun des lobes telsoniques. Cette espèce apparaît un élément constant de la faune des herbiers des côtes nord est américaines et est très voisine de A. abdita avec laquelle elle peut prêter à confusion (cf. Fox et Bynum, 1975). Elle s´en distingue (Bousfield, 1973, par le bord latéro-dorsal de l'urosomite 2 + 3 anguleux, l'uropode 1 qui n'atteint pas l'apex des uropodes 2 et la rame externe des uropodes 2 armée de 3 épines. Matériel: St. 1 (1); 4 (2): Herbiersd'Halodule. Récoltes UNAM, groupe d'Ecologie côtière: Estero Pargo 16/08/ 1982 (2), herbiers deThalassia. AMPHILOCHIDAE
Gitanopsis laguna McKinney, 1978 Figure 1BMcKinney, 1978: 140, fig. 3-4. ? G. petulans: Karaman, 1980: 53, fig. 1-4. Karaman décrivant l'espèce G. petulans la différencie des quatres du genre par son gnathopode 1 subchéliforme, la telson court (moins de deux fois plus long que large et à apex largement arrondi), le carpe du gnathopode 2 recouvrant presque totalement (3/4) le bord inférieur du propode, le lobe interne des MX2 peu soyeux (2 à 5 soies), les yeux plus grands que le diamètre du premier article du pédoncule atennulaire, l'épimère 3 peu anguleux. 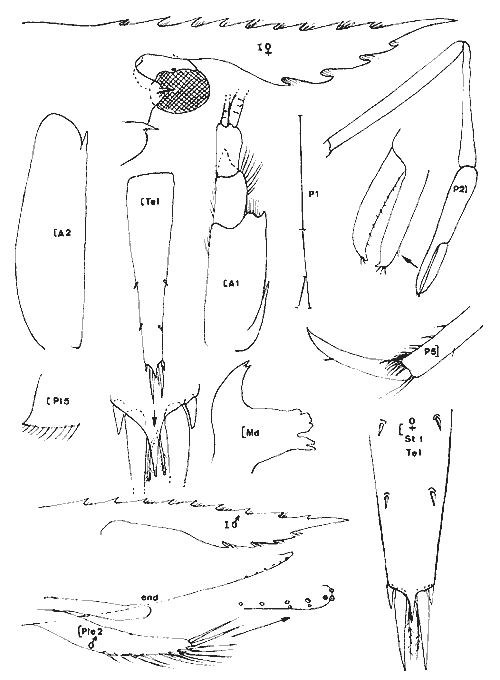 Fig. 1. A: Ampelisca vadorum, Mills, 1963: (femenino) de 11 mm, Estero-Pargo, 16/08/82. (réc. UNAM)B: Gitanopsis laguna McKinney, 1978: (femenino) de 3 mm, St. 17 (réc. Ledoyer). Il différencie l'espèce de G. lagunaessentiellement par les lobes latéraux céphaliques arrondis chez la première et légèrement anguleux chez la seconde: l'observation des figures de McKinney montre que cette différence est très ténue. Les spécimens de la Lagune de Términos par certains caractéres (coxales 1 à 3 avec une microencoche, lobes latéraux céphaliques parfaitement arrondis, palpe mandibulaire relativement grêle, lèvre antérieure bien dissymétrique) correspondet à G. petulans. Par l'ornementation des gnathopodes 2 (épines du bord dorsal du propode), la dilatation apparemment plus marquées de l'article méral des P5 à P7, l'espèce correspond à G. laguna. Décrivant G. petulans, Karaman n'a observe qu'un unique spécimen et en définitive, je me demande si les deux espèces citées ici ne sont pas synonymes d´autant plus que nos spécimens présentent le même type de répartition que G. laguna. Finalment par antériorité, si la synonymie s'avère juste, je considère que l'espèce est référable à G. laguna. G. tortugae Shoemaker, 1933; souvent signalée des herbiers de Floride, (Stoner, 1983, Lewis et Stoner, 1983) se différencie aisément de cette espèce par ses yeux réduits et surtout par le gnathopode 2 à bord palmaire beaucoup plus étroit. Matériel:L'espèce apparaît comme l'un des éléments constants de la faune mobile des feuilles des herbiers de la Lagune de Términos (voir Tableau). Elle semble aussi présenter une forte activité nocturne: 23 et 359 individus capturés au même endroit, dans le même volume et à une heure d'intervalle, la seconde récolte ayant été réalisée lors du passage du soleil derrière la ligne d'horizon. AMPHITHOIDAE
Cymadusa compta (Smith, 1873) Figure 2Grubia compta: Holmes, 1905: 510 (non vu) - Kunkel, 1918: 151, fig. 45 (non vu). Not Pearse, 1913: 376, fig. 6. Amphithoides comptus, Stebbing, 1906: 645. Cymadusa compta: Mills, 1964: 21, fig. 5 - 6 - Bousfield, 1973: 182, fig. 55 (2). Description: Le (masculino) adulte (7 mm et plus) de l`espèce est parfaitement caractérisé dans le genre par son gnathopode 2 à carpe légèrement plus long que le propode et à angle palmaire nettement marqué. En effet, chez les espèces du genre le carpe du gnathopode 2 du (masculino) est toujours nettement plus court que le propode, à l'exception de C. hawaiensis et de C. oceanica chez lesquelles le carpe du gnathopode 2 du (masculino) bien que relativement long, est toutefois plus court que le propode. Chez la (femenino) ou le (masculino) juvénile de l'éspèce, le carpe du gnathopode 2 est nettement moins long que chez le (masculino) adulte mais il demeure cependant subégal au propode ce qui est aussi inhabituel chez les (femenino) ou (masculino) juvéniles des autres espèces du genre. Chez le (masculino) juvénile le bord dorsal du propode et du carpe du gnathopode 2 est plus pileux que chez la (femenino) et ces deux articles sont comparativement plus dilatés; de plus l'angle palmaire tend á se dessiner. Comme chez la (femenino), par contre, le bord antérieur de l'article basal des gnathopodes est quasiment glabre. Matériel: L´espèce apparaît de façon constante dans tous les types d'herbiers prospectés dans ce travail et y constitue l'élément dominant parmi les gammariens sauf, peut-être, au niveau desHalodule du fond de la Lagune de Términos oùGammarus (Mucrogammarus) sp semble prendre le dessus. L'animal enfin semble présenter de remarquables variations nycthémèrales (cf. Tableau 1). COROPHIIDAE
Corophium cf. insidiorum Crawford, 1937 Figure 3BShoemaker, 1947: 53, fig. 6-7; Bousfield, 1973: 203, pl. 63(1), 66(3), 67(C); Myers, 1982: 193, fig. 128. La signalisation de cette espèce est basée sur l'observation d'un unique spécimen (masculino) incomplet (métasome et urosome manquant). Elle doit donc être considérée avec circonspection. Je référe l'animal à cette espéce pour les raisons suivantes, étant entendu que la structure de l'urosome n'est pas connue: — le rostre est aigu et bien prolongé; — le premier article du pédoncule d'A1 est armé d`une dent latéro-interne. 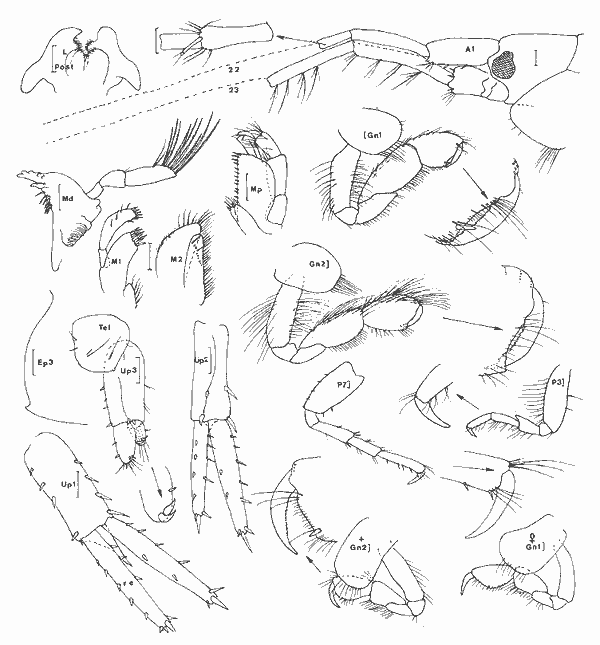 Fig. 2. Cymadusa compta (Smith, 1873): (masculino) de 8. 5 et (femenino) de 9.5 mm, St. 17 (réc. Ledoyer). Remarquons toutefois que cette espéce pourrait correspondre à C. lacustredont le rostre chez le (masculino) est bien développé (cf. Shoemaker, 1947: Fig. 10) du fait de la pilosité de son A1 et de la forte dilatation du pédoncule de l'A2. Cette espèce d'autre part est fréquemment citée des herbiers de la région Est américaine. Toutefois, la morphologie du pédoncule de l'A1 s'oppose à cette détermination: le premier article du pédoncule de l'A1 (masculino) chez C. lacustre est totalement inerme. Quant à C. ellisiqui apparait fréquent dans certains herbiers (Stoner, 1983), il posséde (masculino) une A1 à premier article du pédoncule bien particulier: il est bidenté sur son bord inférieur (cf. Shoemaker, 1943: 3, Fig. 1 a et c) et le dactyle du gnathopode 2, portant une rangée de soies spiniformes, ne posséde qu'une unique dent apicolatérale., Matériel: St. 16(1 (masculino) unique et incomplet). 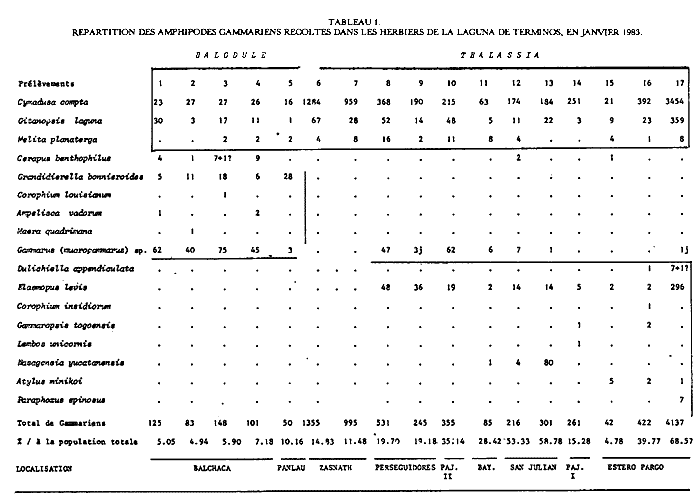 TABLEAU 1. REPARTITION DES AMPHPODES GAMMARIENS RECOLTES DANS LES HERBIERS DE LA LAGUNA DE TERMINOS, EN JANVIER 1983. 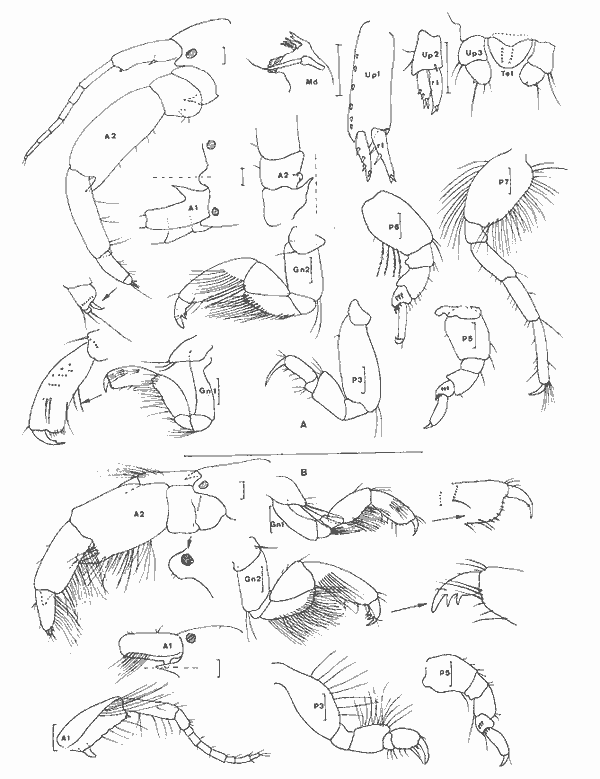 Fig. 3. A: Corophium louisianum Shoemaker 1934: (masculino) incomplet, St. 16 (réc. Ledoyer). B: Corophium cf. insidiorum Crawford, 1937: (masculino) de 2.6 mm, Zasnath 10/06/82 (réc. UNAM). Corophium louisianum Shoemaker, 1934 Figure 3AShoemaker, 1934: 30; 1947: 63, fig. 11; Heard, 1982: 39, fig. 44 (a-c). Description:(masculino)de 2.6 mm, réc. UNAM, Zasnath 10/ 06/ 82. L'espèce posséde un (masculino) aisément identifiable: le premier article du pédoncule de l'A1 est glabre et armé d'une dent latéro-interne; le rostre peu développé est arrondi; l'A2 glabre présente un cône excréteur particulier: il est remarquablement saillent sur le bord interne et ventral. Matériel: St. 3 (1) Réc. UNAM, Zasnath, 10/06/82. Gammaropsis togoensis (Schellenberg, 1925) Figure 4Eurystheus togoensis: Schellenberg, 1925: 177, fig. 23 - 1939: 136, fig. 29; Sivaprakasam, 1970: 283, fig. 1. Gammaropsis togoensis: Krapp-Schickel & Myers, 1979: 459, fig. 12-13. Description:(masculino) de 3.3 mm, Estero Pargo, 16/ 08/1982. L'espèce est bien caractérisée par ses lobes latéraux céphaliques remarquablement développés et le bord inférieur de la tête très encoché (deeply recessed). Le bord dorsal est lisse: au niveau de l'urosome 1 et 2 il y a uniquement une petite sétule latéro-dorsale. Le gnathopode 2 est très particulier avec son carpe presque totalment télescopé et son propode très allongé, piriforme, à bord palmaire trés oblique présentant un processus prédactylaire arrondi et finement crénelé et un angle palmaire dentiforme que n'atteint pas le dactyle. Chez les grands mâles la dent de l'angle palmaire peut être remarquablement saillante (voir figure). Les A1 et A2 sont subégales, nettement plus courtes que le corps; flagelle accessoire de 3 à 4 articles (sur le même individu) et flagelle des A1 et A2 de 7 segment. Cône éteur situé légérement sur le bord interne du pédoncule de l'A1 A2 mais très anguleux. Pas d'épistome saillant. Palpe mandibulaire robuste à second article à peine plus long que le 3ème. Lobe interne de la MX1 garni de 6 petites soies, celui des MX2 avec des soies médiolobulaires (une dizaine environ). Mxp classique, à palpe de 4 articles sans saillie au niveau apical du 3ème article. Coxales 1 à 4 arrondies. P3 et P4 identiques, relativement robuestes et à article méral bien dilaté. P5 à article basal ovalaire, élargi, presentant un petit lobe inféro-postérieur; bord postérieur à peine identé. P6 et P7 quasiment semblables à article basal piriforme, sans lobe inféro-postérieur. Epimère 3 arrondi. Uropodes biramés, épineux, trapus; rames subégales au pédoncule qui au niveau des uropodes 1 et 2 se prolonge par une forte dent apicale; cette dernière dépassant ou atteignant la moitié de la rame interne correspondante. Telson entier avec une épine émoussée au niveau des angles latéraux. (femenino): Comme le (masculino) mais avec un gnathopode 2 beaucoup moins développé: dactyle denticulé et bord palmaire légèrement échancré; angle palmaire armé d'une forte épine. Coloration: espèce portant des bandes colorées verticales (brunâtre après conservation à l'alcool) généralement au niveau des mésosomites 3, 5 et 7 du métasomite 1. Cependant cette disposition n'est pas absolue. Répartition: Seules deux espèces du genre étaient signalées des côtes atlantiques Nord Américaines:G. maculata et G. sutherlandi cette dernière récemment décrite par Nelson (1981) de la Caroline du Nord. La répartition de G. togoensis est assez surprenante puisqu'elle a été signalée des côtes du Togo, du Congo, des Indes et d'Israel en Méditerranée. Les spécimens de la Lagune de Términos sont parfaitement identiques aux individus décrits de Méditerranée. Matériel:St. 14 (masculino); 17 (2 (femenino)). Récoltes UNAM, groupe d'Ecologie côtière: Estero Pargo 16/08/1982 (3 (masculino), 1 (femenino)); Punta Gorda, 03/ 12/ 1881 (4 (masculino), dont 1 (masculino)(masculino) ). L'espèce n'apparaît pas rare dansles herbiers de Thalassia de la Laguna de Términos j'ai pu l'observer dans diverses récoltes faites par l'équipe d'Ecologie côtière. Grandidierella bonnieroides Stephensen, 1948 Figure 5BMyers, 1970: 141, fig. 1-2 (références); Fox & Bynum, 1975: 225; Ledoyer, 1982: 245, fig. 89 (références). Descriptión: (masculino) de 3 mm environ, Balchacáh, St. 3 (réc. Ledoyer). 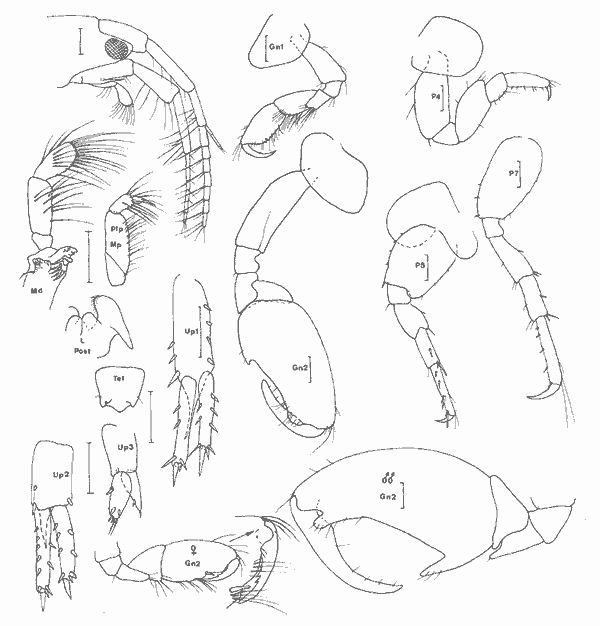 Fig. 4. Gasmmaropsitogoensis (Schellenberg, 1925): (masculino) de 3.3 mm et (femenino) Estero Pargo, 16/08/82 et (masculino) (masculino) Punta Gorda, 03/12/81 (réc.UNAM). Caractérisée par sa coxales l arrondie, les processus sternaux dentiformes sous les mésosomites 1 et 2, le gnathopode 1 carpochéliforme à bord palmaire bifide et à bord postérieur du carpe avec une dent accessoire. Uropode 3 uniramé, à rame nettement plus longue (3 fois environ) que le pédoncule. Chez cette espèce, le flagelle accessoire apparaît absent dans le cas des spécimens de la Lagune de Términos. De plus, le flagelle de l'A2 est beaucoup plus robuste et plus épineux que pour les spécimens que Myers (1970) figure du Venezuela ou de Colombie; il semble assez identifique à celui des individus des Indes occidentales. 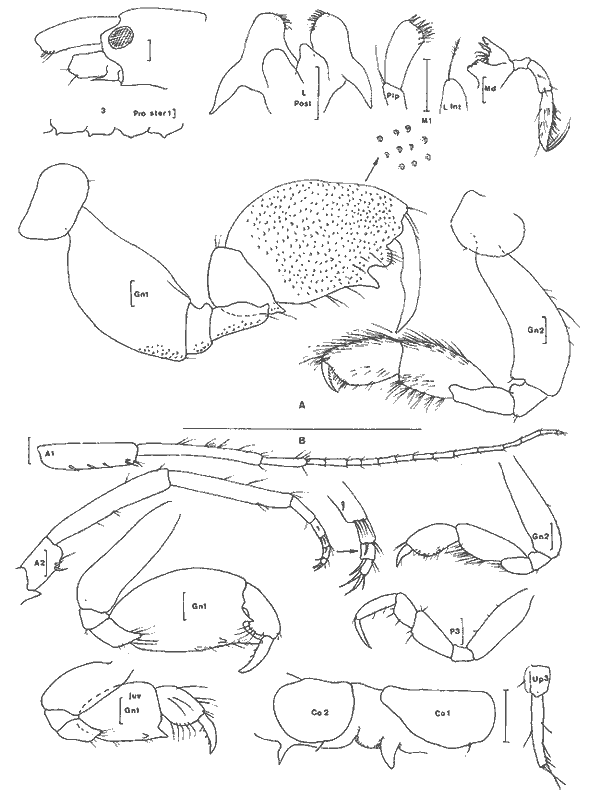 Fig. 5. A: Lembos unicornis Bynum et Fox, 1977: (masculino) de 5.4 mm, Punta Gorda, 03112181 (réc. UNAM)., B: Grandidierella bonnieroides Stephensen, 1948: (masculino) de 3 mm, St. 3 (réc. Ledoyer). Matériel:St. 1 (5); 2 (11); 3 (18); 4 (6); 5 (28); 17 (1? en très mauvais état). Notons que l'espèce semble se cantonner essentiellement dans la partie la plus saumâtre de la Lagune de Términos. Cependant, les herbiers d'Halodule n'ayant pas été étudiés dans les autres secteurs plus directement soumis aux influences marines, il serait actuellement dangereux de conclure de façon absolue quant à ce type de répartition. Lembos unicornis Bynum et Fox, 1977 Figure 5AMicrodeutopus anomalus Kunkel, 1910: 76, fig. 29 (not Rathke, 1843) Lembos sp Williams et Bynum, 1972: 178; Fox et Bynum, 1975: 225 Lembos unicornis: Bynum et Fox, 1977: 23, fig. 15-16; Myers, 1977: 146, fig. 148-152; Myers, 1981: 26, fig. 10-11, pl. 1C. Description: (masculino) de 5.4 mm, Punta Gorda, herbiers de Thalassia, récolte du 3/12/81, UNAM. Dans nos récoltes nous n'avons pas recontré l'espèce de façon certaine. Toutefois, l'observation de divers prélèvements faits par l'équipe d'Ecologie côtière de l'UNAM montre que cet animal peut parfois se recontrer assez abondamment dans la faune mobile des herbiers deThalassia et il convient donc de le signaler. Comme l'indique Myers (1981), cet animal est bien caractérisé d'une part, par sa coloration (tête, mésosomites 2 à 5 et métasomites 1 à 3 brunâtres: cette coloration persistant sur nos échantillons stockés dans l'alcool, dépuis deux ans et demi) et d'autre part, par son gnathopode 1 à dactyle dépassant très largement la dent de l'angle palmaire et à tégument orné de nodosités régulièrement disposées, surtout au niveau du propode. De plus, les processus sternaux sont présents sons les mésosomites 1 à 4 mais ils apparaisent très émoussés. Matériel:Punta Gorda, 3/ 12/ 1981, quadrant, 6(masculino) , tous dépourvus d'Al, A2 et P3 à P7. St. 14 (1(femenino) ?). DEXAMINIDAE
Atylus minikoi (Walker, 1905) Figure 6Nototropis minikoi: Shoemaker, 1932: 199; Pillai,. 1957: 47, fig. 8. Atylus minikoi:Sivaprakasam, 1968: 97, fig. 7; Bynum et Fox, 1977: 15, fig. 9-11; Ledoyer, 1982: 336, fig. 124 (avec réferences). ?Atylus urocarinatus Mc Kinney, 1980: 84, fig. 1-2. Description: (femenino) de 5.4 mm, Estero Pargo, St. 15 (réc. Ledoyer). Atylidae fort bien caractérisé par son bord dorsal pourvu d'une dent médio-dorsale uniquement au niveau de l'urosomite 1 (femenino) et l'article basal de la P5 dépourvu de toute trace de lobe inféro-postérieur (différence avecA. swammerdami). Seule l'espèce de Nouvelle Zélande, A. taupo J.L. Barnard, 1972 a peut prêter à confusion avec A. minikoi, toutefois chez la première l'article basal de la P7 est apparemment plus dilaté et le lobe inféro-postérieur semble nettement plus accusé. Dans le cas de la (femenino) que nous avons figurée, l'une des A2 était aberrante: les deux derniers articles du pédoncule étaient nettement plus court que la normale. Au niveau des MX1 les lobes internes portaient respectivement 3 et 4 soies et les palpes 5 et 8 dents apicales. Chez le (masculino) l'urosomite 2 + 3 est aussi dentiforme au niveau médio-dorsal et les articles distaux du pédoncule de l'A2 portent de petites touffes de soies. Mc Kinney (1980) décrivant du golfe du MexiqueAtylus urocarinatus ne compare malheure usement pas l'espèce à A. minikoi pourtant signalée du secteur par Shoemaker, (1932) et Bynum et Fox, (1977). Il semblerait en fait et à première vue que l'espèce citée des côtes Est américaines soit A. urocarinatuscaractérisée essentiellement par son telson à deux paires d'épines Sivaprakasam (1968) des Indes; Ledoyer de Madagascar figurent un telson à une seule paire d'épines. Cependant Pillai, 1957 figure et écrit que chez ses spécimens des Indes (masculino) le telson est orné de 2 paires de "soies". Ce caractère ne peut donc être retenu. Quant au lobe interne des MX1 il porte 3 ou 4 soies plumeuses suivant les descriptions: Pillai des Indes, Ledoyer de Madagascar, Mc Kinney du golfe du Mexique en signalent 4; Bynum et Fox de la Caroline du Nord en comptent 3; enfin, dans le cas présent, chez un même spécimen nous avons trouvé 3 et 4 soies. Cet autre caractère ne permet donc pas actuellement une discrimination spécifique. 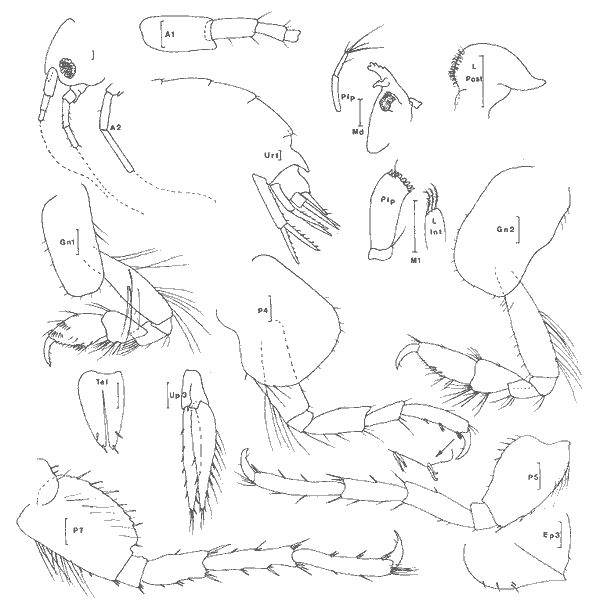 Fig. 6. Atylus minikoi (Walker, 1905):(femenino) de 5,4 mm, St. 15 (réc. Ledoyer). Nous terminerons en signalant que l'article basal de la P7 des spécimens de la laguna de Términos semble présenter un bord postérieur plus épineux que celui des spécimens malgaches. Revoyant les rares individus de Madagascar, j'ái observé qu'ils pouvaient présenter un bord postérieur de l'article basal plus épineux que celui que j'ai figuré (1982, Fig. 124). Enfin, dans leur figuration, Bynum et Fox ont trés certainement interverti les P6 et 7 (Fig. 10 D et E). Matériel: St. 15 (5); 16 (2 (masculino)); 17 (1). EUSIRIDAE
Genre Nasageneia J.L. Barnard et Karaman, 1982Le genre Portogeneia Boeck, 1871 a été scindé depuis 1972 en quatre genres distincts:Pontogeneia Boeck, 1871, Gondogeneia etTethygeneia J.L. Barnard, 1972b etNasageneia J.L. Barnard et Karaman, 1982. Les genres Gondogeneia etTethygeneia se distinguent essentiellemente par leur rostre court chez le premier et relativement développé chez le second. J.L. Barnard et Karaman, 1982 créent le nouveau genreNasageneia pour distinguer les espèces deTethygeneia à épimère 3 denticulé (serrate) et y incluent N. nasa (J.L. Barnard, 1969) etN. quinsana (J.L. Barnard, 1964). Il ne fait aucun doute que l'espèce recontrée dans la Lagune de Términos doit être référée à ce genre actuellement connu des côtes du Pacifique (Basse Californie et Golfe de Californie) et apparaît remarquablement voisine de ces deux espèces. Pontogeneia longleyi Shoemaker, 1933 etP. bartschi Shoemaker, 1948 décrites de Floride et de Cuba sont actuellement référées au genreTethygeneia car elles possédent un épimère 3 parfaitement lisse. Toutefois, denticulations de l'épimère 3 mises à part il est remarquable de constater l'étonnante analogie morphologique de T. longleyi, N. nasa et N. yucatanensis n. sp. Nasageneia yucatanensis n. sp. Figure 7Description: (femenino) holotype de 4.2 mm (P5 et A2 incompletes), San Julian, St. 13. Al sans flagelle accessoire, plus courte que le corps et un peu plus courte que l'A2 (d'après une (femenino) paratype). Bord dorsal apparaissant denté médiodorsalement au niveau des trois métasomites: en réalité, le bord dorsal est lisse mais il présente de grands décrochements à ce niveau de sorte, qu'à première vue, l'espèce semble dorsalement dentée. Rostre très recourbé, bien développe. Lobes latéraux céphaliques largement arrondis. Oeil développé, circulaire, brun froncé. Lévre antérieure entière. Lévres postérieures sans lobes internes (ceux-ci sont à peine indiqués par deux petits replis). Mandibule à processus inciseur et lacinia mobilis très dentelés; processus molaire saillant, triturant et très crénelé; 3 épines molaires environ; palpe triarticulé, robuste, avec le second article le plus développé et orné de soies barbelées apicolatérales, troisième article falciformes à soies (3) spiniformes distales et à soies (4) barbelées latérales. MX1 á palpe biarticulé portant de petites (6 à 7) épines distales; lobe externe armé d'épines très crénelées; lobe interne à 3 soies apicales plumeuses. MX2 bilobée, ornée de soies apicales; lobe interne avec deux grosses soies apicolatérales et 6 + 7 soies apicales. Mxp à palpe robuste, de 4 articles; lobe externe bien développé, bordé d'une dizaine d'épines et de soies; lobe interne distalement armé de soies, de 2 épines arrondies et d'une épine interne aigue. Coxales arrondies, plus hautes ou aussi hautes que larges (1 à 4), à peine festonnées sur leur bord inférieur. Lobes branchiaux des P5 et P6 très plissés. Gnathopodes 1 et 2 subchéliformes, subégaux, peu robustes, à carpe nettement plus court que le propode régulièrement ovalaire et à angle palmaire défini par des épines; carpe du gnathopode 2 légèrement saillant au niveau antéro-inférieur. P3 et P4 semblables, à article basal relativement court et à article méral presque aussi large que long; propode le plus développé; dactyle robuste, orné d'une rangée de spinules. P5 à P7 de longueur régulièrement croissante et morphologiquement identiques: article basal dilaté, ovalaire, à lobe inféro-postérieur arrondi et bien développé; bord postérieur orné de légers festons. Epimère 2 armé d'un petit redent inféropostérieur. Plaque épimérale 3 régulièrement arrondie mais présentant sur son bord postérieure 5 à 6 fines indentations. Uropodes biramés. Uropodes 1 et 2 à rames dissymétriques, peu épineuses et á bords spinuleux; rame interne la plus développée, subégale au pédoncule. Uropode 2 nettement plus court que le premier. Uropodes 3 à rames subégales, épíneuses, ornées de nombreuses soies plumeuses, étroitement lancéolées et beaucoup plus longues que le pédoncule qui est armé, sur son bord interne, de 4 à 5 longues épines. Telson un peu plus long que large, fendu sur la moitié de sa longueur et à apex des lobes telsoniques régulièrement arrondi. (masculino): très semblable à la (femenino) mais avec des yeux beaucoup plus volumineux, presque coalescents dorsalement. Pédoncule antennaire plus élargi. 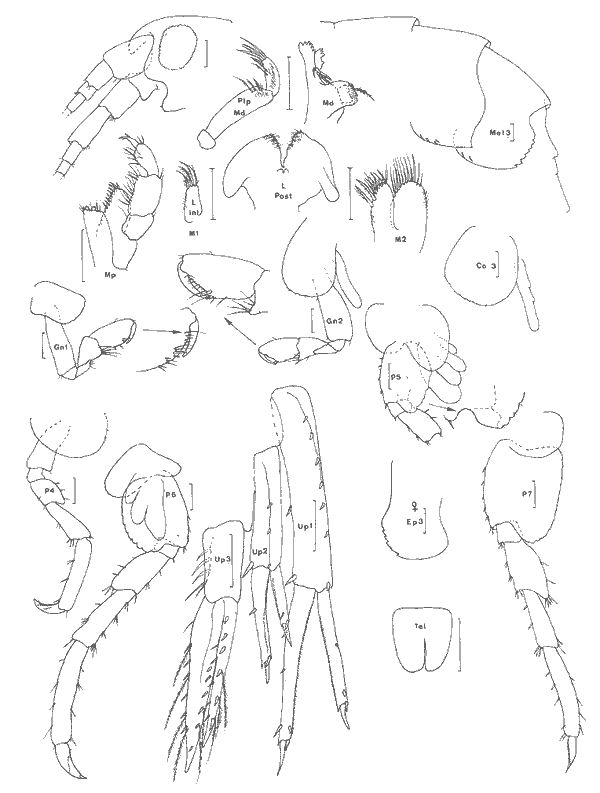 Fig. 7. Nasageneia yucatanensis n. sp. (femenino) holotype de 4.2 mm, St. 13 (réc. Ledoyer). Materiel: St. 11 (1), 12 (4), 13 (80). Notons que l'espèce paraît localisée dans la zone la plus marine de la Lagune de Términos. Affinités: Comparativement àN. quinsana l'espèce des eaux de la Lagune de Términos possède des lobes latéraux céphaliques plus largement arrondis, un carpe des gnathopodes ( (femenino)uniquement) plus court et un telson plus profondément fendu (l/2 et non 1/4). Comparée à N. nasa on notera chez N. yucatanensis que l'angle inférieur de la tête est moins anguleux, le lobe de l'article basal de la P7 est plus arrondi et les P3 et P4 sont plus robustes. Toutefois, ce sont essentiellement des raisons géographiques qui m'incitent à distinguer cette espèce atlantique de celle du Pacifique. Remarquons, que, mis à part la denticulation de la plaque épimérale 3, cette espèce apparaît quasimement identique à Tethygeneia longleyi de Floride . GAMMARIDAE
Dulichiella appendiculata (Say, 1818) Figures 8-9Melita fresnelii: Kunkel, 1910. 31, fig. 11 Karamant et Barnard (1979) rétablissent le genre Dulichiella Stout, 1912. De plus, ils signalent que les mises en synonymies antérieures sont à revoir et probablement abusives. Ils estiment que le genre compte environ 5 espèces:D. appendiculata (Say, 1818), D. australis (Haswell, 1879), D. exilii(Müller, 1864), D. fresneli (Audouin, 1826) et D. spinosa (Stout, 1912). Cette opinion est suivie par Berents (1983). Karamar (1981) adjoint à cette liste D. grandimana (Chevreux, 1908). Melita subchelataSchellenberg, 1925 (sensu K.H. Barnard, 1932) entre elle-même, nous semble-t-il, dans ce genre et apparaît bien caractérisée par l'article basal de ses P6-P7. Quant à Melita excavata Ledoyer, 1979 sa position reste incertaine puisque l'unique spécimen holotype n'était en possession que d'un de ses gnathopodes 2; toutefois, la structure de son palpe mandibulaire est assez particulière et sa formule médiodorsale ne présente aucune interférence avec le complexe D. appendiculata.
En effet, J.L.
Barnard (1955) mettait en synonymie
Stebbing (1906) avait mis en
synonymie les diverses espèces de Melita à gnathopodes 2
(masculino) dissymétriques avec M. fresneli et considérait M.
appendiculata comme une espèce douteuse. Il fut suivi dans cette optique
par Spandl (1924) etc.
A l'origine D. appendiculata a été décrite des côtes de Georgie alors queD. fresneli l'était d'Egypte (Mer Rouge). On pourrait donc penser que les formes Indopacifiques sont réferables à la seconde espèce, les formes atlantiques l'étant à la première. Quant à Dana (1852) il décrit deux espèces du genre: D. valida de Singapoure etD. setipes de Rio de Janeiro. Il signale et figure aussi, de cette même localité, D. anisochir(Kröyer) qu'il estime identique à D. pilosusqu'il avait signalée auparavant. Malheureusement ces descriptions restent très sommaires et on ne posséde pas d'indication sur les formules mediodorsales si ce n'est que les pléonites 1 à 5 sont dentés.
Le gros
problème concernant ce genre reste la définition des deux
espèces D. appendiculata(Nord Atlantique) et
D. fresneli (Mer Rouge: signale l'espèce d'Egypte
mais celle-ci n'ayant pas été trouvée à ce jour en
Méditerranée, il est probable qu'elle provient, à
l'ongine, de la Mer Rouge). En ce qui concerne cette seconde espèce, on
peut penser que la description de Spandl peut être prise comme base.
Pour D. appendiculata,pourtant fréquente le long des
côtes atlantiques nord américaines et particulièrement
dans les herbiers de phanéreogames marines (Nelson (1980) signale que
l'espèce est commune dans les, Zostera marina de la
Caroline du Nord; Stoner (1983) la récolte, parfois abondanment, dans
lesThalassia de Floride) onane posséde,
d´ailleurs sous le nom de D. fresneli, que des citations
Pearse (1913), Shoemaker (1935) et les figurations de Kunkel (1910)
Kunkel
figure et décrit les processus du ord palmaire du grand gnathopode 2 du
(masculino) comme arondis. Toutefois, comme cour spécimens, c'est la
dent prédactylaire qui est la plus saillante et la 3éme
apparaît la plus rétduite.
On peut raisonnablement penser que ces références s'appliquent à l'espèce décrite par Say et que l'espéce rencontrée dans la Lagune de Términos est représentative de D. appendiculata d'autant plus que les gammariens accompagnateurs de l'espéce dans ce secteur sont identiques à ceux des herbiers de Floride ou de la Caroline(Cymadusa compta, Elasmopus levis, Gammarus "mucronatus"). 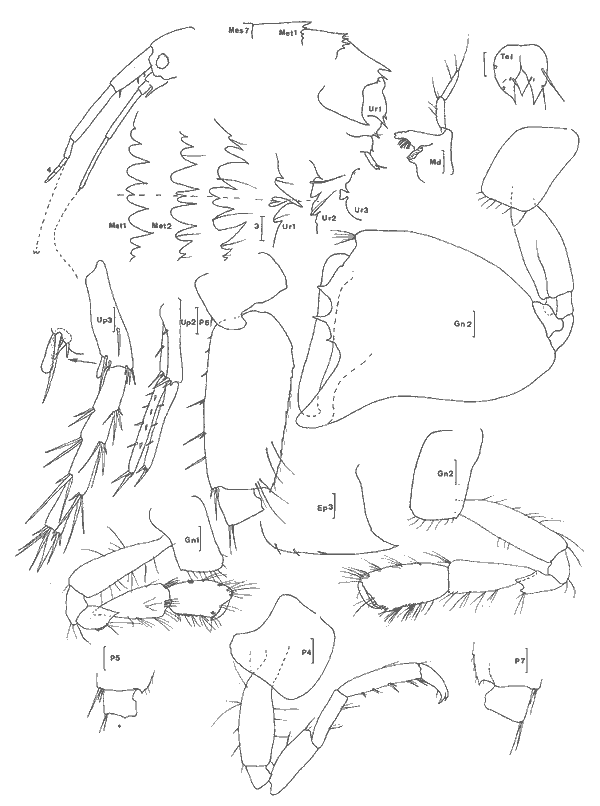 Fig. 8. Dulichiella appendiculata (Say, 1818): (masculino) de 6 mm, St. 17 (réc. Ledoyer). 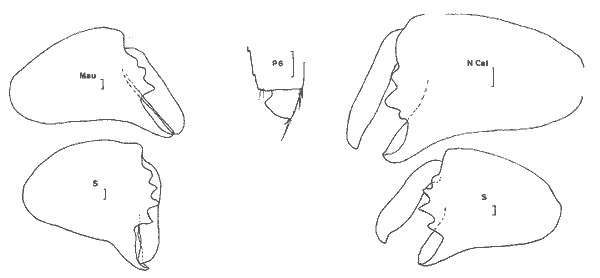 Fig. 9. Dulichiella appendiculata sensu lato. S: spécimen du Sénégal, 7 m, 15/12/80 (réc. Leung Tack); N. Cal. spécimen de Nouvelle-Calédonie, St. 7 (réc. Ledoyer), 4 m, récif Abore, 12/04/79; Mau. spécimen de l'il Maurice, St. 16 (réc. Peyrot-Clausade), 25 m, pente externe du Trou aux Biches, 1974. Considérant l'espèce de la Lagune de Términos comme représentative de D. appendiculata (Say, 1818), je l'ai comparée a des spécimens du Sénégal (Réc. Leung Tack, en cours d'étude), de Madagascar, de l'île Maurice et de la Nouvelle Calédonie. Dans le cas de ces deux derniers secteurs, je n'avais pas figuré de spécimens de cette espèce (Ledoyer, 1978: 282 et sous-presse). J´ai cru bon ici de donner quelques indications sur ces individus. Quoiqu'il en soit, l'espèce recontrée dans les herbiers de la Lagune de Términos se caractérise, comparée aux individus du Sénégal, de Madagascar, de l'île Maurice et de Nouvelle Colédoenie par divers caractéres dont le plus important et, à mon sens, le moins suggestif est: — la structure de l'article basal des P5 à P7 qui présente un petit éperon dentiforme inféropostérieur alors que chez “les autres espèces” ce bord est arrondi. Ce caractère est parfaitement figuré par Barnard (1970) pour les spécimens d'Hawaii et par Reid (1951) pour ceux des côtes de Guinée et de Sierra Leone. — le bord dorsal du métasome, de plus, parait plus épineux (cela mériterait évidemment d'être vérifié sur des séries d'individus) puisqu'il y a 9 dents dont une petite médio-dorsale au niveau des trois métasomites. Formule dorsale du métasomite l à l'urosomite 3 = 2 (2) 1 (2) 2 - 1 (3) 1 (3) 1 - 1 (3) 1 (3) 1 - 1 (1) 1 (1) - 1 1 1 - 2 - Chez les spécimens des divers secteurs que j'ai revus, on observe 7 dents dusmétasomite 1 au métasomite 3. Ajoutons toutefois que parfois, au niveau du métasomite 3 la dent la plus latérale apparaît légèrement bifide. — Chez les (masculino) bien développés (4), les dents du bord palmaire du grand gnathopode 2 sont aigues et la dent proximale (par rapport au dactyle) est la plus développée. Chez les representants des autres secteurs géographiques citées ci-dessus, ces dents sont toujours émoussées, au nombre de trois, et c'est la dent médiane ou distale qui apparaît la plus développée. — le palpe mandibulaire apparaît, tout au moins chez le spécimen disséqué, peu pileux. Pour le reste on observe un premier article du pédoncule antennulaire portant sur son bord inférieur trois épines, un lobe interne des MX1 à deux soies distales plumeuses, un lobe interne des MX2 avec une rangée de soies médiolobulaires. Ces caractéres sont donc identiques avec ceux des autres "formes". Matériel: St. 16 (1(femenino) en maivais état); 17 (5 (masculino) dont 1 juvénile et 2 (femenino)). L'espéce n'a été rencontrée, de façon significative, que dans le prélèvement effectué en début de nuit. En ce qui concerne la forme des côtes du Sénégal comparativement aux formes indo pacifiques (Madagascar, île Maurice, Nouvelle -Calédonie), toutes quatre à article basal des P5 à P7 à bord inféro-postérieur arrondi, elle posséde un bord dorsal du métasome beaucoup plus lipleux de même que l'article méral des P5 à P7. Il est probable que ces diverses formes qui demandent à être revues, d'une part en détail et d'autre part sur un certain nombre de spécimens, pourront ultérieurement être séparées avec plus de précision.
Actuellement, dans le cas des citations
où " l'espèce" a été figurée (grand
gnathopode 2 du (masculino)), je pense que l'on peut raisonnablement assimiler:
D. fresneli, dans sa forme typique d'après la
description de Spandl (1924), serait caractérisée par un
gnathopode 2 à dent intermédiaire dominante. Ceci correspond aux
descriptions de Berents (1983)
Forme décrite sous le nom de D.
appendiculata par cet auteur comme l`ont été toutes les
descriptions ou citations faites après 1970 à la suite de la mise
en synomymie faite par J.L. Barnard.
Cette modeste mise au point ne constitue en rien une revision, mais a pour seul but d'attirer l'atterition sur cette "espèce" ou plutôt sur ce groupe d'espèces du complexe fresneli et d'ébaucher une première mise en place des diverses "formes". Elasmopus levis Smith, 1873 Figure 10ASmith, 1873. non consulté; Barnard J.L., 1965. 497 (dans une clé); Bousfield, 1973: 63, pl. X (2). Non E. levis Kunkel. 1918 = E. Pocillimanus par Schellenberg, 1938: 56 et Berents, 1983: 117. NonE. levis K.H. Barnard, 1916: 200, pl,. 27 (15) =Maera vagans par K.H. Barnard, 1940: 459. Description: (masculino) de 7 mm, St. 17 (réc. Ledoyer), Estero Pargo. Parmi le genre Elasmopus seules les espèces E. levis et E. pocillimanus présentent un mâle à gnathopode 2 à bord palmaire oblique, regulièrement ovalaire et dépourvu de sinus, mais possédant une cavité latéro-interne dans laquelle peut se loger le dactyle. Ces deux espèces sont aisément différenciables: — chez E. levis le propode des P3 et 4 ne présente pas d'épines postéro-distales remarquables et striées comme chez E. pocillimanus; — chez E. levis les lobes du telson sont nettement anguleux et pourvus d'épines (1 + 2) à l'apex alors que chez E. pocillimanus les lobes telsoniques sont arrondis et les épines (1 + 0 à 2) sont latérales; — Enfin, chez E. levisl'épimère 3 est nettement anguleux. Chez E. pocillimanus il est de type arrondi bien qu'orné d'une petite dent inféropostérieure. Matériel: l'espèce est apparemment absente dans les herbiers d'Halodule du fond de la Lagune de Términos. L'espèce semble liée (voir Tableau 1) plus particulièrement aux zones soumises aux influences marines et paraît présenter des variations nycthémèrales importantes (2 et 296 individus capturés le même jour, pour le même volume traité au même endroit, à une heure d'intervalle. Le prélèvement considéré comme nocturne a été effectué juste après le passage du soleil sous la ligne d`horizon). 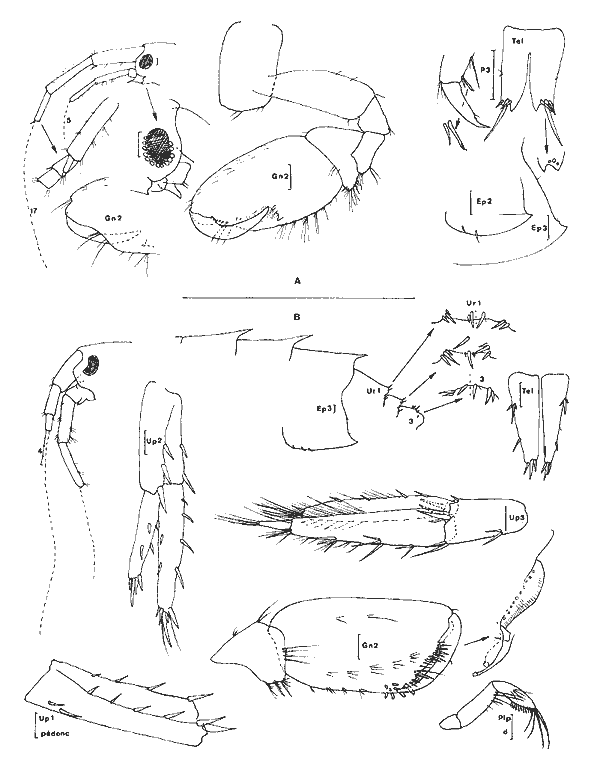 Fig. 10. A: Elasmopus levis Smith, 1873: (masculino) de 7 mm, St. 17 (réc. Ledoyer). B: Gammarus (Mucrogammarus) sp. (masculino) de 7 mm, St.3 (réc. Ledoyer). Gammarus (Mucrogammarus) sp. Figure 10BGammarus mucronatus Say, 1818 (=Mucrogammarus mucronatus) présente une très large distribution depuis le golfe du Saint Laurent jusqu´au golfe de México (Bousfield, 1973: 55, pl. 3 - 1 -) et a été signalé dans de très nombreux travaux traitant des gammariens des eaux estuariennes ou de la faune de herbiers de la côte atlantique Nord américaine. Toutefois, Farrell (1970) signale une espèce voisine de G. mucronatus despuis le Nord de la Floride jusqu'à l'embouchure du Mississipi et caractérisée par des dents médiodorsales beaucoup plus développées que chez cette derniére. Thomas (1976), Heard (1979) retrouvent cette forme"macromucronatus" dans les zones oligohalines. Dans le cas de nos récoltes, dans la Lagune de Términos il s'agit en fait de cette forme caractérisée comparativement à la description de Bousfield (1973) par: — une plaque épimérale 3 apparemment plus anguleuse; — un telson comparativement moins épineux; — des dents médiodorsales du métasome beaucoup plus saillantes; — enfin, contrairement à ce que figure Bousfield pour G. mucronatus, l'antenne est légèrement plus courte que l'antennule (vérifié sur divers spécimens des deux sexes). Je n'insisterai pas plus sur cette espèce qui paraît être en cours de description. Matériel: Ce gammare paraît curieusement présenter une double répartition (cf. Tableau 1). Présent dans les herbiers d'Halodule du fond de la lagune qui correspondent approximativement à la région la plus dessalée, il semble absent dans la zone intermédiaire et réapparaît dans la partie la plus soumise aux influences marines. Ceci semble assez contradictoire. Pourtant il s'agit bien de même espèce: mucronations dorsales, rapports A1/A2 identiques. Maera quadrimana (Dana, 1853) Figure 11Ortiz, 1978: 8; Ledoyer, 1982: 542, fig. 206 ( avec références); Berents, 1983: 128, fig. 22 (part) (avec références). Desirription:(masculino) de 7 mm, Punta Gorda, 03/12/1981. Espèce caractérisée par son gnathopode 2 (masculino) à bord palmaire transverse creusé de 3 sinus. Le telson à longues soies spiniformes (6 par lobe dans le cas du spécimen disséqué ici, mais il y en a moins chez les spécimens de plus petite taille) et les uropodes 1 et 2 à longues épines distales apparaissent aussi assez caractéristiques. Remarque:Cette espèce apparait très polymorphe quant à la structure de l'article basal des pattes 5 à 7 et la forme de la Lagune de Términos semble bien différente de celle de Mádagascar sur ce point et elle est intermédiaire entre les deux formes figurées par Schellenberg (1938, Fig. 21-22). L'espèce, essentiellement indo-pacifique, a été signalée de Cuba par Ortiz (1968) sans autre précision. Comparativement aux diverses formes indo-pacifiques décrites, j'ai noté, au niveau du gnathopode 2 du (masculino), chez l'espèce atlantique que le sinus précédant l'angie palmaire était toujours peu marqué (4 (masculino) observés) et que le dactyle présentait toujours une nette dilatation médiane. Le cas de cette "espèce" au sens large semble devoir être rapproché de celui de Dulichiella appendiculata sensu lato. Matériel: St. 2(1 (masculino)). Récoltes UNAM, groupe Ecologie côtière: Punta Gorda, 03/12/1981 (3 (masculino), 1(femenino)). Genre Melita sensu latoKaraman et Barnard (1979) scindent le genre d'ailleurs fort mal défini (cf. Ledoyer, 1982) en rétablissant le genre Dulichiella Stout (1912) qui regroupe les espèces à gnathopodes 2 très dissymétriques chez le mâle. Karaman (1981) redécrivant l´espèce Melita planatergaKunkel, 1910, scinde les espèces restant alors dans le genreMelita (Dulichiella exclues) en deux: — genre Melita sensu stricto Leach 1814 caractérisé par, d'une part un uropode 3 à rame externe simple et d'autre part un lobe interne des MX2 dépourvu d'une rangé de soies médiolobulaires. — genreAbludomelita Karaman, 1981 caractèrisé par les critères inverses: uropode 3 à rame externe biarticulée et lobe interne des MX2 pourvu d'une rangée de soies médiolobulaires. Ces diverses distinction paraissent parfaitement justifiées et fort utiles au niveau de ce genre complexe. 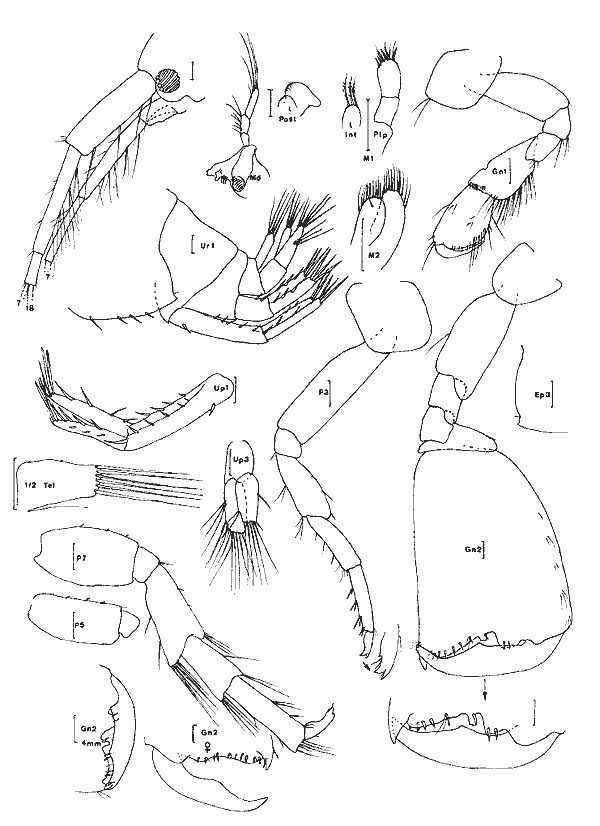 Fig. 11. Maera quadrimana: (masculino) de 7 mm et (femenino), Punta Gorda 03/12/81 (réc. UNAM) et (masculino) de 4 mm, St. 2 (réc. Ledoyer). Dans les herbiers de la Lagune de Térninos, deux espèces du genre, au sens large, ont été récoltées. L'une appartient au genre Dulichiella (voir précédemment), l'autre au genre Melita sensu stricto. Melita planaterga Kunkel, 1910 Figure 12Kunkel, 1910: 35, fig. 12; Karaman, 1981: 44, fig. 1-4, 5 (1-4).
Cette espèce, sensu Karaman, 1981 non Barnard,
1962 (107)
J.L. Barnard, 1962 considère que l'espèce ne
posséde aucune dent médiodorsale à quelque niveau que ce
soit. A ce sujet, la seule indication donnée par Kunkel reste ambigue:
"abdominal segments with posterior margin smooth".
Discussion: M. planterga Kunkel, 1910 sensu. Karaman, 1981 (44, Fig. I-IV, V (1-4) non sensu. Barnard, 1962 qui considére que l'espèce no posséde aucune dent médiodorsale à quelque niveau que ce soit, entre dans le groupe des Melita à urosomite l pourvu d'une dent médiodorsale et à urosomite 2 à une dent + une épine latérodorsales par coté. M. palmata (Montagu, 1804), M. festiva (Chilton, 1884), M. hergensis Reid, 1939, M. abyssorum, 1944 constituent ce groupe (cf. Barnard, 1962).M.festiva entre dans le genre Abludomelita etM. abyssorum est une espèce aveugle. M. reidi Hamond, 1965 et M. celericula Croker, 1971, décrites ultérieurement, si elle; possédent un urosomite l unidenté, elles montrent un urosomite 2 bidenté de chaque coté et s'intégrent dans le groupe M. orgasmos, M. solada etc. La première, de plus, est aveugle et est référable au genreAbludomelita. Karaman (1981) rétablit l'espèce M. sulca (Stout, 1912) que Shoemaker, 1941 estime synonyme de M. palmata. Shéridan (1980), de même que Barnard (1962), considere que M. planaterga est une espèce entrant dans le complexe des Melita, au sens large, à ucosomite l inerme telles que Barnard (1962) les énumère. A cette liste, il faut ajouter M. pahuwaii Barnad, 1970. M. elongata, M. intermedia, M. setosa toutes trois décrites par Shéridan (1980) qui propose d'ailleurs une clé pour la totalité des espèces de ce groupe. En définitive, dans le cas qui nous préoccupe plus particulièrement, trois espèces seulement restent comparables à celle des herbiers de la Lagune de Términos: M. planaterga sensu. Karaman, 1981: le telson présente des épines latérales sur son bord interne, les lobes apicaux sont relativement aigu et mis à part le plus grand nombre d'articles (4) au flagelle accessoire, il n'y a aucune différence entre la forme des Bermudes et celle du Yucatan. M. palmata présente un gnathopode 2 (masculino) très transverse, un telson à apex très anguleux avec les èpines en position très apicales et une èpine sur le bord interne des lobes (cf. Karaman, 1982, in Ruffo). M. hergensis a des lobes telsoniques peu anguleux ornés de deux paires d'épines qui sont en position plus médiane qu'apicale. Ceci nous semble la seule différence notable avec l'espéce du Yucatan (cf. Karaman, 1982, in Ruffo, Fig. 228). ISCHYROCERIDAE
Cerapus benthophilus Thomas et Heard, 1979 Figure 13Thomas et Heard: 98, fig. 1-4. Description: (masculino) de 4.8 mm (du rostre á la base du telson). Punta Gorda, 03/12/81, réc. UNAM, Ecologia costera. Animal maculé de brun, présentant un petit rostre anguleux, des lobes latéraux céphaliques arrondies et occupés par les yeux circularies et noirâtres. Péndoncules des A1 et A2 les articles distaux pourvus ventralement de longues soies. Al Légèrement plus courte que l'A2, toutes deux avec un flagelle pincipal de 7 segments; les deux ou tróis dermiers segments de ces flagelles portent une longue épine antéro-ventrale. 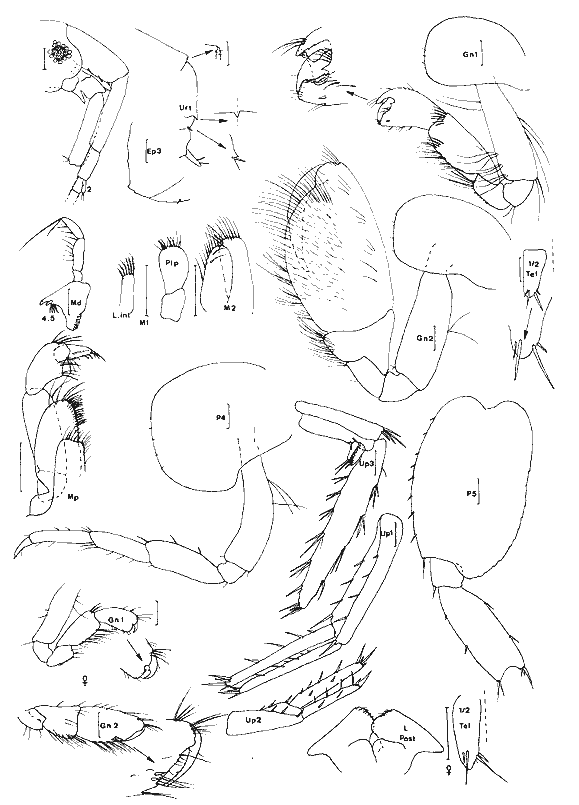 Fig. 12. Melita planalerga Kunkel, 1910: (masculino) de 6 mm et (femenino), St. 17 (réc. Ledoyer). 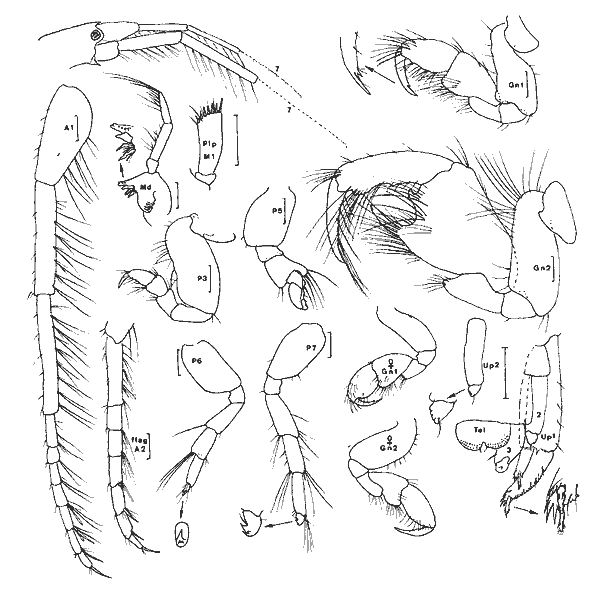 Fig. 13. Cerapus benthophilus Thomas et Heard, 1979: (masculino) de 4.8 mm, Punta Gorda, 03/12/81 (réc. UNAM). Pieces buccales classiques, mais mandibule a palpe relativement grête et à article distal simple, non spatulé et relativement peu pileux; deux ou trois épines molaires. Palpe de la MX1 biarticulé, portant 7 épines distales. MX2 bilobée, sans soie médiolobulaire. Mxp à palpe de 4 articles et purvu d'une griffe teminale. Coxales 1 à 4 plus larges que hautes, contigues. Gnathopode 1 subchéliforme, nettement moins robuste que le second, à carpe et propode subégaux, ce dernier sans bord palmaire bien défini; dactyle long présentant une dent accesoire apicale. Gnathopode 2 puissant, carpochéliforme et très pileux: bord antéro-proximal de l'article basal orné d'une groupe de longues soies; carpe dilaté, à bord antéro-inférieur élargi et armé de deux dents à pointe arrondie, la dent externe, étant la plus développée, est entourée à sa base de longues soies; propode étroit, pileux et présentant une petite saillie prédactylaire finement denticulée; dactyle aussi long que le propode, orné de longues soies et de nombreuses denticuations. P3 à P7 classiques pour le genre: lobe postérieur de l'article méral de la P5 portant 5 longues soies. Pléopodes 1 á 3 biramés, de taille nettement décroissante et à rame interne plus réduite que l´externe. Uropode 1 biramé, rame externe deux fois plus longue que l'interne, denticulée sur le bord externe et armée à lapex de fines dentes atteignant la moitié de l'épine terminale dont l'apex est barbelé; péndoncule orné de 4 petites soies latérales externes et trés finement denticulé á l'apex. Uropode 2 simple: rame courte, portant une petite soie et présentant 3 à 4 crénelures. Uropode 3 uniramé, simple; rame armé d'une petit crochet apical. Telson légèrement fendu, plus large que long, apex des lobes largement arrondi et orné d'une vingtaine de denticules disposées sur deux rangs. (femenino) ovigère d'environ 4 mm, St. 4 (réc. Ledoyer). Elle est identique au (masculino) au niveau des uropodes et du telson. Le flagelle principal des A1 et A2 compte 5 articles. Gnathopode 1 subchéliforme, assez semblable à celui du (masculino) mais àrticle basal moins élargi. Gnathopode 2 subégal au précédent, subchéliforme, à bord palmaire non défini. Dactyle du gnathopode 1 avec une dente accessoire, celui du gnathopode 2 denticulé. Chez de jeunes spécimens j'ai pu observé que le flagelle de l'A1 pouvait compter 4 segments, celui de l'A2 en présentant 5. Matériel: St. 1 (4); 2 (1); 3 (7); 4 (9); 12 (2); 15 (1). Réc. UNAM, Punta Gorda, 03/12/81: (masculino). L'espèce semble préférer les zones les plus déssalées de la Laguna de Términos. Ceci correspond aux observations de Thomas et Heard (1979) décrivant l'espèce de Louisiane. PHOXOCEPHALIDAE
Paraphoxus spinosus Holmes 1905 Fig. 14Bousfield, 1973: 125, pl. 34 (1) Description: (femenino) de 2.6 mm, St. 17 (réc. Ledoyer), Estero Pargo. Rostre large, proéminent, Arrondi en vue dorsale. Oeil bien conformé, brunâtre. Al à flagelle accessoire de 5 articles, le principal en comptant 6, second article du pédoncule portant quelques longues soies sur le bord inférieur. A2 à flagelle de 7 articles; les deux derniers articles du pédoncule sont armés de quelques épines sur le bord dorsal. Mandibule à palpe triarticulé avec l'artcle terminal obliquemet tronqué à l'apex oú sont implantées 7 spies; processus inciseur peu denté; épines molaires petites et peu nombreuses (4); processus molaire réduit à une petite protubérance armée de 3 ou 4 courtes soies spiniformes. MX1 à palpe biarticulé arné de 4 soies á l'apex; lobe intene avec 2 ou 3 courtes soies distales. Lobes des MX2 réduits et peu soyeus (6 à 8 soies à l'apex de chacun des lobes). Mxp classique rais à lobe externe peu développé et bordé de rares (4) longues soies spiniformes. Coxales 1 à 3 plus hautes que larges, arrondies et ornées de quelques longues soies sur le bord inféropostérieur. Gnathopodes 1 et 2 subchéliformes, de même taille et de même morphologie. Bord palmaire oblique, arrondi; angle palmaire armé d'une longue épine. P3 et P4 semblables; carpe portant une longue épine presque aussi longue que le propode armé d'epines. P5 à article basal dilaté, ovalaire; carpe et mérus élargí et épineux. P7 á article basal largement ovalaire; carpe et merus élargi et épineux. P7 à article basal largement ovalaire, très lobè au nive au inféropostérieur; bord postérieur à peine crénelé; bord anterieur praesque glabre. Epimère 3 arrondi, bord postérieur finement festonné et orné de 4 o 5 petites soies. Uropodes birmés; rames des uropodes 1 et 2 peu épineuses et subégales. Uropode 3 à rame trés dissymétriques (femenino): rame externe, la plus développée, biarticulée et ornée de quelques soies sur le bord interne de l'article proximal; rame extene réduite, quasiment glabre si cen n'est une petite soie distale. Telson fendu apex des lobes arrondi et armé d'une coutre épine et d'une petite soie. 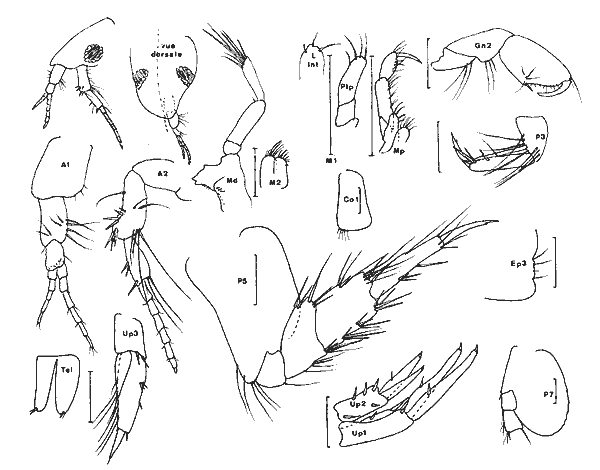 Fig. 14. Paraphoxus spinosus Holmes, 1905: (femenino) de 2.6 mm, St. 17 (réc. Ledoyer). Materiel: Dans nos récoltes cette espèce n'a été récolté que dans le prélèvement réalisé au coucher du soleil. On peut penser que l`espéce est surtout activve la nuit alors que le jour elle reste enfouie dans le sédiment. L'observation rapide de prélèvements réalisés par l'équipe d'écologie côtière de l'UNAM indique que cet animal semble assez largemente distribué dans les herbiers de Thalassia. ConclusionesCONCLUSIONSEn ce qui concerne le peuplement amphipodologique des herbiers de la Lagune de Términos, il ne fait aucun doute qu'il est référable aux peuplements que l'on rencontre dans les divers hebiers littoraux depuis la Caroline du Nord. Dans les herbiers de Zostera marina de Caroline du Nord, Nelson (1980) signale comme espèces communes de l`épifaune:Ampithoe longimana, Cymadusa compota, Elasmopus levis etMelita appendiculata. Fox et Bynum (1975) donnent un inventaire des amphipodes des eaux estauriennes de la Caroline du Nord. Dans cette liste on rencontre beaucoup des espèces récoltées dans la Lagune de Términos et Cymadusa compta etElasmopus levis y sont recensées como des espèeces communes à large distribution dans les euplements végétaux. Dans les peuplements d'Halodule wrightii, sur la côte Est de Floride, Nelson et al. (1982) ont étudié la faune des amphipodes dans cinq stations sur une période de quatre ans. Ils ont rencontré un total de 15 espèces dont Cymadusa comta, Grandidierella bonnieroides, Melita elongata, Corophium ellissi, Ampelisca abdita, Gammarus mucronatus, Corophium lacustre et Ampithoe longinana. Melita appendiculata et Ampelisca vadorum y apparaissent rares. Dans les peuplements d'Halodule, de Syringodium et deThalassia, sur la côte Est de Floride, Stoner (1983) constate que Cymadusa compta reste l'élérnent dominat de l'épifaune avec Ampithoe longimana etMelita elongala. Gammarus mucronatus, Melita appendiculata apparaissent plus occasionnellment. Quant àGitanopsis tortugae, assez bien représenté, il paraît absent dans les Halodule. Sur la côte Ouest de Floride, Stoner (1980) étudie durant 15 mois le peuplement amphipodologique des herbiers deThalassia intertidaux. Ce sont en réalité, comme le signale l'auteur des herbiers mixtes à Thalassia, Syringodium et Halodule (stations F12, E10, E12). L'auteur y trouve 30 espèces (St. F11 exclue). L'élement dominant y est Elasmopus levis puis Lembos sp. A,Rudilemboides naglei, Cymadusa compla, Ampelisca vadorum(cette dernière surtout abondante en F11: peuplement sans végétation). Melita appendiculata, Gitanopsissp. A et Gammarus macromucronatus y apparaissent respectivernent en 12, 13 et 24ème position. Dans le même secteur, Sherindan et Livingston (1983) étudient l'endofaune et l'épifaune des herbiers d'Halodule writhtiioù ils signalent Ampelisca vadorum en tant qu'élérment dominant de l'endofaune alors que Cymadusa compta et Mucro gammarus mucronatus le sont pour l'épifaune. A ces espèces s'ajoutent Cerapussp., Gitanopsis sp Grandidierella bonnieroides et Melita elongata. Ces diverses données montrent qu'il y a une relative homogénéité de la faune des amphipodes depuis la Caroline du Nord jusqu'aux herbiers littoraux de la côte nord cuest du Yucatan. L'uniformité semble presque totale dans le cas des herbiers de la côte du golfé de México. En effet, comme dans le cas des récoltes réalisées dans la Lagune de Términos, sur côte Ouest de la Floride il y a apparemment disparition d'Amphithoe longimana. Ampelisca vadorum paraît se subsistuer à A. abdita quoique Fox et Bynum (1975) ne la signalent pas formellement de Caroline du Nord, et, Corophium louisianum à C.ellisi. En ce qui concerne la côte atlantique de la région de l'Amérique centrale, au sud du Yucatan, il n'y a pas de données à ma connaissance. J'ai, par contre, en ma possession les gammariens récoltés dans les herbiers d'Halodule et deThalassia par R. Vasquez Montoya dans la région de Punta Galeta (Panama). La faune d'amphipodes est peu variée puisque 5 ou 6 espèces y sont inventoriées: 2 Ampeliscadifférentes mais á P7 de type brevicornis (2 individus); 1 Talitridae (1 spécimen (femenino)), 1Elasmopus (2 individus), Melita planaterga (2 individus dans 2 prélèvements distincts) et dans 16 récoltes différentes l'espèce Cymadusa filosa sensu lato. La présence de cette dernière espèce indique que dans ce secteure la faune des herbiers présente un caractère tropical mettement marqué. Comparativement aux autres herbiers dont nous venons de donner un aperçu, ceux de la lagune de Términos paraissent se singulariser uniquement par la faible abondance (peut-être de caractère saiseonnier) enDulichiella ( = Melita)appendiculata. On note aussi la présence assez constante de Melita planaterga et de Gitanopsis laguna qui semblent remplacer les espèces plus nordiques Melita elongata et Gitanopsis tortugae. Toutefois Stoner (1980) signale de la côte Ouest de Floride un Gitanopsis sp. A qui est, peut-être, identique à celui de la lagune de Términos. Dans le cas de la lagune de Términos proprement dite, à coté d'un lot commun d'espèces (Tableau 1), on observe approximativement, quoique de façon moins précise le mème schéma de distribution que dans le cas des Caridea: 1) groupe des herbiers d'Halodule en zone dessalée (Sud de la lagune) avec essentiellement Grandidierella bonnieroides. 2) gróupe des herbiers les plus soumis aux influences marines avec Nasageneia yucatanensis et Elasmopus levis qui est aussi un hôte des autres herbiers deThalassia (groupe 3). LITERATURABARNARD, J.L., Bernice P. Bishop Mus. Bull., Gammaridean Amphipoda (Crustacea) in the collections of Bishop Museum. 1955. 1-46. 215: BARNARD, J.L., Pacif. Natur.Benthic marine Amphipoda of southern California: Families Tironidae to Gammaridae. 1962.75-115, 23 fig.(2): 3 BARNARD, J. L., Pacif. Natur. Marine Amphipoda of Bahia de San Quintin, Baja California. 1964. 55-139, 21 fig., 17 chartd.(3): 4 BARNARD, J.L., Proc. US. Nat. Mus.,Marine Amphipoda in Micronesia.1965. 459-552, 35 fig. (3516):117 BARNARD, J. L. Trans. San Diego Soc. nat. Hist. A bilogical survey of Bahia de los Angeles, gulf of California, Mexico, IV. Benthic Amphipoda (Crustacea). 1969.175-228, 30 fig.(13): 15 BARNARD, J.L., Smith. Contrib. Zool., Sublittoral Gammaridea (Amphipoda) of the Hawaiian Islands. 1970. I-IV; 1-286, 180 fig.34: BARNARD, J.L., The marine fauna of New Zealand: Algae-living littoral Gammaridea (Crustacea, Amphipoda). Dept. Sci. Indus. Res. N.Z, 1972a. 1-26, 109 fig. 210: BARNARD, J.L., Smith. Contrib. Zool.Gammaridean Amphipoda of Australia. Part. I. 1972b. 333 p., 194 fig.103: BARNARD, J.L. et G. KARAMAN, Proc. Biol. Soc. Wash. Classification revisions in gammaridean Amphipoda (Crustacea). Part 2.1982. 167-187, 1 fig. (1):95 BARNARD, K.H., Ann. S. Afr. Mus. Contribution to the crustacean Fauna of South Africa. 5: The Amphipoda. 1916. 105-3025 3 pl.(3): 15 BARNARD, K.H., Discovery Report Amphipoda. 1932.326 p., 174 fig. 1 pl.5: BARNARD, K.H.., Ann. S. Afr. Mus. Contribution to the crustacean Fauna of South Africa, XII: Further additions to the Tanaidacea, Isopoda and Amphipoda, together with keys for the identification of hitherto recorded marine and fresh-water species.1940. 381-543, 35 fig. 32: BERENTS, P.B., Rec. Austral. Mus., The Melitidae of Lizard Island and adjacent reefs, the Great Barrier Reef, Australia (Crustacea, Amphipoda). 1983, 101-143, 29 fig. 35: BOECK, A. Forh. Vedensk. Selsk. Krist., Crustacea Amphipoda borealia et arctica. 1871.83-279. 1870 BOUSHELD, E.L., Shallow-water gammaridean Amphipoda of New England. (Cornel University press) Ithaca and London1973. 312 p., 69 pl. BYNUM K.H. et R.S. Fox New and noteworthy Amphipod crustaceans from Nort0h Carolina,Chesapeake Sci.U.S.A.1977. 133, 18 fig. (1):18 CHEVREUX, E., Bull. Inst. océanogr. Monaco Diagnoses d'Amphipodes nouveaux provenant des campagnes de la "Pricesse-Alice" dans l'Atlantique Nord. 1908.18, 4 fig. 122: CROKER, R.A., Pacif. Sci. A remarkable new Amphipod genus (Crustacea, Gammaridea) from Eniwetok Atoll lagoon.1971. 382-386, 2 fig. (3): 25 DANA, J.D., Amer. Acad. Arts Sci. Conspectus crustaceorum quae in orbis terrarum circumnavigatione Carolo Wikles e classe reipublicae faederatae duce, lexit et descripsit Jacobus D. Dana. Pars III: Amphipoda n° I. Proc 1852. 201-220. 2: DANA, J.D., Crustacea. Part II. US. Expl. Exped., 1853-55.689-1618, 96 pl. 14: FARRELL, D.H., Thesis, Ecology and seasonal abundance of littoral amphipods from Mississipi. M.S. Miss. State Univ., State Ollege, Miss. 1970. FOX, R.S. et, K.H. BYBUM, The Amphipod crustaceans of North Carolina estuarine waters. Chesapeake Sci., 1975.223-237. (4): HAMOND, R., On some Amphipods from the coast of Norfolk.J. mar. biol. Ass. U.K1965.153-160. 45: HASWELL, W. A., On australian Amphipoda. Proc. Linn. Soc. N.S. Walles,1880. 245-279, pl. 7-12. 4: HEARD, R.W., Guide to common tidal marsh invertebrates of the Northeastern gulf of Mexico. Mississipi Alabama Sea Grant Consort. MASGP 79-004:1979. 82 p. 73 fig. HIRAYAMA, A. et T. KIKUCHI, The first record of Melita appendiculata (Say, 1818), (Crustacea: Amphipoda, Gammaridae) from japan. Publ. Amakusa Mar. Biol. Lab., 1979. 67-77, 6 fig. 5(1): HOLMES, S.J., Bull. U. S. Bur. Fish. The Amphipoda of Southern New England. 1905.459-529, pl. 1- 13 (non consulté). 24: KARAMAN, G.S., Poljoprivreda i Sumarstvo,Revision of the genus Gitanopsis Sars 1895 with description of new genera Afrogitanopsis and Rostrogitanopsis n. gen. (fam. Amphilochidae) 1980.43-69, 4 fig. 26: KARAMAN, G.S., Poljoprivreda i Sumarsivo, Redrescription of Melita planaterga Kunkel, 1910 from Bermuda islands with revision of genera Melita Leach and Abludomelita n. gen. 1981.29-50, 7 fig.(1): 27 KARAMAN, G.S., Mem. Inst. ocianogr. Family Gammaridae (245-364) in The Amphipoda of the Mediterranean, Ruffo Ed. Monaco,1982. 364 p., 243 fig. 13: KARAMAN, G.S. et J.L. BARNARD, Classificatory revisions in gammaridean Amphipoda (Crustacea), part I. Proc. Biol. Soc. Wash.,1979. 106-165.(1): 92 KRAPP-SCHICKEL, G. et A.A.MYERS Boll. Mus. Civ. St. Nal. The mediterranean species of Gammaropsis Liljeborg (Crustacea, Amphipoda). Verona1979. 441-467, 15 fig. 6: KUNKEL, B.W., Trans. Connect. Acad. Arts Sci. The Amphipoda of Bermuda. 1910. 1-116, 43 fig.16: KUNKEL, B. W., Connect. Geol. Nal. Hist. Surv. The Arthrostaca of Connecticut. 1918.15-181, 55 fig. (non consulté).(26) 6, Amphipoda: LEDOYER, M.., Bull. Mauritius Inst. Amphipodes gammariens (Crustacea) des biotopes cavitaires organogènes récifaux de l'île Maurice (0céan Indien). 1978. 197-332, 43 fig. (3):8 LEDOYER, M., Mem. Mus. Civ. St. Nat. Verona, Les Gammariens de la pente externe du Grand Récif de Tuléar (Madagascar) (Crustacea, Amphipoda). sér. 2. Sez. Sci. della Vita n° 2:1979. 1-150, 91 fig. LEDOYER, M. Faune de Madagascar. Crustacés Amphipodes Gammariens. Pars 1. Families des Acanthonotommatidae a Gammaridae. Editions, du C.N.R.S. Paris.1982. 598 p, 226 fig. (1).59 LEDOYER, M., Mém. Mus. natn. Hist. nat Les Gammariens (Crustacea, Amphipoda) des herbiers de phanérogames marines de Nouvelle Calédonie (Région de Nouméa). Sér. A, Zool. 1984. 113 p., 48 fig. 129: LEWIS, F.G. et A.W. STONER, Bull. Mar. Sci.,Distribution of macrofauna within sea grass beds: an explanation for petterns of abundance. 1983. 296-304.(2): 32 MCKINNEY, L.D., Gulf. Res. Rep.,Amphilochidae (Crustacea: Amphipoda) from the Western gulf of Mexico and Carribean Sea.1978. 137-143, 4 fig. 6: MCKINNEY, L.D., Proc. Biol. Soc. Wash.Four new unusual Amphipods from the gulf of Mexico and Carribean Sea. 1980.83-103, 9 fig.(1): 93 MLLS, E.L., Can. J. Zool. A new species of Ampelisca (Crustacea, Amphipoda) from eastern North America, with notes on other species of the genus.1963. 971-989, 5 fig. 41: MILLS, E.L., Postilla Yale Peabody Mus. nat. Hist.,Noteworthy Amphipoda (Crustacea) in the collection of the Yale Peabody Museum. 1964. 1-41, 6 fig. 79: MYERS, A.A. Bull. Mar. Sci. Taxonomic studies on the genus Grandidierella Coutière (Crustacea: Amphipoda) with descript ion of G. dentimera sp. nov. 1970. 135-147, 4 fig. (1):20 MYERS, A.A., Boll. Mus. Civ. St. Nat. Verona, Studies on the genus Lembos Bate. VI. Atlantic species: L. dentischium sp. nov., L. kunkelae sp. nov., L. rectangulatus sp. nov: L. unicornis Byrnum & Fox. 1977. 125-154, 20 fig. 4: MYERS, A.A., Amphipod Crustacea. I. Family Aoridae. Mem. Hourglass Cruises, 1981. 75 pp., 34 fig., 1 pl.(5): 5 MYERS, A.A., Mém. Inst. Ocianogr. MonacoFamily Corophiidae (185-208) in The Amphipoda of the Mediterranean, Ruffo Ed. 1982. 364 pp., 243 fig. 13: NAYAR, K., Bull. Madras Govt. Mus. natn. Sci. nat. Hist.The Amphipoda of the Madras coast. 1959. 1-56. 16 fig. 6: NELSON, W. G.,Crustaceana, The biology of eelgrass (Zostera marina L.) Amphipods. 1980. 59-89. (1): 39 NELSON, W.G., Proc. Biol. Soc. Wash., A new species of the marine Amphipod genus Gammaropsis from the South eastern United Stated (Photidae).1981.1223-1229, 3 fig.(4): 93 NELSON, W.G., CAIRNS, K.D. et R.W VIRSTEIN, Bull. Mar. Sci.Seasonality and spatial paperns of seagrass- associated Amphipods of the Indian River lagoon.1982.121-129.(1): 32 ORTIZ, M., Invest. Mar. Cienc. Serv.Invertebrados marinos bentonicos de Cuba. 1. Crustacea, Amphipoda, Gammaridea. 1978. 3-10.(38): 8 PEARSE, A.S., Proc. U.S. Nat. Mus. Notes on certain Amphipods from the gulf of Mexico, with descriptions of new genera and new species. 1913. 369-379, 8 fig.(1936): 43 PEARSE, A.S., Proc. U.S. Nat. Mus. Notes on certain Amphipods from the gulf of Mexico, with descriptions of new genera and new species. 1913. 369-379, 8 fig.(1936): 43 REID, D.M., Allantide Rep. 2 Report on the Amphipoda (Gammaridea and Caprellidae) of the coast of tropical West Africa. Sci. Res. Danish Exp. Coasts tropical West Africa, Copenhagen:1951. 189-291, 50 fig. 1945-1946 SCHELLENBERG, A., Beitr. Kenntn. Meeresfauna Westafrikas III; Crustacea VIII: Amphipoda. Michaelsen. 1925.113-204, 27 fig. SCHELLENBERG, P.F., Handl. Kgl. Svenska Vet. Akad., Litorale Amphipoden des Tropischen Pazifiks. 1938.105 pp., 48 fig. 16: SHERIDAN, P.F., Northeast Gulf. Sci.Three new species of Melita (Crustacea: Amphipoda), with notes on the Amphipod fauna of the Apalachicola estuary of Northwest Florida. 1980.60-73, 12 fig.(2): 3(1979). SHERIDAN, P.F. et R.J. LIVINGSTONE, Estuaries Abundance and seasonality of infauna and epifauna inhabiting a Halodule wrightii meadow in Apalachicola Bay, Florida. 1983.407-419.(4): 6 SHOEMAKER, C., The amphipod Nototropis minikoi on the cast coast of the United States. Proc. Biol. Soc. Wash.1932.199-200. 45: SHOEMAKER, C., Pap. Tortugas Lab., Two new genera and six new species of Amphipoda from Tortugas. 1933.245-256, 8 fig.(15): 28 SHOEMAKER, C., Proc. Biol. Soc. Wash.,The amphipod genus Corophium on the esat coast of America. 1934. 23-32. 47: SHOEMAKER, C., Scient. Surv. Porto Rico and Virgin Isl, New York Acad. Sci. The amphipods of Porto Rico and Virgin Islands. 1935. 229-253, 5 fig.(2): 15 SHOEMAKER, C., Charles-ton Mus. Leaf.,A new Amphipod of the genus Corophium from Florida.1943. 3-6, 1 fig. 18: SHOEMAKER, C., J. Wash. Acad. Sci., Further notes on the amphipod genus Corophium from the east coast of America. 1947. 47-63, 12 fig.(2): 37 SHOEMAKER, C., Smith. Misc. Coll.,The amphipods of the Smithsonian Roebling expedition to Cuba in 1937. 1948.1-15, 3 fig.(3): 110 SIVAPRAKASAM, T.E., J Mar. Biol. Ass. India, Amphipoda from the east coast of India. Part. l Gammaridea. 1968. 82-122, 14 fig.(1):, (66): 8 SIVAPRAKASAM, T.E., J, Mar.biol. Ass. India,Eurystheus togoensis Schellenberg, a new record of Amphipoda from the Madras coast.1970. 283-285, 1 fig. (2):, (68): 10 SMITH, S.I., Report upon the invertebrate animals of Vineyard Sound and adjacent waters, with an account of the physical features of the region. (Crustacea: Amphipoda, in Verrill A.E.: 1873. 1545-1558 (non consulté). SPANDL, H., Zool. Ergebn. 35. Denk. Akad. Wissensch. Wien, Mat. Nat. Die Amphipoden des Roten Meeres. Exped. S.M. Schiff "Pola" in das Rote Meer nordliche und sudliche hälfe 1895-96, 1897-98.1924. 19-73, 23 fig. 99: STEBBING, T.R.R., Das TierreichAmphipoda I: Gammaridea.1906. 806 p., 127 fig. 21: STONER, A.W., Contr. Mar. Sci., Abundance, reproductive seasonality and habitat preferences of Amphipod crustaceans in seagrass meadows of Apalachee bay, Florida.1980. 63-77. 23: STONER, A.W., Mar. Ecol. Prog. Ser. Perception and choice of substratum of epifaunal Amphipods associated with seagrasses. 1983.105-111. 3: STONER, A.W., J. Crust. Biol., Distribution ecology of Amphipods and Tanaidaceans associated with three seagrass species. 1983.505-518.(4): 3 STOUR, V.R., First Ann. Rep. Laguna Mar. Lab. Studies in Laguna Amphipoda.1912. 134-149, 10 fig. THOOMAS, J.D., Contr. Mar. Sci. A survey of gammarid Amphipods of the Barataria Bay, Louisiana region. 1976. 87-100, 1 fig. 20: THOMAS, J. D. et R. W. HEARD, Proc. Biol. Soc. Wash., A new species of Ce ra pus Say, 1817 (Crustacea: Amphipoda) from the northern gulf of Mexico, with notes on its ecology. 1979. 98-105, 4 fig. (1):92 WILLIAMS, A.B. et K.H. BYNUK Ciesaleale Sci. A ten-year study of meroplankton in North Carolina estuaries: Amphipods.1972.251-266.(3): 13
|