|
FAUNE MOBILE DES HERBIERS DE
PHANÉROGAMES MARINES (HALODULE ET THALASSIA) DE LA LAGUNA DE
TÉRMINOS (MEXIQUE, CAMPÉCHE) I. LES CARIDEA (CRUSTACEA DECAPODA)
ET APERCU SUR LA FAUNE GLOBALE
Trabajo recibido el 23 de abril de l985 y
aceptado para su publicación el 9 de septiembre de l985.
MICHEL LEDOYER
Station marine
d'Endoume, rue de la Batterie des Lions, 13007 Marseille.
Francia.
Cuantitativamente los
Caridea representan 60% de la población móvil de los pastos de
fanerógamas marinas de la Laguna de Términos (63%
enHalodule y 57% en Thalassia). En este
estudio estos animales alcanzan una abundancia de 2484 espécimenes para
una superficie cerca de 12 m² . Cualitativemente, Alpheidae
excluídos, 11 especies se encuentran en estos dos biotipos vegetales que
principalmente se presentan, pero en proporciones casi
inversasHippolyte zostericola y tres especies
dePalaemonetes. Estas cuatro especies unicamente viven en
los pastos de Halodule de la parte más interna de la
laguna donde, además, las poblaciones de Hippolyte
zostericola parecen el límite de su extensión (ausencia
de hembras ovigeras). Cualitativemente se nota un enriquecimiento distinto de
los Caridea en la parte de la laguna mas cercana de la influencia
marina.
Palabras claves: Caridea- Pastos
-Halodule - Thalassia -Laguna de Términos,
México.
Quantitativement les Caridea constituent environ 60% de la population
vagile des herbiers de phanérogames de la Lagune de Términos (63%
dans lesHalodule et 57% dans les
Thalassia). Cette population est remarquablement abondante
puisque pour les récoltes réalisées suivant la technique
du fauchage, l'abondance moyenne sur une surface d'environ 12 m² est de
2484 individus. Qualitativement, Alpheidae exclus, 11 espèces ont
été rencontrées dans ces deux peuplements
végeétaux qui présentent essentiellement, mais dans des
proportions quasiment inverses,Hippolyte zostericola et
trois espèces dePalaemonetes. Notons que seules ces
quatre espèces se trouvent dans les herbiers d‘
Halodule du fond de la lagune où de plus les
populations de‘ Hippolyte zostericola apparaissent
à leur limite (absence de femelles ovigères par ailleurs bien
reprèsentées). On observe qualitativement un net enrichissement
de la faune de Caridea vers la zone de la lagune la plus soumise aux influences
marines.
Paroles clef: Caridea - Herbiers
-Halodule -Thalassia -Lagune de Términos,
México.
RESUMEN
Cuantitativamente
los Caridae representan 60% de la población móvil de los pastos
de fanerógamas marinas de la Laguna de Términos (63% en
Halodule y 57% Thalassia). En este
estudio estos animales alcansan una abundancia de 2484 espécimenes para
una superficie cerca de 12 m². Cualitativamente, Alpheidae
excluídos, 11 especies se encuentran en estos dos biotipos vegetales que
principalmente se presentan, pero en proporciones casi inversas
Hippolite zostericola y tres especies de
Palaemonetes. Estas cuatro especies unicamente viven en los
pastos de Halodule de la parte más interna de la
laguna donde, además, las poblaciones deHippolyte
zostericola parecen el límite de su extensión (ausencia
de hembras ovigeras). Cualitativamente se nota un enriquecimiento distinto de
los Caridea en la parte de la lagunamas cercana de la influencia
marina.
Palabras
claves:Cariea-Pastos-Halodule-Thalassia-Laguna de
Términos, México.
Cette étude effectuée dans le cadre des accords de
coopération franco-mexicains et financée conjointement par le
CONACYT (Mexique)*
* Projet: "Mecanismos de Produccion en ecosistemas
lagunares costeros". Clave CONACYT: QCMABFR-001698.
Ce travail est très ponctuel dans le temps puisqu'il a été réalisé dans un délai de quatre jours et couvre diverses stations réparties sur le pourtour de la Laguna de Términos qui a une superficie d'environ 2000 km² et une profondeur moyenne de 3.5 m. Ce bref délai, lié à la grande superficie de la zone concernée, indique clairement, que cette étude ne constitue en fait qu'un coup d'oeil sur la faune mobile de ces types de biotopes. Toutefois, les prélèvements ayant été réalisés à la même époque, l'aspect saisonnier ne peut pas êste évoqué pour expliquer les différences observées entre les divers sites échantillonnés. Il est donc possible de dégager certains caractères originaux pour divers groupes de stations prospectées. Nous avons choisi d'étudier en priorité les Crustacés Caridés car, dans le cas desHalodule ils représentent en abondance de 14 à 79% de la population et dans le cas des Thalassia de 11 à 86 % de la population avec une moyenne de 57%. Ils constituent donc l'élément faunistique dominant du peuplement vagile de ces herbiers. Nous espérons que ce modeste travail permattra aux études en cours sur la faune mobile des herbiers de ce secteur d'aboutir plus rapidement en facilitant la détermination de cet ordre de Crustacés qui paraît constituer la base du peuplement. Signalons que les phénomènes nycthémèraux semblent revétir une grande importance. Nous terminerons en donnant un aperçu sur la faune mobile globale afin de situer l'importance des diverses classes d'animaux constitutifs de cette faune. Le second groupe, en abondance, est représenté par les Crustacés Amphipodes dont l'étude suivra. MÉTHODE D’ECHANTILLONNAGE ET LIEUX DE PRÉLÈVEMENTSLa technique d'échantillonnage est simple et consiste en l'utilisation d'un filet fauchoir (troubleau des entomologistes) d'un diamètre de 20 cm d'ouverture et à maille, de 0.7 mm. L'ouverture du filet, introduite verticalement entre les feuilles du peuplement végétal prospecté et ce jusqu'au niveau du sédiment, est déplacée latéralement sur une distance d’environ 1 m. Cette opération est répetée 60 fois, le récolteur déplaçant le filet á chaque coup successif. Une surface d'environ 12 m² est ainsi prospectée dans un plan moyen au niveau des feuilles, étant entendu que les coups de filets successifs ne repassent pas au méme endroit. Le volume d'eau théoriquement filtré avoisine les 2 m³ (1.88): colonne d'eau suffisante (égale ou supérieure à 20 cm) et verticalité de l`ouverture du filet toujours parfaitement maintenue. 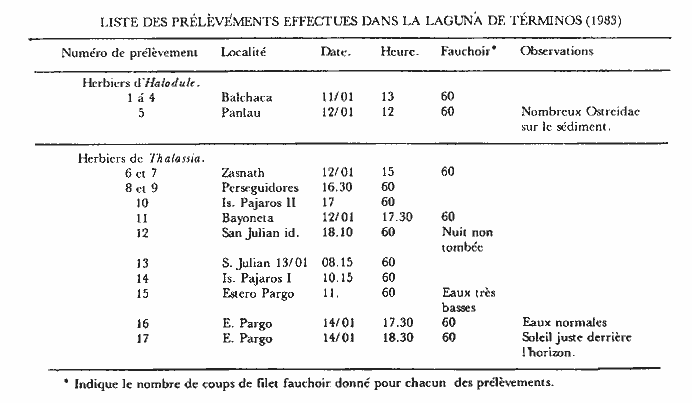 LISTE DES PRÉLÈVEMENTS EFFECTUES DANS LA LAGUNA DE TÉRMINOS (1983) 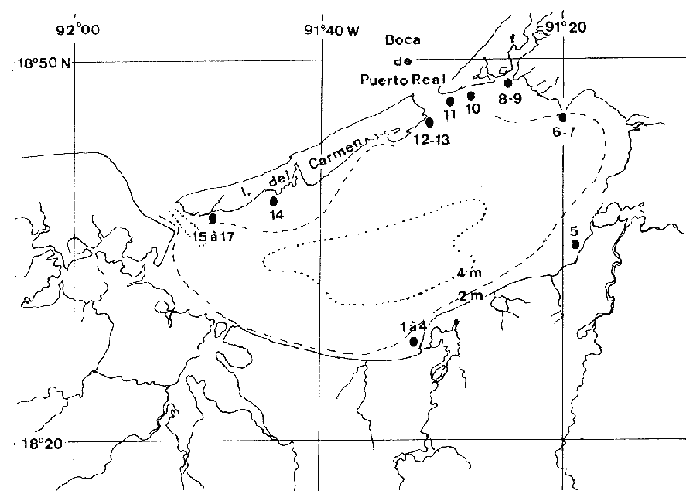 Fig. 1. Localisation des stations prospectées dans la laguna de Términos (janvier 1983). Quant aux stations de prélèvement elles ont été reportées sur la carte ci-jointe (Fig. 1). Elles se répartissent approximativement sur le pourtour de la Lagune de Términos. ÉTUDE SYSTÉMATIQUELa faune des Caridea des herbiers de la Laguna de Términos est constituée d'espèces appartenant essentiellement à deux familles: celle des Hippolytidae et celle des Palaemonidae. Nous ne traiterons pas des Alpheidae qui sont d'ailleurs trés peu représentés (6 individus). A titre indicatif et dans le cas présent, les deux familles qui nous préoccupent peuvent ici se différencier très aisément par l`observation de caractères facilement accessibles. - le carpe de la patte 2 (P2) est respectivement pluriarticulé chez les Hippolytidae et simple chez les Palaemonidae. - le dactyle des pattes locomotrices est respectivement pluridenté et épineux chez les Hippolytidae et simple à bifide chez les Palaemonidae. HIPPOLYTIDAEQuatre genres fréçuentes apparemment les herbiers de phanérogames marines de la laguna de Términos. Ces 4 genres á mandibule dépourvue de palpe peuvent facilement se différencier. 1 - Le carpe de la P2 compte plus de trois articles. Rostre court, inférieur à la longueur de l’écaille antennaire. Epine supraorbitaire vestigiale ou méme inapparente. Mandibule àprocessus inciseur _____________________________________________Thor 1' - Le carpe de la P2 compte trois articles. Rostre aussi long ou plus long que l'écaille antennaire. Epíne supraorbitaire présente ou non. Processus inciseur présent ou non ______________________________________________2 2 - Epine supraorbitaire présente. Bord antérieur du céphalothorax inerme ______________________________________________3 2' - Epine supraorbitaire absente. Bord antérieur du céphalothorax pluridenté. (Mandibule sans processus inciseur) _____ Latreutes 3 - Rostre atteignant l'apex de l'écaille antennaire. Mandibule pourvue d'un processus inciseur _________________Hyppolyte 3 - Rostre beaucoup plus long que l'écaille antennaire. Mandibule sans processus- inciseur _____________________________Tozeuma º (° le caractére épine supraorbitaire n'est valable, dans le genre, que pour l'espéce rencontrée dans les eaux de la laguna de Términos). Genre Hippolyte Leach, 1814Le genre est aisément identifiable par observation de quelques caractéres: épine supraorbitaire présente, carpe des P2 triarticulé et mandibule dépourvue de palpe mais portant un processus inciseur permettent de différencier les espèces de cegenre. Le problème, par contre, devient beaucoup plus ardu en ce qui concerne la détermination spécifique. Chace (1972) revoit les espèces des côtes Nord Ouest Atlantiques qui sont actuellement au nombre de 5. Dans le cas de nos récoltes, il ne fait aucun doute que les spécimens capturés sont référables à l'espèce: Hippolyte zostericola (Smith, 1873) Figure 2Chace, 1972: 118, fig. 49-50 (avec références). Non H. zostericola Williams, 1965: 82, fig. 66. Généralement le rostre porte dorsalement deux épines; très rarement il y en a une ou trois. Le pléonite 5 est dorsalement simple et le bord dorsal du pléonite 3 est nettement saillant par rapport au pléonite 4. Ce caractère est parfaitement figuré par Chace et il m'a paru l'un des caractéres des plus notables et des plus facilement observables pour différencier l'espèce de ses voisines dont les formules rostrales peuvent parfois interférer: H. curacaoensis, H. nicholsoni et surtoutH. pleuracanthus. Chez la (femenino) les pattes 3 à 5, de longueur trés légérement décroissante, sont identiques: le carpe porte une épine 1 atéro proximale. Quant à l'article méral il porte respectivement 4, 2 et 0 épines latérales. Chez le(masculino) la patte 5 n'ést pas essentiellement différente de celle de la (femenino). Par contre les P3 et P4, identiques, ont un propode remarquablement dilaté (Fig. 2). Matériel. L'espèce est constante et remarquablement abondante. Ici nous ne donnerons pas le détail des captures qui sont reportées dans le Tableau 1. Nous indiquerons trois faits qui nous semblént essentiels: Dans la partie Sud de la lagune à herbiers d'Halodule en se trouvé en présence de populations quasiment privées de (femenino) ovigéres. Ces populations sont apparemment juvéniles. Partout ailleurs les populations sont équilibrées et les (femenino) ovigères représentent de 14 à 37% du peuplement. En moyenne, le 1/ 5 dela population est constitué de (femenino) ovigères. Les populations tant dans les Halodule que dans lesThalassia son en général particulièrement abondantes, à l'exception de celles des stations qui encadrent très directement la Boca de Puerto Real. Ce fléchissement peut s'expliquer par les influences marines plus marquées à ce niveau comme le montre la présence d'autres espéces. Mais il est fort possible que ce fléchissement soit aussi en grande partie, du aux conditions météorologiques. En effet, lors de cette période d'échantillonage, la lagune était soumise à un fort vent du Nord qui au niveau de la Boca de Puerto Real conservé toute sa vigueur. Actuellement, il est donc difficile d'indiquer le facteur qui conditionne réellement le fléchissement de population à ce niveau. Dans le premier cas ce fléchissement serait constant et dans le second il serait très temporaire. Genre Latreutes Stimpson, 1860Hippolytidae à mandibule dépourvue de palpe et de processus inciseur. Carpe des P2 triarticulé. 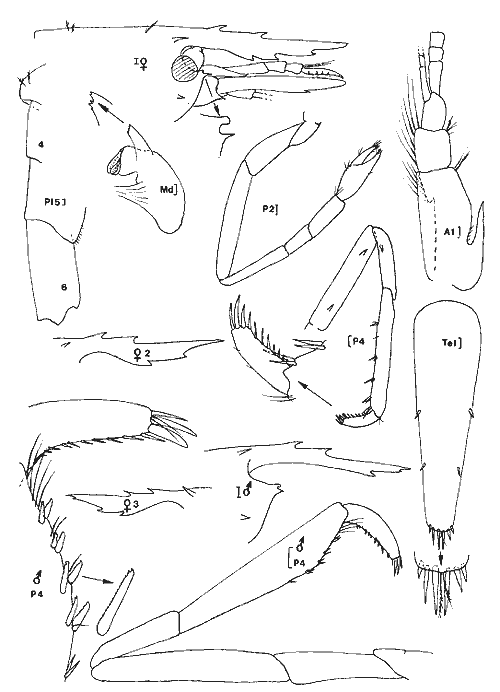 Fig. 2. Hippolyte zostericola,(Smith 1873). (masculino) ovigère de 20 mm et (femenino), St. 10 ( réc. Ledoyer). Longueur rostre et telson exclus. Echelle de 0.1 mm. Maxillipède 3 avec un exopode. Bord antérolatéral de la carapace denticulé. Lame rostrale generalement très dilatée et saillante au niveau postéro-ventral entre la base des antennes. TABLEAU 1 Deux espèces on été capturées dans les herbiers de la Lagune de Términos. 1 Bord dorsal postrostral armé d'une dent. Ecaille antennaire étroite et anguleuse __________________________________L. fucorum 1' Bord dorsal postrostral armé de 5 à 6 dents. Ecaille antennaire spatulée d'une épine disto-latérale _________________L. parvulus Latreutes fucorum (Fabricius 1798) Figure 3Latreutes ensiferus: Stimpson 1860: 27; Bate, 1888: 583, pl, 104 (1); Rathbun, 1902: 114. Latreutes fucorum: Holthuis, 1947: 16; 1951: 131; Siversten et Holthuis, 1956: 31, pl. 1 (1-2); Williams, 1965: 78, fig. 63; Chace, 1972: 121. A l'exception de la dilatation beaucoup plus importante de la lame rostrale de la (femenino) , les deux sexes ne sont pas essentiellement différents. Matériel. St. 12: 4 dont 1 (femenino) ov. ; 8 dont 2 (femenino) ov. Latreutes parvulus (Stimpson, 1866) Figure 4Rhynchocyclus parvulus: Stimpson, 1866: 48; Kingsley, 1878: 56. Concordia gibberosa: Kingsley, 1880: 414, pl. 14 (5). Latreutes parvulus: Holthuis, 1947: 17 et 59; 1951: 131, fig. 28-29 (références); Williams, 1965: 79, fig. 64; Chace, 1972: 124. Très aisément identifiable l'espèce présente un net dimorphisme sexuel qui affecte essentiellement la forme du céphalothorax: chez la (femenino) le bord dorsal du céphalothorax est nettement gibbeux alors qu'il est parfaitement droit chez le (masculino). En arrière de la lame rostrale très élargie, on note chez la (femenino) comme chez (masculino),la présence de 6 épines (parfois 5). Matériel: St. 9: 1; 10: 1 (femenino) ov.; 13: 2 (masculino), 1 (femenino) ov. Genre Thor Kingsley, 1878Une seule espéce du genre (T. floridanus Kingsley) était signalée de l'Atlantique avant le travail de Chace (1972) qui répertorie T. amboinensis (de Man) et décrit deux nouvelles espèces: T. dobkini et T. manningi. Toutes ces espèces, à l'exception de la seconde, sont des hôtes des herbiers. La darnière toutefois présente une répartition plus large. Thor floridanus Kingsley, 1878 Figure 5Kingsley, 1878: 95; 1880: 421, pl. 14 (6); Rathbun, 1902: 116; Holthuis, 1947: 49 (note); Chace, 1972: 136, fig. 58. C'est la seule espèce que nous ayons rencontrée avec certitude dans nos récoltes. Elle apparaît assez polymorphe quant à sa structure rostrale et quant au développement de l'épine supraorbitaire qui, toujours très réduite, apparaît dans quelques cas totalement absente (voir remarque à matériel, St. 7). Cependant, de façon générale, le rostre est bifide à son extrémité (occasionnellement simple à trifide, voir figure 5) et armé dorsalment de 4 à 5 dents (3 et 6 apparaissent les cas extêmes). En ce qui concerne la morphologie de l'article méral de la patte 1, utilisée par Chace pour différencier T. dobkini de T. floridanus et de T. manningi, chez tous les spécimens (masculino) et (femenino) observés avec attention j'ai toujours noté la présence d'une dent latéro-externe proximale. Chez quelques (femenino) (pas chez les (masculino)) j'ai remarqué de plus la présence d'une épine distale bien développée (voir figure 5, (femenino) St. 9). Toutefois, dans ce cas, la pleurite 5 est nettement anguleuse (cf. Chace, 1972: Fig. 58c) et beaucoup moins tronquée que chez T dobkini (cf. Chace, 1972: Fig. 57c) et le dactyle des P 4 et 5 est du type T floridanus à 4 ou 5 denticules latéraux. Le stylocérite, de longueur variable, a un apex qui s'étend de la base de l'épine du second article du pédoncule antennulaire jusqu'à son extrémité distale, ceci suivant les spécimens. Toutefois celui-ci dépasse toujours la cornée (l'oeil étant parfaitement aligné le long du rostre). Chez les spécimens figurés par Chace, le stylocérite atteint juste la partie distale de la cornée. 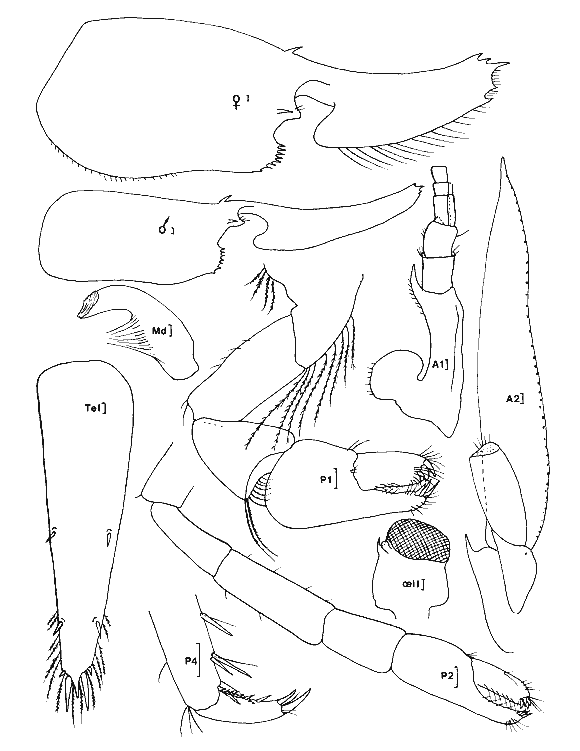 Fig. 3. Latreutes fucorum (Fabricius 1798). (femenino) ovigère de 19 mm et (masculino) St. 12 (réc. Ledoyer). Longueur rostre et telson exclus. Echelle de 0. 1 mm. 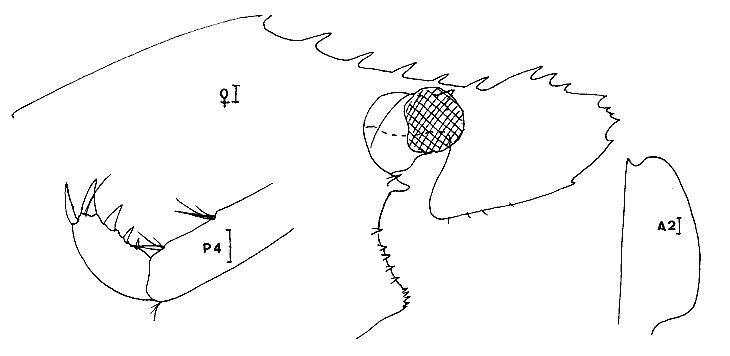 Fig. 4. Latreutes parvulus (Stimpson, 1866). (femenino) ovigère de 13 mm, St. 13 (réc. Ledoyer). En définitive, il est très délicat de séparer T. dobkini de T. floridanus(cf. Chace, 1972: 135, remarks), d'autant plus que cet auteur metThor sp Dobkin, 1968 en synonymie avec T. floridanus et inversement. Enfin, notons que Chace référe à T. floridanus quasiment tous les spécimens récoltés sur la côte Est du Yucatan. Seule la Station 6960 de Bahia de la Ascención présente conjointement les deux espèces. Matériel: St. 7: 1 (femenino) (6/ 2; article méral de la patte 1 inerme distalement; dactyle des P 4 et 5 à 5 denticules latéraux et 2 épines distales); 8: 1 (femenino) ov. (4/ 2; article méral de la P 1 armé distalement); 1 (masculino) (5/ 2; article méral de la P 1 inerme); 9: 2 (femenino) (5/ 2); 1 (femenino) ov. (4/ 2; article méral P 1 armé, cf. figure 5); 10: 1 (femenino) ov. (5/ 2; mérus armé); 1 (femenino) ov. (5/ 2); 1 (masculino) (4/ 2; mérus inerme); 13: 18 spécimens dont 2 (femenino) ov. Animaux non revus en détail; 14: 50 spécimens dont 16 (femenino) ov. 15 individus revérifiés: 5 (masculino), 1 (3/ 2), 2 (4/ 2) et 2 (5/ 2); 8 (femenino) ov., 1 (4/ 1), 6 (4/ 2), 1 (5/ 2); 2 (femenino), 1 (4/ 2), 1 (5/ 2); 15: 18 spécimens dont 3 (femenino) ov., 1 (4/ 3, voir figure 5), 2 (5/ 2); 4 (femenino) (5/ 2); 11 (masculino), 9 (4/ 2), 2 (5/ 2); 16: 3 spécimens, mauvais état, tous (masculino): 2 (4/2), 1 (5/2); 17: 253 spécimens dont 55 (femenino) ov. Animaux non revus en détail. Genre Tozeuma Stimpson, 1860Hippolytidae à mandibule dépourvue de palpe et de processus inciseur. Carpe des P2 triarticulé. Maxillipède 3 sans exopode. Bord antérolatéral de la carapace entier. Généralement il n'y a pas d'épine supraorbitaire, toutefois ce n'est pas le cas pour l'espèce rencontrée dans les herbiers de la Lagune de Términos. Tozeuma carolinense Kingsley, 1878 Figure 6Tozeuma carolinensisKinglsey, 1878: 90; 1880: 413, pl. 14 (8).Tozeuma carolinense: Rathbun, 1902: 114; Ewald, 1969: 510-549, fig. 1-20; Chace, 1972: 141. Espèce aisément identifiable du fait de la structure remarquablement allongée du rostre qui,dorsalement, est inerme. De plus chez cette espèce le bord médiodorsal de l'abdomen est lisse et dépourvu de dents. Remarques: Ce genre compte de espèces à céphalothorax généralement dépourvu d'épine supraorbitaire. Ce caractère est d'ailleurs repris par Chace (1972: 106) dans sa clé. Ceci peut done prêter à confusion. En effet, l'espèce dont il est question ici présente sans conteste une épine supraorbitaire. Kingsley (1880) ne figure pas cette structure, cependant dans sa description originale il parle d'épines suboculaires et branchiostèges: "carapace with sub-ocular and branchiostegal spines; Rostrum narrow. . . a small spine on each side at the base;" 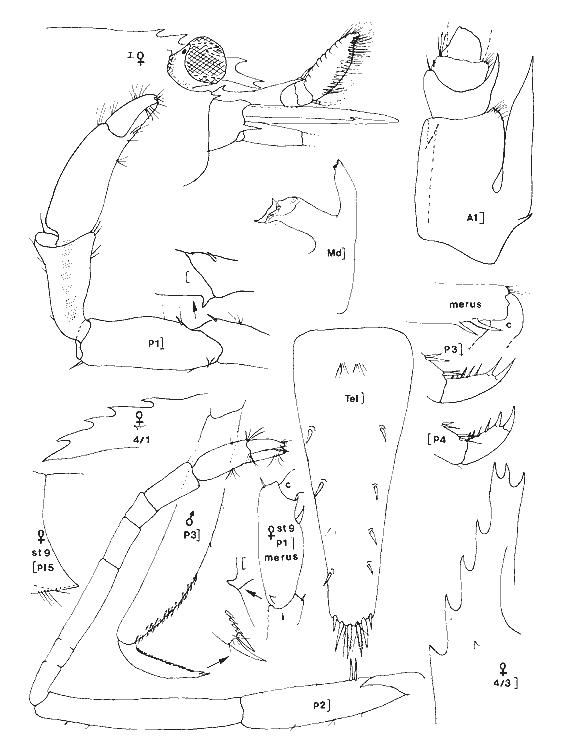 Fig. 5. Thor floridanus Kingsley, 1878. (femenino) ovigère de 17.5 mm, St. 14, (femenino) 4/1 et 4/3, St. 15; (femenino) ovigère St. 9: mérusPl et Pl. 5; (masculino) St. 14: dactyle et propode P3 (réc. Ledoyer). 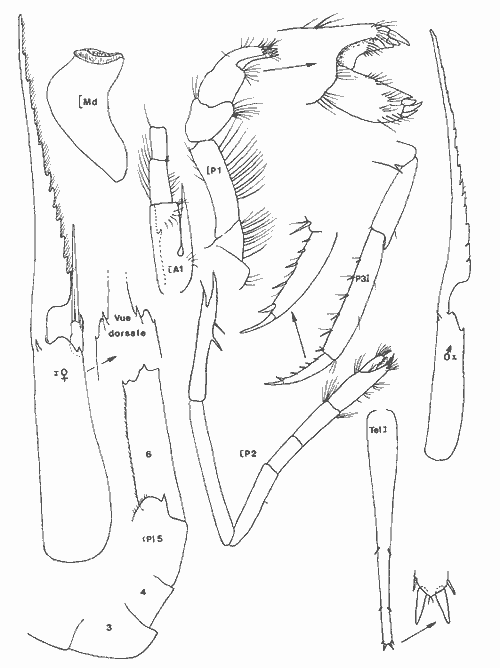 Fig. 6. Tozeuma carolinense Kingsley, 1878. (femenino) ovigère de 47 mm et (masculino) St. 19 -(réc. Ledoyer). Ce caractère est repris mot pour mot par Rathbun (1902) et parfaitement figuré par Ewald. Matériel: St. 9: 5 dont 1 (femenino) ov. PALAEMONIDAEDans les herbiers de la Lagune de Términos nous avons rencontré trois genres appartenant à cette famille, elle même scindée en deux sousfamilles: Palaemoniinae (apex du telson à deux paires d'épines, plus deux longues soies plumeuses) et Pontoniinae (apex du telson à trois paires d'épines). Parmi les Palaemoniinae nous avons identifié deux genres qui se différencient par la présence d'un palpe mandibulaire (Leander) et encore dans le cas de l'espèce de la Lagune de Términos est-il réduit ou par son absence (Palaemonetes). Il est possible de procéder beaucoup plus simplement en observant le propode des pattes 5 qu'il s'agisse de (masculino) ou de (femenino). 1 -Bord apical du propode des P5 orné d'une série de soies disposées en brosse _______________________________________Palaemonetes 1' -Bord apical du propode des P5 orné uniquement de quelques èpines disposèes par paires sèparèes __________________________Leander Parmi les pontoniinae, nous n'avons identifié qu'un genre celui des periclimenes qui se scinde aisément en deux spus-genres. 1 -Dactyle des pattes locomotrices simple _____________________________ P. (Harpilius) 1' -Dactyle des pattes locomotrices bifide _____________________________P. (Periclimenes) Genre Leander Demarest, 1849
Palaemoninae à cornée
colorée et à maxillipèdes pourvus d'exopodes,
caractérisé par son céphalothorax orné d'une
épine antennaire et d'une épine branchiostège et par la
mandibule pourvue d'un palpe.
Le palpe est très réduit dans
le cas de l'espèce dont il est question ici.
Leander paulensis Ortmann, 1897 Figure 7Manning, 1961: 526, fig. 1-2 (avec références). Avec les caractères du genre et comme chez L. tenuicornis l'épine branchiostège est située très nettement en retrait du bord antérieur du céphalothorax (femenino) de 25 mm, non ovée, St. 9. Perseguidores). La partie fusionnée du flagelle interne de l'antennule est inférieure à la partie bifide. Elle se différencie toutefois très aisément de cette espèce par le stylocérite dont le lobe antérieur esttaussi long que l'épine apicale externe qui, ellemême, n'atteint pas l'apex du second article du pédoncule. De plus, le palpe mandibulaire est remarquablement réduit et peut aisément passer inaperçu. Il est biarticulé, l'article distal étant très petit (nous avons observé ce palpe chez divers individus, il ne s'agit pas d´une aberration individuelle). Enfin, le rostre du (masculino) n'est pas essentiellement différent de ceui de la (femenino) et de façon générale, la distance séparant les deux épines postorbitaires est le double de celle qui sépare la première épine postorbitaire de celle qui la précéde. Matériel:St. 8: 1 (femenino) ov., 1 (femenino) ; 9: 1 (masculino), 1 (femenino), 1 juv.; 12: 2 (masculino) , 1 (femenino) ov.; 1 juv.; 13: 2 (masculino) ; 15: 3 (masculino), 1 (femenino) ov. Remarques:Notons que la position de l'épine branchiostège qui est très en retrait du bord anterieur de la carapace, peut inciter à la considérer comme une épine hépatique. Dans ce cas, l'espèce deviendrait référable au genreMacrobrachium. Ceci ne peut pas être envisagé car d'une part, il s'agit de spécimens adultes (présence de (femenino) ovigères) et d'autre part, l'endopode du pléopode 1 du mâle est pourvu d'un appendice interne. De plus, le palpe mandibulaire réduit est biarticulé. Enfin, de même que Manning (1961), j'ai, pendant un certain temps, pensé que ces spécimens étaient des représentants d'une espèce non décrite. 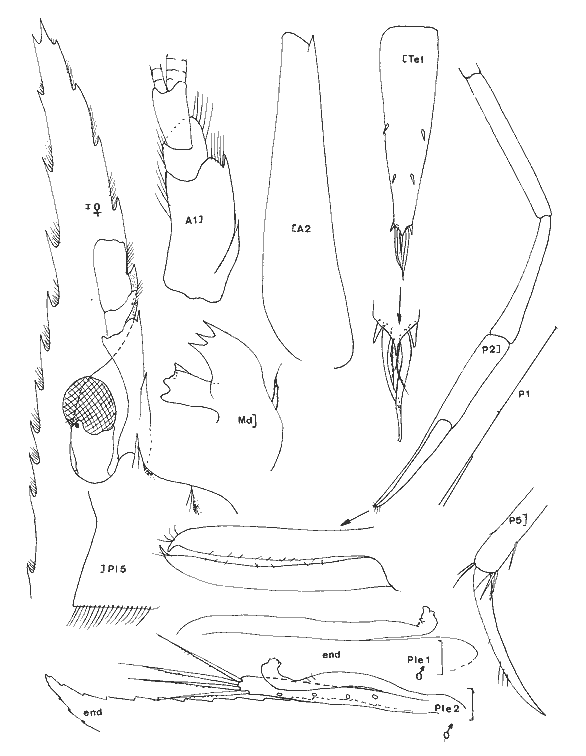 Fig. 7. Leander paulensis Ortmann, 1897.(femenino) et (masculino) (pléopodes 1 et 2), St. 9 (réc. Ledoyer). Genre Palaemonetes Heller, 1869Genre constitué de Palaemininae essentiellement caractérisés par leur mandibule dépourvue de palpe et un céphalothorax armé uniquement d'une épine antennaire et d'une épine branchiostége. Cette dernière se prolonge par un sillon. Chez le mâle le pléopode 1, de plus, est dépourvu d'appendice interne. Holthuis (1952a) fait une étude des espèces américaines qui sont essentiellement dulcaquicoles. Ultérieurement, Chace (1972) décrit une nouvelle espèce du genre des eaux de la Gadeloupe et de Quintana Roo, Mexico. La distinction des espèces du genre fait appel à la longueur de la partie fusionnée du flagelle supérieur de l'antennule relativement à la partie bifide, à la longueur du carpe des P2 comparativement à la pince (propode avec son mors). Dans le cas des spécimens que nous avons recontrés dans la Lagune de Términos, nous avons toujours observés des spécimens reéférables au complexeP. vulgaris (P. vulgarís, P. intermedius etP. pugio) et caractérisés par une partie fusionnée du flagelle superieur de l'antennule plus courte ou subégale à la partie bifide et par un carpe de la P2 plus court ou subégal à la pince. Nous avons éprouvé beaucoup de difficultés à séparer les spécimens de ces populations entre ces trois espèces qui cependant nous ont paru présentes simultanéement. Pour differencier ces trois types dePalaemonetes, nous avons utilisé les critères suivants. 1 - 2 épines postorbitaires. Pleurite 5 spiniforme à arrondi (dernier cas le plus général ). Mors dactylaire de la P2 á 2 dents. Carpe de la P2 nettement plus court que la région palmaire du propode (propode moins son mors)__________________ P. vulgaris 1' - 1 seule épine postorbitaire______________________________2 2 - pleurite 5 nettement anguleuse et spiniforme. Mors dactylaire lisse (P2). Apex du rostre lisse, sans dent provhe de la partie terminale. Souvent 3 épines sous-rostrales. Carpe de la P2 plus long que la région palmaire du propode _______________P. pugio 2' - Pleurite 5 arrondie ou quadrangulaire. Mors dactylaire de la P2 armé d'une dent. Estrémité du rostre souvent pourvue d'une dent proche de l'apex. Carpe de la P2 subégal à la palme___________________________P. intermedius Ces critères résumés sous forme d'une clé dichotomique paraissent satisfaisants. En réalité, le problème de la détermination de ces espèces demeure délicat car il y a apparemment des caractères hybrides de sorte que, pour ma part, j'aboutis beaucoup plus à une approximation qu'à une identification stricte. De plus, je dois signaler que dans le cas des populations importantes de Palaemonetes seul un certain contingent de spécimens a été observé avec attention afin d'aboutir à une estimation du pourcentage respectif des diverses espèces. J'ai enfin noter que, de façon générale, les grandes femelles ovigères appartenaient à l'espèce P. vulgaris quelque soit la composition apparente du reste de la population. L'examen des caractères sexuels du mâle traités par Fleming (1969) n'apparaît pas non plus très satisfaisant quant à la différenciation des espéces. En définitive, nous avons considérer comme référable à P. vulgaris, les individus possédant deux dents nettement postorbitaires. Les autres, à une seule dent postorbítaire, ont été référés à P. pugio lorsqu´ils possédaient une pleurite 5 nettement anguleuse et á P. intermedius dans le cas contraire. Dans ce dernier cas, il faut toutefois signaler que très fréquemment l'apex du rostre de ces individus est du typepugio. Quant aux jeunes spécimens, ils sont, à l'observation de leur pleurite 5, presque exclusivement référables à l'espèce P. intermedius. Finalmente, dans le cas de ces "trois espèces" je ne chercherai pas à les traiter séparement. J'indiquerai de brèves références pour chacune d'elles et je donnerai tout simplement un décompte soit réel, soit estimé de leurs diverses abondances. Palaemonetes intermedius Holthuis, 1949 Figure 8Holthuis, 1952a: 241, pl. 55 (a-f) avec reférences et synonymies. Chace, 1972: 22. 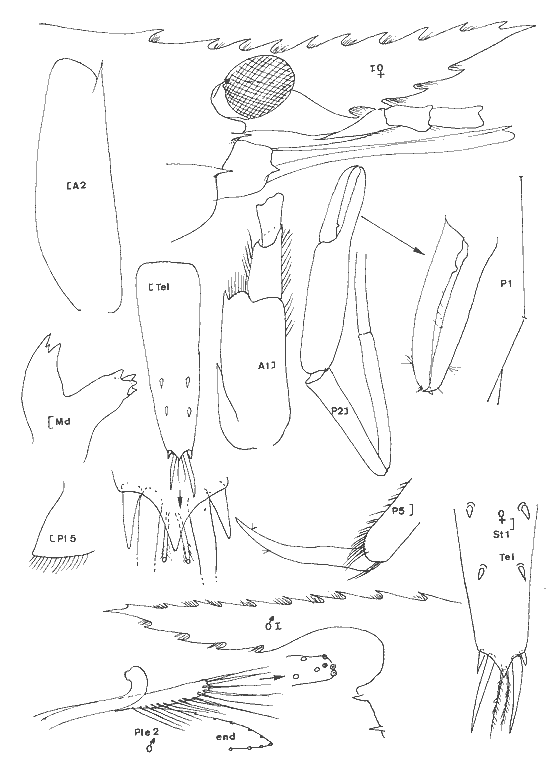 Fig. 8. Palaemoneles intermedius Holthuis, 1949. (femenino) ovigère de 24 mm et (masculino), St. 17 (réc. Ledoyer) et (femenino) St. 1 (Telson). Palaemonetes Pugio, Holthuis, 1949 Figure 9Holthuis, 1952a: 244, pl. 55 (g-l) avec références et synonymies; Heard, 1979: 46, fig. 51 (a-b). Palaemonetes vulgaris (Say, 1818) Figure 10Holthuis, 1952a: 231, pl. 54(f-1); Heard, 1979: fig. 51 (c-d). Matériel: St. 1: population dePalaemonetes estimée à 1519 individus dont 5 (femenino) ovigères. sur 16 individus observés en détail: 4 P. pugio ondt 2 (femenino) ovigères et 12P. intermedius dont 1 (femenino) ovigère; 2: population estimées à 907 individus dont 1 (femenino) ovigère. Sur 25 spécimens observés avec soin tous sont référables à P. intermedius; 3: population de 1450 individus dont 2 (femenino) ovigères. Sur 20 spécimens bien observés, 7 sont référables á P. pugio,les autres à P. intermedius dont les 2 (femenino) ovigères; 4: population de 730 spécimens dont 4 (femenino) ovigéres. Sur 20 individus, 6 P. pugiodont 3 (femenino) ovigères et 14 P. intermedius dont 1 (femenino) ovigére; 5: population de 30 individus, pas de (femenino) ovigère, tous P. pugio; 6: 28 spécimens dont 4 (femenino) ovigères: 11 (dont 3 (femenino)) P. pugio,17 (dont 1 (femenino) ovigère) P. intermedius; 7: 72 individus dont 5 (femenino) ovigères. Sur 20 spécimens observés dont 2 (femenino) ovigères: 4 P. pugio,12 P. intermedius et 1 (masculino), 1 (femenino) et 2 (femenino) ov. P. vulgaris,. 8: 1 (masculino)P. vulgaris avec 1 (femenino) et 1 (femenino) ov. deLeander brevipalpus; 9: 1 (masculino) P. vulgaris avec 1 (masculino) 1 (femenino) et 1 juv. de Leander brevipalpus;10: Pas dePaleamonetes: 11: 7 (masculino), 2 (femenino) ov., 1 (femenino) P. vulgarisavec 1 (masculino) de Periclimenes americans. L'un des (masculino) deP. vulgaris aberrant: 2 dents postorbitaires très nettes mais bord inférieur du rostre à 8 dents: 12 et 13: Pas dePalaemonetes sauf en 13 ou 1 juvénile indéterminé; 14: 29 individus se répartissant en: 1 (masculino), 1 (femenino) P. pugio, 23 P. intermedius et 2 (masculino), 2 (femenino) P. vulgaris. Aucun spécimen ovigère; 15: 6 individus: 1 (femenino) P. pugio, 3 (masculino) P. intermediuset 1 (femenino) et 1 (femenino) ov., de P. vulgaris; 16: 1 juvenile, de P. intermedius; 17: 31 individus dont 6 (femenino) ov. 24P. intermedius et 6 (femenino) ov. et 1 (femenino) postov. deP. vulgaris. Il est possible, d'après ces quelques données, de dégager certains aspects de la répartition de ce groupe d'espèces (Tableau 2). Il est évident que ces conclusions sont très partielles et demanderaient à être étayées par des donnèes saisonnières. Toutefois, pour l'époque concernée, on note d'une part la grande richesse des herbiers d'Halodule de la partie Sud de la lagune en Palaemonetes (P. vulgaris par contre en semble exclu), d'autre part au niveau de la passe Est de la lagune qui constitue la zone la plus directement soumise aux influences marines, cette population de Palaemonetes s'effondre (il y a apparition deLeander paulensis, Periclimenes americanus morphologiquement remarquablement voisins). Enfin, dans la partie Nord Ouest de la lagune, la population de Palaemonetes est réduite, il y a quasiment disparition de P. pugio par contre P. vulgaris apparaît régulièrement présente. La première préfèrerait les zones les plus déssalées contrairement à la seconde. Cependant, nous ne disposons pas de données sur les herbiers d'Halodulequi se trouvent juste au dessus de herbiers de Thalassiadans la zone Nord, c'est à dire le long de la côte Sud de l'île del Carmen et il ne faudrait pas tirer des conclusions trop hâtives. Signalons que cette même remarque est applicable aux Penaeidae (Tableau 2). PONTONIINAE
Periclimenes (Harpilius) americanus (Kingsley, 1878) Figure 11Anchistza americana,Kingsley, 1878: 96. Periclimenes americanus Holthuis, 1951: 60, pl. 18-19 (a-e) et synonymies; 1952b: 10 et 57, fig. 21; Williams, 1965: 43, fig. 36; Chace, 1972: 31. Le rostre est armé dorsalement ((femenino) ovigère de 19 mm) de 9 épines dont l'une est située près de l'apex et une seule ètant postorbitaire. II y a seulement 2 èpines sousrostrales (observées chez tous les spécimens). Le céphalothorax est armé d'une épine antennaire et d'une épine hépathique (Mingsley parle d'épines branchiostège et hépathique). L'angle ptérygostomien est arrondi. Les maxillipèdes sont pourvus d'exopode. La mandibule, sans palpe, posséde un processus inciseur tridenté; le processus molaire est calcifié et triturant. Les pattes 3 à 5 ont un dactyle simple et la partie distale du propode de la patte 5 est dépourvue de rangées de soies (différences avecPalaemonotes). Les pattes 1 et 2 sont chéliformes. 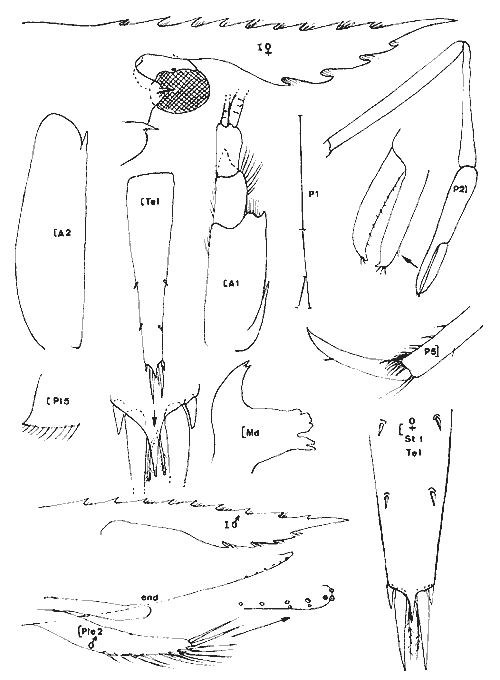 Fig. 9. Palaemonetes Pugio Holthuis, 1949. (femenino) non ovée de 23 mm et (masculino), St. 5 et (masculino) ovigère (Telson), St. 1 (réc. Ledoyer). 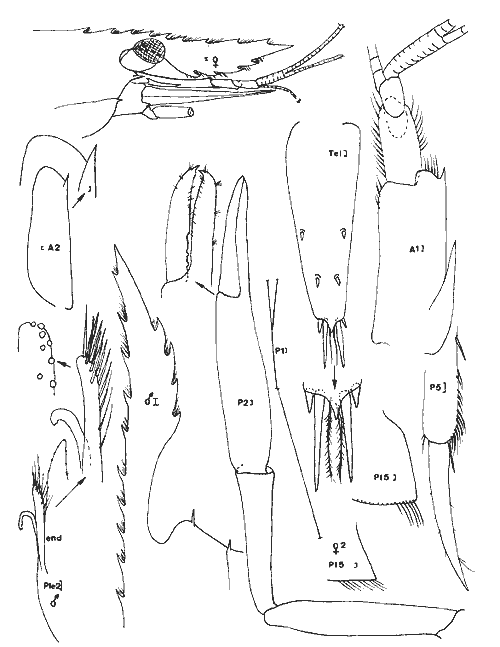 Fig. 10. Palaemonetes vulgaris (Say, 1818). (masculino) Ovigère de 33 mm, (masculino) 2 et (masculino), St. 14 (réc. Ledoyer). 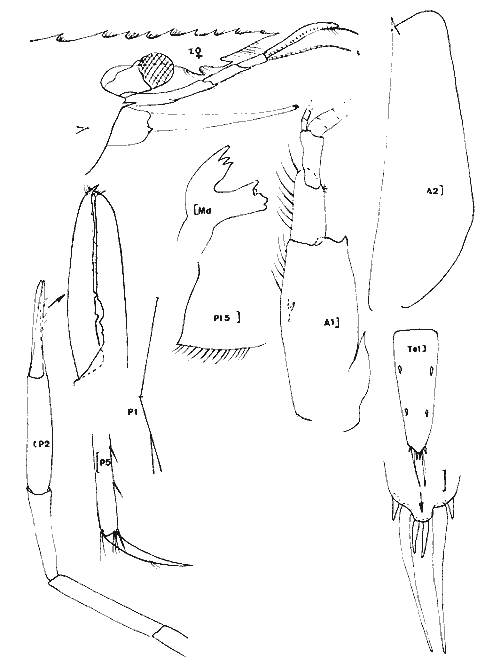 Fig. 11. Periclimenes americanus (Kingsley, 1878). (masculino) Ovigère de 19 mm, St. 10 (réc. Ledoyer). 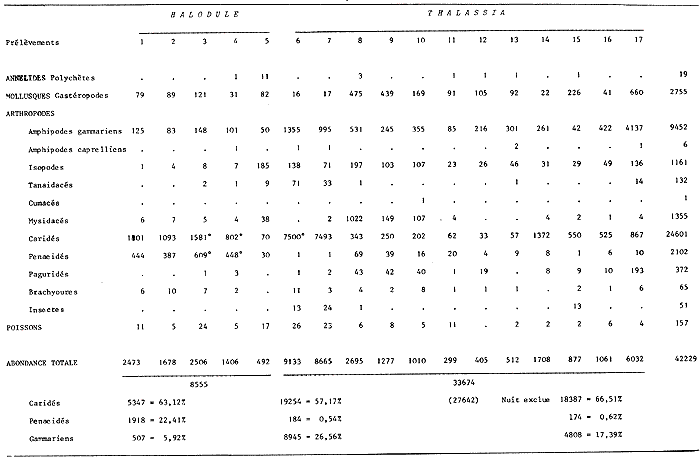 TABLEAU 2 ABONDANCE DES DIVERS GROUPES D'ANIMAUX CONSTITUTIFS DE LA FAUNE VAGILE DES HERBIERS DE LA LAGUNA DE TERMINOS (DU 11 AU 14 JANVIER 1983). Les pattes 2 symétriques sont plus développées que les pattes 1. Ces dernières ont un carpe beaucoup plus long que la pince. Pour les pattes 2 c´est l'inverse. Lemors des pattes 2 représentent environ la moitié de la longueur de la pince. Le mors dactylaire est armé de deux petites dents et le mors propodial est creusé d'une encoche. La pleurite 5 est anguleuse et légèrement dentiforme. Le telson posséde deux paires d'épines médiodorsales, la première paire se situe au 1/3 proximal de la longueur du telson. Ce dernier présente un apex pourvu d'une saillie arrondie et il est armé de 3 paires d'épines la paire médiane étant nettement la plus développée. (masculino): identique à la (femenino) , il présente un pléopode 1 à endopode simple, dépourvu d'appendice interne et un pléopode 2 à endopode portant un appendice interne et un appendice masculina. Cette espèce, dans le contexte écologique observé, peut être confondue dans les abondantes populations dePalaemonetes (plus occasionellement quelquesLeander). Elle s'en différencie par l'épine hépatique et aussi par la grande fusion de la partie basale du flagelle supérieur de l'antennule: en effet ce flagelle n'apparaît bifide que dans sa portion terminale. Or, chez les autres espéces dePalaemonetes et de Leander que nous avons rencontrées dans la Lagune de Términos, la portion bifide du flagelle superieur de l'antennule est plus longue ou subégale à la portion fusionnée. Matériel: St. 10: 1 (masculino), 3 (femenino) ov. 11: 1 (masculino). Periclimenes (Pertclimenes) longicaudatus (Stimpson, 1860) Figure 12Urocaris longicaudata: Stimpson, 1860: 39; Kingsley, 1878: 330; 1880: 424;? Rathbun, 1902: 1916 (voir Holthuis, 1951), Periclimenes longicaudatus: Kemp, 1922: 141; Holthuis, 1951: 26, pl. 6 et 8 (m) (avec références); Williams, 1965: 42, Fig. 35; Chace, 1972: 37. Cette espèce est parfaitement caractérisée par son angle antennaire prolongé en un lobe arrondi dépourvu de toute trace d'épine. Le rostre qui atteint l'extrémité distale du second article du pédoncule antennulaire, porte 8 ou 9 dents dorsales et généralement une petite dent distale sur son bord inférieur; il y en a parfois deux ou elle peut être inexistante. L'article basal du pédoncule antennulaire est armé, sur son bord latéro-distale externe, d'une unique dent. Le dactyle des P 3 à P 5 est nettement biungulé. Les pédoncules oculaires sont grêles et le pléonite 5 est deux fois plus court que le suivant et le pléonite 3, dorsalement est nettement saillant par rapport au quatrième. Matériel: St. 9: 46 dont 30 (femenino) ov. 11: 1. 12: 17 dont 2 (femenino) ov. 13: 17 dont 7 (femenino) ov. L'espèce apparaît localisèe, dans le cas de la Lagune de Términos, dans les herbiers de Thalassia sitúes aux environs de la Boca de Puerto Real où il y a une entrée des eaux marines (cf. Botello, 1978). ConclusionesCONCLUSIONSLes observations qui précédent indiquent clairement qu'il existe une très nette individualité entre les herbiers deThalassia et les herbiers d'Halodule du fond de la lagune de Términos (Tableux 1 et 2). Ceci est d'ailleurs accru si l'on se référe à la faune totale récoltée par fauchage (Tableau 2): les Penaeidae (juvéniles) sont beaucoup plus abondants dans les herbiers d'Halodule de Balchacah. On peut aussi citer, à titre indicatif, que dans le cas des Isopodes dans lesHalodule (Balchacah et Panlau) ce sont les Idotheidae qui tendent à dominer (l/ 1, 3/ 4, 4/ 4, 3/ 7 et 69/ 185 respectivement dans les satations 1 à 5) alors que dans les Thalassia ce sont les Sphaeromidae qui dominent totalement puisque seuls 4, 3 et 1 Idotheidae ont été récoltés respectivement aux stations 6, 7 et 17 (nuit quasiment tombée) ce qui est négligeable comparativement au nombre d'Isopodes recueillis (Tableau 2). Toutefois, si cette démarcation apparaît satisfaisante, il est probable qu'elle correspond à deux cas et qu'elle mérite actuellement une sérieuse restriction. 1) En effet, seules les Halodule de la zone la plus dessalée de la Lagune de Términos (cf. Botello, 1978) ont été observées et dans ce secteur il n'y a pas trace deThalassia. On peut done considérer qu'à ce niveau on est en présence de populations supportant ou adaptèes à de fortes dessalures. 2) Dans la zone Nord Est et Nord de la lagune, seuls les herbiers de Thalassia ont été prospectés étant donné le niveau remarquablement bas des eaux à l'époque de ces récoltes. Mais là, ces herbiers sont le plus souvent bordés vers leur haut niveau par des herbiers d'Halodule dont on ne connait pas précisément la faune mobile. Il est donc possible que dans ce dernier biotope se retrouvent plus abondamment les jeunes Penaeidae, les Idotheidae, les Palaemonetes. Toutefois ceci paraît peu évident étant donné l'asséchement occasionel du biotope sauf peut-être pour les Idotheidae qui très petits sont incapables de se maintenir le long des frondes de Thalassia.Tout cet aspect demanderait toutefois à être confirmé. Cette restriction mise à part, on peut, d'après ces quelques données, scinder l'ensemble de ce peuplement en trois groupes:  Fig. 12. Periclimenes longicaudatus (Stimpson, 1860). (femenino) ovigère de 23 mm, St. 9 (réc. Ledoyer) 1) Groupe des herbiers d'Halodule en zone dessalée (Sud de la lagune) et peut-être des herbiers d'Halodule en général (voir ci-dessus), spécifiquement pauvre, avec fortes populations de Palaemonetes (pugio et intermedius), absence de (femenino) ovigères d'Hippolyte zostericola par ailleurs présentes et, comme nous le verrons ultérieurement abondance de certains gammariens (Gammarus et Grandidierella). 2) Groupe des herbiers de Thalassia de la côte Est de la lagune et de la côte Sud, Sud Ouest de l'île del Carmen à très fortes populations d'Hippolyte zostericola bien équilibraées ( (femenino) ovigères présentes ), présence de Thor floridanus, déclin des Palaemonetes et des gammaridae du genre Gammarus. Cette zone est spécifiquement plus riche que la précédente. 3) Groupe des herbiers de Thalassia de la zone de la Boca de Puerto Real (St.8 à 13) qui est la plus directement soumise aux influences marines par suite du fort courant entrant à ce niveau (Yáñez-Arancibia et Day, 1983). Caractérisé par une richesse spécifique plus élevée (Periclimenes americanus, Periclimenes longioaudatus, Latreutes fucorum, Latreutes parvulus, Tozeuma carolinense). Dans ce groupe d'herbiers, ajoutons que l'on note la présence régulière d'un gammarien du genreElasmopus totalement exclu de la première zone, et que l'abondance totale est relativement basse. Le secteur Perseguidores paraît correspondre à une zone charnière entre le groupe 2 et 3. Ce schéma général correspond d'ailleurs aux observations de Yáñez-Arancibia et Day (1983) au sujet des populations de Foraminifères. Il est certain toutefois que ces trois grands groupes d'herbiers, conditionnés essentiellement, semble-t-il, par des paramètres hydrologiques, correspondent à un même ensemble faunistique caractérisée par le CaridaeHippolyte zostericola et par quelques gammariens comme nous le verrons ultérieurement. Les Palaemonetes(P. vulgaris exclu) sont mieux représentés dans les zones les plus dessalées qu'inversement un gammarien du genreElasmopus ne semble pas fréquenter. N'oublions pas cependant que cet assemblage faunistique ne correspond qu'à un aspect saisonnier (periode de dessalure) et qu'il est peut-être très différent durant la saison opposée. Nous terminerons en signalant que cette faune est classique en ce qui concerne les herbiers de phanérogames marines de côtes Nord Est américaines (Caroline du Nord au Yucatan) et qu'elle apparaît très vicariante de la faune de Caridea des herbiers intertropicaux, tropicaux et méditerranéens de diverses régions avec la coexistence des familles Hippolytidae et Palaemonidae (Palaemoniinae principalement), souvent au genre près. Pource qui est du compartiment faunistique nocturne que constituent les Processidae, l'absence de prélèvements, effectués la nuit étant réellemerit établie, ne permet pas de préciser si ces animaux présents dans la région des Caraibes et du Golfe du Mexique sont effectivement installés dans la Lagune de Términos. Toutefois la très forte poussée de Thor floridanus,observée à la tombée du jour, semble indiquer que si les Processidae habitent ces biotopes quelques spécimens auraient logiquement du être capturés, à moins qu'ils ne se cantonnent que dans la zone de la Boca de Puerto Real. LITERATURABATE, C.S., Rep. Scient. Res. Voyage H.M.S. Challenger, 1873-1876. Report on the Crustacea Macrura collected by the "Challenger" during the years 1873-1876. 1888. XC + 942 p., 76 fig. 157 pl. 24: BOTELLO, A.V., An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Variación de los pararmetros en las épocas de sequía y lluvias (Mayo y Noviembre de 1974) en la Laguna de Términos, Campeche, México; 1978.159-178.(1): 5 CHACE, F.A., Smith. Contrib. Zool. The shrimps of the Smithsonian - Bredin Caribbean Expedition with a summary of the West Indian shallow water species (Crustacea: Decapoda: Natantia). 1972. 179, 61 fig. 98: EWALD, J.J. Crustaceana, suppl. Observations on the biology of Tozeuma carolinense (Decapoda, Hippolytidae) from Florida U.S.A.1969. 18 p. 9 fig. 2: FLEMING, L.E., Proc. biol. Soc. Wash. Use of male external genitalic details as taxonomic characters in some species of Palaemoneles (Decapoda, Palaemonidae). 1969. 443-452, fig. 1-14. 82: HEARD, R.W., Guide to common tidal marsh invertebrates of the Northeastern Gulf of México.Mississippi Alabama Sea Grant Consortium. MASGP-79-004: 1979. 82 p., 73 fig. HOLTHUIS, L.B., Siboga Exped. The Flippolytidae and Rhynchocinetidae collected by the Siboga and Snellius expeditions. With remarks on other species.1947. 100 p. 15 fig. (140): Mono. 39 a 8 HOLTHUIS, L. B., Allan Hancock Found. Publ. Occ. Pap. A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. I. The subfamilies Euryrhynchidae and Pontoniinae. 1951. 332 p. 63 pl. 11: HOLTHUIS, L.B., Allan Hancock Found. Occ. Pap.A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. II. The subfamily Palaemonidae.1952a. 396 p. 55 pl. 12: HOLTHUIS, L.B., Siboga Eped. The Palaemonidae collected by the Siboga and Snellius expeditions. With remarks on other species. 2. Subfamily Pontoniinae. 1952b.253 p. 110 fig. (25),(143). Mono. 39 a 10 KEMP, S., Rec. Ind. Mus., Notes on Crustacea Decapoda in the Indian Museum. XV. Pontoniinae.1922. 113-288, pl. 3-9. (2):24 KINGSLEY, J.S., Proc. Acad. Nat. Sci. Philad., Notes on the North American Caridea in the Museum of the Peabody Academy of Sciences at Salem, Mass. 1878 89-98. (1)1878. KINGSLEY, J.S. Proc. Acad. nat. Sci. Philad., On a collection of Crustacea from Virginia, North Carolina and Florida, with a revision of the genera of Crangonidae and Palemonidae. 1880. 383-427, pl. XIV (Caridea: 411-427).(3): 1979 MANNING, R.B., Bull. Mar. Sci. Gulf Caribbean,A redescription of the Palaemonid shrimp Leander paulensi's Ort mann, based on material from Florida.1961. 525-536, 2 fig. (4):11 RATHBUN, M.J., Bull. U.S. Fish. Comm. The brachyura and Macrura of Porto Rico. 1902.1-127, 24 fig. pl. 1-2 (Caridea: 104-127). (2):20 1900: SIRVERSTEN, E. et L.B. HOLTHUIS, Rep. Scient. Res. "Michael Sars "North Atlant. Deep-Sea Exped. Crstacea Decapoda (the Penaeidae and Stenopida excepted). 1956.1-54. 1910 STIMPSON, W., Proc. Acad. Nat. Sci. Philad.Prodomus descriptionis animalium evertebratorum, quae in Expediti ne ad Oceanum Pacificum Septentrionalem, a Republica Federata Missa, Cadawaladar Ringgold et Johanne Rodgers Ducibus, observavit et descripsif.1860.22-47. 1860: STIMPSON, W., Proc. Chicago Acad. Sci.Descriptions of new genera nad species of Macrurous Crustacea fr the coasts of North America.1866. 46. (non consulté).(1): WILLIAMS, A.B., Fish. Bull. Fish & Wildlife Ser. Marine Decapod Crustaceans of the Carolinas. 1965.298 p. 252 fig.(1): 65 YAÑEZ-ARANCIBIA, A. et J.W. DAY, Jr. Oceanol. Acta. Nº. sp. Ecological characterization of Términos lagoon, a tropical lagoonestuarine system in the southern gulf of Mexico. 1983. 431-440. 5:
|