|
ANÁLISIS DEL CRECIMIENTO DE LA BRÓTOLA PHYUS
BLENNOIDES (BRÜNNICH, 1768) EN EL MEDITERRÁNEO OCCIDENTAL
(PISCES: GADIDAE)
Trabajo recibido el 16 de abril de 1985 y aceptado para
su publicación el 27 de agosto de 1985.
M. GALLARDO-CABELLO
Universidad
Nacional Autónoma de México, Instituto de Ciencias del Mar y
Limnología. Contribución 461 del Instituto de Ciencia del Mar y
Limnología, UNAM.
En el presente trabajo, se
presenta un análisis sobre la metodología empleada por diversos
autores, para determinar las constantes de la ecuación de von
Bertalanffy, y se establece la ecuación del crecimiento que más
se ajusta a las poblaciones de la brótola Phycis
blennoides en el Mediterráneo Occidental. Asimismo, se
determina el crecimiento por sexos, ponderal, compensatorio y la longevidad.
Los parámetros del crecimiento son: L∞ = 574.39 mm; W∞ =
1337.356 gr k = 0. 104769; to = - 0.4637287; A(0.95) = 28 años. Los
parámetros de crecimiento por sexo son: machos L∞ = 366.68 mm; k =
0. 217161 - to = 0. 626438; A(0.95)13 años) hembras L∞ = 677.81
mm; k = 0.0905388; to 1.37965; A(0.95) = 31 años). Los resultados
obtenidos muestran la existencia de un crecimiento compensatorio, en esta
especie, a partir de los tres años de edad.
In this study,
several methods to determine von Bertalanffy's parameters are analysed and the
growth equation forPhycis blennoides (greater forkbeard) in
the Western Mediterranean sea is established. Differences in growth between
sexes, weight growth, growth compensation and longevity were also determined.
The growth parameters were: L∞ - 574.39 mm, W∞ 1337.356 gr, k - 0.
104769, to 0.4637287, A(0.95 161 28 years. The growth parameters between sexes
were: males L∞ - 366.68 mm, k - 0. 219 6 1, to - - 0. 626438, A(O. 95) -
13 years) females (L∞ = 677.81 mm, k - 0.0905388, to = - 1.37965, A(0.95
m 31 years). The data obtained showed that growth compensation occurs in
individuals aged three years or older.
El crecimiento de los organismos ha sido y es en la actualidad- uno de los temas de investigación más importantes e interesantes que han ocupado gran parte del quehacer de numerosos científicos en distintas disciplinas de las Ciencias Biológicas Durante el desarrollo de las investigaciones sobre este fenómeno, se ha logrado conocer bastante acerca de detalles sobre los procesos biológicos del crecimiento orgánico, tales como: los mecanismos de la nutrición, la influencia de las vitaminas y de las hormonas durante el proceso del crecimiento, entre otros, sin embargo, pocos científicos han estudiado el crecimiento a partir de principios y leyes cuantitativas. Brody (1927) observó en sus estudios sobre el crecimiento en animales domésticos, que al elaborar un gráfico de edadlongitud obtenía una curva del tipo S, es decir, que presentaba una parte creciente que puede aplicarse al crecimiento en las primeras etapas de vida de un organismo un punto de inflexión y otra parte decreciente, que corresponde a los estadios finales del crecimiento. Para esta última etapa, Fabens (1965) establece que cuando la edad se incrementa indefinidamente, la longitud tiende a ser asintótica (Loo). Von Bertalanffy (1938) llegó a la misma expresión que Brody, estableciendo que un organismo es análogo a un sistema de reacciones químicas que obedecen a la ley de acción de masas. Por lo tanto, consideró los procesos fisiológicos de los cuales depende la biomasa de un organismo en dos procesos: uno de síntesis de materia (anabolismo) y otro de destrucción de la misma (catabolismo). Asimismo, estableció que dichos procesos son proporcionales a la superficie de absorción de nutrientes, mientras que el catabolismo podría considerarse como proporcional a la biomasa total que es destruida por unidad de tiempo. De acuerdo a lo anterior, formulé la siguiente ecuación para el crecimiento en longitud: Lt = L∞ (1 -e-k(t - to)) que en términos del peso se escribe de la siguiente manera: Wt = W∞(1 -e-k(t - to))³ (Este método teórico, y los datos experimentales necesarios, han permitido establecer conclusiones interesantes. Los organismos no son estáticos, sino el resultado de un gran número de eventos, en los que intervienen, la disponibilidad del alimento, la depredación, la competencia, el almacenamiento de substancias de reserva, la madurez gonádica, las migraciones, entre otros. Asimismo, desde el punto de vista de la fisiología comparada, pueden establecerse distintos tipos de crecimiento animal y derivarse matemáticamente a partir de las constantes metabólicas . Siendo posible calcular y verificar experimentalmente la renovación proteica a partir de las curvas de crecimiento animal establecidas por von Bertalanffy. En el presente estudio, se realiza un análisis mínucioso de los métodos empleados por diversos autores para obtener las constantes de la ecuación de von Bertalanffy (1938) y se establece la ecuación que mejor se ajusta, al crecimiento de las poblaciones de Phicisblennoides en el Mediterráneo Occidental. Asimismo, se aportan datos sobre el crecimiento ponderal, por sexos, compensatorio y longevidad de las poblaciones en estudio. MATERIALES Y MÉTODOSLos ejemplares se obtuvieron de los muestreos biológicos y de la captura comercial realizados frente a las costas de Cataluña, entre Blanes y Vilanova i la Geltru, de septiembre de 1976 a septiembre de 1978. El análisis de las frecuencias de tallas de 5397 ejemplares, por medio de la aplicación de los métodos de Petersen (1892) Cassie (1950, 1954, 1963) y Bhattacharya (1967) permitió la identificación de 5 grupos de edad (Gallardo Cabello, 1986a). El examen de 1132 otolitos ("sagitta" derecha) de Phycisblennoidespertenecientes a ejemplares de 70 a 490 mm de longitud total, permitió establecer ocho grupos de edad. Asimismo, se encontraron diferencias en la relación edad-longitud por sexos (Gallardo-Cabello, 1986b). El método de Bhattacharya fue el que proporcionó los resultados más próximos a los determinados por la lectura de los otolitos (Gallardo-Cabello, 1986a). En base a los resultados obtenidos, se empleó la ecuación de Brody (1927) y von Bertalanffy (1938) para el análisis del crecimiento en longitud. La obtención de los parámetros de la ecuación mencionada, se llevó a cabo por medio de la aplicación de los métodos de Ford (1933) y Walford (1946), Gulland (1964), Tomlinson y Abramson (1961), Allen (1966) y Beverton (1954). Asimismo, se determinó el crecimiento para cada sexo. Para el estudio del crecimiento en peso se determinó el peso entero y desviscerado de 4016 ejemplares. Estos datos se obtuvieron registrando las series de los pesos para cada intervalo de longitudes de 10 en 10 milímetros y posteriormente se calculó el peso promedio de cada una de las clases de tallas. Para la obtención de la relación peso-longitud se aplicó la función exponencial W = a Lb. Empleando los datos de crecimiento en longitud y la relación peso-longitud se obtuvieron los pesos para cada edad. El incremento instantáneo en peso se obtuvo por medio de la aplicación de la ecuación: K = (log e Wt - log e W0) ÷ t El crecimiento teórico en peso se obtuvo substituyendo, en la ecuación de Brody-Bertalanffy, Lt y L∞ por Wt y W∞, obtenidos a partir de la ecuación de la relación peso-longitud. El estudio sobre el crecimiento compensatorio, se llevó a cabo mediante el análisis de la amplitud de los rangos de las clases de tallas obtenidas por los métodos de Cassie y Bhattacharya (Gallardo Cabello, 1986a). La edad límite o longevidad se obtuvo aplicando la ecuación de Taylor (1958, 1959, 1960, 1962). RESULTADOS Y DISCUSIÓN
CRECIMIENTO EN LONGITUD
Método de Ford-WalfordLas longitudes 1t y 1t + 1, se obtuvieron empleando los valores determinados por medio de la lectura de los otolitos, siendo éstas las siguientes:  A continuación se procedió a ajustar la ecuación de la recta de regresión para todos los puntos por el método de mínimos cuadrados, obteniéndose: lt + 1 = 57.9 + 0.89919 lt Teniendo en cuenta que el coeficiente angular K, tiene por logaritmo natural el coeficiente k de la ecuación de Brody Bertalanffy, en nuestro caso resulta que: K = 0.106261 Asimismo, L(sigma) = 574. 64 mm. En la figura 1, se expone la gráfico Ford Walford obtenida para Phycis blennoides, entre un intervalo de tallas de 80 a 340 mm y para los grupos de edad de 0 a 7 años. El método de Ford-Walford, puede presentar serios inconvenientes, si en la obtención de L∞ se emplea el método gráfico. Siendo éste muy sensible a las variaciones de k, pueden obtenerse estimaciones muy diferentes para L∞ Por lo que resulta más práctico obtener el valor de este pará metro, aplicando la ecuación de regresión correspondiente y señalarlo posteriormente en el gráfico. Método de GullandEl valor de to obteniendo mediante la aplicación de este método es el siguiente: to = - 0.39104 La dificultad de este método surge a partir de que los valores de to pueden no estar debidamente representados, tanto en los ejemplares de mayor como de menor edad, debido a un error de muestreo producido por la selectividad del arte empleando. Una pequeña diferencia de lt, daría grandes diferencias en la obtención de la to correspondiente, por ello se recomeinda aplicar la fórmula en los grupos de edad más ampliamente muestrados (Gulland, 1964). 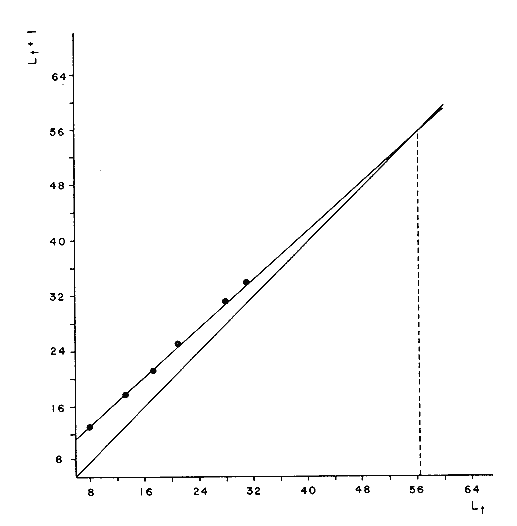 Fig. 1. Gráfica de Ford-Walford. Método de Tomlinson-AbramsonEn las Tablas 1 y 2 se muestra el método empleado. A partir de los resultados obtenidos para diferentes valores de z y por interpolación lineal, se obtiene: z - 0. 9006 Aplicando la ecuación: y = 1÷q · loge z-¹ se tiene que: k = 0.1047 Los valores deΣZ mj, ΣZ ²mj, Σ1j y Σ1j Z mj aparecen en la Tabla 3. Aplicando las ecuaciones: 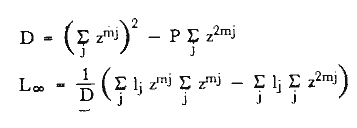 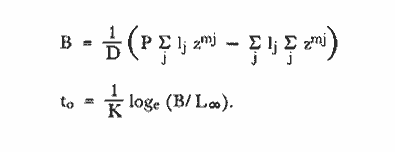 se obtienen los valores de L∞ = 574.39 mm y to -0.4671  TABLA 1 MÉTODO DE TOMLINSON Y ABRANZON. CÁLCULO DE z. 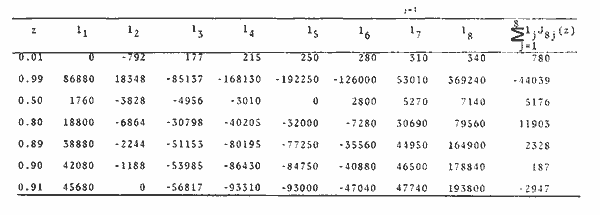 TABLA 2 MÉTODO DE TOMLINSON Y ABRANZON. CÁLCULO DE £ ljJ8j(z) j - 1 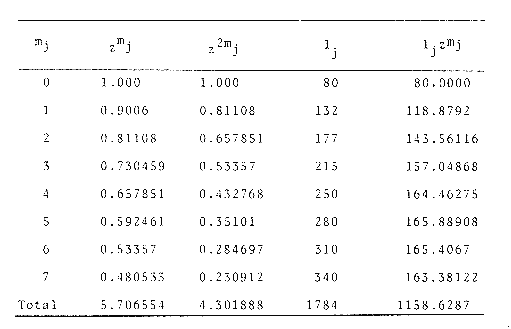 TABLA 3 MÉTODO DE TOMLINSON Y ABRAMSON, RESOLUCIÓN DEL SISTEMA DE ECUACIONES El método de Tomlinson-Abramson, presenta ciertas desventajas al requerir que las muestras estén dispuestas en intervalos regulares de tiempo y que sean del mismo tamaño. Si las muestras no cumplen este requisito, este método puede aplicarse, pero la corrección a realizar requiere de un proceso muy largo, para la cual las tablas que proporcionan Tomlinson-Abramson no son adecuadas. El cálculo de las varianzas de los diferentes parámetros por medio del método de Stevens (1951), puede en algunas ocasiones, no presentar una buena estimación (Tomlinson y Abramson, 1961). Método de AllenTomando como valor de k, el estimado por el método de Tomlinson y Abramson, (k = 0. 1047), se obtienen las siguientes cantidades: A = 8 B = 1784 C = 5.139207 D = 1043.4321 E = 3.489037 F = 5096.4904 G = 20.33454 H = 12.035817 I = 57.490549 J = 16.50434 L = 29723.447 Las expresiones siguientes son: M = 862.0285 P = -820.88848 Q = 1.500896 R = 9.2168743 S = -4495.1038 T = -40.993069 U = 44783.883 V = 4465.8359 α= (1.500896 x -4495.1038) - (820.88848 x 8.2168743) = - 1.545859 DK (α) = (1.500896 x 44783.883) + (8.2168743 x -4495.1038) = (-820.88848 x -40.993069) = 1132.1229 La estimación para la corrección de k, está dada por: -α÷DK (α) = -1.545859 ÷ 1132.1229 = 0.0013654 el valor para k2 es el siguiente: k2 = k1 - α ÷ DK (α) = 0.1047 + 0.1047 + 0.0013654 = 0.1060654 Determinando las primeras y segundas cantidades para este valor de k2 = 0. 1060654, se obtienen:
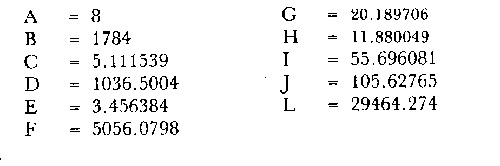 y por lo tanto, 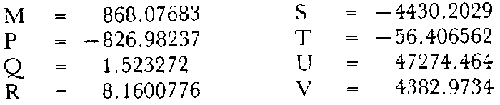 De donde se obtiene: α = (1.5232272 x -4430.2029) - (-826.98237 x 8.160076) = -0.163718. Dk (α) = (1.523272 x 47274.464) +(8.1600776 x -4430.2029) (-826.98237 x -56.406562) = 35394.595. -α ÷ DK (α) = -0.1637718 ÷ 35394.595 = -0.0000046225 siendo el valor de k3: k3 = k2 - α ÷ Dk (α) = 0.1060654 -0.0000046225 k3 = 0.10607 Como la corrección obtenida para k2, da un valor muy pequeño, prácticamente k2 = k3, se procede a estimar to y L∞ to = 6.71778 - 6.77628 ÷ 0.1060654 = -0.457265 L∞ = 868.07683 ÷ 1.523272 = 569.87644 mm El método de Allen, presenta ciertas ventajas con respecto al método empleado por Tomlinson-Abramson (1961), ya que no es necesario mantener constante los intervalos de tiempo y el tamaño de las muestras para cada edad. Sin embargo, el desarrollo del método resulta bastante más largo, debido a la dificultad que representa el elegir arbitrariamente un valor de k, que resulte próximo al valor esperado, sin lo cual, se pueden repetir los cálculos de corrección de este parámetro muchas veces, hasta determinar el valor más ajustado. Por lo que resulta más práctico, desarrollar cualquiera de los métodos anteriores, con el fin de obtener un valor de k, aplicarlo en el método de Allen, y obtener un buen ajuste. Método de BevertonEn la resolución de esta ecuación, se utilizaron los valores de L∞, obtenidos mediante los métodos de Ford-Walford, Tomlinson-Abramson y Allen, en la Tabla 4 se observan los resultados obtenidos. Selección y valoración de los métodos empleadosA partir de las constantes de la ecuación de von Bertalanffy obtenidas por los métodos anteriormente citados, se han obtenido las curvas de crecimiento dePhycis blennoides y aplicado la suma de los cuadros de las diferencias entre los valores. calculados y los observados. Los resultados obtenidos aparecen en la Tabla 5. Los parámetros L∞, y k, obtenidos por medio del método de Ford-Walford, no presentaron una gran diferencia en comparación a los obtenidos por medio de las otras técnicas. Sin embargo, al aplicar estos parámetros en el desarrollo de la curva de von Bertalanffy se obtuvieron valores diferentes a los observados, es decir, no presentó un buen ajuste con respecto a los otros métodos. Lo anterior puede deberse a que en el desarrollo de dicha curva se empleó el valor de to determinado por el método de Gulland. Dicho valor presentó una mayor diferencia de aquellos obtenidos por Tomlinson, Abramson y Allen; y una mayor corrección en el ajuste de la ecuación de Beverton que transformó el valor de to, de - 0. 39104 a -0.4642858, valor éste, más próximo al encontrado por los otros métodos. 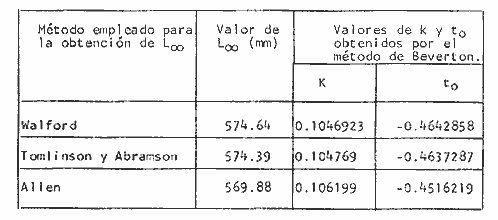 TABLA 4 APLICACIÓN DEL MÉTODO DE BEVERTON PARA LA OBTENCIÓN DE k Y to 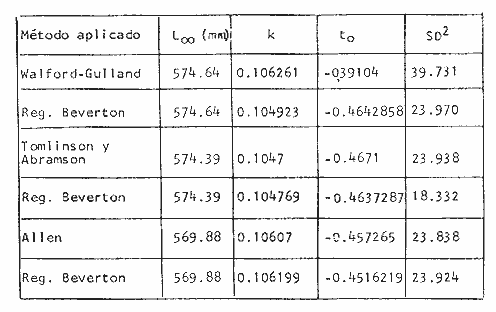 TABLA 5 VALORACIÓN DE LOS MÉTODOS EMPLEADOS Para la obtención del valor de to, Gulland recomienda la aplicación de la fórmula en los grupos de edad que estén mejor representados, con el objeto de evitar errores relacionados con el muestreo, al obtenerse algunos grupos de edad poco representativos de la población en estudio. De acuerdo con lo anterior, se repitieron los cálculos para los grupos de edad "0" a "4" y de "0" a "5" años, obteniéndose como valores de to: -0.43555015 y -0.4232032, respectivamente. No obstante, el desarrollo de las curvas de von Bertalanffy mostró un mejor ajuste de los valores calculados, correspondientes a todos los grupos de edad, cuyos valores de to, se emplearon en la obtención de la to promedio. Es decir, al desarrollar las curvas de crecimiento con los valores de to. obtenidos para los grupos de edad de "0" a "4" y de "0" a "5", se obtuvieron estimaciones de la longitud promedio para las edades 6 y 7 muy diferentes a las observadas. Por lo tanto, el método de Gulland tan sólo proporciona buenos resultados para una parte de la curva de crecimiento de Phycis blennoides. Los métodos de Tomlinson-Abramson y Allen porporcionan mejores valores de to en relación al método de Gulland, lo que disminuye los desajustes que se pueden presentar entre los datos observados y la curva calculada. El avance que presentan los métodos de Tomlinson Abramson y Allen, respecto a los de Ford-Walford y Gulland, radica en el desarrollo de ciertos sistemas que reducen el error cuadrático total entre la curva teórica obtenida y los datos del muestreo. Por otra parte, para la aplicación de los métodos de Tomlinson-Abramson y Allen, las muestras deben de cumplir necesariamente ciertos requisitos para lograr un buen ajuste. Estos métodos tienden a corregir los datos más rigurosamente que las técnicas de Ford-Walford y Gulland al modelo de crecimiento propuesto por von Bertalanffy. En, Phycis blennoides, tanto la técnica de Tomlinson y Abramson, como la de Allen, proporcionaron una buena corrección de los datos, aunque el valor de L∞. obtenido por este último método, presentó mayor desviación respecto a los otros sistemas aplicados. La aplicación de la ecuación de Beverton a los métodos anteriormente citados, proporcionó una mejora de los valores calculados por los métodos de Ford-Walford, Gulland y Tomlinson-Abramson - En el caso del método de Allen, no se observó un mayor ajuste, lo que podría suponer que el valor de L∞, obtenido por este procedimiento, no es del todo correcto en comparación con los otros métodos. Ecuación del crecimiento de Phycis blennoidesLa curva calculada que más se ajusta a los valores observados es la correspondiente a los coeficientes: L∞ = 574.39 mm, k = 0. 104769 y to = -0.4637287 años, calculados por medio del método de Tomlinson y Abramson y ajustados mediante la ecuación de Beverton. Por lo tanto, la ecuación de Brody- Bertalanffy adoptará para la brótola, Phycis blennoides del mar Catalán, la forma siguiente: It = 574.39 (1 - e-0.104769(t + 0.4637287)) Empleando dicha ecuación, se han calculado las longitudes teóricas de la brótola a diferentes edades (Tabla 6). En la figura 2, se expone la curva teórica del crecimiento en longitud, de Phycis blennoides, para las edades 0 a 7 años. Durante las primeras etapas de desarrollo la brótola presenta un crecimiento rápido en longitud. La gran mayoría de los ejemplares de 130 mm de longitud, obtenidos durante los muestreos del mes de julio de 1977, presentan un otolito constituido por el núcleo y las primeras bandas hialina y opaca, perteneciendo, por lo tanto a la clase de edad "1 ". 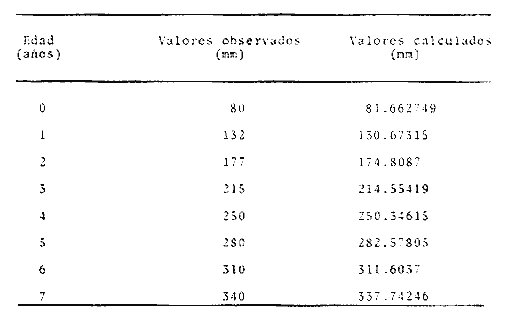 TABLA 6 RELACIÓN EDAD-LONGITUD DE PHYUS BLENNOIDES 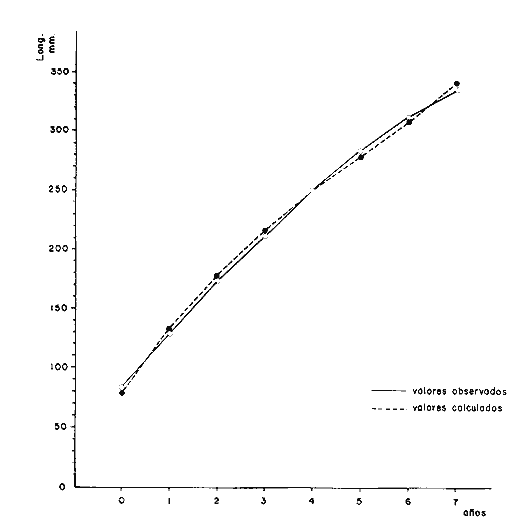 Fig. 2. Curva teórica de crecimiento en longitud de Physcis blennoides. A partir del primer año de vida la velocidad de crecimiento disminuye, incrementándose 45 mm de la longitud total entre el primero y segundo año, 40 mm entre el segundo y tercer año y 36 mm entre el tercero y el cuarto. A partir de este último, el crecimiento es aún más lento con un incremento medio de talla de 30 mm por año. Valores pequeños comparados con los alcanzados durante las primeras etapas del crecimiento (Fig. 3). Para analizar las características del crecimiento de Phycis blennoidesy establecer algunas relaciones con otras poblaciones de peces, se han elaborado las Tablas 7 y 8. En las cuales se hace una exposición de los valores de los parámetros de la ecuación de von Bertalanffy, estimados para varios miembros de la familia Gadidae y diversas especies del Mediterráneo Occidental. Incluyendo los determinados para la brótola en el presente trabajo. En las Tablas 7 y 8, se observa que en la mayoría de los casos, cuanto menor es el valor de L∞, mayor es el coeficiente k, por lo tanto las especies que presentan este tipo de crecimiento alcanzarán más rápidamente los valores de L∞, como ocurre con Sardina pilchardus. 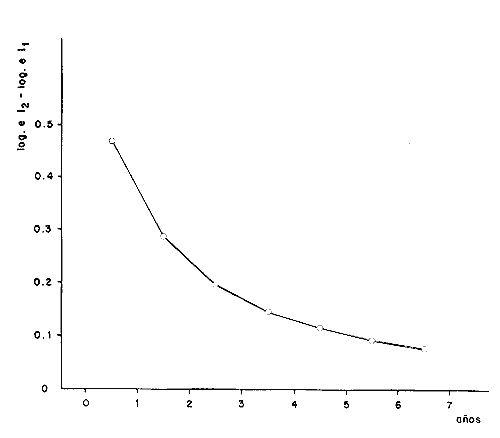 Fig. 3. Rango instantáneo del incremento en longitud. Por lo tanto, los valores elevados de k, producirán curvas de crecimiento muy convexas. Mientras que los bajos valores de k, producirán curvas suaves, en las que el crecimiento es casi constante a través de una serie de años y la longevidad más elevada. Este es el caso dePhycis blennoides. 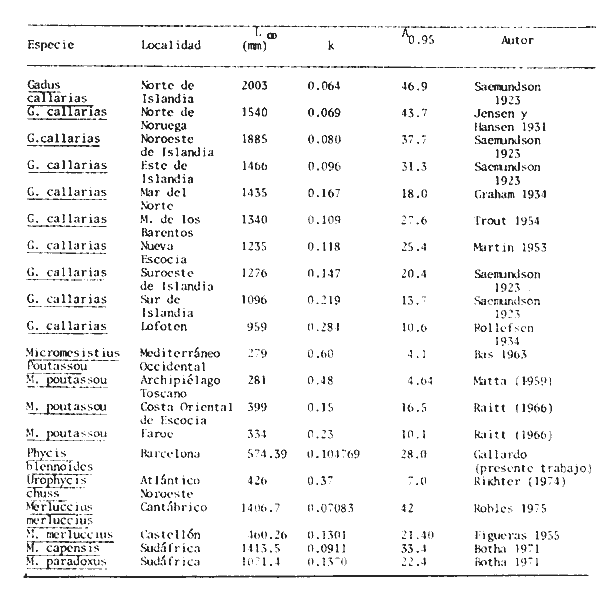 TABLA 7 PARÁMETROS DE CRECIMIENTO PARA DIFERENTES ESPECIES DE LA FAMILIA GADIDAE. (MODIFICADO Y AUMENTADO DE TAYLOR, 1958) 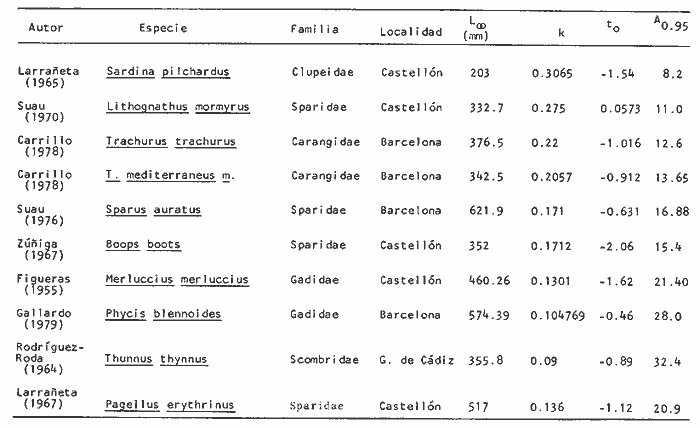 TABLA 8 PARÁMETROS DEL CRECIMIENTO PARA DIFERENTES ESPECIES DEL MEDITERRÁNEO OCCIDENTAL El valor del índice k, determinado para Phycis blennoides, ocupa una posición inferior con respecto a las demás especies del Mediterráneo Occidental (Fig. 8). Una situación muy semejante ocurre con Merluccius merluccius. Con respecto a otros miembros de la familia Gadidae, (Tabla 7) se observa que Phycis blennoidesocupa una posición intermedia en lo que respecta al valor del coeficiente k. Una situación muy semejante ocurre conMerluccius merluccius del área de Castellón yMerluccius capensis del área del Cabo, Sudáfrica. Las diferencias de los parámetros del crecimiento que se señalan para las diversas especies, pueden estar relacionadas con lo expresado por Wheaterly (1966). Este autor establece que las tasas potenciales de crecimiento pueden llegar a estar determinadas genéticamente para las familias o especies. De acuerdo con las condiciones ambientales de un área determinada como son: la temperatura, la disponibilidad del alimento y la intensidad de competencia, entre otras-las tasas potenciales de crecimiento pueden variar dentro de cada especie o población. Dichas variaciones dentro de una familia o especie, pueden observarse en Gaduscallarias en las diferentes áreas del Atlántico Norte. En Merluccius merluccius en las áreas de Castellón y el Cantábrico y entre Phycis blennoides yUrophycis chuss, en el Mediterraneo Occidental y en el Atlántico Noroeste, respectivamente, a pesar de ser especies muy semejantes, presentan valores del coeficiente k muy diferentes. Asimismo, la explotación pesquera puede producir importantes cambios en las tasas de crecimiento de una familia o especie. Una prueba de ello es la disminución de la talla comercial de las especies sometidas a una intensa explotación comercial. CRECIMIENTO SEGÚN EL SEXOEn la determinación de las constantes de la ecuación de Brody-Bertalanffy, para el estudio del crecimiento entre sexos, se aplicaron los métodos de Tomlinson-Abramson y la ecuación de Beverton. Ya que son las técnicas que presentaron un mejor ajuste en la determinación de la curva general de crecimiento de Phycis blennoides. Ecuación del crecimiento en los machos de Phycis blennoidesEmpleando los datos obtenidos por medio de la lectura de otolitos (Tabla 9), se obtuvieron los valores de la ecuación de von Bertalanffy. A continuación se muestran los valores obtenidos así como la suma de las diferencias entre los valores observados y los calculados.  TABLA Los parámetros obtenidos por medio del método de Tomlinson-Abramson fueron los que proporcionaron valores más próximos a los observados, por lo tanto, se propone como ecuación del crecimiento en longitud de los machos de Phycis blennoides, la forma siguiente: 1t = 366.68 (1- e- 0.217161 (t + 0.6264384))  TABLA 9 RELACIÓN EDAD-LONGITUD PARA LOS MACHOS DE PHYCIS BLENNOIDES Las longitudes teóricas a diferentes edades se muestran en la Tabla 9. En la figura 4, se expone la curva teórica del crecimiento en longitud para los machos de Phycis blennoides para las edades de 2 a 6 años. 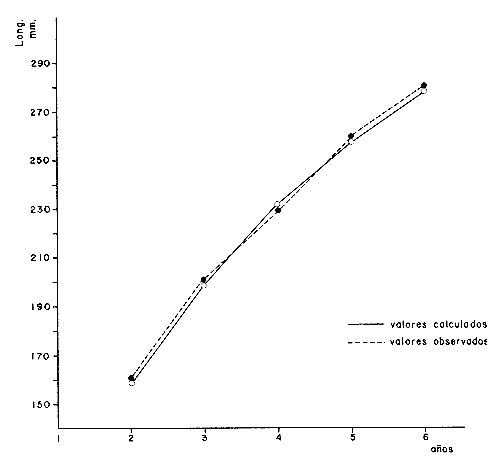 Fig. 4. Curva de crecimiento en machos de Phycis blennoides. Ecuación del crecimiento para las hembras de Phycis blennoidesUtilizando los datos obtenidos por medio de la lectura de los otolitos (Tabla 10) se obtuvieron los parámetros de la ecuación de crecimiento. Asimismo, se desarrollaron las curvas correspondientes y se compararon con los datos observados, por medio de la suma de los cuadrados de las diferencias, los resultados son los siguientes: 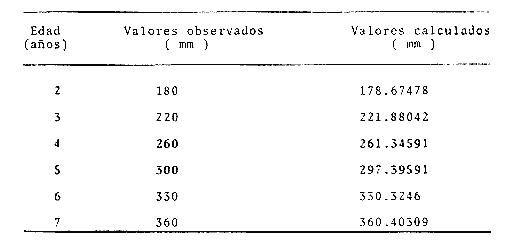 TABLA 10 RELACIÓN EDAD-LONGITUD PARA HEMBRAS DE PHYCIS BLENNOIDES  TABLA La curva calculada que más se ajusta a los valores observados es la correspondiente a los coeficientes: L(sigma) = 667.81 mm, k = 0.905388, to = - 1.3796504, que son los que se toman como definitivos. Por lo tanto, la ecuación de Brody Bertalanffy adoptará para las hembras dePhycis blennoides la forma siguiente: 1t = 667.81 (1 — e -0.0905388 (t + 1.3796504)) Las longitudes teóricas a diferentes edades, aparecen en la Tabla 10. En la figura 5, se observa la curva teórica de crecimiento en longitud para las hembras de Phycis blennoides para las edades de 2 a 7 años. 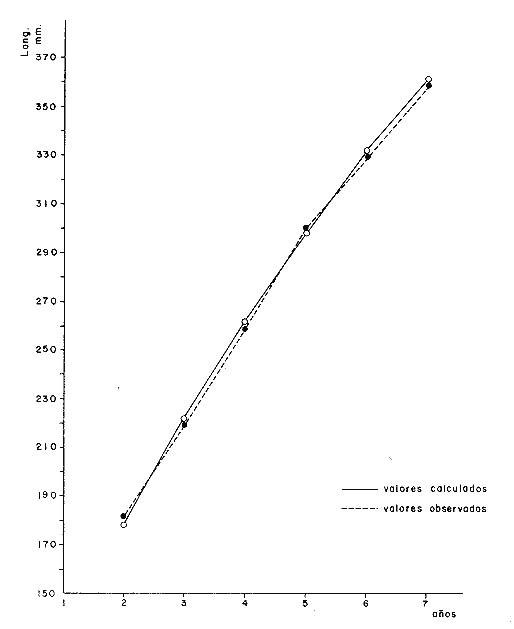 Fig. 5. Curva de crecimiento en hembras de Phycis blennoides. Diferencias del crecimiento entre sexosLas diferencias en los patrones de crecimiento entre sexos se presentan frecuentemente en los miembros de la familia Gadidae. Generalmente, las hembras presentan una tasa de crecimiento mayor que los machos. Hickling (1933) demostró que las hembras de la merluza europea crecen más rápido que los machos desde el tercer año de edad, a partir del cual, esta diferencia se incrementa con la edad. Hickling relacionó este hecho con las etapas de madurez gonádica para cada sexo, estableciendo que los machos maduran sexualmente antes que las hembras, empleando la energía en la formación de los productos sexuales, mientras que las hembras continuan empleando dicha energía en el crecimiento, hasta que alcanzan la edad de madurez sexual. Nelson y Larkins (1970) encontraron que las hembras de la merluza del Pacífico, presentaban un mayor crecimiento que los machos a partir de los cuatro años de edad. Nichy (1969) observó este mismo fenómeno en Merluccius bilinearis. Botha (1971) observó diferencias en el crecimiento de machos y hembras del Merluccius capensis y M. paradoxus, en las áreas del Cabo y Lüderitz (Sudáfrica). Los machos presentaron una tasa de crecimiento menor que las hembras de la misma edad, esta diferencia aumentó conforme se incrementó la edad. Alvariño y Rodríguez (1955) observaron este mismo fenómeno en Gadus minutus. Wheeler (1969) encuentra diferencias entre el crecimiento de hembras y machos en: Gadus merlangus, Trisopterus minutus, Molva molva, Phycis blennoides yMolva dypterygia, en todos los casos la tasa de crecimiento de las hembras es mayor que la de los machos. Los resultados obtenidos para Phycis blennoides se muestran en las Tablas 9 y 10 y en la figura 6. Los parámetros del crecimiento son los siguientes: 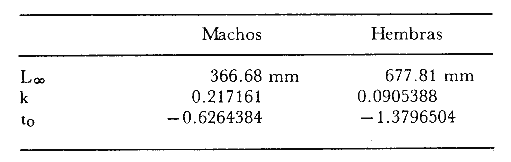 TABLA 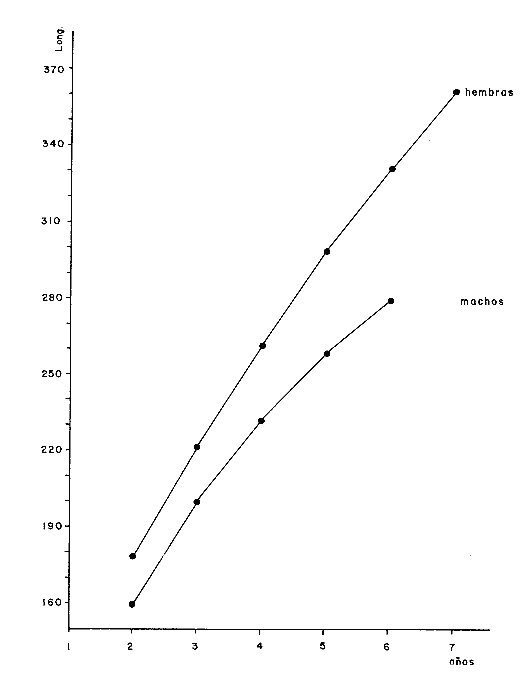 Fig. 6. Diferencias de crecimiento entre sexos. Se disponen de datos para el estudio de la edad y el crecimiento por sexos a partir de los dos años, debido a que es la edad en que las gónadas pueden ser diferenciadas para uno y otro sexo. Las edades máximas que se emplearon para el estudio del crecimiento entre sexos son seis años para los machos y siete años para las hembras. Sin embargo, han aparecido en los muestreos, ejemplares pertenecientes a grupos de edad muy superiores, un macho de ocho a nueve años de 320 mm de longitud total y una hembra de 540 mm, cuya edad correspondería a más de 20 años. Hart (1951) y Wheeler (1969) han encontrado valores semejantes y aún mayores en ejemplares de Phycis blennoides en las costas de Inglaterra. Con respecto a los parámetros del crecimiento entre sexos, se observa que el valor de k es mucho más elevado en los machos que en las hembras, por lo que éstos alcanzarán más rápido una talla cercana a L(sigma), produciéndose una curva de crecimiento más convexa. Mientras que en las hembras se presenta una curva de crecimiento más suave, con una serie de años con una tasa de crecimiento casi constante así como una longevidad mayor. Durante el segundo año de vida de Phycis blennoides, la velocidad de crecimiento es similar en ambos sexos, incrementándose la longitud de los ejemplares 40 mm al tercer año de vida. A partir de esta edad el crecimiento disminuye en los machos, con un incremento de talla media de 30 mm entre los 3 y 4 años, mientras que en las hembras de esa misma edad, la tasa de crecimiento es de 40 mm, valor que se mantiene constante hasta los 5 años. En los machos a partir de los cuatro años el crecimiento es aún más lento, con un incremento promedio de talla de 20 mm cada año. En las hembras a partir del quinto año de vida disminuye la velocidad del crecimiento, aumentando en promedio 30 mm cada año. Como puede observarse, a partir de los 3 años, el crecimiento de los machos es mucho menor que el de las hembras a esa misma edad, esta diferencia aumenta en proporción directa a la edad. Al igual que en las experiencias realizadas por Hickling (1933) para la merluza, es muy posible que en la brótola, debido a que la edad de madurez sexual entre uno y otro sexo es diferente, se presenten diferencias en el crecimiento de machos y hembras. La mayor parte de los machos maduros, colectados en los muestreos pertenecían a los grupos de edad 2 y 3, mientras que las hembras maduras pertenecían a los grupos de edad 3 y 4. Wheeler (1969) observa características muy similares en la velocidad de crecimiento en los machos y hembras de Phycis blennoides, en las costas de Inglaterra. En la Tabla 11, se establece una comparación, de la relación edad-longitud, de la brótola de las costas de Inglaterra y del Mediterráneo Occidental. Como puede observarse, las longitudes para cada edad que alcanzan los machos de Phycis blennoides, en el norte del océano Atlántico, son mayores que las que presenta esta misma especie en el mar Mediterráneo. Sin embargo, la velocidad de crecimiento durante las edades 3 a 5 años es mayor en la brótola del Mediterráneo que en la del Atlántico Norte. A partir del quinto año de edad la tasa de crecimiento es muy similar para esta especie en ambas zonas. Las hembras presentan un patrón de crecimiento muy semejante al señalado por Wheeler (1969). Aunque este autor no proporciona en su trabajo los valores de las constantes de crecimiento de la ecuación de von Bertalanffy para esta especie, establece que las hembras pueden llegar a alcanzar longitudes de 760 mm, valor mayor que el obtenido para L(sigma). = 668 mm en las hembras de Phycis blennoides en el Mediterráneo Occidental. 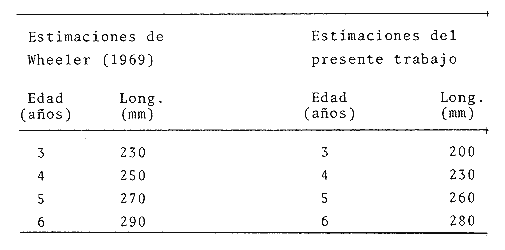 TABLA 11 RELACIÓN EDAD-LONGITUD DE PHYCIS BLENNOIDES (MACHOS) SEGÚN WHEELER (1969) Y GALLARDO (PRESENTE TRABAJO) La talla máxima encontrada para las hembras en los muestreos del Mediterráneo, corresponde a un ejemplar de 540 mm de longitud total, colectado durante el mes de agosto de 1978 en el caladero de la Malica, Blanes, a 500 m de profundidad. Es muy posible que esta especie en el océano Atlántico presente un crecimiento más regular y lento que en el Mediterráneo Occidental. Asimismo, los ejemplares del Atlántico Norte presentan tallas mayores que los del Mediterráneo a una misma edad, así como una mayor longevidad. Todo ello está relacionado con las temperaturas más bajas y una mayor productividad presentes en el océano Atlántico. Por otra parte, la plataforma continental de este último presenta una superficie mucho mayor que la del Mediterráneo, por lo que la disponibilidad y cantidad del alimento y de espacio son mucho mayores. Lo que trae como consecuencia que las poblaciones de peces se desarrollen dentro de un ambiente mucho más estable, alcancen un mayor crecimiento y sean más abundantes. También es importante señalar, que esta especie es menos afectada por la explotación pesquera en el Atlántico Norte que en el Mediterráneo (Wheeler, 1969). Por lo tanto, los ejemplares podrán alcanzar tallas mayores en comparación con los del Mediterráneo, en donde la captura comercial de esta especie está constituida en su mayoría por ejemplares de 2 y 3 años. CRECIMIENTO EN PESO
Relación peso-longitudLas líneas de regresión obtenidas tuvieron las siguientes ecuaciones: W = 0.000004 L 3.094 para los peces enteros, y W = 0.000003 L ³.¹² para los peces desviscerados. En la Tabla 12, aparece la relación entre la talla y el peso para los peces enteros y desviscerados. En las figuras 7 y 8 aparecen las rectas de regresión de las relaciones entre los logaritmos del peso y la longitud. Peso a cada edadEn las Tablas 13 y 14, se muestra el crecimiento en peso absoluto y relativo, observándose que el coeficiente relativo disminuye conforme la edad se incrementa. Crecimiento teórico en PesoLas ecuaciones correspondientes para el peso son:
Wt = 1377.356 (1 — e -0.104769 (t + 0.4637287))³
para el entero y
Wt = 1218.558 (1 — e - 0.104769 (t + 0.4637287))³
para el desviscerado.
A partir de las ecuaciones anteriores se han obtenidos los pesos teóricos, entero y desviscerado de la brótola, Phycis blennoides, a diferentes edades (Tabla 15). En las figuras 9 y 10, se exponen las curvas teóricas del crecimiento en peso, entero y desviscerado de Phycis blennoides, para las edades de 0 a 7 años. CRECIMIENTO COMPENSATORIOEl crecimiento compensatorio ha sido estudiado por Sund (1911), Gilbert (1914), Watkin (1927), Ford (1933), Scott (1949) y Ricker (1969), entre otros. Se define como el fenómeno mediante el cual, los peces, que durante sus primeros años de vida han alcanzado una longitud o peso relativamente bajos, tienden posteriormente a presentar un crecimiento mayor que aquellos peces de su misma edad que crecieron normalmente. De tal manera que la mayoría de los peces tenderán hacia una talla similar y a una reducción en la dispersión de tallas conforme se hacen más viejos. Lo que mostrará una disminución de la desviación standard al incrementarse la edad. 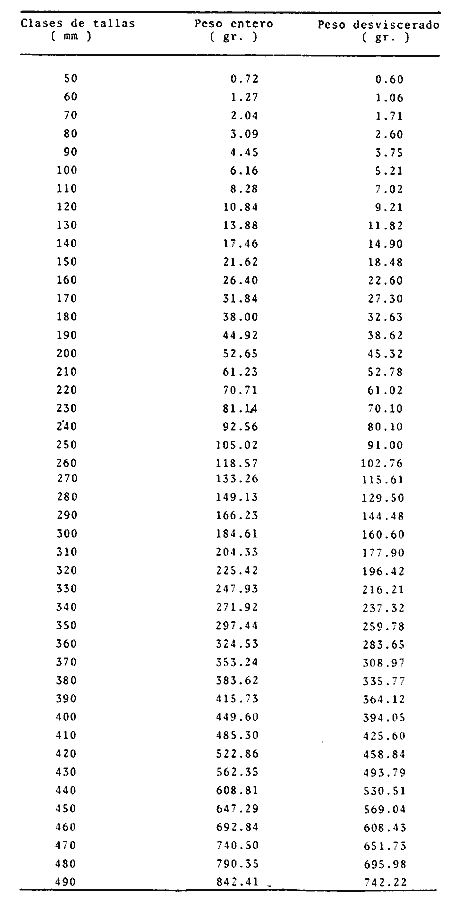 TABLA 12 RELACIÓN PESO ENTERO Y DESVICERADO- LONGITUD PARA PHYCIS BLENNOIDES 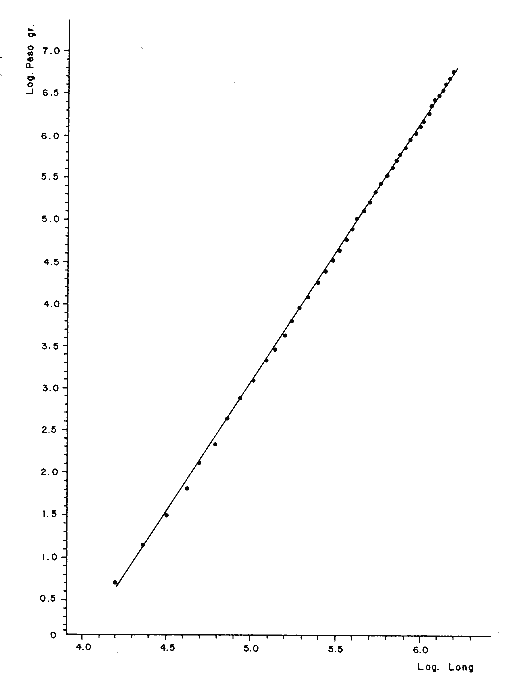 Fig. 7. Relación logaritmo del peso (entero) y de la longitud total del pez. 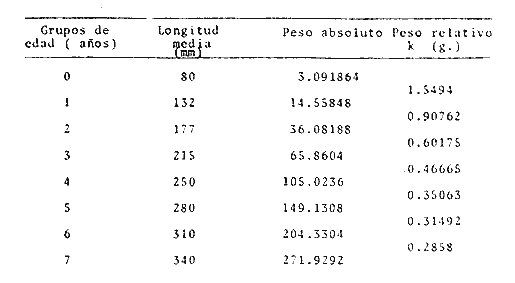 TABLA 13 CRECIMIENTO EN PESO (ENTERO) ABSOLUTO Y RELATIVO DE PHYCIS BLENNOIDES 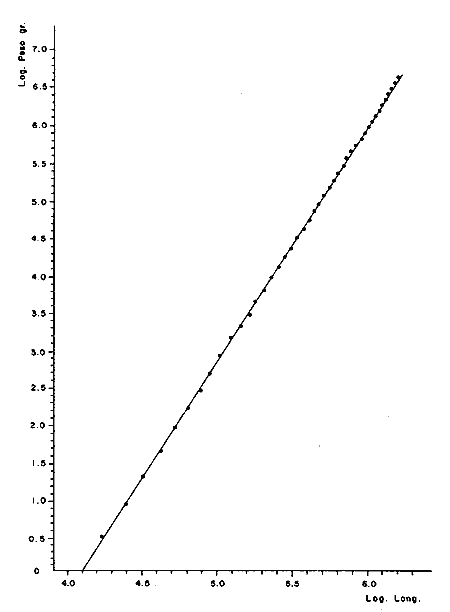 Fig. 8: Relación logaritmo del peso (desviscerado) y de la longitud total de pez. En la Tabla 16, se muestra la amplitud de los rangos de las clases de tallas para los distintos grupos de edad. Aunque los valores obtenidos por el método de Cassie son diferentes a los calculados por la técnica de Bhattacharya, en ambos casos se observa que en las edades 0, 1 y 2 años la variabilidad es mucho mayor que en los demás grupos de edad. Y que a partir de los 3 años de edad ésta se reduce. Aunque luego vuelve a aumentar la desviación standard, las diferencias no son tan marcadas. 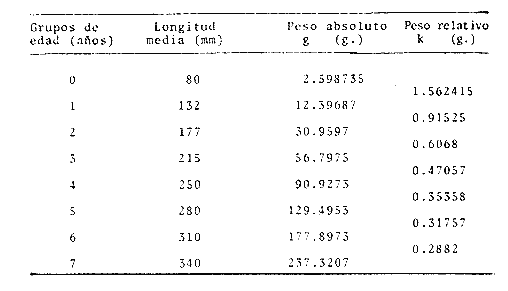 TABLA 14 CRECIMIENTO EN PESO (DESVISCERADO), ABSOLUTO Y RELATIVO DE PHYCIS BLENNOIDES  TABLA 15 RELACIÓN ENTRE LA EDAD (AÑOS) Y EL PESO (GR) SEGÚN LA ECUACIÓN DE VON BERTALANFFY PARA PHYCIS BLENNOIDES Lo anterior puede interpretarse de la siguiente manera: A. El crecimiento compensatorio se presenta a partir del tercer año, edad en que se alcanza el punto de inflexión en la curva de longitud-edad, es decir, que aquellos peces que crecieron poco durante sus primeros años, posteriormente crecieron más intensamente. 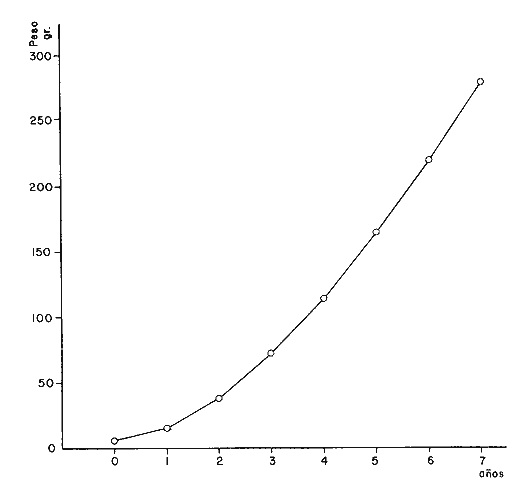 Fig. 9 Curva teórica de crecimiento en peso (entero) de Phycis blennoides. 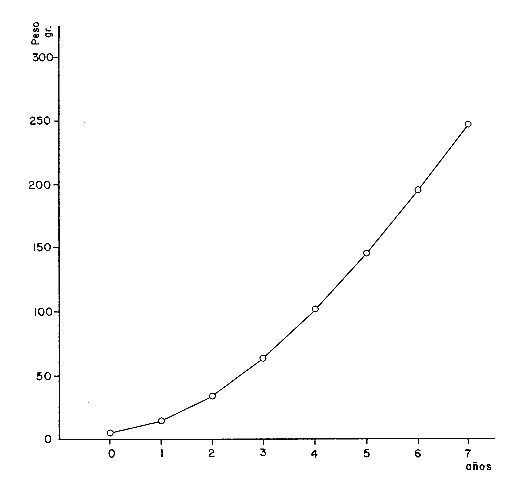 Fig. 10. Curva teórica de crecimiento en peso (desviscerado) de Phycis blennoides.  TABLA 16 AMPLITUD DE LOS RANGOS DE TALLAS EN LOS GRUPOS DE EDAD DE PHYCIS BLENNOIDES También es importante señalar que la disminución de la variabilidad en ejemplares de más de 3 años, puede deberse a que la captura comercial de esta especie es ejercida principalmente en individuos de 2 y 3 años. Por lo tanto, los ejemplares mayores presentarán una variabilidad de tallas más baja debido a una menor abundancia. B. Durante las primeras etapas, edades 0 a 1 (método de Cassie) y 0 a 2 años (método de Bhattacharya) se observa una mayor amplitud en el rango de tallas, tratándose de un crecimiento compensatorio inverso. Este mismo fenómeno ha sido observado por Neave (1954). Esta misma variabilidad en el rango de tallas que va en aumento durante las primeras edades, amplía la dieta alimenticia y el nicho ecológico de la brótola en un medio donde se presenta la escasez de alimento y la competencia intraespecífica puede aumentar. Por lo tanto los individuos de menor talla, ampliarán su nicho ecológico, alimentándose de otro tipo de organismos disponibles en el medio (Gallardo-Cabello y Gual-Frau, 1984). De acuerdo con lo anteriormente expuesto, se puede establecer que la brótola durante sus primeras etapas de desarrollo presenta un crecimiento compensatorio inverso. A la edad de 3 años, en que alcanza el punto de inflexión en la curva de edad-longitud, muestra un crecimiento compensatorio. EDAD LÍMITE O LONGEVIDADLa edad límite de la brótola corresponde a:
A 0.95 = 28 años
La edad límite encontrada para cada sexo es la siguiente:
Machos = 13 años Hembras = 31 años
La edad límite encontrada para Phycis blennoides, 28 años, coincide con los datos expuestos por Nikolsky (1963), quien establece para peces cuya L(sigma) es de 500 a 700 mm, una longevidad de 20 a 30 años. Este valor se aproxima al establecido por Wheeler (1969) para Phycis blennoides en las costas de Inglaterra. Con respecto a la edad límite por sexos, se observa una gran diferencia entre la longevidad que pueden llegar a alcanzar las hembras con respecto a los machos. Un fenómeno muy semejante, se presenta en Merluccius capensis y M. paradoxus, en el área del Cabo, Sudáfrica, en donde las hembras presentan una edad límite que es poco más del doble que la de los machos. Aplicando, la ecuación de Taylor a los datos de Botha (1971), se tiene: ***********ELEMENTO SIN ESTILO ********* tablaEdades semejantes han sido establecidas para Gadus callarias, en algunas áreas del Atlántico Norte, (Tabla 7). Las variaciones de la longevidad en las poblaciones de una misma especie, son el resultado de la adaptación de dichas poblaciones a las condiciones del medio ambiente en que habitan. Nikolsky (1963), establece que aquellos peces que presentan grandes longevidades son grandes predadores bentófagos que muestran un corto periodo de rápido almacenamiento de grasas, fenómeno que se presenta en la brótola así como en la mayoría de los Gádidos. Los peces de mares cálidos presentan un crecimiento inicial más rápido, pero una menor longevidad, que los de la misma especie en mares fríos, lo que quiere decir que tienen un coeficiente k más elevado y una edad límite menor, (Tablas 7 y 8). Esto ha sido confirmado directamente por Taylor (1958) en el bacalao de una serie de localidades del Atlántico Norte, observándose que el logaritmo de la temperatura promedio del agua superficial, muestra una relación directamente proporcional con el coeficiente k, —por lo tanto— una relación inversamente proporcional con respecto a L(sigma) y la longevidad. Como ya se ha mencionádo anteriormente, este fenómeno se presenta en Phycis blennoides, en las áreas del Mediterráneo Occidental y del Atlántico Norte. Siendo menores los valores de L(sigma) en el Mediterráneo que en el Atlántico Norte. Con respecto a las temperaturas que se presentan en el Mediterráneo Occidental, se observa que los fondos en que vive la brótola, 220 a 600 m de profundidad, la temperatura oscila entre 12.75°C a 13.4ºC a través de un ciclo anual, variaciones muy pequeñas como para que puedan representar una variación sobre la curva de crecimiento de estas especies. Por lo que la distribución batimétrica dePhycis blennoides, es uniforme entre los 220 a 600 m de profundidad para los grupos de edad de 2 a 4 años. (Gallardo-Cabello, 1979). Los niveles de abundancia de esta especie, disminuyen a partir de los 3 años de edad. Esta desaparición de los peces de mayor tamaño y edad de la población de la brótola, está más relacionada con los efectos de la explotación pesquera que con las variaciones de los parámetros físico-químicos que puedan presentarse en el Mediterráneo Occidental. Asimismo, la longitud y la longevidad dependen de la disponibilidad del alimento, madurez gonádica y acumulación de substancias de reserva. (Gallardo-Cabello y Gual-Frau 1984). ConclusionesLa curva teórica de crecimiento que más se ajusta a los valores observados por medio de la lectura de los otolitos, "sagitta", es la que se obtuvo por medio de la técnica de Tomlinson y Abramson y la aplicación del método de Beverton. Los parámetros del crecimiento obtenidos, son los siguientes: Longitud máxima (Lsigma) = 574.39 mm Peso entero máximo (Wsigma) = 1337.356 gr. Peso desvicerado máximo (Wsigma) = 1218.588 gr. Coeficiente k = 0. 104769 Edad teórica de la longitud cero (to) = —0.4637287 Durante los primeros meses de vida de la brótola Phycis blennoides, se presenta la etapa de mayor crecimiento lineal. La velocidad de crecimiento disminuye considerablemente conforme la edad se incrementa. El crecimiento de los machos es menor que el de las hembras a partir de los tres años de edad, esta diferencia aumenta en proporción directa con la edad. Los parámetros de crecimiento por sexos son los siguientes: , ***********ELEMENTO SIN ESTILO ********* tablaLa relación peso-longitud es:
peso (W) = 4.0 x 10 -6 x longitud (mm) 3.094 para los peces enteros, y
peso (W) = 3.0 x 10 -6 x longitud (mm) ³·¹²
para los desviscerados
Los datos obtenidos sugieren la existencia de un crecimiento compensatorio a partir de los tres años de edad, es decir, que los peces que en los primeros años alcanzaron una talla y/o un peso relativamente bajos, después crecieron más intensamente. El límite de longevidad (A 0.95) es de 28 años, o sea la edad requerida para alcanzar el 95% de L(sigma). La brótola, en comparacion con otras especies de peces del Mar Catalán, presenta un crecimiento lento y una longevidad elevada. La brótola del Mar Catalán presenta un ritmo de crecimiento mayor, tallas menores y una menor longevidad en comparación con la brótola de las costas de Inglaterra, única estudiada, aunque escasamente. La edad límite encontrada para los machos es de 13 años y 31 años para las hembras AgradecimientosA Ramón Margalef, catedrático de Ecología de la Universidad Central de Barcelona, por la revisión del presente trabajo. A Conchita Allué y a Jaime Rucabado miembros del Instituto de Investigaciones Pesqueras de Barcelona por su cooperación en la realización del mismo. A Alfredo Laguarda, del Instituto de Ciencias del Mar y Limnología de la UNAM por su apoyo y estímulo para la publicación del presente trabajo. A Alicia Durán, del Instituto de Ciencias del Mar y Limnología de la UNAM quien amablemente elaboró las gráficas de este estudio. LITERATURAALVARIÑO A. y O. RODRIGUEZ, Publ. Inst. Esp. Ocean. La merluza, el bacalao y especies afines. 1955.125-127. 1: ALLEN, K. R., A method of fitting growth curves of the von Bertalanffy type to observed data. J. Fish. Res. Bd. Canadá1966.163-277.(3): 23 BAS, C., Proc. Tech. Pap. Gen. Fish. Coun. Mediterr.Fluctuations de la pêche de Merlangus poutassou et quelques considerations sur son controle.1963.417-420. 7: BEVERTON, R. J. H., Notes on the use of theoretical models in the study of the dynamics of exploited fish populations. U S. Fish and Wildl Serv. Fish. Lab., Beaufort, Misc. Contrib., 1954. 1-181.2: BHATTACHARYA, C. G., Biometrics A simple method of resolution of a distribution into gaussian components.1967.115-135. 23: BOTHA, L., Growth and otolith morphology of the cape hakes.Investl. Rep. Div. Sea Fish. S. Afr., 1971.1-32. 97: BRODY, S. Univ. Missouri Agric. Exp. Sta. Bull. Growth rates. 1927.101-215. 97: CARRILLO, J. Tesina. Univ. de la Laguna.Biología del Jurel (Trachurus trachurus (L) y Trachurus mediterraneus mediterraneus Steindachner) del Mar Medilterráneo Catalán.España1978 103 p. CASSIE, R.M. N. Z. Sci. Rev. The analysis of polymodal frecuency distributions by the probability paper method.1950.89-91. 8: CASSIE, R. M., Aust. J. Mar. Freshw Res. Some uses of probability paper in the analysis of size frecuency distributions 1954. 513-522. (3): 5 CASSIE, R. M., N. Z. J. Sci. Tests of significance for probability paper analysis. 1963.474-482.(4): 6 FABENS, A. J. Growth Properties and fitting of the von Bertalanffy growth curve. 1965. 265-289. 29: FIGUERAS A., Inv. Pesq. Datos sobre la edad y crecimiento de la pescadilla (Merluccius merluccius L.) del Levante (Sector Castellón) determinados por medio de los otolitos.1955.73-86. 1: FORD, E., J. Mar. Biol. Ass. Uk. NS. An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933. 1933.305-384. 19: GALLARDO-CABELLO, M.,. Características biológicas de Phycis blennoides (Brünnich, 1768) con especial referencia a algunas modificaciones debidas al efecto de la explotación pesquera. Tesis Doctoral. Universidad Central de Barcelona España1979. 420 p GALLARDO-CABELLO, M., An. Inst. Cienc. Del Mar y Limnol. Univ. Nal. Autón. México, Análisis de las frecuencias de tallas por medio de los métodos de Petersen, Cassie y Bhattacharya, para la determinación de la edad de la brótola Phycis blennoides (Brünnich 1768) en el Mediterráneo Occidental (Pisces: Gadidae) 1986a.187-196.(2): 13 GALLARDO-CABELLO, M., An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México,Determinación de la edad de la brótola Phycis blennoides (Brünnich, 1768) en el Mediterráneo Occidental (Pisces: Gadidae) 1986b.207-222. (2): 13 GALLARDO-CABELLO, M. y A. GUAL-FRAU, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoConsideraciones bioecológicas durante el crecimiento de Phycis blennoides (Brünnich, 1768) en el Mediterráneo Occidental (Pisces: Gadidae) 1984.225-238.(1): 11 GRAHAM, M., Fish. Invest. Lond. Ser. Report on the North Sea Cod. 1934. 13-22.2: GILBERT, C. H., Rep. B. C. Comm. Fish. Constributions to the life history of the sockeye salmon. 1914.53-78. 1: GULLAND, J. A., FAO Fish. Tech. Pap.Manual of Methods of fish populations analysis. 1964.1-60. 40: HART, T. J., J. Cons. Perm. Int. Explor. Mer. A note on growth zones in bones of Phycis blennoides.1951.335-340. 16: HICKLING, C. F., Fishery Invest. Lond. Ser.The natural history of the hake. Age determination and the growth rate. 1933. 1-120.(2): 213 JENSEN, A.S. y P. M. HANSEN, Cons. Int. Explor. Mer. Rapp. et. Proc. - Verb. Investigations of the Greeland cod (Gadus callarias L.) with and introduction on the history of the Greeland cod fishery. 1931.120-134.(1): 72 LARRAÑETA, M. G., Doc. Tech. VIII C. G. P. M., Les constantes de la croissance de la sardina de Castellón.1965.1-5. 39: LARRAÑETA, M. G., Inv. Pesq., Crecimiento del Pagellus erythrinus de las costas de Castellón.1967.185-258.31 (2): MATTA, F. Boll. Pesca Piscis. Idrobiol. La pesca a strascico nell'Arcipelago Toscano.1959.23-326.(1): 13 MARTIN, W. R., , ICNAF., 3rd Ann. Meet. Vital statistics of the cod and haddock in Subarea 4.1953. 1-20. 39: NEAVE, F., J. Fish. Res. Board. Can. Principles affecting the size of pink and chum salmon populations in Brittish Columbia. 1954. 450-491. 9: NELSON, M. O y H. A. LARKINS, Distribution and biology of the Pacific hake: A synopsis, Circ. Fish. Wildl. Serv., Wash. 1970.23-33. 332: NIKOLSKY, G. W., Vopr. Ikhtiol.Biological basis of mathematical simulation of fish population dynamics 1963.1-29.(4): 3 NICHY, F. E., Res. Bull. Int. Comm. Northw. Atlant. Fish. Growth patterns of otoliths from young silver hake, Merluccius bilinearis (Mitch.). 1969. 107-117.6 PETERSON, C. G. J., Berentning fra de Danske Biologiske Station for 1890 (91),Fiskenbiologiske forhold i Holboek Fjard, 1890-911892.121-183. 1: RAITT, D. F. S., In: Symposium on Ecology of Pelagic Fishes in Arctic Waters, The biology and commercial potential of the blue whiting (Micromesistius poutassou) in the North East Atlantic.ICES,1966. September, 1966. RICKER, W.E., J. Fish. Res. Board Can. Effects of size selectivite mortality and sampling bias on estimates of growth, mortality, production, and yield. 1969.479-541. 26: RIKHTER, V. A., J. Ichitol. A study of the Dinamics of Read Hake (Urophycis ch uss) catches from the Northwest Atlantic by the Method of Simulation. 1974.484-49114 (4): ROBLES, R., S. IGLESIAS, F. J. PEREIRO y J. A. PEREIRO, Bol. Inst. Esp. Ocean. Primeros datos sobre la dinámica del "stock" de merluza europea (Merluccius merluccius L) frente al litoral gallego 1975. 1-21. 196: RODRIGUEZ-RODA, J. Inv. Pesq.Biología del atún, Thunnus thynnus L., de la costa sudatlántica de España. 1964. 33-146.25: ROLLEFSEN, G., Cons. Int. Explor. Mer. Rapp. et Pro. Verb. The cod otolith as a guide to race, sexual development and mortality.1934.177-190. 88 (22): SAEMUNDSON, B., Med. Komm. Danmarks Fisk. og Havunders, Ser. Fish. On the age and growth of the cod (Gadus callarias) in Iceland waters.1923.32-50.(3): 7 SCOTT, D. C., Invest. Indiana Lakes streams, A study of a stream population of rock bass.1949.169-234. 3: STEVENS, W. L., BiometricsAsymptotic regression. 1951. 247-267.(3): 7 SUAU, P., Inv. Pesq. Contribución al estudio de la biología de Lithognathus mormyrus L. (Peces espáridos). 1970. 237-265. (2): 34 SUAU, P. y J. LÓPEZ, Inv. Pesq.Contribución al estudio de la dorada, Sparus auratus L. 1976.169-199.(1): 40 SUND, O., Aarsbereining Vedkommende Norges Fiskereí (1910), Undersokelser over brislingen i norske farvand vaesentling paa grundlag av 'Miichael Sars' togt 1908. 1911.357-410. 3: TAYLOR, C.C., J. Conseil Cod growth and temperature. 1958.366-370(3): 23 TAYLOR, C. C. J. Conseil Temperature and growth - The Pacific razor clam. 1959. 93-101.(1) 25 TAYLOR, C. C. J. Conseil Temperature, growth and mortalilty. The Pacific cockle. 1960.117-124.(1): 26 TAYLOR, C.C., J. Conseil Growth equations with metabolic parameters.1962.270-286.(3): 27 TOMLINSON, P. K. y N. J. ABRAMSON, Fish. Bull. Calif. Fitting a von Bertalanffy growth curve by least squares. 1961.3-69. 116: TROUT, G. C., Rapp. p. -V Reun. Cons. Int. Explor. Mer.Otolith growth of the Barents sea cod.1954. 89-102. 136: VON BERTALANFFY, L., Human Biology A quantitative theory of organic growth.1938.181-213.10 (2): WALFORD, L. A., Biol. Bull. A new graphic method of describing the growth of animals.1946. 141-147. 90 (2): WATKIN, E.E. Rep. Mar. Freshwater Invest. Investigations of Cardigan Bay herring Univ. Coll. Wales, (Aberystwyth) 1927.80-127. 2: WEATHERLY, A. H., Nature Ecology of fish growth. 1966.1321-1324. 212: WHEELER, A.,. The fishes of the Brittish Isles and N. W. Europa. Michigan State Univ. Press. 1969.613 p ZUÑIGA, L. R., Inv. Pesq. Estudio del crecimiento de Boops boops L. del Levante Español.1967.383-418.(3): 31
|