|
DETERMINACIÓN DE LA EDAD DE LA
BRÓTOLA PHYCIS BLENNOIDES (BRUNNICH 1768) EN EL MEDITERRÁNEO
OCCIDENTAL (PISCES: GADIDAE)
Trabajo recibido el 25 de septiembre de 1984
y aceptado para su publicación el 30 de enero de 1985.
MANUEL GALLARDO-CABELLO
Instituto de Ciencias del Mar y Limnología. Universidad
Nacional Autónoma de México. Contribución 482 del
Instituto de Ciencias del Mar y Limnología, UNAM
En el presente trabajo se establecen ocho grupos de edad, así
como la relación edad-longitud, mediante el análisis del otolito
sagitta de las poblaciones de Phycis
blennoides (brótola) que habitan en el Mediterráneo
Occidental. Asímismo, se describe la anatomía del otolito y se
estudia la relación entre la longitud, la anchura y el peso de la
sagitta, la cual queda expresada por los valores siguientes:
y=0.382 x 0.96 (longitud-anchura); y = 0.074 x
3.034 (longitud-peso). Las bandas de crecimiento
rápido en los bordes indican que éste fue mayor en los meses de
marzo, abril y mayo (primavera) mientras que en las de crecimiento lento en los
bordes fue mayor durante los meses se septiembre, octubre y noviembre
(otoño). El desarrollo de dichas bandas se relacionó con la
temperatura, la edad, la alimentación y el desove. Asímismo, se
relaciona el crecimiento del otolito con el del pez. El poreentaje de otolitos
cristalinos fue de 1. 32 %. Se encontraron diferencias de crecimiento entre
sexos, siendo este mayor en las hembras que en los machos para cada grupo de
edad considerado.
ln this study, by means of the
sagitta otolith analysis eight age groups and the relacion
age length of populations of Phycis blennoides (Greather
Forkbeard) in the Western Medíterra nean Sea are established. In
adition, the anatomy of the sagitta is described and the growth in length,
width and weigth is studied: its relation is expressed in the following values:
y = 0.382 x 0.96 (length-width) and y = 0.074 x
3.034 (length-weigth). The fast growing bands on the
edges show that their formation was higher during the months of march, april
and may (spring) while the slow growing bands on the edges were formed during
the months of september, october and november (autumn). The development of
those bands was rela ted to temperature, age, feeding and spawning. Also,
growth of the otolith is related to growth of the fish. The percentage of
crystalline otoliths was 1.32%. Also, differences in growth between sexes were
detected: the females reach a bigger size than males for each group
age.
Cuando el ritmo de crecimiento de un pez presenta fluctuaciones, se producen marcas o impresiones en estructuras duras como son: huesos, escamas, opérculos, otolitos u otros, que pueden servir para reconocer la edad y el crecimiento del animal. En los otolitos se forman dos marcas anuales o anillos cada año. Una corresponde a la época de crecimiento rápido de aspecto visual opaco y otra correspondiente a la época de crecimiento lento, mucho más translúcida, a la que se denomina banda hialiana. En el otolito la determinación de la edad se basa en la formación regular de una banda hialina y otra opaca, en torno al núcleo. El análisis de la edad de los peces es uno de los parámetros más importantes para el estudio de la dinámica de poblaciones. El conocimiento de la misma permite llevar a cabo el cálculo del crecimiento por medio del cual se puede estudiar la mortalidad, el reclutamiento y otros importantes parámetros de la población. MATERIAL Y MÉTODOSLos ejemplares se obtuvieron frente a las costas de Cataluña, entre Blanes y Vilanova i la Geltrú, de profundidades que oscilaron entre 150 y 650 m, de septiembre de 1976 a septiembre de 1978. Para el estudio del desarrollo del otolito de Phycis blennoides se examinaron un total de 1132 otolitos ("sagitta"' derecha), pertenecientes a ejemplares de 70 a 490 mm de longitud total. Los otolitos fueron observados, sumergidos en agua, por medio de una lupa binocular provista de un ocular graduado. De cada uno de ellos se registró la longitud total, la anchura y el peso. Para la determinación de los anillos de crecimiento se empleó la siguiente metodología a). La "sagitta" fue sumergida en xilol como líquido aclarante y observada por medio de una lupa binocular con iluminación directa o transmitida. b). Se obtuvieron por simple rotura, secciones transversales del otolito a nivel del núcleo que fueron sumergidas en xilol y observadas por medio de una lupa binocular. c). Las secciones del otolito fueron quemadas con un méchero Bunsen al nivel del núcleo. (Método de Christensen, 1964). d). Las secciones de los otolitos fueron teñidas con benzoato de metilo. (Técnica de Aker, 1961). El método que proporcionó mejores resultados fue el primero, empleando luz transmitida de gran intensidad. Así, en todos los casos se decidió observar los otolitos por transparencia. En estas condiciones las zonas hialinas aparecen como bandas claras y las opacas como bandas oscuras, al inverso de como se ven con la luz reflejada. En cada caso se anotó si los bordes del otolito presentaban bandas de crecimiento rápido o lento, lo que permitió conocer el momento de aparición de las mismas y relacionarlas con las variaciones ambientales. A fin de conocer las variaciones en el tamaño del núcleo y en la amplitud de las bandas del otolito de Phycis blennoides, se obtuvieron las mediciones correspondientes. Dicho examen se realizó con una lupa binocular provista de un objetivo graduado, colocando la "sagitta" en sentido transversal sobre el borde dorsal de la cara interna. Todas las mediciones fueron efectuadas en la "sagitta" derecha, para eliminar posibles diferencias debidas a variaciones existentes entre los otolitos del mismo ejemplar. La edad de los ejemplares se estableció contando el número de anillos hialinos que rodean al núcleo. De los 1, 132 otolitos examinados, el 70 % fueron analizados sin dificultad, un 20 % fueron revisados varias veces y un 10 % fueron descartados porque los anillos de crecimiento no pudieron observarse con claridad. RESULTADOS Y DISCUSIÓN
DESCRIPCIÓN DE LAS "SAGITTAS" DE PHYCIS BLENNOIDES Y SU VARIACIÓN CON LA EDADEn la brótola, Phycis blennoides, las "sagittas" son bastante grandes, carácter común a la familia de los gádidos. Son gruesas, lo que dificulta la identificación de las bandas de crecimiento rápido y lento, problema que se acentúa conforme el animal envejece y las dimensiones de los otolitos aumentan en tamaño y en grosor. La "sagitta" es alargada, la altura máxima está contenida de 2.71 a 3.1 veces en la longitud (Figs. 1 y 2). El border anterior presenta un "rostrum" saliente, redondeado o con pequeñas denticiones; la "excisura major" es reducida y el "antirostrum" está poco desarrollado. 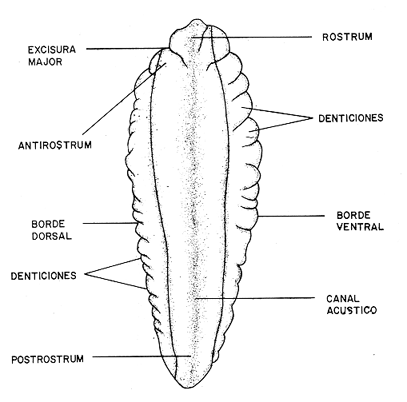 Fig. 1. Otolito de Phycis blennoídes ("sagitta" derecha, cara interna) mostrando sus características distintivas (aumentado 8 veces). 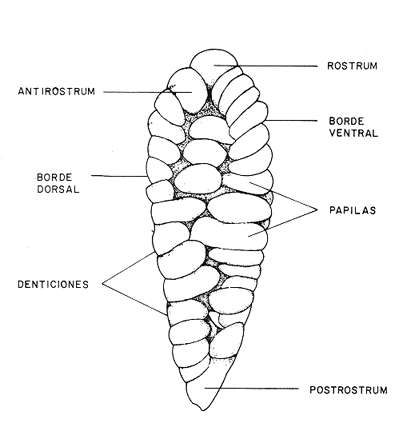 Fig. 2. Otolito de Phycis blennoides ("sagitta" izquierda, cara externa) mostrando sus características distintivas (aumentado 16.2 veces). El borde dorsal es ascendente, desde el rostrum hasta el centro del otolito y desciende con suave inclinación hasta el borde posterior; tiene pequeñas denticiones bastante regulares en forma de sierra, que tienden a desaparecer conforme el animal envejece, observándose un borde dorsal liso, en individuos de 360 mm de longitud total. El borde posterior presenta un "postrosturm" agudo y bien desarrollado, carece de "excisura minor" y "pararostrum". El borde ventral es ligeramente curvo desde el "rostrum" hasta el centro del otolito, y rectilíneo conforme asciende desde el centro del otolíto hasta el borde posterior. Presenta denticiones, las cuales tienden a hacerse irregulares o bien a desaparecer cuando el animal envejece (Fig. 3). La cara interna del otolito es bastante convexa, carácter que se acentúa con la edad, su superficie es plana y está recorrida en su totalidad por el "sulcus" que es ancho, poco profundo y uniforme, sin presentar una clara diferenciación en "ostium" y "cauda".  Fig. 3. Selección de "sagittas" de Phycis blennoides ("sagitta" izquierda, cara interna) mostrando las diferencias en forma y tamaño de acuerdo a las diferentes clases de tallas y grupos de edad(aumentado 1.6 veces). La cara externa es cóncava y está recorrida en su totalidad por numerosas papilas o granulaciones de diferente tamaño, las cuales están menos marcadas en los otolitos de mayor tamaño; el borde dorsal presenta una disminución brusca del espesor en sentido longitudinal, a nivel de la línea de papilas. DESARROLLO DEL OTOLITOLos datos obtenidos se muestran en la Tabla 1. De acuerdo a los resultados se trazaron las gráficas de relación entre la longitud y la anchura del otolito, y entre la longitud y el peso del mismo, empleando el método de mínimos cuadrados. En la figura 4, se observa un punto de inflexión en la línea de regresión entre la longitud y la anchura del otolito, que corresponde a la longitud total del pez de 170 mm. La diferenciación de las glándulas sexuales de Phycis blennoides se inicia a partir de dicha talla (Gallardo-Cabello, 1979), razón por la cual se han desdoblado las series de datos en dos partes: 70-170 mm y 180-490 mm de la longitud total del pez. La relación alométrica existente entre la longitud y la anchura del otolito, viene expresada por el valor del exponente o índice alométrico, k = 0.96. Se trata por lo tanto, de una relación de crecimiento muy próxima a la isometría. El índice de correlación que se ha hallado para comprobar la relación entre las dos series de datos es igual a 0.998. La fórmula hallada es la siguiente: y = 0.382 x 0.96 Donde: y representa la anchura y x la longitud del otolito. En la primera parte del periodo estudiado (70-170 mm de longitud total) el otolito muestra un crecimiento significativamente isométrico; el valor del índice alométrico es k = 1.010. Durante el resto de la vida de Phycis blennoides (180-490 mm de longitud total) el crecimiento del otolito muestra una clara tendencia hacia una alometría negativa, obteniéndose un valor del índice de regresión de k = 0.889. 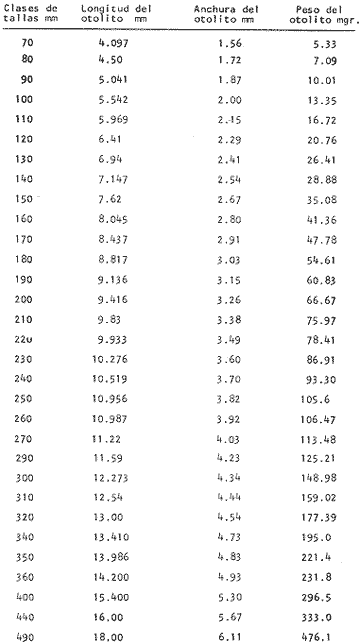 TABLA 1 RELACIÓN CLASES DE TALLAS-LONGITUD-ANCHURA Y PESO DEL OTOLITO DE PHYCIS BLENNOIDES De lo anterior se concluye que en la primera etapa, el crecimiento del otolito es isométrico sin variaciones a lo largo del desarrollo, con lo que la forma de los mismos se mantiene constante y una vez que comienza la diferenciación sexual de esta especie, el otolito muestra una clara tendencia hacia una alometría negativa. Esto puede deberse a que a partir de esta época la actividad metabólica del pez y particularmente el metabolismo del calcio, se encuentran orientados hacia su desarrollo gonádico y a la formación de los productos sexuales. 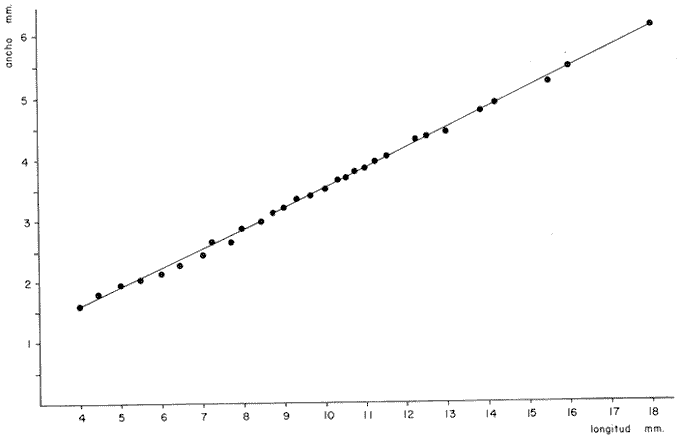 Fig. 4. Línea de regresión entre la longitud total y el ancho total del otolito de Phycis blennoides. Así, se presenta un considerable gasto de energía en la formación de los mismos, que se traduce en la disminución de las dimensiones del otolito. La relación entre la longitud y el peso del otolito, se caracteriza por el valor del índice alométrico k = 3.034; se trata por lo tanto, de una relación isométrica de crecimiento. El índice de correlación obtenido para comprobar la relación entre estas dos series de datos, es de 0.997. La fórmula encontrada es la siguiente: y = 0.074 x 3.034 Donde: y representa el peso y x la longitud del otolito. En la figura 5, se observa un punto de inflexión en la línea de regresión, entre el log de la longitud y el log del peso del otolito, que corresponde a la longitud total del pez de 140 mm. Se han desdoblado las series de datos en dos partes: 70-140 mm y 150-490 mm de longitud total del pez, obteniéndose en la primera parte del periodo estudiado, un valor de k = 4.32 que corresponde a un crecimiento alométrico positivo, mientras que el valor obtenido para el segundo periodo estudiado es de k = 2.85, que corresponde a un índice alométrico negativo. Como se muestra más adelante, la brótola Phycis blennoides alcanza el primer año de edad a la longitud de 140 mm, por lo que podría deducirse que el primer periodo (70-140 mm) corresponde a la formación del núcleo del otolito y de las primeras bandas de crecimiento rápido y lento, cuya amplitud es mayor. Esto trae como consecuencia un mayor depósito de sales de calcio. El resto de las bandas formadas durante el transcurso de la vida del pez (140-490 mm) presentan un crecimiento mucho más limitado. 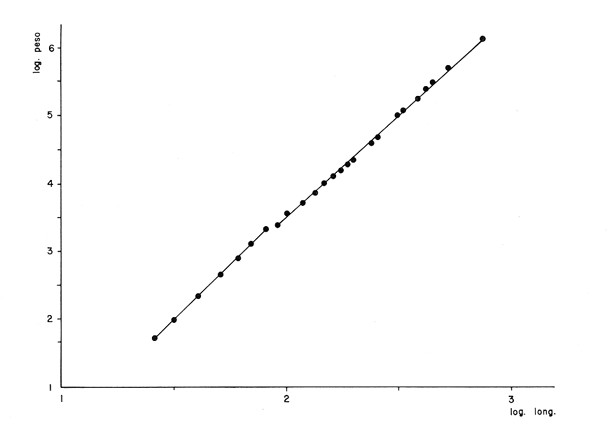 Fig. 5. Línea de regresión entre el logaritmo de la longitud y el logaritmo del peso del otolito de Phycis blennoides. ESTRUCTURA: ANILLOS DE CRECIMIENTOLas interpretaciones de los anillos de crecimiento de la "sagitta" de Phycis blennoides, aparecen en la Tabla 2. La anchura de los núcleos examinados varía dentro de una amplitud de 0.83 a 1.30 mm, siendo la anchura promedio de 1. 10 mm. El núcleo es excéntrico en cada plano del otolito, fenómeno que se acentúa conforme el pez envejece y alcanza la madurez sexual. Un patrón muy similar se observa en los otolitos de otros Gáditos (Bas, 1966; Blacker, 1974). La primera banda de crecimiento lento (hialina) o sea la que se forma después del núcleo, se halla totalmente delimitada y completamente desarrollada; el valor de la amplitud media es de 0.29 mm. (Tabla 2). En la banda siguiente, que es la primera de crecimiento rápido (opaca), éste es más intenso y alcanza una anchura promedio de 0.39 mm. Por el contrario, en la segunda banda de crecimiento lento el descenso es considerable, como se puede deducir del valor medio observado, 0.19 mm. La segunda banda de crecimiento rápido presenta un tamaño medio de 0.27 mm, valor inferior con respecto a la primera banda opaca pero superior al señalado para la banda anterior. En el resto de los anillos de crecimiento se observa una disminución progresiva de la anchura, con la particularidad de que esta disminución no aparece de forma contínua, sino más bien describiendo un ritmo ondulatorio con tendencia a desaparecer. Este ritmo presenta los valores máximos en la época de crecimiento rápido y los mínimos en la época de crecimiento lento. 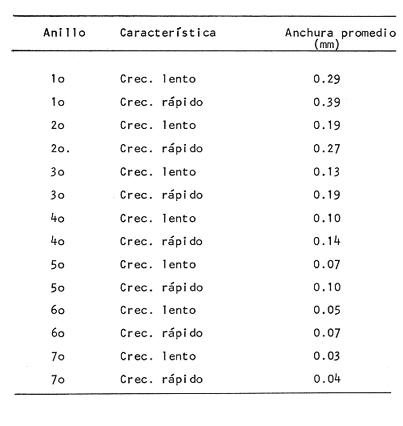 TABLA 2 DIAGRAMA DEL OTOLITO DE PHYCIS BLENNOIDES En el interior del núcleo de algunos ejemplares jóvenes (80 a 140 mm de longitud total) se observó una fina banda hialina, que podría corresponder al cambio de habitat pelágico al bentónico durante el periodo juvenil. Esta banda hialina tiende a desaparecer conforme los otolitos aumentan sus dimensiones, para formar un núcleo de aspecto completamente denso y compacto. OTOLITOS CRISTALINOSEl análisis de los otolitos de Phycis blennoides revela la formación completa o parcial de otolitos cristalinos (Fig. 6) en la proporción del 1.32%. Los otolitos cristalinos se presentan muy frecuentemente en los miembros de la familia Gadidae (Mankevich, 1966), como una disrupción parcial o alguna alteración en los mecanismos que controlan el crecimiento del otolito. Pueden producirse cuando la formación de la matriz orgánica es interrumpida por alguna substancia presente. Generalmente, uno de los otolitos es parcial o completamente cristalino, mientras que el otro es normal (Blacker, 1974). En Gadus morhua, Pleuronectes platessa y Gadus aeglefinus los otolitos cristalinos, formados parcial o totalmente, alcanzan un valor de 5 % (Blacker, 1974). El porcentaje de otolitos cristalinos observados en Phycis blennoides no representa un valor muy elevado comparado con otros Teleósteos. 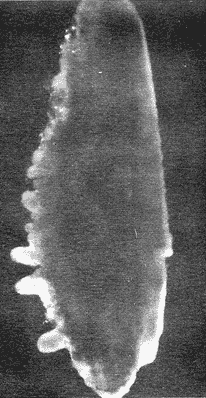 Fig. 6. Otolito cristalino de Phycis blennoides. PATRÓN DE CRECIMIENTO ESTACIONAL
Tiempo de Formación de las Bandas de Crecimiento Rápido y LentoDe acuerdo con la bibliografía consultada (Tabla 3) existe una gran confusión en cuanto a lo que los diversos autores consideran como términos hialino y opaco. Lo anterior, puede deberse al empleo de diversos sistemas de iluminación (Blacker, 1974). En la mayoría de los casos las bandas de crecimiento lento aparecen durante el otoño y el invierno, correspondiendo a la época en que las condiciones son desfavorables y el crecimiento del pez se reduce (Hickling, 1933; Trout, 1954). Las bandas de crecimiento rápido aparecen durante la primavera o el verano, que corresponden al periodo de máximo crecimiento (Dannevig, 1933; Molander, 1947). 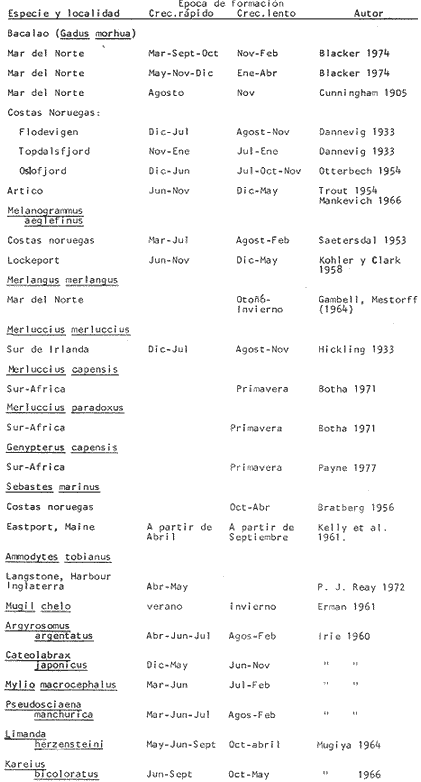 TABLA 3 ÉPOCA DE APARICIÓN DE LAS ZONAS DE CRECIMIENTO RÁPIDO Y LENTO DE OTRAS ESPECIES Por otro lado, Irie (1957 y 1960) establece que en varias especies, las bandas de crecimiento lento aparecen durante el verano, cuando los peces se alimentan activamente, mientras que las de crecimiento rápido se forman en invierno-primavera cuando se reduce la actividad alimenticia del pez. En el caso de la brótola, Phycis blennoides, los resultados han sido claros, en la figura 7 y en la Tabla 4 se observa que el porcentaje de otolitos con bandas de crecimiento rápido en los bordes es mayor durante los meses de marzo, abril y mayo (primavera), mientras que el mayor porcentaje de otolitos con bandas de crecimiento lento en los bordes aparece durante los meses de septiembre, octubre y noviembre (otoño). Por lo tanto, cada año se depositan una banda de crecimiento rápido y otra de crecimiento lento, permitiendo que la estimación de la edad basada en el número de bandas de crecimiento rápido y lento sea válida para Phycis blennoides RELACIÓN ENTRE LA FORMACIÓN DE BANDAS DE CRECIMIENTO Y PARAMETROS FÍSICOS Y BIOLÓGICOSLas causas de la formación regular de las bandas de crecimiento rápido y lento, a través de un cielo anual, han sido estudiadas por diversos autores. TemperaturaDannevig, (1956) establece que la formación de las bandas de crecimiento rápido y lento en otolitos y escamas de bacalao, está directa o indirectamente influida por la temperatura. Por otro lado, los experimentos de Molander (1947) e Irie (1960) muestran que los peces que han sido mantenidos bajo condiciones constantes de temperatura y salinidad, presentan cambios estacionales en el patrón de crecimiento de las bandas del otolito y que el desarrollo de las mismas es independiente de estos parámetros. 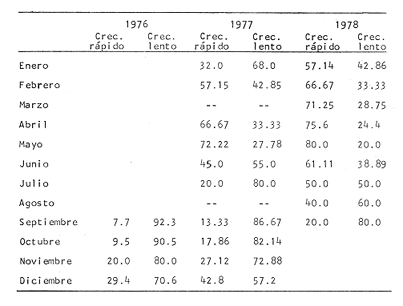 TABLA 4 PORCENTAJES MENSUALES DE OTOLITOS CON BANDAS DE CRECIMIENTO RÁPIDO O LENTO EN LOS BORDES 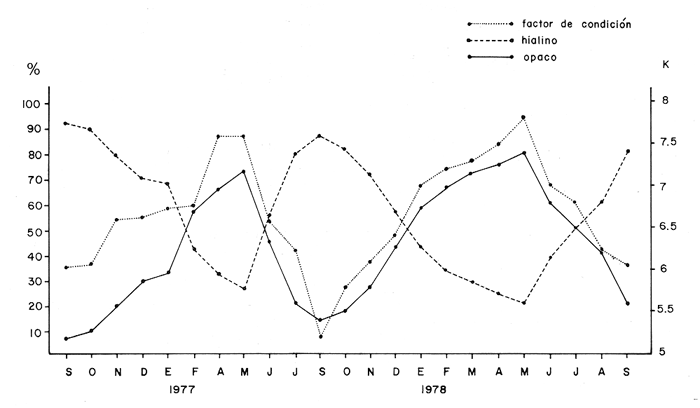 Fig. 7. Frecuencia de otolitos con el borde opaco e hialino; y valores del factor de condición. Blacker (1974), observa en la formación de las bandas de crecimiento del otolito, la repetición de un patrón estacional a través de una sucesión de años, resultados que no podrían obtenerse si la formación de dichas bandas dependiese absolutamente de factores ambientales. La influencia de la temperatura en la formación de las bandas de crecimiento rápido y lento en la "sagitta" de Phycis blennoides, parece no tener un efecto directo. Los registros de la temperatura en los fondos de la plataforma y talúd continentales donde habita la brótola, no presentan cambios estacionales marcados a partir de los 200 metros. Hasta los 700 metros de profundidad la temperatura oscila entre 12.75°C y 13.5°C en invierno y verano, respectivamente (Gallardo-Cabello, 1979); dicha variación no se considera significativa como para influir directamente en la formación de las bandas del otolito de Phycis blennoides. Alimentación y DesoveBilton y Robbins (1971) encuentran una gran influencia entre el nivel alimenticio y la formación de bandas de crecimiento en las escamas del salmón (Oncorhinchus nerka). Hickling (1933), Molander (1947), Botha (1970 y 1971) y otros, establecen una fuerte correlación entre las bandas de crecimiento rápido y lento y el nivel alimenticio, por estaciones, de diversas especies. Demuestran que las bandas de crecimiento rápido aparecen en aquellos periodos en que el pez se alimenta activamente, mientras que las bandas de crecimiento lento se depositan en las épocas en que el pez se encuentra en condiciones desfavorables y su alimentación disminuye. En el caso de Phycis blennoides, se encontró una fuerte correlación entre la formación de bandas de crecimiento rápido y lento y el nivel y época de alimentación del pez, durante cada mes; determinado por medio del factor de condición: K = P/L³ x 100 (Fulton) (P = peso y L longitud total del pez) En la Tabla 5 y en la figura 7, se observan los resultados obtenidos. El índice del factor de condición comienza a incrementarse a partir del mes de noviembre, alcanzando sus máximos valores en los meses de abril y mayo época en que se forma la banda de crecimiento rápido. 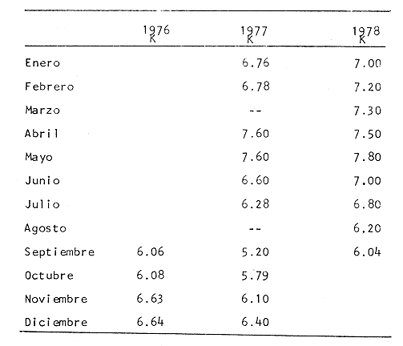 TABLA 5 VALOR MENSUAL DEL FACTOR DE CONDICIÓN Este periodo durante el cual el pez se alimenta más activamente, coincide con dos épocas de máxima productividad en el mar Catalán, correspondientes a los meses de noviembre-diciembre y febrero-abril (Margalef, 1967). Este aumento de la productividad frente a las costas de Barcelona, se debe a una mayor disponibilidad de nutrientes, principalmente nitratos y fosfatos, procedentes de los niveles profundos donde son removidos por medio de dos movimientos ascensoriales de corrientes verticales turbulentas, relacionadas con los vientos de la componente Suroeste (Gallardo- Cabello, 1979). Lo anterior trae como consecuencia un aumento del fitoplancton, así como del zooplancton que se alimenta de él y de los diferentes organismos que integran la red trófica, incrementándose de esta manera las fuentes alimenticias de Phycis blennoides durante estas épocas. El índice del factor de condición decrece a partir del mes de junio, alcanzando su valor mínimo durante el mes de septiembre, época en que se forma la banda de crecimiento lento. Este descenso del valor del factor de condición puede estar asociado con la época de desove, que en esta especie ocurre durante los meses de julio y agosto, coincidiendo con la formación de la banda de crecimiento lento, con lo cual se forma una sola banda. Es muy probable que durante esta época el desgaste corporal del pez sea muy grande, ya que su metabolismo se encuentra orientado hacia la formación de los productos sexuales y su peso disminuye considerablemente. Por otra parte, el valor del índice de replección gástrica alcanza un valor mínimo durante esta época (Gallardo-Cabello, 1979), lo que demuestra que el pez se alimenta menos. EdadPueden presentarse amplias variaciones en el tiempo de formación de las bandas de crecimiento rápido y lento, conforme se incrementa la edad del pez. Dannevig (1949) establece que en los peces de mayor edad, la zona opaca comienza a formarse dos meses después que en los peces jóvenes. Williams et al. (1974) observan en el bacalao del Mar Artico, que en individuos de dos años la formación de la banda de crecimiento rápido comienza en abril, mientras que en ejemplares de siete años ésta comienza a formarse a partir de junio. Blacker, (1974) observa algo similar en el bacalao del Mar del Norte (Tabla 3). En el caso de Phycis blennoides no se registró ningún cambio en las épocas de formación de las bandas de crecimiento a través de los grupos de edad, en todos los casos se observó que los valores obtenidos coincidían para cada una de las épocas mencionadas anteriormente. RELACIÓN ENTRE EL CRECIMIENTO DEL OTOLITO Y EL DEL PEZRelacionando la longitud y la anchura del otolito con la longitud total del pez se encontró la relación existente entre estos dos aspectos del desarrollo. En la Tabla 6 y en las figuras 8 y 9, pueden observarse los resultados obtenidos, así como el cambio de ritmo cuando los peces alcanzan la talla de 170 mm de longitud total. 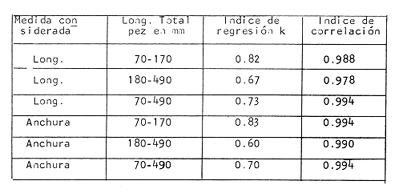 TABLA 6 RELACIÓN ENTRE EL CRECIMIENTO DEL OTOLITO Y DEL PEZ Los valores son negativos, k = 0.73, para la longitud y k = 0.70 para la anchura del otolito, respectivamente. Sin embargo, en el primer periodo los valores alométricos de k son de 0.82 y 0.83 respectivamente. Esto indica una alometría que tiende hacia la isometría, mientras que en el segundo periodo la relación alométrica es totalmente negativa, k = 0.67 y k = 0.60, respectivamente. Como ya se ha mencionado, el rango en el incremento de la longitud del pez decrece con la edad. Esta variación puede asociarse con el comienzo de la época de la diferenciación gonádica. Tales relaciones ocurren frecuentemente en muchas especies, particularmente en muchos miembros de la familia Gadidae (Hickling, 1933; Trout, 1954; Templeman y Squires, 1956; Mina, 1967; Harden Jones, 1969). En la bacaladilla, Micromesistius poutassou, esta diferencia es aún más acentuada que en Phycis blennoides; el valor del índice alométrico, en la relación entre la longitud del otolito y la longitud total del pez cuando este alcanza la talla de 150 a 160 mm de longitud total, pasa de 1.617 (120-168 mm de longitud total) a 0.706 (170-300 mm de longitud total; Bas, 1966). RELACIÓN EDAD LONGITUD
Interpretación de los Anillos de CrecimientoEn la Tabla 7 se muestran las longitudes promedio obtenidas para cada edad y en la Tabla 8 se muestran las longitudes medias para cada edad, tanto para los machos como para las hembras. Como puede observarse, los machos alcanzan una longitud menor que las hembras a una misma edad. Este fenómeno se presenta frecuentemente entre los miembros de la familia Gadidae (Hickling, 1933; Botha, 1971; Wheeler, 1966; Blacker, 1974).  TABLA 7 RELACIÓN EDAD-LONGITUD DE PHYCIS BLENNOIDES 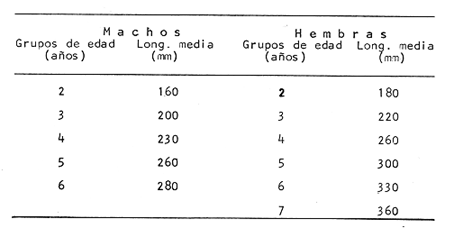 TABLA 8 RELACIÓN EDAD-LONGITUD POR SEXOS 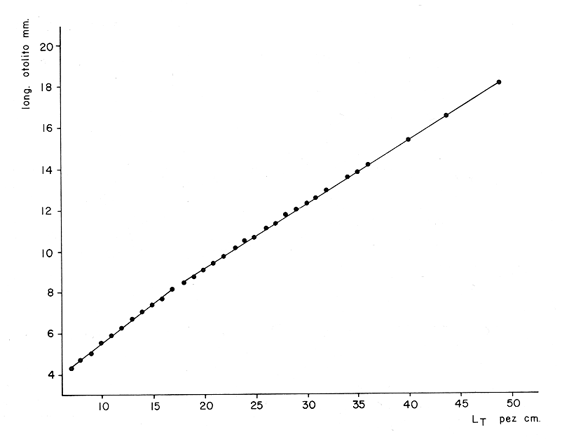 Fig. 8. Línea de regresión entre la longitud total de Phycis blennoides y la longitud total del otolito. 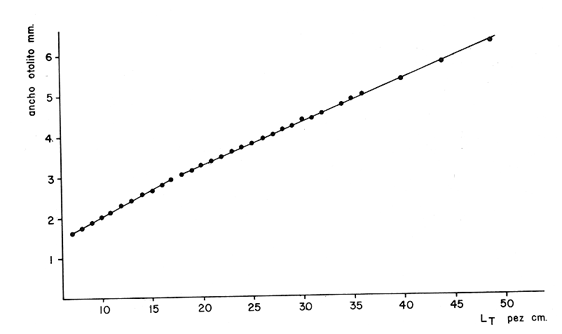 Fig. 9. Línea de regresión entre la longitud total de Phycis blennoides y la anchura total del otolito. ConclusionesEl mejor material para el estudio de la edad y el crecimiento son los otolitos. El método que proporcionó mejores resultados para la identificación de las bandas de crecimiento rápido y lento, fue el de analizar las "sagittas" sumergidas en xilol, por medio de una lupa binocular, con iluminación transmitida de gran intensidad. La forma del otolito varía conforme aumenta la edad del pez. La relación entre la longitud y la anchura del otolito está dada por dos periodos distintos: la primera etapa, 70-170 mm de longitud del pez, corresponde a un crecimiento isométrico; la segunda etapa, a partir de los 180 mm de longitud total del pez "momento en que comienza la diferenciación sexual de esta especie" corresponde a un crecimiento alométrico negativo. La relación entre la longitud y el peso del otolito está expresada por dos periodos: el primero entre los 70 y 140 mm de longitud total, con un valor del índice alométrico k = 4.32; el segundo periodo entre los 150 y 490 mm, expresado por un valor del índice k = 2.85. La relación entre la longitud y el peso de la "sagitta" indica un mayor incremento en peso durante los primeros años de la vida del pez, correspondiendo a la época de la formación del núcleo y de las primeras bandas de crecimiento. Durante el crecimiento en longitud de la "sagitta" es mayor el número de bandas que se forma en la parte posterior que en la anterior, crece más rápido el lado dorsal que el ventral y se deposita mayor cantidad de material en la cara interna que en la externa. La amplitud de las bandas concéntricas que se forman en torno al núcleo, disminuye conforme el pez incrementa su longitud. Las bandas son más anchas durante el periodo de crecimiento rápido que en el de crecimiento lento. El porcentaje de otolitos cristalinos es de 1.32%. El porcentaje de otolitos con bandas de crecimiento rápido en los bordes, es mayor durante los meses de marzo, abril y mayo (primavera); el mayor porcentaje de otolitos con bandas de crecimiento lento en los bordes, aparece durante los meses de septiembre, octubre y noviembre (otoño). Durante cada año se depositan una banda de crecimientc rápido y otra de crecimiento lento. Las variaciones de la temperatura en los fondos de la plataforma y talúd continentales, no son significativas para la formación de las bandas de crecimiento del otolito. La formación de las bandas de crecimiento rápido y lento, está fuertemente relacionada con la alimentación. A través de la edad, no se observó ningún cambio en las épocas de formación de las bandas de crecimiento rápido y lento. La relación entre la longitud total del pez y la longitud total del otolito está representada por dos periodos: en la primera etapa el valor del índice alométrico k es de 0.82 (70-170 mm de longitud total) y en la segunda etapa el valor de k es de 0.67 (180-490 mm de longitud total). La relación entre la longitud total del pez y la anchura total del otolito está representada por los siguientes periodos: 0.83 como valor del índice k para la etapa de 70-170 mm de longitud total del pez, y 0.60 para la etapa de 180-490 mm de longitud total. El análisis de los anillos de crecimiento de las "sagittas" permitió la identificación de ocho grupos de edad. Se encontraron diferencias de crecimiento entre sexos, siendo éste mayor en las hembras que en los machos para cada grupo de edad considerado. AgradecimientosA Ramón Margalef, catedrático de Ecología de la Universidad Central de Barcelona, por la revisión del presente trabajo. A Conchita Allué miembro del Instituto Investigaciones Pesqueras de Barcelona por su cooperación en la realización del mismo. A Alfredo Laguarda- Figueras, del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México por su apoyo y estímulo para la publicación del presente trabajo y Alicia Durán, quien amablemente elaboró las gráficas de este estudio. LITERATURAAKER, VON E. Untersuchungen über díe Biologie der Makrele (Scomber scombrus L.) in der Nordsee. Ber. Dt. Wiss. Komm. Meeresfursh. 1961 105-128 16 (2) BAS, C., y E. MORALES Inv. Pesq. Crecimiento y desarrollo de Micromesistius poutassou. I. Desarrollo del otolito. 1966 179-195 30 BILTON, H. T. Effects of starvation, feeding, and light period on circulus formation on scales of young Sockeye salmon. Fish. Res. Bd. Can. 1971 1749-1755 28 BLACKER, R. W. Sea Fisheries Research. Recent advances in otolíth studies J. Harden-Jones London 1974 67-90 BOTHA, L. The growht of the Cape Hake Merluccius capensis. InvestI. Rep. Div. Sea Fish. S. Afr. 1970 1-9 82 BOTHA, L. Growth and otolith morphology of the Cape hakes Merluccius capensis y M. paradoxus Investl. Rep. Div. Sea Fish. S. Afr. 1971 1-32 97 BRATBERG, E. On the interpretation of the opaque and hyaline zones in the otoliths of immature redfish. (Sebastes marinus L.) J. Conseil 1956 66-74 22 (1) CUNNINGHAM, J. T. Zones of growth in the skeletal structures of Gadidae and Pleuronectidae. Rep. Fish. Bd. Scot. 1905 125-140 23 (3) CHRISTENSEN, J. M. Burning of otoliths, a technique for age determination of soles and other fish. J. Cons. Perm. Int. Explor. Mer. 1964 73-81 29 (1) DANNEVIG, A. On the age and growth of the cod (Gadus callarías) from the Norwegian Skagerrack Fisk. Dir. Skr. Ser. Havunders 1933 1-145 4 (1) DANNEVIG, A. The variation in growth of young codfishes from the Norwegian Skagerrack coast. Cod seales as indicator of local stoks. Fisk Dir. Skr. Ser. Havunders 1949 122 9 (6) DANNEVIG, E. H. Chemical composition of the zones in cod otoliths. J. Cons. Int. Exploe. Mer. 1956 156-159 21 ERMAN, F. On the biology of thick-lipped grey mullet (Mugil chelo Cuv.) Rap. Proc-Verb. Reu. CIESM 1961 277-285 16 (2) GALLARDO-CABELLO, M. Caracteristicas biológicas de Phycis blennoides (Brünnich 1768) con especial referencias a algunas modificaciones debidas al efecto de la explotación pesquera. Universidad Central de Barcelona 1979 420 p. GAMBELL, R., y J. MESSTORFF Age determination in the Whiting (Merlangus merlangus L.) by means otoliths. J. Conseil 1964 393-404 28 (3) HARDEN JONES, F. R. Observations of the behaviour of fish made with the bifocal sector scanner. FAO. Fish. Rep. 1969 667-670 62 (3) HICKLING, C. F. The natural history of the hake. Age determination and the growth rate. Fish. Invest. Lond. Ser. 1933 1-120 13 (2) IRIE, T. On the forming season of anual rings (opaque and translucent zones) in the otoliths of several. Marine Teleost. J. Fac. Anim. Husbandry, Hiroshima Univ. 1957 311-317 1 (3) IRIE, T. The growth of the fish otolith J. Fac. Fish Anim. Husb. Hiroshima Univ. 1960 203-229 3 KELLY, G. F. y A. M. BARKER Observations on the behaviour, growth and migration of redfish at Eastport Maine. Sp. Publ. ICNAF 1961 263-275 3 KOHLER A. C., y J. R. CLARK Haddock scaleotolith for Haddock (Melanogrammus aeglefinus L.) from the Lockeport, N. S. area. J. Fish. Res. Bd. Canada 1958 1229-1238 15 (6) MANKEVICH, E. M. Methods of taking and reading the age samples of cod. Mater. Rybokhoz. Issled. Sev. Boss. 1966 53-56 1 MARGALEF, R. y J. CASTELLVÍ Inv. Pesq. Fitoplancton y producción primaria de la costa catalana, de julio de 1966 a julio de 1967. 1967 491-502 31 (3) MINA, M. V. Study of the relation between the weight of an otolith (sagitta) and the length of an individual in the cod population of the Barents and White Seas. Nauch. Dold. Vyssh. Shk. 1967 26-31 9 MOLANDER, A. R. Observations on the growth of the plaice and on the formation of annual rings in its otoliths. Svenska. Hydrogr. Biol. Konin. Skr. N. S. Biol. 1947 11 p. 2 (B) MUGIYA, Y. Bull. Jap. Soc. Scient. Fish. Calcification in fish and shell-fish-Seasonal ocurrence of a pre-albumin fraction in the otolith fluid of some fish corresponding to the period of opaque zone formation in their otoliths. 1964 445-467 30 MUGIYA, Y. Bull. Jap. Soc. Scient. Fish. Calcification in fish and shell-fish. A study on paper electrophoretic patterns of the acid mucopolysaecharides and Pas-positive materials in the otolith fluid of some fish. 1966 117-129 32 MUGIYA, Y. Bull. Jap. Soc. Scient. Fish. Calcification in fish and shell-fish Seasonal change in calcium and magnesium concentration of the otolith fluid in some fish with special reference to the zone formation of their otolith 1966 549-557 32 OTTERRECH, F. The cod population of the Oslofjord Rapp. P. V. Réun. Cons. Int. Explor. Mer 1954 15-21 136 PAYNE, A. Stock differentiation and growth of the Southern African kingklip Genypterus capensis. Investl. Rep. Div. Sea Fish. S. Afr. 1977 1-32 113 REAY, P.J. The seasonal pattern of otolith growth and its aplications to back-calculations studies in Ammodyctes tobianus L. Jour, Cons. Int. Explor. Mer 1972 485-505 34 (3) SAETERSDAL, G.S. The haddock in Norwegian waters. II Methods in age and growth investigations. Rep. Norw. Fish. Mar. Investl. 1953 5-46 10 (9) TEMPLEMAN, W., H. I. SQUIRES Relationship of otolith lengths and weigths in the haddock Melanogrammus aeglefinus (L.), to the rate of growth of the fish. J. Fish. Res. Bd. Can. 1956 467-487 13 TROUT, G. C. Otolith growth of the Barents Sea Cod. Rapp. P. V. Reun. Cons. Int. Explor. Mer 1954 89-102 136 WHEELER, A. The Fishes of the Bittih isles and N. W. Europe. Michigan State Univ. Press. 1966 613 p. WILLIAMS, T., y B. C. BEDFORD Ageing of Fish. The use of otoliths for age determination T. B. Bagenal Unwin Brothers Ltd. Surey 1974 114-123
|