|
ESTUDIO DE LA ULTRAESTRUCTURA. DEL
OTOLITO "SAGITTA" DE LA BRÓTOLA PHYCIS BLENNOIDES (BRUNNICH 1768) EN EL
MEDITERRÁNEO OCCIDENTAL (PISCES: GADIDAE)
Trabajo recibido el 25 de
septiembre de 1984 y aceptado para su publicación el 30 de enero de
1995.
MANUEL GALLARDO-CABELLO
Instituto de Ciencias del Mar y
Limnología, Universidad Nacional Autónoma de México.
Contribución 481 del Instituto de Ciencias del Mar y Limnología,
UNAM.
En el presente trabajo se realiza un
análisis sobre la ultraestructura del otolito sagitta
de Phycis blennoides (brótola) en el
Mediterráneo Occidental. La sagitta
estáformada por carbonato cálcico en forma de argonita y por una
matriz protéica de fibras radiales y concéntricas en torno al
núcleo. La internación entre las fibras protéicas y los
cristales de argonita produce en éstos una forma de crecimiento en
zig-zag. Los incrementos de crecimiento diario están representados por
dos finas bandas, cuyo grosor oscila entre 0. 5 y 2.5 µ, que
corresponden a una capa con mayor proporción de proteína y otra
con predominio de argonita. Las zonas de crecimiento rápido presentan
incrementos de crecimiento diario más anchos con fibras protéicas
más gruesas y cristales de argonita más grandes que las de
crecimiento lento.
In the present paper, an analysis of
the ultrastructure of the otolith, sagitta, of
Phycis blennoides (Greater Forkbeard) in the Western
Mediterranean Sea was made. Sagitta is compo sed of calcium
carbonate in the form or aragoníte erystals and an organic matrix or
radials and concentric fibers around the nueleus. Interaction between protein
fibers and aragonite fibri1s shows an intergrowth pattern in zig-zag. Daily
growth layers are represented by two thin bands: one, 0.5 µ wide and
one, 2.5 µ wide. Those bands correspond to two different production
rates of fibrous material. The first contains the highest amount of protein,
the second, contains the highest amount of aragonite. Fast growth layers are
formed by daily growth increases with protein fibers and argonite erystals much
wider than slow growth layers.
Los estudios sobre la estructura molecular de los otolitos fueron iniciados por Immermann (1908), quien describió la ultraestructura del otolito de Pleuronectes platessa. Ese mismo año, Maier realizó un análisis similar en Gadus morhua. Hickling (1931), estudió los otolios de Merluccius merluccius, llegando a la conclusión de que éstos estaban formados por una matriz de materia orgánica, con fibras radiales y concentricas y carbonato cálcico en forma de argonita, Irie (1955 y 1960) y Carlström (1963), determinaron por medio de la difracción de rayos X, que los otolitos estaban compuestos en su mayor parte por carbonato cálcico en forma de argonita. Morris y Kittleman (1967), encontraron que el sodio está presente en los otolitos de los peces óseos en una concentración de 1-2 % en un mol de calcio, por lo que sugirieron la presencia de cristales de shortita Na2, Ca2 (Co3) y pirssonita (Na2 CO3 · CaC03 · 2H20) como un componente de los otolitos de los teleósteos. Carlström (1963) reporta vaterita como el componente del otolito de Acipenser sturio, concluyendo que la presencia de la vaterita pura en el laberinto de esta especie, indica, asimismo, la presencia de algún componente estabilizante. Degens (1969), analiza la estructura y composición de los otolitos de veinticinco especies, determinando las cantidades de aminoácidos y distribución de isótopos de oxígeno y carbono en la fracción carbonato. La materia orgánica estudiada por este autor corresponde a una proteína fibrosa de elevado peso molecular y bioquímicamente única, a la que denomina otolina. Pannella (1971 y 1974), relacionó las bandas de otolína con los incrementos de crecimiento diario, utilizándolas para determinar la edad de los peces con una frecuencia menor a la estacional. Rannou y Thiriot (1975), encuentran resultados semejantes en otolitos de la especie abisal Coryphaenoídes guentheri, demostrando que en esta especie abisal están presentes las mismas marcas de crecimiento diario que en las especies de zonas fóticas. Brothers et al. (1976), realizan un estudio de los otolitos de diferentes especies, en adultos y larvas, llegando a la conclusión de que los incrementos de crecimiento diario pueden utilizarse para determinar la edad de los peces durante la etapa larvaria y adulta, tanto si son de aguas tropicales como frías. Struhsaker y Uchiyama (1976), analizan los otolitos de larvas y adultos de la anchoveta, Stolephorus purpureus, determinando la curva de crecimiento de esta especie en los primeros seis meses de vida. Estos ritmos diarios que se presentan en la mayoría de las especies, se traducen a nivel de marcas anuales, como bandas hialinas y opacas, empleadas en la determinación de la edad. MATERIAL Y MÉTODOSLos ejemplares de la brótola, Phycis blennoídes, se obtuvieron frente a las costas de Cataluña entre Blanes y Vilanova i la Geltrú en profundidades que oscilaron entre 150 a 650m, durante el período de septiembre de 1976 a septiembre de 1978. La extracción de los otolitos, "sagitta", se efectuó levantando el basioccipital mediante un corte de tijera por su parte posterior, con lo que quedaban al descubierto las cápsulas óticas. Los otolitos fueron limpiados con agua y guardados en seco en sobres de papel, en donde constaba el número del ejemplar y la fecha de su captura. Para poder estudiar la composición cristalográfica de la "sagitta" de Phycis blennoides, se empleó un difractómetro de polvo de rayos X marca Philips, PW 1001, perteneciente al Departamento de Cristalografía y Mineralogía de la Universidad Central de Barcelona. Para el estudio de las marcas de crecimiento de ritmo inferior al anual presentes en las "sagittas" de Phycis blennoides, se empleó un microscopio electrónico de barrido, marca Stereoscan 180, Cambridge Instruments, perteneciente al Instituto de Investigaciones Pesqueras de Barcelona. RESULTADOS Y DISCUSIÓN
ULTRAESTRUCTURA
Composición CristalográficaEn la figura 1, se muestra el diagrama de difracción obtenido en el presente trabajo. Los ángulos de incidencia (2θ) aparecen al pie del diagrama, con valores de 20 a 69. En el resto de la figura se observan los picos correspondientes a las intensidades ópticas (I) obtenidos para cada ángulo. En la Tabla 1, se muestran los valores de los ángulos de incidencia, a partir de los cuales se obtuvieron los valores de las distancias del retículo de amstrong (dA) por medio de la ley de Bragg. A continuación se compararon los valores obtenidos en los análisis, con los correspondientes a las fichas características de otras sustancias que aparecen en el Joint Committee on Powder Difraction Standars, consultándose los referentes a la calcita y a la aragonita. Siendo esta última (Tabla 2, ficha 5-0453 del JCPDS) la que muestra valores de dA (distancia del retículo en amstrongs) muy similares a los obtenidos en el presente estudio. 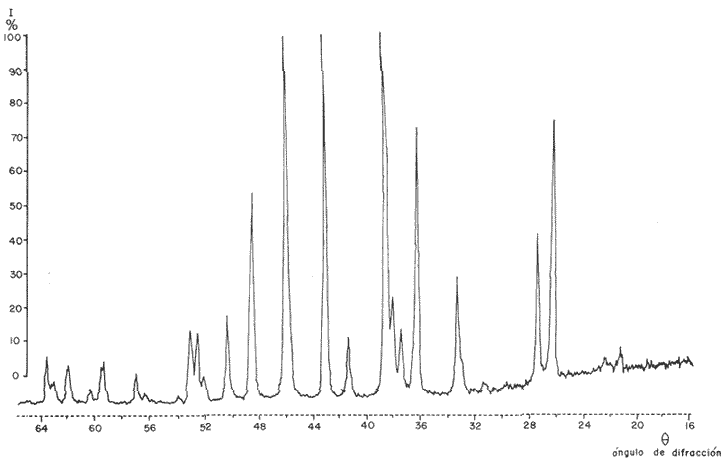 Fig. 1. Diagrama de difracción obtenido del análisis cristalográfico del otolito de Phycis blennoides. En la Tabla 1, se observa que en los resultados obtenidos, aparecen dos valores de dA: 3.969 y 1.6182 correspondientes a los ángulos de incidencia 22.40 y 56.90, respectivamente, que no a aprecen en la argonita en estado puro (Tabla 2). Estos valores pueden corresponder a ciertas sustancias impuras presentes en los otolitos de Phycis blennoides. Para esclarecer el significado de estos picos de escasa intensidad, se consultaron las fichas de JCPDS, correspondientes a la shortita y a la pirssonita, pero en ninguno de los casos se observó alguna correspondencia entre estos valores con los de las sustancias comparadas. Por lo que podrían atribuirse a alguna otra sustancia constituyente del otolito. De lo anterior podemos concluir que el material inorgánico presente en las "sagittas" de Phycis blennoides está constituido por Ca CO3, en forma de aragoníta con un alto grado de pureza. Los resultados obtenidos coinciden con los hallados por Immermann (1908), Brandenberger y Schintz (1945), Sasaki y Miyata (1955 y 1960) y CarIström (1963), en las "sagittas" de otros teleósteos marinos. No se registró la presencia de carbonato de calcio en forma de calcita, reportada por Devereux (1967) como componente de los otolitos de los teleósteos marinos. Tampoco se registró la presencia de cristales de shortita y pirssonita, citadas por Morris y Kittleman (1967), como probables componentes del otolito de los teleósteos. Esto puede ser debido a las bajas concentraciones de Na en las "sagittas" (1-2 % en un mol de calcio) que impiden detectar estas sustancias por las técnicas de difracción de rayos X. 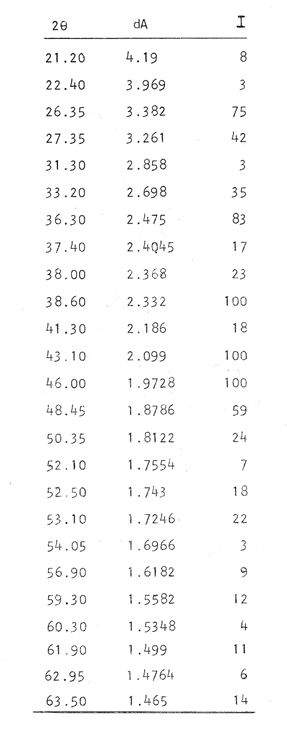 TABLA 1 VALORES OBTENIDOS EN EL ANÁLISIS CRISTALOGRÁFI DEL OTOLITO DE PHYCIS BLENIVOIDES Incrementos de CrecimientoEl otolito está formado por una matriz protéica mineralizada, compuesta por proteína fibrosa de elevado peso molecular y por carbonato cálcico cristalizado en forma de aragonita. La proteína ha sido denominada otolina por Degens (1969). Su peso molecular es superior a 150 000 y está formada principalmente por grandes concentraciones de ácido glutámico y aspartico, cistina e hidroxiprolina y pequeñas cantídades de aminoácidos básicos y aromáticos. El contenido total de la materia orgánica en los otolitos de las diferentes especies varía entre 0.2 a 10% del total, siendo de mayor concentración en aquellas especies cuyos otolitos son de menor tamaño y grosor (Degens, 1969). La otolina se dispone formando una matriz, con fibras radiales y concéntricas. En las figuras 2, 3 y 4 puede observarse la sección de una "saggita" de Phycis blennoídes, perpendicular al núcleo, mostrando las fibras radiales orientadas del núcleo al margen del otolito y las fibras concéntricas dispuestas en torno al núcleo. Estas últimas son de dos tipos, unas más finas y otras más gruesas formadas por conjuntos de las primeras. La argonita se forma por crecimiento epitaxial sobre la matriz protéica, uniéndose a ésta por puentes de hidrógeno. En las figuras 5, 6, 7 y 8, se muestran diversas fases de los cristales de aragonita, obtenidas a partir de la sección transversal de un otolito de Phycis blennoides. Los ejes de los cristales de aragonita están orientados perpendicularmente a la superficie de las bandas concéntricas de proteína y se prolongan del centro al margen del otolito, sin ser interrumpidas en su crecimiento por las bandas de otolina. Cada cristal de argonita puede estar formado por miles de segmentos individuales de crecimiento. La interacción entre las fibras protéicas y los cristales de argonita, da a estos últimos una forma de intercrecimiento en zigzag (Fig. 6). Las bandas transversas a los cristales de argonita de las figuras 2, 3 y 4, tienen la apariencia de suturas, pero no interfieren la orientación de los segmentos individuales de crecimiento (Degens, 1969). Los cristales de argonita se forman entre las fibras radiales y se prolongan a través de las fibras concéntricas de otolina, desde el centro del otolito a la periferia (Hickling, 1931).  TABLA 2 VALORES DE dA (DISTANCIA DEL RETÍCULO EN AMSTRONGS) E I (INTENSIDAD ÓPTICA DE LA ARAGONITA EN ESTADO PURO. (FICHA CARACTERÍSTICA)  Figs. 2 y 3. Electromierofotografías (Scanning) de la sección longitudinal de una "sagitta" de Phycis blennoides. mostrando las bandas radiales y concéntricas de otolina, aumentado 100 y 137 veces respectivamente (superficie pulida). Las bandas de otolina han sido consideradas por Kelly (1961), Dannevig (1956) y Degens (1969), como marcas de crecimiento que corresponden a períodos estacionales por medio de los cuales se puede llegar a determinar la edad de los peces. Pannella (1971), realiza un estudio sobre el crecimiento de Merluccius bilinearis, Gadus morhua y Urophycis chuss, tratando de encontrar el significado de la formación de las bandas en relación a un período menor al estacional. Dicho autor encuentra que en el otolito se depositan cada día, una capa de proteína y otra de argonita, cuya periodicidad diaria responde a un ritmo metabólico interno del pez. Resultados semejantes han sido observados por Rannou y Thiriot (1975), en especies abisales. 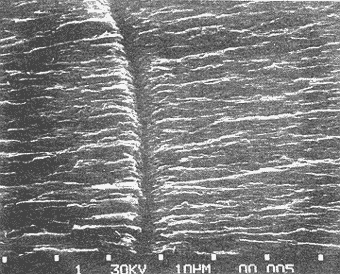 Fig. 4. Electromicrofotografía (Scanning) de la sección longitudinal de una "sagitta" de Phycis blennoides, mostrando una marca de detención del crecimiento y los cristales de argonita desmineralizados, aumentado 1 640 veces (superficie pulida). Estas bandas empiezan a formarse al eclosionar la larva y al absorberse el saco vitelino (Brothers, 1976; Struhsaker, 1976), pudiéndose determinar la edad de las formas larvarias y juveniles con exactitud. Los distintos tipos de bandas corresponden a dos períodos diferentes de producción del material fibroso; durante los períodos de crecimiento rápido se producen incrementos de crecimiento diario más finos, densos y numerosos que los del período de crecimiento lento. La formación diaria de estas bandas se repite muchas veces durante la formación de los anillos estacionales. Durante el período de formación de bandas de crecimiento rápido tiene lugar una mayor producción de fibras orgánicas y cristales de argonita que en el de crecimiento lento, siendo mayor la producción de argonita (un 90 % del total) en relación a las bandas de otolina. En el período de formación de bandas de crecimiento lento, la producción de bandas de otolina y argonita es menor que en la fase anterior,  Figs. 5 y 6. Electromicrofotografía (Scarming) de la sección transversal de una sagitta" de Phyc blennoides, mostrando los cristal de argonita, aumentado 1 000 y 1 870 veces, respectivamente. La razón del grosor de las bandas de crecimiento rápido y lento es de 1:20 en la etapa juvenil y de 1:3 en la adulta. En las figuras 9, 10 y 11 se aprecia la sección de una "sagitta" de Phycis blennoides, cercana al núcleo, mostrando los incrementos de crecimiento diario. Obsérvense las capas de otolina en color oscuro y los cristales de argonita en un tono más claro. De acuerdo con Pannella (1971), en Merluccius bilinearis, Gadus morhua y Urophycis chuss, las bandas de otolina llegan a alcanzar entre 0.5 y 2.5 µ de grosor y las bandas de argonita entre 0.5 y 1.0 . La amplitud de estas bandas puede variar entre las distintas especies.  Fig. 7. Electromicrofotografía (Scanning) de la sección transversal de una "sagitta" de Phycis blennoides, mostrando los cristales de argonita, aumentando 3 200 veces.  Fig. 8. Electromicrofotografía (Scanning) de la sección transversal de una "sagitta" de Phycis blennoides, mostrando los cristales de aragonita desmineralizados, aumentado 3 600 veces (superficie pulida) En la figura 11, se denotan las zonas de crecimiento lento (borde inferior derecho de la figura, en un tono más claro) y rápido (resto de la figura, tono más oscuro). En esta última los incrementos de crecimiento diario son más anchos y posiblemente con fibras de otolina más gruesas y cristales de aragonita más grandes, en relación a los incrementos de las zonas de crecimiento lento. 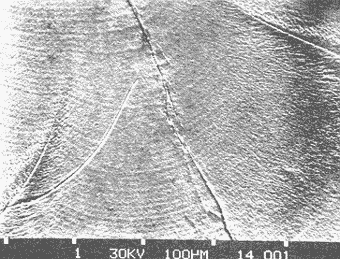 Fig. 9. Electromicrofotografía (Scanning) de la sección longitudinal de una "sagitta" de Phycis blennoides, mostrando los incrementos de crecimiento diario, aumentado 218 veces.  Figs. 10 y 11. Electromicrofotografía (Scanning) de la sección longitudinal de una "sagitta" de Phycis blennoides, mostrando los incrementos de crecimiento diario, aumentado 360 y 600 veces respectivamente ConclusionesLa "sagitta" está constituida por una matriz protéica, que forma fibras radiales y concéntricas en torno al núcleo, y por material inórganico correspondiente a carbonato cálcico en forma de aragonita con un alto grado de pureza. Los ejes de los cristales de aragonita están orientados perpendicularmente a la superficie de las bandas protéicas, prolongándose del centro al margen del otolito sin ser interrumpidas en su crecimiento por las bandas de proteína. La interacción entre las fibras protéicas y los cristales de aragonita producen en éstos una forma de crecimiento en zig-zag. No se registró la presencia de carbonato cálcico en forma de calcita, ni cristales de shortita ni de pirssonita, citados por otros autores como probables componentes del otolito de los teleósteos. Los incrementos de crecimiento diario están representados por dos finas bandas, cuyo grosor oscila entre 0.5 y 2.5 µ, correspondientes a una capa con mayor proporción de proteína y otra con predominio de aragonita. Las zonas de crecimiento rápido presentan incrementos de crecimiento diario más anchos con fibras protéicas más gruesas y cristales de aragonita más grandes que las de crecimiento lento. AgradecimientosA Ramón Margalef, catedrático de ecología de la Universidad Central de Barcelona, por la revisión del presente trabajo. A Luisa Cros y a Conchita Allué, miembros del Instituto de Investigaciones Pesqueras de Barcelona y a José Chinchón del Departamento de Cristalografía y Mineralogía de la Universidad Central de Barcelona por su cooperación en la realización del mismo. A Alfredo Laguarda-Figueras, investigador del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, por su apoyo para la publicación del presente trabajo. LITERATURABRANDENBERGER, E. y H. R. SCHINTZ Uber die Natur der Verkalkungen bei Mensh und das Verhalten der anorganischen Knochensubstanz im Falle der Hauptsáchlichen menschlichen Knochenkrankheiten. Helv. Med. Acta. Suppl. 1945 1-63 16 BROTHERS, F., S. MATHEWS y R. LASKER Fish. Bull. Daily growths increments in otoliths from larval and adult fishes. 1976 70-75 74 (1) CARLSTROM, D. Biol. Bull. Mar, Biol. Lab. A crystallographic study of vertebrate otoliths. Woods Hole 1963 441-463 125 DANNEVIG, E. H. Chemical composition of the zones in cod otholiths. J. Cons. Int. Explor. Mer. 1956 156-159 21 DEGENS, E. T., W. G. DEUSER y R. L. HAEDRICH Mar. Biol. Molecular structure and composition of fish otoliths. 1969 105-113 2 (2) DEVEREUX, I. Science Temperature measurements from oxygen isotope ratios of fish otoliths. 1967 1684-1685 1955 HICKLING, C. F. The structure of the otolith of the hake. Q. Jl. Microsc. Sci. 1931 547-561 74 IMMERMANN, F. Die innere Struktur der SchollenOtolithen. Wiss. Meeresunters. Abt. Helgoland. N. F. 1908 129-176 8 (6) IRIE, T. The crystal texture of the otolith of a marine Teleost Pseudoscia nena. J. Fac. Físh. Anim. Husb. Hiroshima Univ. 1955 1-14 1 IRIE, T. The growth of the fish otolith J. Fac. Fish Anim. Husb. Hiroshima Univ. 1960 202-229 3 KELLY, G. F. y A. M. BARKER Observations on the behaviour, growth and migration of redfish at Eastport Maine. Sp. Publ. ICNAF 1961 263-275 3 MAIER, H. N. Die Altersbestimmung noch den Otolithen bei Scholle und Kabeljau. Wis Meeresunters. Abt. Helgoland N. F. 1908 56-115 8 MORRIS, R. W. y L. R. KITTLEMAN Science Piezoelectric property of otoliths. 1967 368-370 158 PANNELLA, G. Science Otoliths dayly growth layers and periodical patterns. 1971 1124-1127 173 PANNELLA, G. Ageíng of Fish. Otolith growth patterns: An aid in age determination in temperate and tropical fihes. T. B. Bagenal Unwin Brothers Ltd. Surrey 1974 28-39 RANNOU, M. y C. THIRIOT-QUIEVREUX Ann. Inst. Ocean. Structure des otolithes d'un Macrouridae bathyal. Etude au mieroscope electronique a balayage. 1975 195-201 51 (2) SASAKI, H. y J. MIYATA Experimentelle Studien Uber Otolithen. Zeitschr. Laryngol. Rhinol. Otol. 1955 740-748 34 STRUHSAKER, P. y J. L. UCHIYAMA Fish. Bull. Age and growth of the melin, Stolephons purpureus (Pisces; Engraulidae) from the Hawaiian islands as indicated by dayly growth increments of sagittas. 1976 9-17 74 (1)
|