|
ANÁLISIS DE LOS HÁBITOS ALIMENTICIOS DE LA BRÓTOLA
PHYCIS BLENNOIDES (BRUNNICH 1768) EN EL MEDITERRÁNEO OCCIDENTAL (PISCES:
GADIDAE)
Trabajo recibido el 16 de julio de 1984 y aceptado para su
publicación el 12 de noviembre de 1984.
MANUEL GALLARDO-CABELLO
Universidad Nacional Autónoma de México. Instituto
de Ciencias del Mar y Limnología. Contribución 479 del Instituto
de Ciencias del Mar y Limnología, UNAM.
En el presente
trabajo se analizaron las variaciones en el crecimiento de las diferentes
partes del encéfalo y de los órganos de los sentidos de
Phycis blennoides, asociados con cambios en su habitat
(migraciones verticales) realizadas a través de su cielo de vida. Se
obtuvieron los valores del índice alométrico k
para las distintas partes del encéfalo en relación a la longitud
total del pez, siendo de: 6.78 x 10-³ para el
telencéfalo; 5.32 x 10-³ para el
mesencéfalo; 15.87 x 10-³ para el
metencéfalo y 3.94 x 10-³ para el
mielencéfalo. El número de laminillas de la estructura en roseta
de esta especie es de 26 a 28. Durante las edades 0 a 2 años la
percepción visual tiene mayor importancia que la olfativa, apareciendo
presas pelágicas o bentopelágicas en el contenido estomacal. A
los tres años, el telencéfalo presenta un mayor desarrollo que
los lóbulos ópticos y la alimentación está
constituida por organismos que viven en los fondos. La relación
alométrica existente entre la longitud total del pez y la longitud de
las aletas pélvicas es de y = 0. 109379 x
1.216. La tasa de crecimiento de las aletas
pélvicas se in crementa en proporción directa a la edad y a la
distribución batimétrica de esta especie.
This paper
deals with data about the growth of the brain and the sense organs associated
with life cycle's changes (vertical migrations) of the greater forkbeard
Phycis blennoides. Data about allometric indexes
(k) of the length in different parts of the brain in
relation to total length of fish was obtained as follows:6.78 x
10-³ for the telencephalon, 5.32 x
10-³ for the mesencephalon, 15.87 x
10-³ for the metencephalon and 3.94 x
10-³ for the myelencephaIon. The fo1ds number
of the rosette structure is 26 to 28 for this species. During ages 0 to 2 years
the visual perception is more important than the olfactory sense, as is clear
by the presence of pelagic or benthopelagic preys in the stomacal content. At 3
years the telencephalon is greater than the optic lobes and feeding is composed
by benthic organisms. The allometric relation between the total length of the
fish and the total length of the pervil fins is y = 0. 109379 x
1.216; the growth rate of the pelvic fins increases
in direct proportion to age and bathymetrie distribution of this
species.
El estudio de la morfología y desarrollo de los órganos de los sentidos en los peces, resulta de gran interés, ya que son sistemas que pueden alcanzar un alto grado de desarrollo y diversidad en relación al comportamiento alimenticio de muchas especies. La importancia de la recepción olfatoria, gustatoria y visual en los hábitos alimenticios de los peces, ha sido estudiada por diversos autores como son: Baglioni, 1909; Burden, 1945; Da Franca, 1952; Hasler, 1950, 1957; Healy, 1957 y Olmsted, 1918; entre otros. MATERIAL Y MÉTODOSLos datos biológicos fueron obtenidos de ejemplares de Phycis blennoides, procedentes de los muestreos biológicos y de la captura comercial, de septiembre de 1976 a septiembre de 1978. Los ejemplares se obtuvieron en el Mediterráneo Occidental frente a las costas de Cataluña, entre Blanes y Vilanova i la Geltrú, en profundidades que oscilaron entre 150 y 650 m. Los muestreos de la captura comercial se efectuaron varias veces a la semana y una vez al mes se realizó un muestreo biológico, efectuando arrastres a distintas profundidades: el primero entre 150 y 300 m, el segundo entre 300 y 400 m y el tercero entre 400 y 600 m, utilizando para ello una red comercial (bou) con un copo de 35 mm y un sobrecopo de 10 mm de abertura de malla. Los datos obtenidos fueron los siguientes: 1) Longitudes de las diversas partes del encéfalo: telencéfalo, mesencéfalo, metencéfalo y mielencéfalo de un total de 250 ejemplares. 2) Longitud total de las aletas pélvicas de 1,000 ejemplares. 3) Dimensiones de los sacos nasales y conteo del número de laminillas de la estructura en roseta, de 56 ejemplares. Para analizar y comparar la importancia de la recepción olfatoria con respecto a la recepción visual, en los hábitos alimentarios de Phycis blennoides, se calculó el coeficiente ecológico propuesto por Teichmann (1954). Los grupos de edad fueron establecidos para esta especie por Gallardo-Cabello (1979) y los datos reales para el análisis del contenido estomacal fueron proporcionados por E. Macpherson. RESULTADOS Y DISCUSIÓN
RECEPCIÓN OLFATORIA
Desarrollo del TelencéfaloComo en la mayoría de los peces que dependen de su olfato para su alimentación, en la brótola Phycis blennoides, los lóbulos se presentan bien desarrollados y ocupan una importante proporción en comparación con el resto del cerebro (Fig. 1). Los bulbos olfatorios son pedunculados, como en el resto de los gadiformes, lo que constituye una importante característica filogenética dentro de la clase Pisces (Devitsyna, 1972). La recta de regresión obtenida entre la longitud total del pez y la longitud del telencéfalo (Fig. 2), viene expresada por la siguiente ecuación: y = 6.78 x 10-³ x + 2.86 donde x es la longitud total de pez y y la longitud total del telencéfalo. Los valores de la longitud del telencéfalo para cada clase de tallas y grupos de edad, se muestran en las Tablas 1 y 2. Con respecto a la edad (Fig. 3) se observa una disminución en la tasa del crecimiento del telencéfalo, conforme el pez envejece. La longitud total del telencéfalo se incrementa 0.35 mm entre las edades 0 y 1 año; y 0.35 mm entre los años 1 y 2. A partir de esta edad, la velocidad de crecimiento disminuye, con un incremento promedio de talla de 0.23 a 0.20 mm, por cada año. Sacos NasalesLos sacos olfatorios de la brótola descansan en la parte dorsal anterior de la cabeza, oralmente a las órbitas. Los orificios nasales anteriores y posteriores, están bien separados y se abren en el techo epidérmico del saco olfatorio. Su forma es oval. El diámetro de dichos orificios se incremeta conforme aumenta la edad del pez. En la Tabla 3, se muestra la longitud de los orificios nasales anterior y posterior, a través de los grupos de edad. El orificio anterior, muestra un diámetro mayor que el posterior y presenta un apéndice en la parte exterior- posterior, que facilita la entrada del agua en el saco olfatorio. 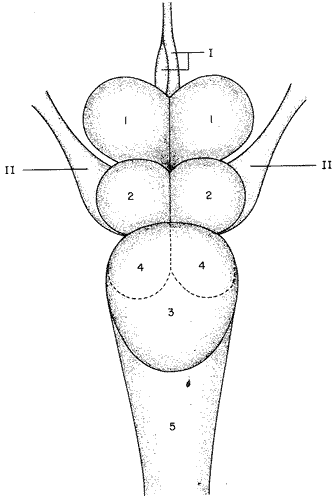 Fig. 1. Organización del encéfalo de Phycis blennoides. (1, Tecencéfalo; 2, Mesencéfalo; 3, Macrocéfalo; 4, Mielencéfalo; 5, Médula espinal. Pares craneales: I: olfatorios; II ópticos). Los orificios nasales anteriores, conducen directamente a los sacos nasales, que en la brótola son el aspecto abombado. Al igual que en otras especies, presentan una pared membranosa, cubierta por una capa mucosa, que segrega el mucus. El tamaño de estos sacos varía a través de la edad, como puede verse en la Tabla 4. Los sacos olfatorios de Phycis blennoides son de menor tamaño en comparación con otros miembros de la familia Gadidae. En el bacalao, Gadus morhua, alcanzan una longitud y anchura de 5.5 x 4.0 mm, en ejemplares de 265 mm de longitud total. En la navaga, Elegínus navaga, llegan a medir 6.0 mm de longitud por 4.0 mm de ancho, en ejemplares de 310 mm; y en Lota lota, 6.5 x 4.0 mm, en ejemplares de 500 mm. 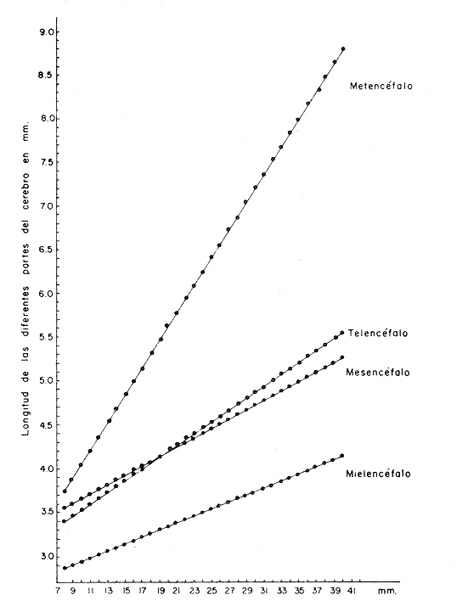 Fig. 2. Relación entre la longitud total del pez y, las longitudes de las diferentes partes del cerebro de Phycis blennoídes. Placas NasalesEn el fondo de los sacos nasales, la pared membranosa se pliega, formando una estructura laminar, donde se ordenan el epitelio olfatorio y las terminaciones nerviosas (Fig. 4). Estas estructuras se conocen con el nombre de placas olfatorias o rosetas. El número de laminillas de las placas olfatorias, es un carácter taxonómico para cada especie (Devitsyna, 1972). La roseta de Phycis blennoides, es de forma alargada, en dirección antero-posterior, presenta de 26 a 28 laminillas, que se disponen paralelamente a cada lado del septo central. La forma de las laminillas varía en dirección rostrocaudal, siendo las primeras de forma más irregular. Las laminillas incrementan su área de recepción, tanto en longitud como en anchura, conforme el pez envejece, pero no se observó que aumentara el número de éstas a través de los grupos de edad. 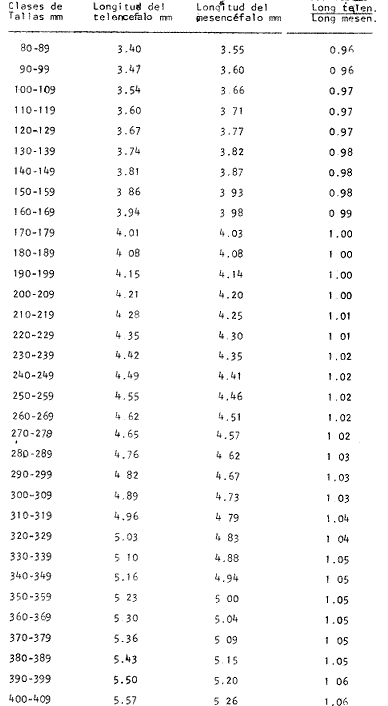 TABLA 1 RELACIÓN LONGITUD TOTAL DEL PEZ -LONGITUD DEL TELENCÉFALO-LONGITUD DEL MESENCÉFALO Las laminillas olfatorias muestran una gran vascularización y cada una de ellas es recorrida por capilares de la arteria facial. A diferencia de otros Gádidos las laminillas no presentan pigmentos, sin embargo, éstos se presentan en el tejido conectivo adyacente a la roseta. 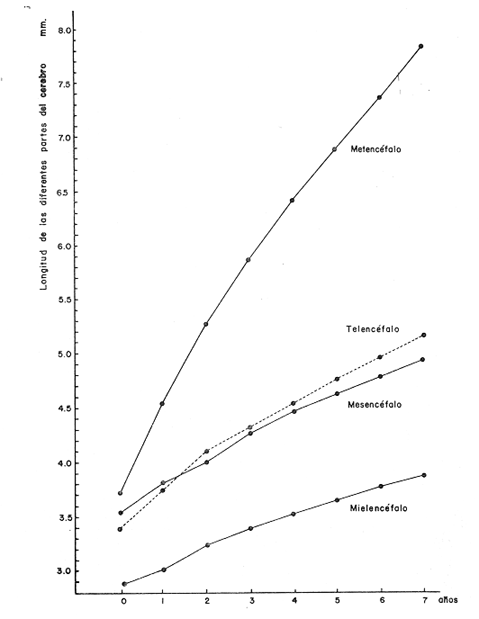 Fig. 3. Relación edad-longitud de las diferentes partes del cerebro de Phycis blennoides. RECEPCIÓN VISUAL
Desarrollo del MesencéfaloLa recta de regresión obtenida entre la longitud total del pez y la longitud del mesencéfalo (Fig. 2), está expresada por la siguiente ecuación: y = 5.32 x 10-³ x + 3.18 donde x es la longitud total del pez, y y la longitud del mesencéfalo; Los valores de la longitud del mesencéfalo para cada clase de tallas y grupos de edad se muestran en las Tablas 1 y 2. 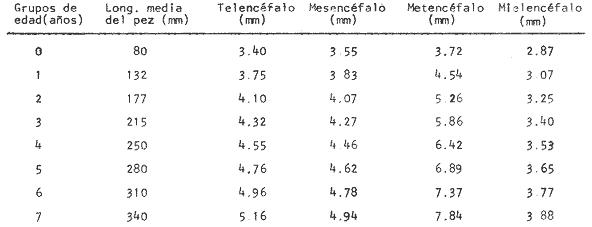 TABLA 2 RELACIÓN EDAD-LONGITUD DE LAS DIFERENTES PARTES DEL ENCÉFALO 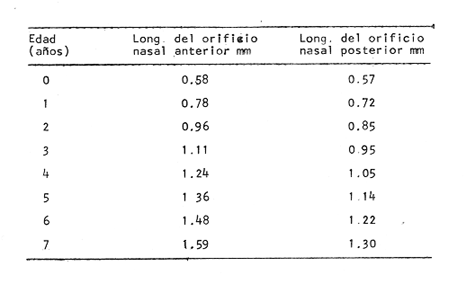 TABLA 3 RELACIÓN EDAD-LONGITUD DE LOS ORIFICIOS NASALES, ANTERIOR Y POSTERIOR DE PHYCIS BLENNOIDES 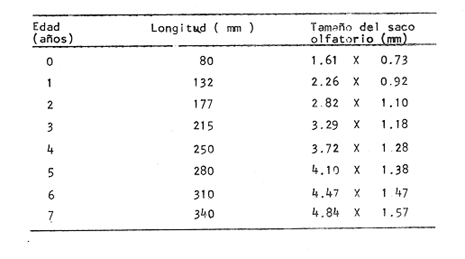 TABLA 4 RELACIÓN EDAD-TAMAÑO DE LOS SACOS OLFATORIOS DE PHYCIS BLENNOIDES Con respecto a la edad (Fig. 3), se observa que los incrementos en la longitud del mesencéfalo descienden conforme el pez envejece. La longitud total del mesencéfalo se incrementa 0.28 mm entre las edades 0 y 1 años; 0.24 mm entre 1 y 2 años; 0. 20 mm entre 2 y 3 años; 0. 19 mm entre 3 y 4 años. A partir de esta edad, la velocidad del crecimiento disminuye con un incremento medio de talla de 0.16 mm. En las Tablas 1 y 5 se muestran los valores del coeficiente de Teichmann, obtenidos para clases de tallas y grupos de edad. Como puede observarse en dichas tablas, y en las figuras 2 y 3, la recepción visual muestra una mayor importancia en relación al olfato, durante las primeras etapas de vida de Phycis blennoides, o sea, durante las edades 0 a 1 años. A los dos años de edad, el desarrollo de los lóbulos olfatorios y ópticos, es muy similar, presentando una longitud muy semejante. A partir de los tres años de edad (215 mm de longitud total) se presenta un mayor desarrollo del telencéfalo en relación al mesencáfalo. Se pueden distinguir dos períodos importantes en el cielo de vida de Phycis blennoides, con respecto al desarrollo de la vista y el olfato y relacionarlos con sus hábitos alimenticios: a) Primer período (0 a 2 años). El sentido de la vista se encuentra más desarrollado que el olfato y probablemente presenta una mayor importancia en la detección de las presas. Durante este período, Phycis blennoides, se alimenta de presas pelágicas o bentopelágicas, como son: Copépodos (Aetideopsis sp), Amphypodos (Vibilia armata), Misidáceos y Gammaroideos, que no se presentan en los individuos de mayor edad. De acuerdo con lo propuesto por Teichmann, durante este período, Phycis blennoides, se comporta como un depredador mierosomático. 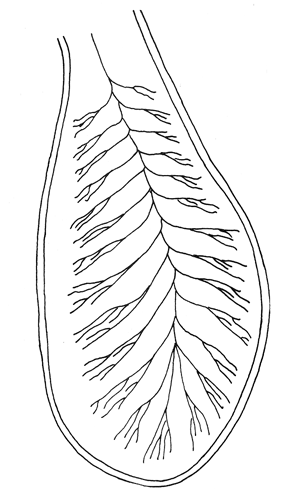 Fig. 4. Placa olfatoria o estructura en roseta de Phycis blennoides. 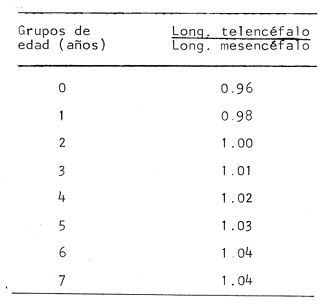 TABLA 5 ÍNDICE DE TEICHMANN PARA CADA GRUPO DE EDAD b) Segundo período (3 años en adelante). El telencéfalo presenta un mayor desarrollo que los lóbulos ópticos. Incrementándose esta diferencia conforme aumenta la edad. En este período, la alimentación de Phycis blennoides, está sólo constituida por organismos que viven en los fondos, como son: Alpheus glaber, Calocaris mecandreae, Goneplax rhomboides y otros, los cuales serán más fácilmente detectados por medio del olfato y del gusto, que por la vista. La presencia de Engraulis encrasicholus en el contenido estomacal de los individuos de gran talla, puede sugerir que éstos o sus restos son capturados cerca del fondo. En la Tabla 6, aparecen los valores del índice de Teichmann para diversas especies de la familia Gadidae. En Phycis blennoides y Lota lota, el coeficiente ecológico, indica la predominancia de la recepción olfatoria sobre la recepción visual en los hábitos alimenticios de estas dos especies. Siendo una característica de la subfamilia Lotinae a la cual pertenecen la brótola y la lota. Mientras que en Gadus morhua y Eleginus navaga (subfamilia Gadinae) el receptor visual se presenta mejor desarrollado que el olfatorio. Este mismo fenómeno ocurre en Merluccius merluccius y M. senegalensis. En la Tabla 7, se muestra el número de laminillas de la estructura en roseta para los diferentes miembros de la familia Gadidae. La brótola presenta un número de laminillas muy siriffiar a Gadus morhua y Merluccius merluccius. Un número superior a Eleginus navaga y un número inferior a Lota lota. En Merluccius merluccius y M. senegalesis, Geiger (1954), encuentra que el número de laminillas se incrementa a través de los grupos de edad. Fenómeno que no ocurre con otras especies de Gádidos (Devitsyna, 1972 y GallardoCabello, presente trabajo). Por otra parte, Phycis blennoides, presenta los orificios nasales anteriores, provistos de una estructura dérmica que regula el flujo del agua a través de los orificios nasales, lo cual está evidentemente relacionado con el gran desarrollo de la sensibilidad olfatoria.  TABLA 6 ÍNDICE DE TEICHMANN EN DIVERSAS ESPECIES DE LA FAMILIA GADIDAE  TABLA 7 NÚMERO DE LAMINILLAS DE LA ESTRUCTURA EN ROSETA DE DIFERENTES ESPECIES DE LA FAMILIA GADIDAE Ha sido establecido por medio de métodos electrofisiológicos (Bondreau, 1962), que un incremento en el rango del flujo a través de los nostrilos, o cualquier cambio en la dirección del flujo del agua, conduce a una intensificación de la respuesta a los estímulos olfatorios. Por lo que el número de laminillas de la roseta y la estructura de los orificios nasales de Phycis blennoides y Lota lota, incrementan la reacción a los estímulos del olfato. Mientras que Gadus morhua y Eleginus navaga presentan receptores olfatorios menos desarrollados. El desarrollo de la recepción olfatoria y visual está estrechamente relacionado a las características ecológicas y hábitos alimenticios de estas especies. Mientras que Lota lota, es un depredador solitario, que se alimenta durante la noche o a bajas iluminaciones, Gadus morhua y Eleginus navaga, son especies capaces de formar cardúmenes y se alimentan durante el día (Sonina, 1957). Es muy posible que Phycis blennoides, a partir de los tres años de edad presente un comportamiento muy semejante a Lota lota, es decir, que se alimenta durante la noche y no llega a formar grandes agregaciones, fenómeno que también ocurre en Phycis Phycis. LOCOMOCIÓN
Desarrollo del MesencéfaloEl metencéfalo se presenta bien desarrollado, y como en algunas otras especies es la parte que ocupa la mayor proporción en tamaño del cerebro. En la figura 2, aparece la recta de regresión entre la longitud total del pez y la longitud del metencéfalo. La línea de regresión está representada por la siguiente ecuación: y = 15.87 x 10-³ x + 2.45 donde x es la longitud del pez, y y la longitud del metencéfalo. En la Tabla 8, se muestran los valores de la longitud del metencéfalo para cada clase de tallas. 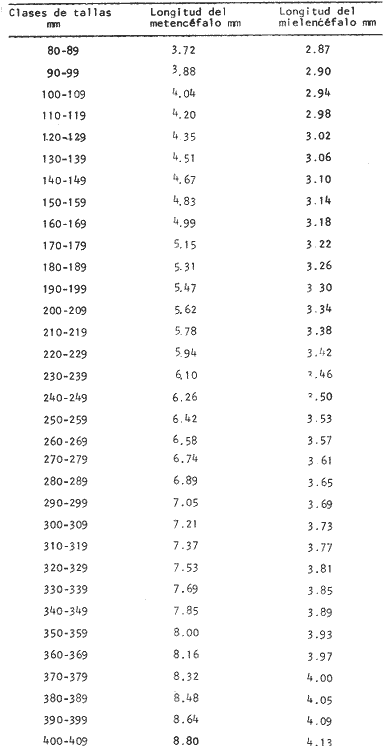 TABLA 8 RELACIÓN LONGITUD TOTAL DEL PEZ-LONGITUD DEL METENCÉFALO - LONGITUD DEL MIELENCÉFALO El valor del índice alométrico k = 15.87 x 10-³, es mayor que los valores de los índices obtenidos para las demás partes del cerebro (Tabla 9). Lo que indica la importancia de este órgano durante el desarrollo del cielo de vida de Phycis blennoides.  TABLA 9 VALORES DEL ÏNDICE ALOMÉTRICO K PARA DISTINTAS PARTES DEL ENCÉFALO Con respecto a la edad (Fig. 3, Tabla 2), se observa que los incrementos en la longitud del metencéfalo descienden conforme el pez envejece. Durante las edades 0 a 1 años, la tasa de crecimiento es mayor, desciende en las edades 2 y 3, y a partir de los 4 años la tasa de crecimiento decrece y se mantiene constante. El desarrollo del cerebelo de Phycis blennoides, puede estar relacionado con las experiencias realizadas por Franz (1911, 1912 y 1920); y Tilney (1923), quienes establecen una estrecha relación entre el desarrollo del cerebelo y la actividad locomotora del pez. Dichos autores encontraron que el metencéfalo se encuentra poco desarrollado en peces que presentan una escasa locomoción, como son: Hippocampus, Lophius y los peces planos. Mientras que en aquellos peces que son buenos nadadores, se presenta bien desarrollado, como es el caso: Thunus y Scomber. Por lo que sería posible, que en la brótola, el gran tamaño del cerebelo estuviese relacionado con una mayor locomoción en relación a otras especies que ocupan su mismo habitat, como son: Lophius piscatorius y Lepidorhombus boscii. Y que represente, una ventaja adaptativa en las relaciones de competencia con esta última especie. Por otra parte, Herrick (1905) establece que algunas especies de peces presentan un cerebelo bien desarrollado, aunque su actividad locomotora sea escasa (Fam. Mormyridae) y que el desarrollo de este órgano está relacionado con la recepción de los estímulos químicos, debido a las conexiones que existen entre el cerebelo, y el par craneal VII. Fenómeno que puede presentarse en la brótola, debido al gran desarrollo del sentido del gusto. RECEPCIÓN GUSTATORIA
Desarrollo del MielencéfaloEn Phycis blennoides, el mielencéfalo se presenta bien desarrollado. A diferencia de la mayoría de las especies, está constituido por dos lóbulos diferenciados que se localizan debajo de la parte media posterior de cerebelo. En la figura 2, aparece la recta de regresión entre la longitud total del pez y la longitud del mielencéfalo. La línea de regresión está representada por la siguiente ecuación: y = 3.94 x 10-³ x + 2.55 donde x es la longitud del pez, y y la longitud del mielencéfalo. Los valores de la longitud del mielencéfalo para cada clase de tallas, se muestran en la Tabla 8. Con respecto a la edad (Fig. 3, Tabla 2), se observa una disminución en la tasa de crecimiento del mielencéfalo conforme aumenta la edad del pez. La longitud total se incrementa 0.20 mm entre las edades 0 y 1 años, 0.18 mm entre las edades 1 a 2 años, 0.15 mm entre los 2 y 3 años y 0.13 mm entre los 3 y 4 años. A partir de esta edad la velocidad de crecimiento disminuye, con un incremento medio de talla de 0.12 mm por año. El valor del índice alométrico 3.94 x 10-3, es el menor de los valores de los índices obtenidos para las demás partes del cerebro. Sin embargo, el hecho de que este centro se presente perfectamente bien desarrollado y diferenciado en Phycis blennoides, indica la gran importancia que tiene el sentido del gusto en los hábitos alimenticios de esta especie. Desarrollo de las aletas pélvicasEn Phycis blennoides, las aletas pélvicas se encuentran reducidas a dos radios libres enfundados conjuntamente en su parte superior, a nivel de su implantación. Son largas y filiformes, ocupan una importante proporción con respecto a la longitud total del pez, entre un 25.5 % y un 41.67 %, lo que indica la gran importancia de su función en relación a los hábitos alimenticios de esta especie. 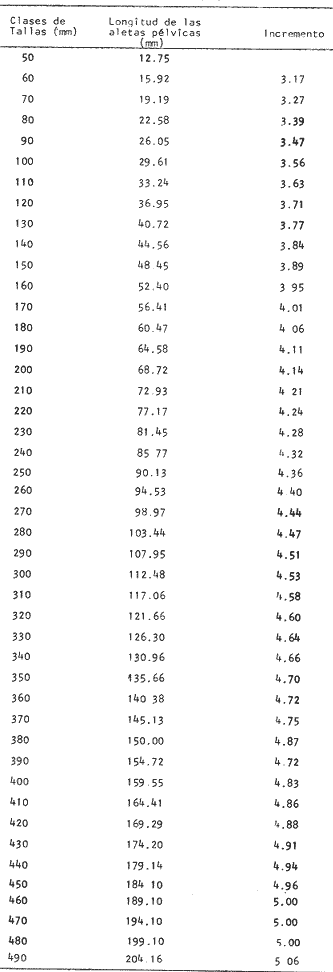 TABLA 10 RELACIÓN CLASES DE TALLAS - LONGITUD DE LAS ALETAS PÉLVICAS La relación alométrica existente entre estas dos series de datos, viene expresada por el valor del índice alométrico k = 1.216, se trata por lo tanto de una relación de crecimiento positiva. El índice de correlación es de 1.00. La fórmula encontrada, es la siguiente: y = 0. 109379 x 1.216 donde y representa la longitud de las aletas pélvicas y x la longitud total del pez. Los valores de la longitud de las aletas pélvicas para cada clase de tallas se muestran en la Tabla 10. En la figura 5, se observan dos puntos de inflexión en la línea de regresión, entre la longitud total de las aletas pélvicas y del pez, razón por la cual se han desdoblado las series de datos de tres partes, encontrándose el valor del exponente o índice alométrico para cada caso. En la Tabla 11, se muestran los resultados obtenidos. Las variaciones del valor de k a lo largo del ciclo de vida de Phycis blennoides, son cada vez mayores conforme aumenta la longitud del pez. Dichas variaciones pueden estar relacionadas con cambios de habitat (migraciones verticales), realizadas a lo largo de su cielo de vida. 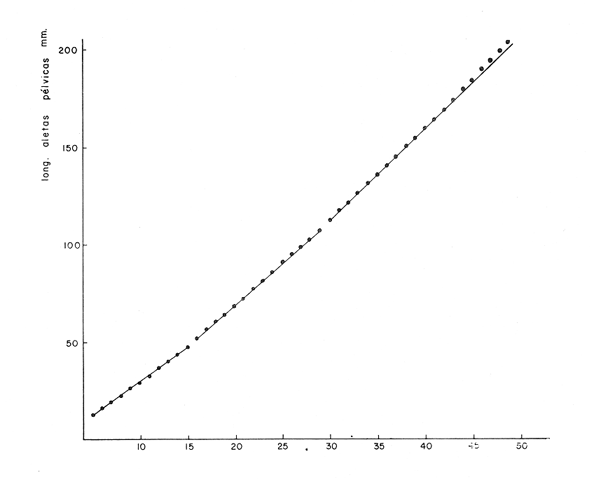 Fig. 5. Relación entre la longitud total del pez y la longitud total de las aletas pélvicas. El gran desarrollo y diferenciación que presentan el mielencéfalo y las aletas pélvicas de Phycis blennoides, indican la importancia que tiene el sentido del gusto en los hábitos alimenticios de esta especie. En la Tabla 10 y en la figura 5, se observa que conforme aumenta la talla o edad de esta especie, se incrementa la tasa de crecimiento de las aletas pélvicas, fenómeno que no se presenta durante el crecimiento de ningún otro órgano o centro de los estudiados en el presente trabajo.  TABLA 11 VALORES DEL ÍNDICE DE REGRESIÓN PARA DISTINTAS CLASES DE TALLAS Y LONGITUD DE LAS ALETAS PÉLVICAS DE PHYCIS BLENNOIDES Como ya se ha mencionado anteriormente, estos incrementos en la tasa de crecimiento de las aletas p é lvicas pueden estar relacionados con cambios de habitat (migraciones verticales) realizadas por esta especie, durante el transcurso de su cielo de vida. En la Tabla 12 y en la figura 6, se observa que conforme aumenta la talla, estos peces tienden a vivir a una mayor profundidad, localizandose las tallas mayores (300-490 mm de longitud total) sólo a profundidades superiores a los 400 m. Lo que puede indicar que conforme aumenta la talla, estos ejemplares viven a profundidades mayores y que las aletas pélvicas juegan cada vez un papel más importante en la detección de las presas, debido en gran parte a la disminución de la luz. Las presas de las que se alimenta Phycis blennoídes: Alpheus glaber, Calocaris mecandreae, Goneplax rhomboides y otras, viven enterradas en el fango, por lo que serán más fácilmente detectadas por medio del gusto y del tacto. 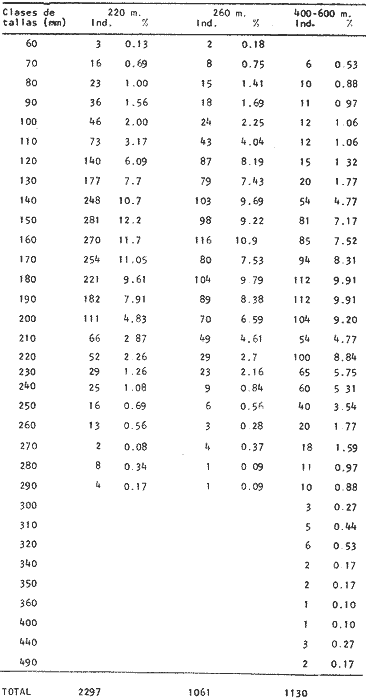 TABLA 12 DISTRIBUCIÓN DE LAS FRECUENCIAS DE TALLAS EN RELACIÓN A LA PROFUNDIDAD (DISTRIBUCIÓN BATIMÉTRICA) ConclusionesLa recta de regresión entre la longitud total del pez y la longitud del telencéfalo viene expresada por la ecuación: y = 6.78 x 10-³ x + 2.86 Los sacos olfatorios de Phycis blennoides son de menor tamaño en relación a otros miembros de la familia Gadidae, como son: Gadus marhua, Eleginus navaga y Lota lota. El número de laminillas de la estructura en roseta de esta especie es de 26 a 28. El desarrollo de esta estructura y de los orificios nasales favorecen la recepción de los estímulos olfatorios. 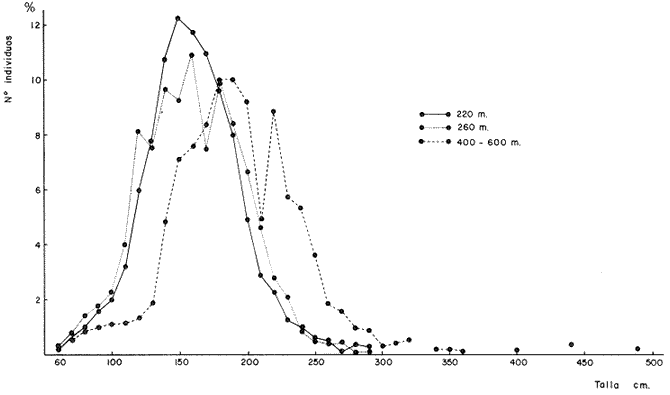 Fig. 6. Distribución batimétrica de Phycis blennoides La recta de regresión entre la longitud total del pez y la longitud del mesencéfalo está expresada por la ecuación: y = 5.32 x 10-³ x + 3.13 En esta especie se presentan importantes cambios en la morfología y desarrollo de las diferentes partes del encéfalo y órganos de los sentidos, en relación con variaciones en la dieta afimentaria. Durante las edades 0 a 2 años, la percepción visual tiene mayor importancia que la olfativa, apareciendo presas pelágicas o bentopelágicas en el contenido estomacal que no se presentan en los individuos de mayor edad. A partir de los tres años, el telencéfalo presenta un mayor desarrollo que los lóbulos ópticos, diferencia que se incrementa conforme aumenta la edad. En este período, la alimentación está constituida por organismos que viven en los fondos. La recta de regresión entre la longitud total del pez y la longitud del metencéfalo es la siguiente: y = 15.87 x 10-³ x + 2.45 Existe una estrecha relación entre el desarrollo y la diferenciación del mielencéfalo y de las aletas pélvicas, como centros de la recepción gustatoria y los hábitos alimenticios de esta especie. La recta de regresión entre la longitud del pez y la longitud del mielencéfalo es la siguiente: y = 3.94 x 10-³ x + 2.55 La relación alométrica existente entre la longitud total del pez y la longitud de las aletas pélvicas es la siguiente: y = 0. 109379 x 1.216 La tasa de crecimiento de las aletas pélvicas se incremeta en proporción directa a la edad y a la distribución batimétrica de esta especie. AgradecimientosA Ramón Margalef, catedrático de ecología de la Universidad Central de Barcelona, por la revisión del presente trabajo. A Conchita Allué, Pilar Sánchez y Enrique Maepherson, miembros del grupo de Recursos del Instituto de Investigaciones Pesqueras de Barcelona, por su coope ración en la realización del mismo. A Alfredo Laguarda-Figueras, investigador del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México por su apoyo y estímulo para la publicación del presente trabajo. LITERATURABAGLIONI, S. Atti. Soc. Ital. Progr. Sci. Alcune esperienze di fisíologia dei sensi negli animalI marini (o1fatto e senso tattile). 1909 1-399 2 BONDREAU, J. C. Japan J. Physiol. Electrical activity of the olfactory tract of the catfish. 1962 28-52 12 (3) BURDEN, W. D. U.S. Aír Surgeon's Bull. Development of a shark deterrent 1945 344-347 2 (10) DA FRANCA P. Notas e estudos Inst. Biol. Marít. Merluccius merluccius (L.) Merluccius senegalensis (Cadenat): seus caracteres distintivos. 1952 1-72 3 DEVITSYNA, G. V. J. Ichthyol. Morphology of the organs of olfaction in Gadidae. 1978 78-125 12 (6) FRANZ, V. Biol. Zentr. Uber das Kleinhirn in der vergleichenden Anatomie. 1911 434-445 31 FRANZ, V. Das KIeinhirn der Knochenfische. Zool. Jahrb. Abt. Anat. Ontog. 1912 401-464 32 FRANZ, V. Zur mikroskipíshen Anatomie der Mormyriden. Zool. Jahrb. Abt. Anat. Ontog. 1920 91-148 42 GALLARDO-CABELLO Características biológicas de Phycis blennoides (Brünnich, 1768), con especial referencia a algunas modificaciones debidas al efecto de la explotación pesquera. Universidad Central de Barcelona 1979 420 p. GEIGER, W. Bull. Inst. des pêches da Maroc Le curveau de Merluccius merluccius (L.) et de Merluccius senegalensis (Cadenat). Contribution a Pétude comparative des deux espéces. 1954 70-112 35 HASLER, A. D., y W. J. WISBY Use of fish for the olfactory essay of pollutants (phenols) in water. Trans. Am. Fish. Soc. 1950 64-70 79 HASLER, A. D. The Physiology of Fishes. The sense organs Olfactory and gustatory senses of fishes. Brown Academic Press Inc. Publ. New York 1957 187-209 2 HEALY, E. G. The Physiology of Fishes. The nervous system. Brown Academic Press Inc. Publ. New York 1957 1-119 2 HERRICK, C. J. The central gustatory paths in the brain of bony fishes. J. Comp. Neurol. 1905 375-456 15 OLMSTED, J. M. D. Experiments on the nature of the sense of smell in the common catfish Ameírus. Am. J. Physiol 1918 443-458 46 SHELDON, R. E. The reactions of the dogfish to chemical stimuli. J. Comp. Neurol. 1909 273-311 19 SHELDON, R. E. The sense of smell in Selachians. 1. Expt. Zool. 1911 51-62 10 SONINA, M. A. The White Sea Cod (Materials for a combined study of White Sea) Leningrad, Akad Nauk. SSSR 1957 68-83 1 TEICHMANN, H. Vergieichende Untersuchungen an der Nase der Fische. Z. Morphol. Oekol. Tiere 1954 171-212 43 TILNEY, F. Genesis of cerebellar functions. Arch. Neurol. Psychiat 1923 137-169 9
|