|
ESTIMACIÓN
DE BIOMASA DE BAIRDIELLA CHRYSOURA POR MEDIO DEL CENSO DE HUEVOS Y ALGUNOS
ASPECTOS DE SUS PRIMEROS ESTADÍOS. LAGUNA DE TÉRMINOS, CAMPECHE
(PISCES: SCIAENIDAE)
Trabajo recibido el 16 de agosto de 1984 y aceptado
para su publicación el 17 de enero de 1985.
ARTURO SÁNCHEZ ITURBE
CÉSAR FLORES COTO
Universidad Nacional Autónoma de México. Instituto
de Ciencias del Mar y Limnología. Contribución 477 del Instituto
de Ciencias del Mar y Limnología, UNAM.
Huevos y
larvas de la corvina Bairdiella chrysoura, fueron colectados
en muestreos mensuales de ictioplancton en la Laguna de Términos, de
febrero de 1980 a abril de 1981, con el fin de determinar áreas y
épocas de desove, crecimiento y mortalidad larvaria, así como
estimar la biomasa adulta a través de la producción anual de
huevos, la que junto con la producción anual de larvas fueron
determinadas por dos métodos. El desove ocurrió de enero a
octubre con su máxima intensidad de mayo a julio en la zona Este de la
laguna, principalmente en áreas de vegetación sumergida. Los
huevecillos se encontraron en salinidades de 13 a 36% y temperaturas de 21.3 a
32°C. A partir de marcas diarias en los otolitos y con ajuste lineal, se
determinó la tasa instántanea de crecimiento en 0.279. La tasa
instántanea de mortalidad calculada f ue de 0. 678, en un rango de edad
de 0. 37 a 16.65 días. La biomasa adulta estimada fue de 1,556 y 1,630
toneladas métricas, de acuerdo a los dos métodos
empleados.
Eggs an larva of Bairdiella chrysoura
of monthly ichthyoplankton samples of Terminos Lagoon, from february 1980 to
april 1981, were obtained in order to determine spawnin season and area, growht
and larvae mortality and adult biomass through the annual eggs production.
Annual eggs and larvae production were stimated by two methods. Spawing season
was from january to october, with a pike on may and july, in the east of the
Lagoon mainly on seagrass beds. Eggs were founded at 13 to
360/00 of salinity and 21.3
to 32.0°C. Instant growht rate of 0.279 was obtained by lineal
regression of daily marks on otholis. Instant mortality rate of 0.678 was
obtained for ages 0.37 to 16.65 days. Adults biomass vary from 1,556 to 1,630 m
ton. aceording with both methods used.
Los estudios de sistemas lagunares en la actualidad son de gran importancia, particularmente aquellos que tienden al análisis global del ecosistema y a la utilización de esta información para la evaluación y manejo racional de los recursos vivos. Parece obvio que esto podrá lograrse en la medida en que se conozca mejor a las especies dominantes; ya que son clave en la dinámica de estos sistemas en virtud de que su presencia no es casual sino resultado de una serie de adaptaciones que les permite el aprovechamiento óptimo de la riqueza productiva de estas áreas. En los diversos estudios que sobre los peces de la Laguna de Términos se han desarrollado (Amezcua-Linares y Yáñez-Arancibia, 1980; Vargas-Maldonado et al., 1981; Yáñez-Arancibia et al., 1982), se encuentra que la corvina Bairdiella chrysoura (Lecépède 1802), es una de las especies más abundantes y frecuentes, lo que aunado a su condición potencial de recurso pesquero ha llevado a abordarla. La corvina es un pez de amplia distribución, reportada desde Nueva York, hasta el sur del Golfo de México (Castro-Aguirre, 1978; Reséndez, 1981). La mayor parte de las publicaciones sobre esta especie se ocupan de los estados adultos y son pocas las que versan sobre sus primeros estadíos de vida, se pueden citar a aquellas sobre embriología y desarrollo larvario de Kunts (1915), WeIsh y Breeder (1923), Powles (1980); sobre sus áreas y épocas de desove por Welsh y Breede (1923); Hildebrand y Schroeder (1982). Hildebrant y Cable (1930); sobre aspectos ecológicos Darnell (1958) y Springer y Woodburn (1960). El presente estudio forma parte de un proyecto tendiente a conocer y evaluar la población de la corvina B. chrysoura en la Laguna de Términos, Campeche. La parte sobre los adultos ha sido terminada y publicada por FloresHernández (1983). Así, el objetivo del trabajo es conocer una serie de aspectos sobre los primeros estadíos de vida de esta especie en el sistema lagunar, tales como: distribución de su abundancia, crecimiento, mortalidad, área y época de desove, así como la estimación de la biomasa desovante a partir de la producción de huevos pretendiendo introducir este método de evaluación de los sistemas estuarinos y lagunares, ya que ha sido probado como un importante auxiliar en el estudio de pesquerías. ÁREA DE ESTUDIOEl área de estudio fue la Laguna de Términos, situada en el estado de Campeche, al sureste del Golfo de México entre los 18°20' y 19°00' Norte y 91°10' y 92°00' Oeste (Fig. 1). Es una laguna costera con amplia comunicación con el mar, gran afluencia de agua dulce a través de diversas descargas de ríos, riachuelos y arroyos; tiene una longitud de 70 km y una anchura máxima de 28 km. Para fines de este estudio se estimó una área de 1,567 Km2. Esta laguna es una de las más estudiadas de México, por lo que para profundizar en su descripción se sugiere acudir entre otros, a los trabajos de: AyalaCastañares, 1963; Gómez Aguirre, 1974; Yáñez-Arancibia et al., 1980; Mancilla y Vargas, 1980; Reséndez, 1981. 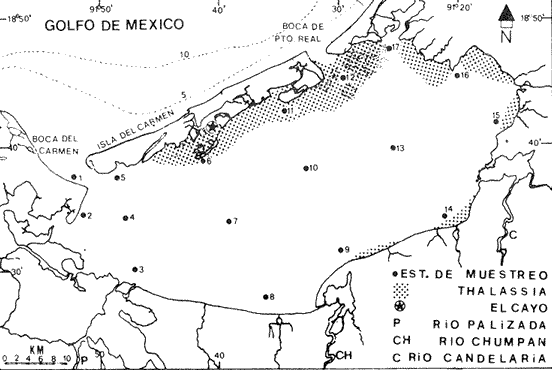 Fig. 1. Área de estudio y ubicación de estaciones de muestreo. MATERIAL Y MÉTODOS
METODOLOGÍA DE CAMPO Y DE LABORATORIOSe estableció una red de 17 estaciones de muestreo, con distancias aproximadas de 10 km entre ellas (Fig. 1), donde se realizaron colectas mensuales de ictioplancton y peces adultos, por lo que el área se cubrió generalmente en tres días. No se realizaron colectas nocturnas. En los muestreos de ictioplancton se utilizó una red tipo trapecio con malla de 500 micras, manga de 1.5 m, boca y copo colector de 50 y 10 cm de diámetro respectivamente. En la boca se montó un flujómetro previamente calibrado. Se hicieron arrastres horizontales de 10 minutos, en trayectoria circular entre 10 y 50 cm bajo la superficie, con una velocidad de arrastre entre 2 y 3 nudos. Las muestras se conservaron en frascos con formalína al 10 % neutralizadas con bórax. Se tomaron datos de salinidad, temperatura y transparencia en cada estación. En el laboratorio, de cada muestra se extrajeron el total de huevos y larvas de peces, de los cuales posteriormente se seleccionaron y procesaron aquellos de Bairdíella chrysoura, basándose para su identificación en el trabajo de Johnson (1978), CRECIMIENTO LARVARIOLa tasa de crecimiento larvario fue estimada en base al incremento en los otolitos de anillos conformados por pares de bandas, una clara y otra oscura, que representan un día en la vida de las larvas. El método ha sido utilizado por Taubert y Coble, 1977; Barkmann, 1978; Methot y Kramer, 1979; Towsend y Graham, 1981; quienes han confirmado la naturaleza diaria de las marcas en los otolitos. Los otolitos fueron extraídos bajo un microscopio de disección, lavados en agua dulce y mon tados sobre un portaobjetos en bálsamo de Canadá. De las tallas de, larvas encontradas que variaron en un rango de 2 a 6 mm, se escogió una secuencia representativa, obteniéndose una serie de 17 otolitos de los que se hicieron 5 lecturas para cada uno. El crecimiento se ajustó a un modelo de tipo lineal de acuerdo a la fórmula: Lp = bt + a donde: Lp = Longitud patrón en mm, t = Número de marcas diarias (anillos claros), b =Tasa instantánea de crecimiento, a = Constante. PROCESO DE DATOSLa metodología para el procesamiento de datos de huevos y larvas se basó en la utilizada por: Sette y AhIstrom (1948); Kramer et al. (1972); Hempel (1973); Houde (1977) y Smith y Richardson (1977). a) Estimación de la abundancía de huevos y larvasLa abundancia (Ab) de huevos y larvas en cada período de colecta se calculó por dos mé,todos (Tabla 1). El primero consistió en conocer la densidad promedio de huevos y/o larvas en la laguna por unidad de volumen y multiplicarla por el volumen total (V), estimado en 3.9 x 109 m³, que resulta de la sumatoria de volúmenes de cada estación de muestreo (vi), asumiendo una profundidad promedio en la laguna de 2.5 m, que aunque menor de la profundidad promedio real que es alrededor de 3.5 m, es aceptable, ya que se estima que el ictioplancton es escaso en el fondo. El segundo consistió en aplicar el método de polígonos de Sette y AhIstrom (1948), en el que se calcula el área de cada estación de muestreo (Tabla 2, Fig. 2), trazando líneas que marcan límites intermedios entre estaciones adyacentes; en el caso del presente estudio, en lugar de expresar los valores por m², se hace por volumen (m3), considerando la misma profundidad promedio; así entonces, se multiplica la densidad de hurvod y/o larvas de cada estación (Cij) por su volumen estimado, obteniendo posteriormente por la sumatoria de los valores de todas las estaciones, la abundancia (Ab) total de la laguna (Tabla 1). En el caso de las larvas, se obtuvieron frecuencias de talla, para lo cual se tomaron intervalos de clase de 1 mm de longitud patrón. La estimación de la abundancia se obtuvo para cada clase de talla. 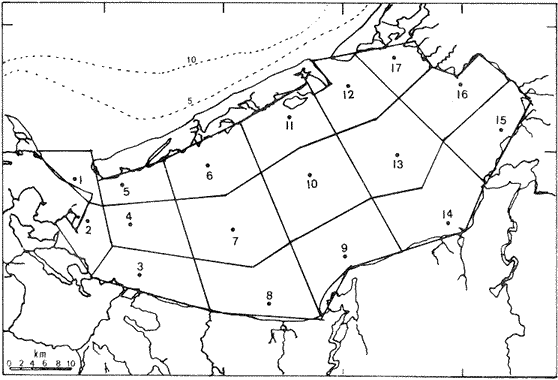 Fig. 2. Delimitación de áreas que representa calda estación de muestreo, bajo el método de polígonos de Sette y AhIstrom (1948). b) Estimación de la producción diaria, por campaña y anual de huevos y larvasLos valores de abundancia de cada período de colecta, fueron convertidos a valores de producción diaria (Pd), dividiendo la abundancia (Ab) entre el tiempo de residencia (Tr) de los huevos y de cada clase de talla de las larvas. El tiempo de residencia de los huevos, es decir el lapso desde el momento de desove hasta la eclosión, fue tomado de johnson (1978), quien lo estima en 0.75 días a 27°C, valor aceptable para el presente trabajo, ya que la variación de temperatura en las estaciones en donde se colectaron huevos de B. chrysoura fue de 22.5 a 32.0°C. Para las larvas, el tiempo de residencia fue estimado a partir del modelo de crecimiento, utilizando la siguiente fórmula: Tr = (L2 - L1) / b donde: Tr = Tiempo de residencia de las larvas (en días), L2 = Límite superior de la clase de talla (en mm), L1 = Límite inferior de la clase de talla (en mm). b = Coeficiente instantáneo de crecimiento. A partir de los datos de producción diaria y siguiendo el método de Sette y AhIstrom (1948), se obtuvo la producción por campaña (Pc), que se logra al multiplicar el valor de producción diaria por el número de días que representa la campaña (Te) y éste se obtiene sumando a los días de muestreo, la mitad de días desde que fue realizada la campaña precedente y la mitad de días antes del inicio de la siguiente (Tabla 3). La sumatoria de la producción de las diferentes campañas, lleva a obtener la producción anual (PA). Un resumen de la secuencia de pasos y fórmulas para los cálculos de abundancia y producción se dan en la Tabla 1.  TABLA 1 SECUENCIA DE PASOS PARA OBTENER POR LOS MÉTODOS DESCRITOS EN EL TEXTO, LA PRODUCCIÓN ANUAL DE HUEVOS Y/O LARVAS 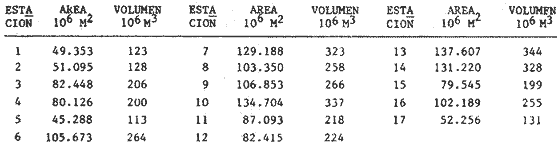 TABLA 2 ÁREA Y VOLUMEN ESTIMADOS PARA CADA ESTACIÓN DE MUSTREO, CONSIDERANDO UNA PROFUNDIDAD PROMEDIO DE 2.5 M 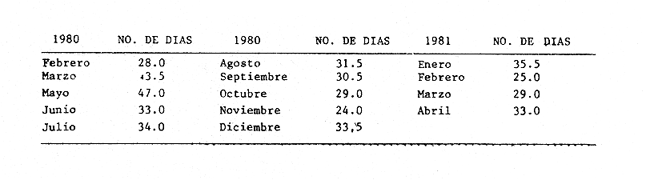 TABLA 3 TIEMPO DE CAMPANIA EN DIAS, ESTIMADO DE ACUERDO AL MÉTODO DE SETTE, Y ALHSTROM (1948) c) Estimación de la biomasa adultala biomasa adulta se estimó de acuerdo a la siguiente fórmula (Houde, 1977): B = PA / Fr K donde: B = Biomasa adulta, PA = Producción anual de huevos, Fr = Fecundidad relativa, K = Proporción de hembras en la población adulta. d) MortalidadLa mortalidad fue determinada por dos formas: la primera a partir de la disminución de la abundancia de huevos y distintas clases de talla de larvas, estimadas en todos los casos como un censo anual (C), obtenido de la sumatoria de sus abundancias mensuales (Ab). El modelo de regresión ajusta a uno de tipo exponencial de la siguiente forma: C = a e -zL donde: C = Censo anual de huevos y clases de talla de larvas, z = Coeficiente instantáneo de mortalidad (por mm de crecimiento), L = Talla (mm), a = Constante. La segunda, fue a partir de la disminución de la producción anual de huevos y larvas a medida que aumenta de edad, ajustando también a un modelo de tipo exponencial: PA = a e -zt donde: PA = Producción anual de huevos y larvas, z = Coeficiente instantáneo de mortalidad (por día), t = Edad (días), a = Constante. De los modelos anteriores se pueden deducir los porcentajes de sobrevivencia y mortalidad de acuerdo a las fórmulas: S = e -z M= (1-e -z) 100 donde: M = Porcentaje de mortalidad, S = Porcentaje de sobrevivencia; S y M se pueden calcular por mm de crecimiento y/o por día. RESULTADOS Y DISCUSIÓN
ÁREA Y ÉPOCA DE DESOVEEn la revisión que sobre B. chrysoura y otros scianidos hace Powles (1980), señala que la especie desova en aguas costeras y estuarinas, por lo que siendo este último el carácter de la Laguna de Términos, no es de extrañar que la especie ocupe esta laguna como área de desove; de hecho la distribución y abundancia de sus huevecillos así lo demuestra. Flores-Coto et al. (1983) encuentra que también desovan en la Laguna de Tamiahua, en el NO del Golfo de México. En la figura 3, en la que se gráfica la abundancia de huevos y larvas contra los meses de muestreo, puede apreciarse parte de dos cielos de desove, correspondiendo a los años de 1980 y 1981. Aunque el desove parece ocurrir todo el año, hubo al menos dos meses, noviembre y diciembre en los que no se hallaron ni huevos ni larvas. Atendiendo a la primera porción de la gráfica resulta claro que la época del máximo desove es de mayo a septiembre con un fuerte descenso hacia los siguientes meses. El nuevo cielo se inicia en el invierno. La época de mayor desove parece tener una estrecha relación con la de ascenso de temperatura, es decir con los meses de mayo a agosto (26.530°C), asimismo con la salinidad, pues el período señalado es el de lluvias y cabe resaltar la correspondencia entre la baja abundancia de huevos y larvas con la época de disminución de salinidad 1229.20%. El período de desove parece comenzar más tarde y ser más corto a mayores latitudes (Powles 1980). En los estudios que sobre los adultos se desarrollé paralelo a éste, Flores-Hernández (1983), señala haber encontrado hembras en etapa de maduración (Fase III), de febrero a abril y unas pocas desovadas (Fase IV) en abril, información con la que el autor propone la idea de que el penodo de mayor intensidad de maduración ocurre de febreroabril y que el desove es realizado por la mayor parte de la población fuera de la laguna, particularmente en los sistemas fluvio lagunares que drenan en ella. La información que sobre huevos y larvas se presentó en los párrafos anteriores no corresponde con la de FloresHernández (1983), tanto porque la mayor abundancia de los estados meroplanetónicos ocurre en primavera-verano, lo cual incluso había sido señalado anteriormente para esta misma laguna por Flores-Coto y Álvarez-Cadena (1980), como tampoco en cuanto a la zona de desove la cual se discute adelante. DISTRIBUCIÓN DE LA ABUNDANCIA DE HUEVOS Y LARVASLa distribución espacial de huevos en la laguna, mostró que en 1980 el mayor poreentaje de ocurrencia (66.8%) se presentó en el tercio este de la laguna y un poreentaje menor (27.8 %) en el tercio medio, mientras que en la porción oeste no se encontraron (Fig. 4A). El patrón de distribución de las larvas da indicios de la deriva que sufren; el mayor poreentaje (57 %) se presentó también en el tercio este de la laguna aunque no se sobrepuso estrictamente con aquella de mayor abundancia de huevos (Fig. 4B) y a diferencia de éstos, las larvas se presentaron no sólo en la parte media sino tembién en la porción oeste, encontrándose incluso en estaciones cercanas a la boca del Carmen. En la figura 4C, puede apreciarse que la casi total concentración de huevos en 1981, ocurrido alrededor de la boca de Puerto Real, esto es, otra vez los mayores valores en el tercio este. Las larvas tuvieron una ocurrencia mis amplia que los huevos abarcando propiamente toda la laguna, con su mayor poreentaje (más del 70 % en la porción sur) (Fig. 4D). Aunque el patrón de distribución de huevos y larvas bosqueja lo ocurrido en el cielo anterior (1980), no cabe una comparación estricta ya que no se muestrearon los meses de máximo desove en 1981. 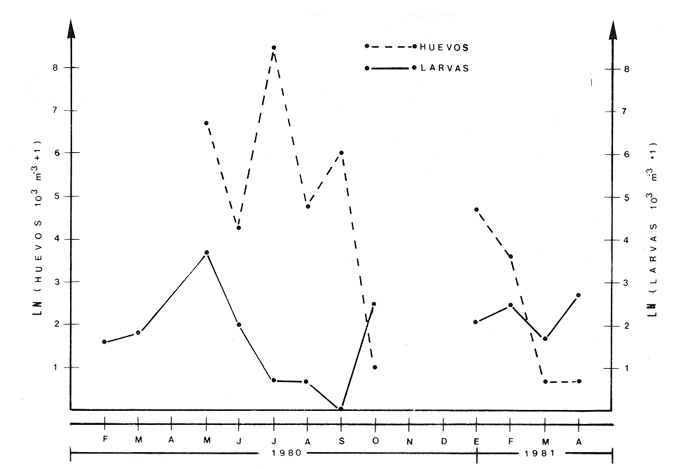 Fig. 3. Abundancia mensual de huevos y larvas de B. chrysoura. En la figura 5, se ha graficado el poreentaje acumulado de huevos y larvas en relación a los parámetros abióticos, encontrándose que el 90 % de los huevos ocurre en salinidad de 26 a 360/oo, temperaturas de 28 a 32°C y transparencias relativas mayores de 20 % ; las larvas por su parte ocurrieron en rangos más amplios, así, el 80 % de ellas se presentaron en salinidades de 18 a 32%, temperatura de 26 a 32°C y transparencias arriba de 20 %. Las áreas de mayor concentración de huevos corresponden muy estrechamente a aquellas con vegetación sumergida, salinidades, temperaturas y transparencias altas, lo que muestra a éstas como las características sobresalientes en las áreas de desove de esta especie. El señalamiento hecho por diversos autores (FloresCoto y Alvarez-Cadena, 1980; Powles, 1980;'entre otros), sobre esta especie como desovante en aguas estuarias y costeras hace suponer que sólo una parte de la población desova en la laguna y el resto en aguas costeras adyacentes. ESTIMACIÓN DE LA BIOMASA ADULTA
Abundancía y Producción de HuevosLos valores obtenidos para la producción anual (PA) de huevos, bajo los dos métodos empleados, fueron de 89.96 X 10 10 y 94.26 x 10 10 en 1980 (Tabla 4), y muestran la diferencia que se origina en el hecho de considerar a la laguna como una sola unidad o fraccionada en volúmenes. Los resultados de producción anual (PA) para 1981, 2.45 x 10 10 y 2.37 x 10 10 resultaron contrastantemente bajos respecto a 1980, como consecuencia que en 1981 el muestreo sólo abarco hasta abril, faltando mayo, junio y julio que fueron los meses de mayor desove en el año anterior. 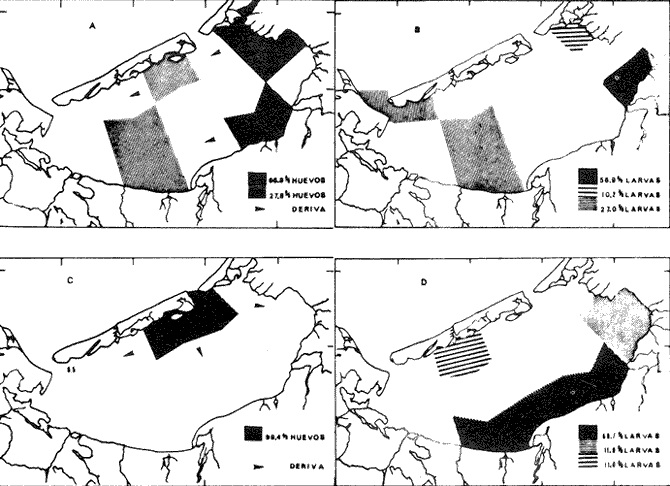 Fig. 4. Distribución espacial de huevos y larvas de B. chysoura durante los ciclos 1980 (A-D) y 1981 (B-D). 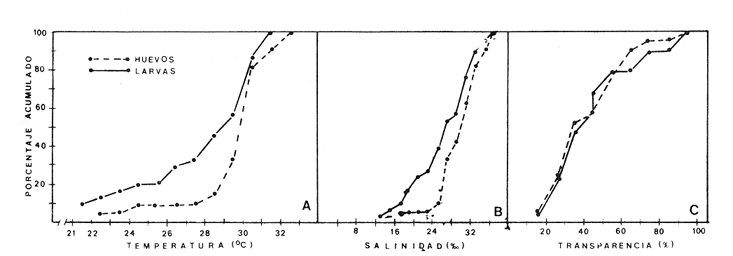 Fig. 5. Porcentaje acumulado de la abundancia de huevos y larvas de B. chrysoura en relación a la temperatura (A), salinidad (B) y transparencia (C). 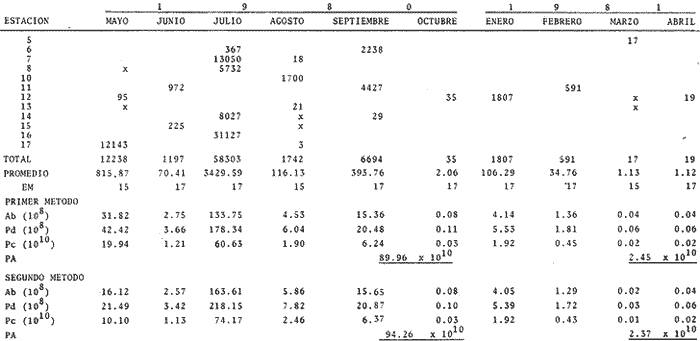 TABLA 4 ESTIMACIÓN DE LA PRODUCCIÓN DE HUEVOS DE BAIRDIELLA CHRYSOURA, POR MEDIO DE LOS DOS MÉTODOS DESCRITOS EN EL TEXTO. LOS DATOS BÁSICOS SON EXPRESADOS COMO: HUEVOS/ 1 000 M3. SE ELIMINARON ESTACIONES DE COLECTA DE NULA OCURRENCIA. (X, NO SE MUESTREO: E.M., NÚMERO DE ESTACIONES MUESTREADAS; Ab, ABUNDANCIA MENSUAL; Pd, PRODUCCIÓN DIARIA. Pc. PRODUCCIÓN POR CAMPAÑA, PA, PRODUCCION ANUAL). TIEMPO DE RESIDENCIA DE LOS HUEVOS 0.75 DíAS. Estimación de la BíomasaEn el cálculo de la biomasa se utilizaron los siguientes valores: fecundidad relativa (FR) 863 huevos/gr de hembra; proporción de hembras en la población adulta (K) 0.67; y producción anual de huevos (PA), para 1980, calculados bajo los dos métodos. Los valores de fecundidad relativa (FR) y proporción de hembras (K) provienen del estudio de Flores-Hernández (1983), cuyo trabajo de campo se realizó conjuntamente a éste. La biomasa estimada fue de 1,556 toneladas métricas bajo el primer método y de 1,630 toneladas métricas bajo el segundo. Cabe señalar que en ambos casos está muy por arriba de las 500 toneladas métricas estimadas por Flores-Hernández (1983), a partir de la captura de adultos, en la que se utilizó una red de prueba camaronera. PRODUCCIÓN LARVARIA
Crecimiento LarvarioA fin de conocer la tasa instantánea de crecimiento de las etapas larvarias de B. chrysoura, se determinó su edad, empleando el método de conteo de marcas diarias en los otolitos. En virtud de los resultados (Tabla 5), se pudo obtener un modelo de crecimiento que ajusta a una regresión lineal, para larvas comprendidas entre 2-6 mm de longitud patrón: LP = 0.279 (t) + 0.308 t = tiempo en días Este modelo, presentado en la figura 6 difiere del encontrado para otras especies, como es el caso de Archosargus rhomboidalis (Chavance et al., 1984), en el que el mejor ajuste fue exponencial. Probablemente el hecho de que B. chrysoura tuviera un ajuste lineal puede deberse al corto rango de longitud; ya que sólo abarcó larvas hasta los 6 mm, en virtud de que las más grandes tienden al fondo. 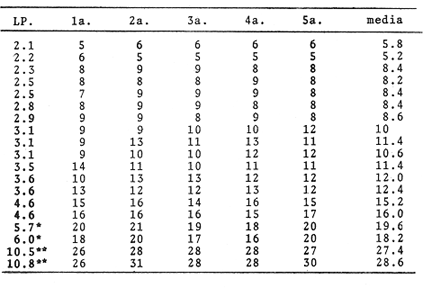 TABLA 5 NÚMERO DE MARCAS DIARIAS EN LOS OTOLITOS DE LARVAS DE B. CHRYSOURA; CINCO REPETICIONES Y SU VALOR MEDIO. (L.P., LONGITUD PATRÓN EN MM; OBTENIDOS DEL FONDO EN MUESTREOS EXTRAORDINARIOS; **NO CONSIDERADOS EN EL MODELO. 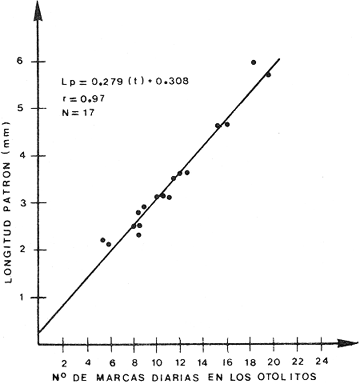 Fig. 6. Crecimiento de las primeras etapas larvarias de B. chrysoura. Sobrevivencia y MortalidadLa probabilidad de que una clase de edad pase a la siguiente, así como la relación que se da entre la cantidad de huevecillos desovados y la abundancia de la última etapa larvaria, depende de la sobrevivencia y mortalidad en estas primeras fases, que sin duda son las m á s sensibles a los cambios del medio ambiente dentro del cielo de vida de los peces; por ello, una buena evaluación de estos parámetros permitirán estimar a una de las principales causas de las variaciones de abundancia de la población adulta. El análisis sistemático de la mortalidad y sobrevivencia podrán servir como indicadores de variaciones en el ecosistema. A fin de estimar la sobrevivencia y mortalídad, los resultados de las abundancias mensuales (Ab), tanto de huevos (Tabla 4), como de cada clase de talla de las larvas (Tabla 6), fueron sumados para cada cielo obteniéndose un censo anual (Tabla 7): C = ΣAb En la figura 7, se han graficado los valores de cada censo anual de huevos y clases de talla de larvas contra el incremento de talla; lo que llevó a un modelo de declinación con un ajuste exponencial. Los resultados para cada cielo se presentan en la Tabla 8 donde puede apreciarse que los valores del coeficiente instantáneo de mortalidad obtenidos para el primer cielo: 2.15 y 2.34, corresponden a un poreentaje de 88 y 90 % de mortalidad; lo que equivale a 12 y 10 % de sobrevivencia respectivamente; y para 1981, 0.77 y 0. 75, representan 54 y 53 % lo que equivale a un 46 y 47% de sobrevivencia. Previo a este trabajo, pero proveniente del mismo programa de muestreo ictioplanetónico, Chavance et al., 1984, estimaron la tasa de mortalidad de A. rhomboidalis, esto ha permitido comparar con lo obtenido para B. chrysoura, encontrándose que para esta última especie los valores son muy altos. Cabe señalar que los estados meroplanetónicos de estas dos especies tienen una distribuci ión muy similar dentro de la laguna. La alta tasa de mortalidad de B. chrysoura, aunada a una distribución restringida al tercio este de la laguna, donde prevalecen condiciones cercanas a las marinas, parece indicar que la laguna no constituye un sitio idóneo para ser utilizado como área de desove y crianza para esta especie. Esta misma idea apoya aquella discutida en párrafos anteriores en relaci ión a considerar que sólo una parte de la población desova en la laguna y la mayor parte lo hace en la zona costera adyacente; y asimismo se contrapone a aquella otra de que el desove ocurre en los sistemas fluvio lagunares propuesto en base a la distribución de las hembras maduras (Amézcua Linares y Yáñez-Arancibia, 1980 y Flores-Hernández, 1983). Producción Anual de LarvasLa producción anual de larvas (PA) fue posible calcular únicamente en virtud de haber podido conocer la edad (t) y tiempo de residencia (Tr) para cada clase de talla. La determinación del tiempo de residencia (Tr) se calculó en base a la fórmula expuesta en la metodología; en virtud de que el modelo de crecimiento fue lineal el tiempo de residencia fue constante para todas las clases de talla siendo éste de 3.23 días. La edad se calculó a partir del modelo de acuerdo al despeje siguiente: LP = 0. 279 (t) + 0. 308 t = LP - 0.308 / 0.279 El cálculo de la producción anual (PA) de cada clase de talla de las larvas siguió la misma metodologá que la de los huevecillos. Los resultados son presentados en las Tablas 6 y 7, donde puede apreciarse que los mayores valores se presentaron en el primer cielo como resultado de que durante 1981 no se muestrearon los meses de mayor desove. A diferencia de la producción anual de huevos, la de las larvas no fue utilizada para la estimación de biomasa desovante, sino para desarrollar un modelo de sobrevivencia más preciso y accesible de manejar, pudiéndose utilizar como un parámetro de monitoreo. Se considera más preciso; ya que mientras el modelo de sobrevivencia que se discutió en párrafos anteriores, sólo utiliza la declinación de abundancia de las tallas de larvas, en este otro, se involucra el tiempo de residencia de las diferentres clases de tallas, edad promedio de cada una de ellas y la extrapolación de lo que representan a través de las campañas de muestreo. Así entonces se construyó un modelo de declinación de la producción de las etapas meroplanetónicas, incluyendo a los huevos, en el que el ajuste fue de tipo exponencial (Fig. 8), con los resultados que se presentan en la Tabla 8. 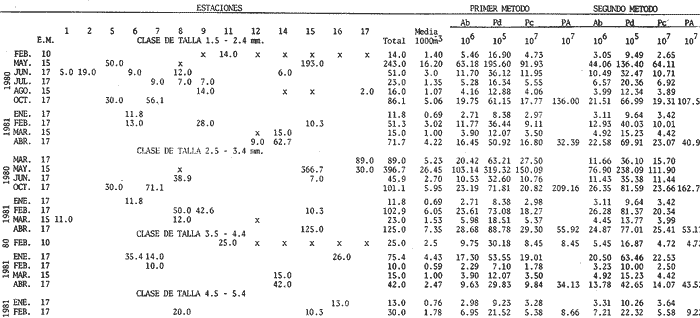 TABLA 6 ESTIMACIÓN DE LA PRODUCCIÓN ANUAL DE LARVAS POR CADA CLASE DE TALLA DE BAIRDIELLA CHRYSOURA POR MEDIO DE LOS MÉTODOS DESCRITOS EN EL TEXTO. LOS DATOS BÁSICOS SON EXPRESADOS COMO: LARVAS/1 000 M3. SE ELIMINARON ESTACIONES DE NULA OCURRENCIA (X, NO SE MUESTREO; E.M. ESTACIONES MUESTREADAS; Ab, ABUNDANCIA MENSUAL; Pd, PRODUCCIÓN DIARIA; Pc, PRODUCCIÓN POR CAMPAÑA; PA, PRODUCCIÓN ANUAL). TIEMPO DE RESIDENCIA IGUAL EN CADA CLASE DE TALLA (3.23 DÍAS) 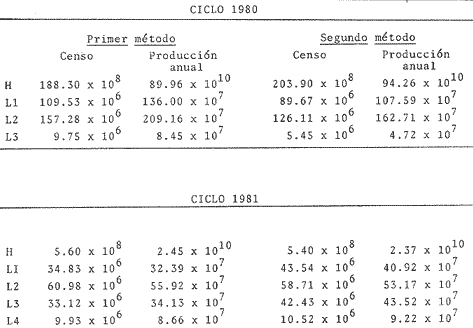 TABLA 7 CENSO Y PRODUCCIÓN ANUAL DE HUEVOS Y LARVAS DE B. CHRYSOURA EN LA LAGUNA DE TÉRMINOS, DURANTE LOS AÑOS DE 1980 Y 1981, BAJO LOS DOS MÉTODOS DESCRITOS EN EL TEXTO. LOS RANGOS DE TALLA Y TALLA MEDIA (MM) EN CADA CASO SON: HUEVO (H) 0.68-088, 0.78; LARVA 1 (L1) 1.5-2.4, 1.95; LARVA 2 (L2) 2.5-3.4, 2.95; LARVA 3 (L3) 3.5-4.4, 3.95; LARVA 4 (L4) 4.5-5.4, 4.95. 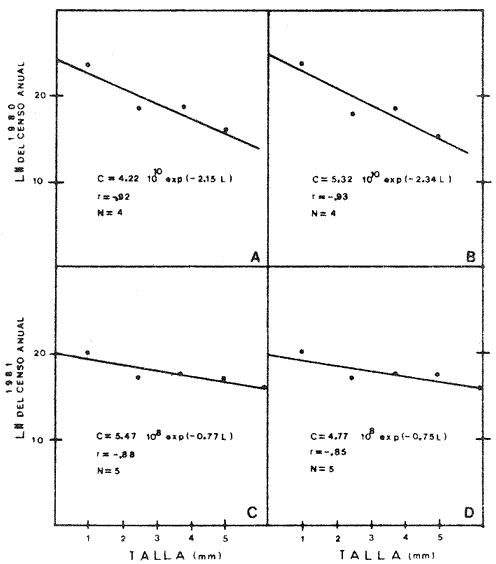 Fig. 7. Tasa instantánea de mortalidad, obtenida como un ajuste exponencial de la relación entre la abundancia censada de huevos y, larvas de B. Chrysoura y su longitud patrón, conforme aumenta de talla. Valores del censo obtenidos bajo los dos métodos descritos en el texto. (A y C, primer método; B y D, segundo método). En los resultados de 1981 por razones ya explicadas, las altas tasas instantáneas de mortalidad de 1980, llevan a las consideraciones que se discutieron al tratar la mortalidad por talla. Este modelo de sobrevivencia ofrece la posibilidad de que en años venideros con solo muestrear en la época de mayor abundancia larvaria mayojulio y tomando únicamente las larvas más grandes, de mayor edad, traerlas a este modelo se podrá estimar la producción anual de huevos y por ende la biomasa desovante. 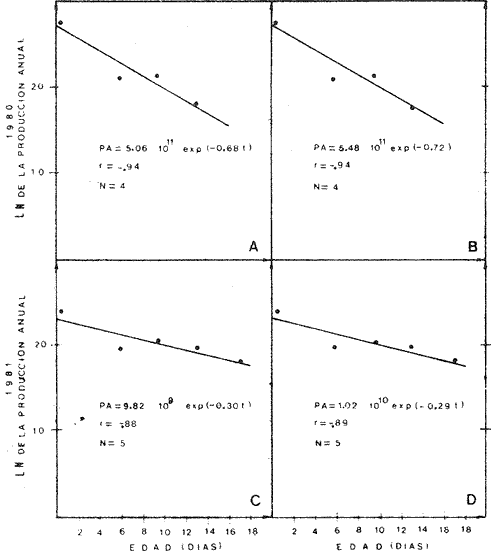 Fig. 8. Tasa instantánea de mortalidad, obtenido como un ajuste exponencial de la relación entre la producción anual de huevos y larvas de B. chrysoura y su edad en días. Valores de producción obtenidas bajo los dos métodos descritos en el texto. (A y C, primer método; B y D, segundo método).  TABLA 8 TASAS INSTANTÁNEAS DE MORTALIDAD (Z) Y PORCENTAJES CORRESPONDIENTES DE MORTALIDAD (M) Y SOBREVIVENCIA (S), OBTENIDAS A PARTIR DEL DECRECIMIENTO DE LA ABUNDANCIA CENSADA (c) Y DE LA PRODUCCIÓN ANUAL (PA) DE HUEVOS Y LAS SUCESIVAS CLASES DE TALLA DE LARVAS DE B. CHRYS0URA. Conclusiones1. Baírdiella chrysoura desova en la Laguna de Términos principalmente de mayo a septiembre que corresponden a los meses de temperaturas altas y resalinización de la laguna; ocupando áreas con vegetación sumergida, salinidades, temperaturas y transparencias altas, encontrándose el 90 % de los huevos en salinidades de 16 a 360/00 y temperaturas de 28 a 32°C. 2. La producción anual de huevos durante 1980, calculada bajo los dos métodos descritos en el texto, fue de 89.96 x 1010 y 94.26 x 1010; lo que en virtud de su fecundidad relativa (FR = 863 huevos/gr (femenino)), proporción de hembras en la población adulta (K = 0.67 (femenino)), llevó a estimar una biomasa de 1,556 y 1,630 toneladas métricas, bajo estos métodos. 3. Empleando el método de conteo de marcas diarias en los otolitos, se llegó a un modelo de crecimiento, que ajustó a una regresión lineal, con larvas comprendidas entre 2 y 6 mm de longitud patrón, obteniéndose un coeficiente instantáneo de crecimiento de 0.279. 4. Basándose en las sumatorias de las abundancias mensuales de huevos y cada clase de talla y bajo dos métodos descritos, se estimó la tasa de mortalidad para 1980, que fue Z = 2.15 y Z = 2.34 correspondiendo al 88 y 90 % de mortalidad; esto es 12 y 10 % de sobrevivencia respectivamente. 5. Conociendo la edad y tiempo de residencia para cada clase de talla de larvas, pudo entonces calcularse la producción anual de larvas, que a diferencia de la de huevos, fue utilizada no para la estimación de la biomasa desovante, sino para desarrollar un modelo de sobrevivencia, cuya virtud sobre el anterior es incluir el factor tiempo, y poder obtener un coeficiente initantáneo de mortalidad por día que ofrece la posibilidad que en futuras campañas, tomado sólo la época de mayor abundancia larvaria, se pueda, considerando únicamente las larvas de mayor edad, estimar la producción anual de huevos (PA) y por ende la biomasa desovante. AgradecimientosLos autores desean expresar su agradecimiento a Virgilio Arenas, por la revisión crítica del manuscrito, a Faustino Zavala por su ayuda durante el desarrollo del trabajo, a Domingo Flores por su compañerismo en campo y laboratorio y al Instituto de Ciencias del Mar y Limnología, UNAM por las facilidades otorgadas. LITERATURAAMEZCUA LINARES, F. y A. YÁÑEZ-ARANCIBIA An. Centro Cienc. del Mar y Limnol. Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces. Univ. Nal. Autón. México 1980 69-118 7 (1) AYALA-CASTAÑARES, A. Bol. Inst. Geol. Univ. Nal. Autón. México Sistemática y distribución de los foraminíferos recientes de la Laguna de Términos, Campeche, México. 1963 1-130 67 (3) BARKMAN, R. C. The use of otolith growth rings to age young Atlantic silverides, Menidia menidia. Trans. Am. Fish. Soc. 1978 790-792 107 (6) CASTRO AGUIRRE, J. L. Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México con aspectos zoogeográficos y ecológicos. Inst. Mal. de Pesca. Ser. Cient. 1978 1-298 19 CHAVANCE, P., C. FLORES-COTO y A. SÁNCHEZITURBE Early life history and adult bíomass of sea bream in the Terminos Lagoon, Southern Gulf of Mexico. Trans. Am. Fish. Soc. 1984 116-177 113 DARNELL, R. M. Food habits of fishes and larger invertebrates of Lake Pontchartrain, Louisiana and estuarine community. Publ. Inst. Mar. Sci. Univ. Tex. 1958 353-416 5 FLORES-COTO, C. y L. ÁLVAREZ-CADENA An. Centro Cienc. del Mar y Limnol. Estudios preliminares de distribución y abundancia del ictioplancton en la Laguna de Términos, Campeche. Univ. Nal. Autón. México 1980 67-78 7 (2) FLORES-COTO, C., F. BARBA-TORRES y J. SÁNCHEZROBLES Seasonal Diversity, Abundance and Distribution of Ichthyoplankton in Tamiahua Lagoon, Western Gulf of Mexico. Trans. Am. Fish. Soc. 1983 247-256 112 (2B) FLORES-HERNÁNDEZ D. Tesis profesional. Ecología y dinámica de las poblaciones de la corvina Bairdiella chrysoura. Lecépkede 1983, en la Laguna de Términos, Campeche. (Pisces: Scianidae). Fac. de Cienc. Univ. Nal. Autón. México 1983 112 p. GÓMEZ-AGUIRRE, S. An Centro Cienc. del Mar y Limnol. Reconocimientos estacionales de hidrología y plancton en la Laguna de Términos, Campeche, México (1964-1965). Univ. Nal. Autón. México 1974 61-82 1 (1) HEMPEL, G. Fish egg an larvae surveys (Contribution to a manual) FAO. Fish Tech. Pap. 1973 1-82 122 HILDEBRAND, S. F. y W. C. SCHROEDER Bull U.S. Bur. Fish. Fishes of Chesapeake Bay. 1982 388 p. 43 (1) HILDEBRAND, S. F. y L. E. CABLE Bull. U. S. Bur. Fish. Development and life history of fourteen teleostean fishes at Beaufort, N. C. 1930 383-488 46 HOUDE, E. D. Fish. Bull. Abundance and potential yield of the round herring Etrumeus teres, and aspects of its early life history in the eastern Gulf of Mexico. 1977 61-89 75 (1) JOHNSON, D. G. Development of fishes of the midAtlantic Bigts, an Atlas of egg, larval and juvenile stages. Fish and Wildlife Service, U. S. Department of the Interior. Biol. Serv. Prog. 1978 172-179 4 KRAMER, D., M. J. KALIN, G. STEvENsE, J. R. THRAILKILL, y J. R. ZWEIFEL Collecting and processing data eggs and larvae in the California current region. NOAA Tech. Rep., NMSF. Circ. 1972 1-38 370 KUNTZ A. Bull. U. S. Bur. Fish. The embiologý and larval development of Bairdiella chrysoura y Anchovia mitchilli. 1915 1-19 33 MANCILLA-PERAZA, M. y M. VARGAS-FLORES An. Centro Cienc. del Mar y Limnol. Los primeros estudios sobre la circulación y el flujo neto de aguas a través de la Laguna de Términos, Campeche. Univ. Nal. Autón. México 1980 1-12 7 (2) METROT, R. D. y D. KRAMER Fish. Bull. Growth of northern anchovy Engraulis mordax, larvae in the sea. 1979 413-423 77 (2) POWLES-HOWARD Fish. Bull. Description of larvai silver perch, Bairdiella chysoura, Banded Drum, Larimus fasciatus, Star Drum, Stellifer lanceolatus (Scianidae) 1980 119-136 78 (1) RESÉNDEZ-MEDINA, A. Biótica Estudio de los peces de la Laguna de Términos, Campeche, México. 1981 239-430 6 (4) SETTE, O. E. y E. H. AHLSTROM estimations of abundance of eggs of the Pacific pilchard (Sardinops caerulea) off southern California during 1940 and 1941. J. Mar. Res. 1948 511-542 7 (3) SMITH, P. E. y S. L. RICHARDSON Standard techiques for pelagic fish egg and larval surveys. FAO Fish. Tech. Pap. 1977 1-100 175 SPRINGER, Y. G. y K. D. WOODBURN An Ecological study of the fishes of Tampa Bay area. Fl. St. Board Conserv. Mar. Lab. Prof. Pap. Ser. 1960 1-104 (1) TAUBERT, B. D. y D. W., COBLE Daily rings in otoliths of three species of Lepomis and Tilapia mossambica. J. Fish. Res. Board. Can. 1977 332-340 34 TOWSEND, D. W. y J. J. GRAHAM Fish. Bull. Growth and age structure of larval Atlantic herring, Clupea harengusharengus, in the Sheepscot River estuary, Maine, as determined by daily growth increments in otoliths. 1981 123-130 79 (1) VARGAS MALDONADO, I., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA- LINARES An. Inst. Cienc. del Mar y Limnol. Ecología y estructura de las comunidades de peces en áreas de Rhizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos, sur del Golfo de México. Univ. Nal. Autón. México 1981 241-266 8 (1) WELSH, W. W. y C. M. BREEDER, Jr. Bull. U.S. Bur. Fish. Contribution to life histories of Scianidae of the eastern United States coast. 1923 141-210 39 YÁÑEZ-ARANCIBIA, A., F. AMEZCUA LINARES y J. W. DAY Estuarine Perspectives. Fish community structure and function in Terminos Lagoon a tropical estuary in the Southern Gulf of Mexico. Kennedy, V. Academic Press Inc. New York 1980 465-482 YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍGUEZ, P. SÁNCHEZ, I. VARGAS, P. CHAVANCE, F. AMEZCUA LINARES, A. AGUIRRE LEÓN y S. DÍAZ Coastal Lagoans. Ecosystem dynamies and nichthemeral and seasonal programming of fish community structure, ln a tropical estuarine inlet, Mexico. Lasserre, P. y H. Postma Oceanológica Acta. 1982 417-429 5 Vol. Spec. (4)
|