|
EVOLUCIÓN OCEANOGRÁFICA DE LA BOCA DEL GOLFO DE
CALIFORNIA
Trabajo presentado en el 27° Congreso Internacional de
Geología, Moscú, Rusia, agosto, 1984. recibido el 27 de
septiembre de 1984 y aceptado para su publicación el 29 de noviembre de
1984.
ADOLFO MOLINA-CRUZ
Universidad Nacional Autónoma de México. Instituto
de Ciencias del Mar y Limnología. Contribución 476 del Instituto
de Ciencias del Mar y Limnología, UNAM.
De acuerdo a
datos mieropaleontológicos y sedimentológicos, el Golfo de
California es una cuenca oceánica "joven" que inició su
formación hace aproximadamente 5 millones de años. Durante ese
tiempo, su régimen climático-oceanográfico fue subtropical
como lo es en el presente. Sin embargo, las incursiones de la Corriente de
California y los procesos de surgencias costeras, que "hoy" son comunes, no
fueron establecidos sino hasta el Pleistoceno temprano. Las surgencias costeras
ocurren tanto en el margen ocidental (en verano) como en el margen oriental (en
invierno) del Golfo. En el lado occidental, cerca de la boca del Golfo, hay una
influencia más directa de la Corriente de California en los procesos de
surgencias, en la formación de "frentes oceánicos- y en el
incremento de la productividad primaría. Este hecho, acoplado a
condiciones de gran preservación de materia orgánica en los
sedimentos (la capa mínima de oxígeno incide sobre el fondo del
talud continental), así como también a una dilución
mínima por clásicos terrígenos, ha inducido a la
acumulación de nódulos de fosforita en varias localidades. Las
deducciones hechas han sido desarrolladas analizando la distribución
geográfica y estratigráfica de conjuntos de radiolarios,
definidos mediante modelos matemáticos.
According to
micropaleontological and sedimetological data, the Gulf of California is a
"young" ocean basin which began its formation approximately 5 millions years
ago. At the time, its clim atical-oceano graphic regime was subtropical as it
is today. However, the incursions of the California current an the seasonal
coastal upwelling processes which are commun at present, were not established
up to approximately the early Pleistocene. Coastal upwelling occurs both in the
western side (summer) an the eastern side (winter) of the Gulf. At the western
side, near the mouth of the Gulf, there is more direct influence of water from
the California Current in the upwelling process, in the formation of "thermal
fronts- in the water mixed-layer, and in the increase of primary productivity.
This fact has induced the aceumulation of phosphorite nodules in several
localities; coupled to conditions of large organic material preservation in the
sediments (the oceanic oxygen minimum layer impinges on the bottom slope), as
weIl as little relative dilution by terrigenous debris. The above deductions
have been developed analyzing the geographic and stratigraphic distribution of
radíolarian assemblages, through mathematical models.
El Golfo de California es uno de los mares más estudiados en el mundo. Schwartzlose y Hendrickson (1980), reportan que ha habido más de 4,000 trabajos sobre el Golfo publicados durante este siglo. No obstante, el tópico relacionado a su evolución oceanográfica aún es modesto y relativamente reciente (ejem. Ingle, 1984). Por consiguiente, en el presente estudio aportamos nuevas ideas al respecto, utilizando técnicas modernas en paleo-oceanografía. Las ideas sobre el origen o formación del Golfo de California, desde un punto de vista tectónico, han cambiado con el tiempo; desde considerarlo una estructura depresional tipo "horst", hasta considerarlo como producto de la dispersión del piso oceánico, de acuerdo al concepto actual de "placas tectónicas" (Curray et al., 1982). Avocados a este último concepto, es de esperarse que el Golfo de California, en la medida que se forma, presente una evolución morfológica y batimétrica, así como una invasión de aguas marinas del adyacente Océano Pacífico (Fig. 1). Dichas aguas han venido modificando sus características oceanográficas en respuesta a factores climáticos, tal como ha ocurrido en otras partes del mundo (CLIMAP, 1976). En el presente estudio, para deducir dicha evolución oceanográfica, así como sus consecuencias interelimáticas se desarrolla la siguiente estrategia: lo. se infiere como las señales oceanográficas son expresadas por el registro sedimentario, definiendo micropaleontológicamente la distribución geográfica de especies y conjuntos de radiolarios recientes, y analizando la asociación que dicho patrón distribuitario guarda con distintos factores oceanográficos de la región. Después, dentro de un marco estratigráfico cuaternario, establecidos en los núcleos BAP 83-7 (22° 55' N; 108° 13' W) y BAP 83-17 (23° 22' N; 108° 49' W), se estructura una historia paleoceanográfica de la región, tomando en cuenta las asociaciones: patrón distribuitario-factor oceanográfico, definidas anteriormente, y asumiendo que las fluctuaciones en abundancia de las especies y conjuntos de radiolarios es causada por variaciones ambientales. 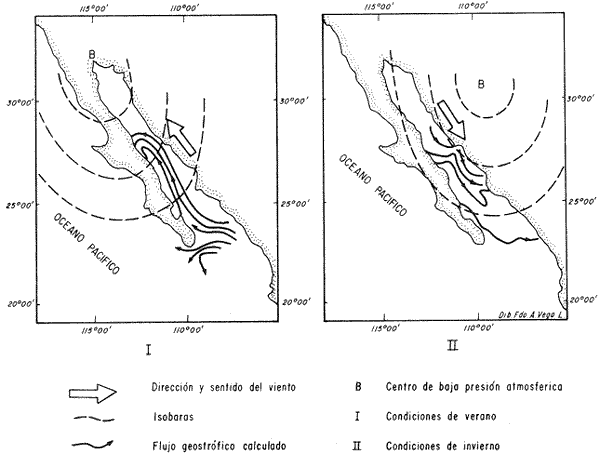 Fig. 1. Estructura barométrica, y circulación del viento y corrientes superficiales en el Golfo de California (según Schrader y Baumgartner, 1983). Por último, el criterio argumentado anteriormente, se extiende a tiempos más remotos, utilizando una columna sedimentaria que casi alcanza el contacto Mioceno/Plioceno: el sitio 476 de la etapa 64 del Proyecto de Perforación Profunda ("DSDP, site 476, leg 64"; 23°02'N; 109°05'W). La distribución y composición específica de conjuntos de radiolarios fue definida mediante la técnica matemática de análisis de factores, en su modo "Q" (Klovan e Imbrie, 1971). La cronoestratigrafía utilizada está basada en tasas de sedimentación definidas regionalmente. MARCO OCEANOGRÁFICOEl Golfo de California es una cuenca alargada de aproximadamente 1,200 km de longitud y localizada en el noreste de México, entre los 23 y 32° latitud Norte y los 107 y 117° longitud Oeste. Este golfo, aun cuando está en estrecha comunicación con el Océano Pacífico por medio de su "boca", tiene un clima continental, debido a su forma tan alargada y a que está situado entre dos zonas áridas: la península de Baja California al "occidente" y en forma general, el desierto de Sonora al "oriente" (Fig. 1). La península de Baja California, ocupada por una larga cordillera, obstaculiza el efecto termorregulador del Océno Pacífico sobre el interior del golfo. Consecuentemente, los rangos en los registros de temperatura, tanto diurnos como anuales, son grandes, relativos a los de otras regiones (Roden, 1964). Durante el final del invierno y principios de primavera, el centro atmosférico anticielónico del Pacífico nororiental concurre latitudinalmente con el centro atmosférico ciclónico del desierto de Sonora (Fig. 1). Tal hecho crea un gradiente barométrico que produce que los vientos geostróficos se hagan paralelos al eje axial del golfo y se intensifiquen, manteniendo así un clima seco, induciendo el transporte de agua superficial al sur y generando surgencias a lo largo de la costa oriental. Durante el verano y principios del otoño, la estructura atmosférica es diferente respecto a la antes descrita: la emigración del centro de baja presión del desierto de Sonora al norte del golfo, origina que los vientos provengan del sureste (Fig. 1). Estos vientos cargados con humedad provocan precipitación en la costa oriental del golfo, principalmente en la porción sur, inducen el transporte de las aguas superficiales hacia el interior del golfo y generan surgencias sobre la costa occidental. En el Golfo de California, el "control" del patrón circulatorio superficial por el campo eólico es evidenciando "claramente" por mediciones directas y cálculos hidrodinámicos. Roden (1964), realizando una sección entre Cabo San Lucas y Bahía Banderas (Fig. 2), descubrió un flujo superficial hacia el sureste con velocidades entre 10 y 15 cm/seg, de febrero a mayo, y hacia el noroeste, con velocidades máximas de 10 cm/seg. entre junio y septiembre. El mismo autor, haciendo un cálculo hidrodinámico, definió una corriente superficial neta de 10 cm/seg saliendo del golfo, en febrero; y una de 21 cm/seg entrando al golfo, en agosto. La distribución geográfica de la temperatura "superficial" (Fig. 2) no sólo ayuda a "delimitar" masas de agua, sino que refleja, en cierto grado, el patrón circulatorio. De este modo, definimos en la entrada del golfo: 1) a la Corriente de California transportando, alrededor de la península de Baja California, aguas relativamente más frías (T < 22 °C) y menos salinas (S < 34.60/00) que las adyacentes; 2) a el transporte noroccidental del agua cálida y medio salina, (T > 25°C; 34.60/00 < S > 34.90/00) del Pacífico tropical oriental, en la porción sureste de la boca, y 3) aunque no muy bien definido, el transporte al sur del agua altamente salina y "templada" (S > 34.90/00; 22 °C < T < 25°C) originada en el interior del golfo. La masa de agua del Pacífico tropical oriental, proveniente del sureste, durante el verano penetra muy al interior del golfo, limitando la influencia superficial de la Corriente de California en dicho interior (Robinson, 1973). En el interior del golfo, mediante la distribución térmica, se distinguen particularmente dos fenómenos circulatorios: 1) el "estrangulamiento" de las corrientes de marea en el "Canal de Ballenas", y 2) los grandes centros de surgencias costeras (Fig. 2). La canalización de las corrientes de marea induce a la intensificación deéstas y a su consecuente mezcla con las aguas relativamente más frías de la subsuperficie; por lo tanto, en la región del Canal de Ballenas observamos temperaturas más frías que en las regiones circundantes (Gaxiola, 1978). 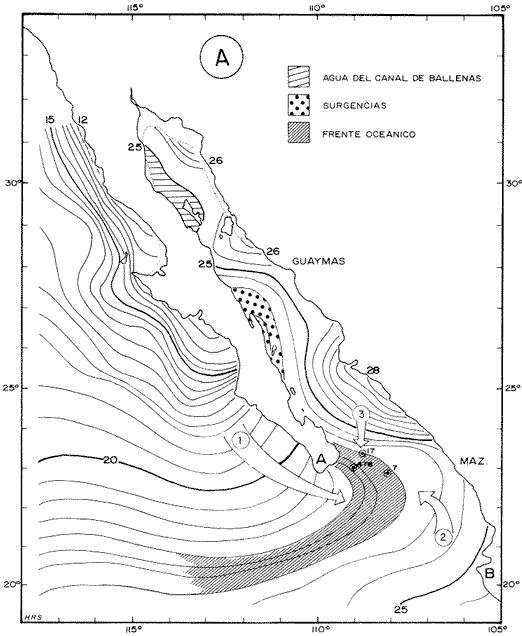 Fig. 2. Distribución de temperatura (°C) a 30 m de profundidad, durante los meses de agosto (A) y febrero (B), según Robinson (1973). Sobre esta distribución, mediante sombreados, se ha definido la ubicación de diversos fenómenos oceanográficos: distribución de masas de agua, frentes oceánicos y surgencias. Mediante flechas se indica el flujo de la Corriente de California (1), del agua Tropical (2), y del agua del Golfo (3). La letra "A" en la península de Baja California indica la localización de Cabo San Lucas, y la letra "B", cerca de los 20° de lat. N, la localización de Bahía Banderas. Presumiblemente, en la región oriental de la cuenca de Guaymas en invierno, y en la región occidental de la cuenca del Carmen en verano, se definen dos grandes centros de surgencias (Fig. 2). Aparentemente, las surgencias de la cuenca del Carmen son alimentadas en parte por incursiones subsuperficiales de la Corriente de California, como se argumentará más adelante. El encuentro de dos o tres tipos de agua forman gradientes térmicos horizontales muy marcados, indicativos de "frentes oceánicos" (Cromwell y Reid, 1956), principalmente en la entrada del golfo (Griffiths, 1963; Warsh et al., 1973). En la subsuperficie de dichos frentes, generalmente se estructura una hidroestratigrafía en la que en agua "fria y densa- se acuña bajo el agua de mayor temperatura y más ligera (Simpson et al., 1979). 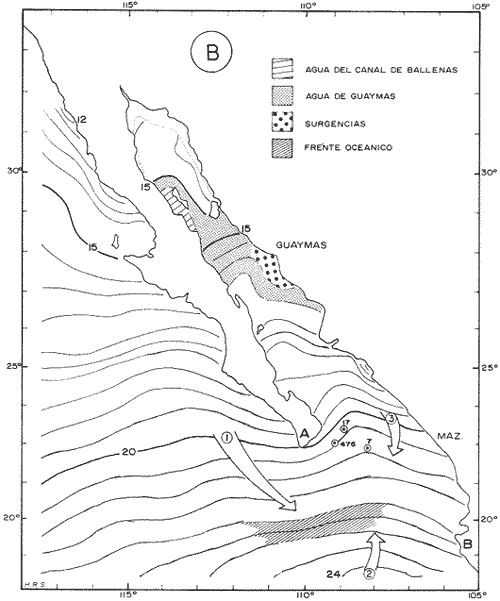 Fig. 2-B La posición de los sistemas de frentes oceánicos varía tanto geográficamente como batigráficamente a través del tiempo, como consecuencia de procesos climático oceanográficos (Wyrtki, 1965); por ejemplo, con la intensificación de las corrientes. La posición de los frentes oceánicos de la entrada del golfo varía batigráficamente entre 0 y 200 m de profundidad (Warsh et al., 1973; Robinson, 1973), y geográficamente están localizados generalmente cerca de los cabos, en el extremo sur de la península de Baja California. Las mezclas de agua que ocurren en el Golfo de California, provocadas por fenómenos de surgencias, "canalización" de corrientes de mareas, y formación de frentes oceánicos, inducen al regeneramiento de nutrientes en la superficie y consecuentemente a el de florecimiento de plancton (Roden y Groves, 1959; Hasting y Turner, 1965; Roden, 1972; Soutar et al., 1981 ; Schrader et al., 1984). Tal condición produce que el talud continental del golfo y la parte norte de la cuenca de Guaymas, principalmente alrededor de las islas Tiburón y Ángel de la Guarda, sean regiones donde hay una gran concentración de material orgánico en los sedimentos (Van Andel, 1964; Molina-Cruz, 1979). Además, de que dicha concentración es preservada por condiciones anóxicas, originadas por un marca grado mínimo de oxí no de la columna hidráulica que incide sobre el fondo marino, entre aproximadamente 400 y 800 m de profundidad (Calvert, 1964; 1966). En la parte sur del golfo, el régimen fluvial, principalmente el de oriente, impone un carácter terrígeno-detrital al fondo de las cuencas (Van Andel, 1964; Aguayo, 1981). Una compleja variación o alternación en el flujo sedimentario descrito, produce lamínaciones ("varves") en una gran parte de los sedímentos de talud continental del golfo (Calvert, 1966: Donegan y Schrader, 1982). MATERIAL Y MÉTODOSLas muestras sedimentarias utilizadas en el presente estudio fueron obtenidas principalmente en dos 'campañas oceanográficas (BAP82 y BAP83), realizadas a bordo del B/O "El Puma" de la Universidad Nacional Autónoma de México (UNAM), en los veranos de 1982 y 1983. En la región de la Cuenca de Guaymas, algunas muestras de la serie "L" de la colección de Scripps fueron utilizadas como complemento. La localización de las muestras es indicada en las Tablas 1 y 2. Las columnas sedimentarias fueron obtenidas con nucleadores tipo caja "Reinck" y de gravedad, en sus versiones: "Kasten" y "Gran diámetro" (11. 5 cm). Estos núcleos fueron submuestreados a bordo, obteniéndose para el estudio de "distribución superficial" aproximadamente 10 cc de sedimentos del primer centímetro de los núcleos. La densidad de submuestreo a lo largo de los núcleos considerados es indicada en la Tabla 2. Para el estudio estratigráfico se eligieron los núcleos BAP83-7 y BAP83-17 porque éstos provienen de regiones cercanas a fronteras entre masas de agua (Fig. 2); es decir de lugares convenientes para observar posibles desplazamientos de esas masas de agua. El núcleo del sitio 476 de la etapa 64 del Proyecto Internacional de Perforación Profunda (IPOD, DSDP) fue escogido para este estudio porque, a la fecha, éste comprende el mayor tiempo geológico, "atravesado" en la columna se dimentaria del golfo; a tal grado, que alcanza el "basamento" más antiguo que se conoce en el Golfo de California (edad aproximada: Mioceno Tardío; Curray et al., 1982). La bioestratigrafía Cuaternaría del Golfo de California generalmente, presenta resoluciones no menores a 200,000 años (Aubry et al., 1982); particularmente en la bioestratígrafía a base de radiolarios (Molina-Cruz, 1982). Por consiguiente, es necesario efectuar una fechación por medios isotópicos (δ 18O) o radiométricos (14C) para estructurar una estratígrafía de referencia e intentar correlaciones entre núcleos. No contando con facilidades locales para efectuar análisis de 14C o δ 18O, se definió una correlación bioestratigráfica entre los núcleos BAP83-7 y BAP83-17, utilizando máximos y minimos de los registro de Druppatractus pyriformís (Figs. 3 y 4), y los siguientes elementos de juicio. En un estudio previo, en la parte media del golfo (Alvarez-Arellano y Molina-Cruz, 1986), se observó que el comportamiento cronoestratigráfico de D. pyriformis es repetitivo en varias localidades (Fig. 5); posiblemente como consecuencia de cambios paleoceanográficos regionales. Consiguientemente, se cree que esta observación puede ser interpolada a otras regiones del golfo, si se consideran los efectos causados por diferencias en tasas de sedimentación entre un lugar y otro. 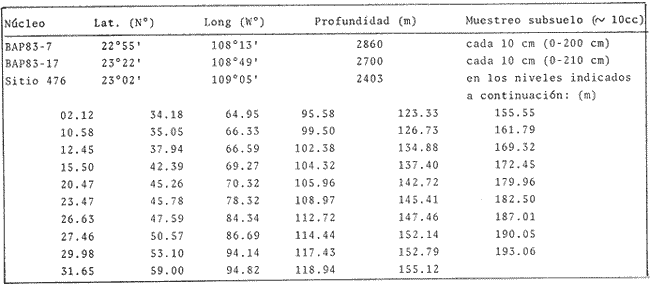 TABLA 2 LOCALIZACIÓN Y MUESTREO DE NÚCLEOS SEDIMENTARIOS Si en los sitios 483 y 484 del DSDP, se considera la profundidad en el subsuelo donde ocurre la "última aparición de Stylatractus universus (Benson, 1983), es posible asumir tasas de sedimentación para los núcleos BAP83-7 y BAP83-17, porque éstos están respectivamente, cerca de esos sitios. La "última ocurrencia" de S. uníversus data de hace 410,000 años (Hays y Shackleton, 1976), y se presenta en el subsuelo del sitio 483 a 30.31 m, y en el subsuelo del sitio 484 a 42.82 m. Por consiguiente, se asume tentativamente una tasa de sedimentación de 7.3 cm/ 1000 años para el núcleo BAP83-7, y una tasa de sedimentos de 10.4 cm/1000 años para el núcleo BAP83-17. Estos valores colocan a la "boca" del golfo con una tasa de sedimentación que es mucho menor que la de su parte medía (compare Figs. 3 y 5); en consecuencia, de acuerdo con lo estipulado por otros autores (Van Andel, 1964; Calvert, 1966, Curray et al., 1979; Aubry et al., 1982; Spiker y Simoneit, 1982). Fechando con las tasas de sedimentación asumidas, las columnas sedimentarias de los núcleos BAP83-7 y BAP83-17, se observa que los "datums" 11,000 años (11 K) y 18,000 años (18 K), indicativos del comienzo del Holoceno y del máximo del último estadio glacial (Shacketon y Opdyke, 1973), corresponden aproximadamente con inflecciones marcadas de los registros estratigráficos de D. pyriformis (Fig. 3); a excepcion del punto 11 K del núcleo BAP83-7. Tal discrepancia puede ser explicada mediante la correlación de los núcleos (Figs. 3 y 4), porque ésta sugiere que la tasa de sedimentación del núcleo BAP83-7 ha sido menor recientemente; luego entonces, "arbitrariamente" se transfirió el datum 11 K de 80 a 50 cm. Con el fin de analizar la tafosenosis de radiolarios, las submuestras sedimentarias (aprox. 10 cc) se trataron con ácido clorhídrico (HCI) y peróxido (H2O2) para disolver los carbonatos y la materia orgánica; tamizándose posteriormente a través de una malla de 62 mieras, con la ayuda de agua corriente filtrada. La porción retenida en el tamiz fue montada en un portaobjetos siguiendo la técnica de "distribución aleatoria" (Randon sampling technique"; Moore, 1973; Molina-Cruz; 1979). Debido a que identificar y definir la distribución de cada una de las especies de radiolarios que habitan en el Golfo de California es una tarea ardua (Benson, 1966), se optó por estructurar un cuadro taxonómico, basado en trabajos previos (Molina-Cruz, 1979; 1982), que comprendiese a las especies más comunes (abundancia relativa mayor al 0. 5 % ). Este cuadro taxonómico es mostrado en la Tabla 3. 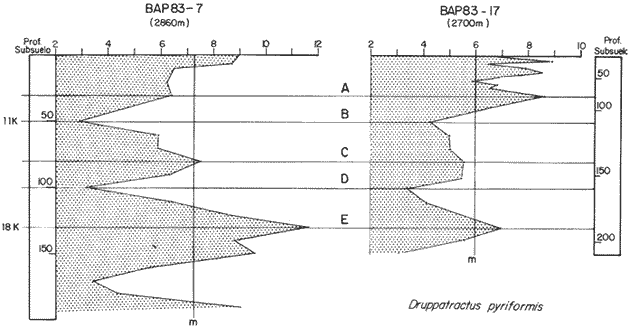 Fig. 3. Correlación estratigráfica entre los núcleos BAP83-7 y BAP83-17, utilizando máximos y mínimos de la abundancia relativa (porcentaje en relación a la población total de radiolarios) de D. pyriformis, Los datums cronoestratigráficos: 11 000 años antes del presente (11 K) y 18 000 años antes del presente (18K) han sido determinados siguiendo el criterio expuesto en el texto. La profundidad en el subsuelo es en cm. El porcentaje de cada una de las especiesconsideradas en el cuadro taxonómico, relativo a la población total de radiolarios, fue evaluado para cada una de las submuestras de la serie "BAP83"; para la serie "BAP82" fue obtenido del trabajo de Álvarez-Arellano y Molina-Cruz (1986). Estos porcentajes sirvieron como datos brutos para aplicar, en el Golfo de California, la técnica paleoceanográfica de: "Funciones transformadas" ("Transfer Function") inventada por Imbrie y Kipp (1971), y después exitosamente empleada por un número significativo de investigadores (Moore, 1973a, Sachs 1973a; 1973b; CLIMAP, 1976; Moorley, 1979; Molina-Cruz, 1984). Dicha técnica, aunque puede ser empleada en varias modalidades (Molina-Cruz, 1977b), generalmente persigue dos propósitos: 1) definir la distribución de conjuntos de mierofósóles, así como su composición de especies, mediante un análisis de factores en su modo "Q" (Imbrie y Van Andel, 1964; Klovan e Imbrie, 1971), y 2) deducir la relación existente entre las distribuciones de conjuntos “antiguos” y las de conjuntos “recientes", mediante un análisis de clasificación-comparación ("Parafactor method"; Sachs, 1973b; 1973c). El análisis de factores fue llevado a cabo mediante el programa computacional "CABFAC" (Pisias et al., 1973) y los resultados son mostrados en las Tablas 4 y 5. Los resultados del análisis de clasificación comparación (programa computacional "THREAD", Pisias et al., 1973) son indicados en la Tabla 6.  TABLA 3 LISTA DE ESPECIES IDENTIFICADAS 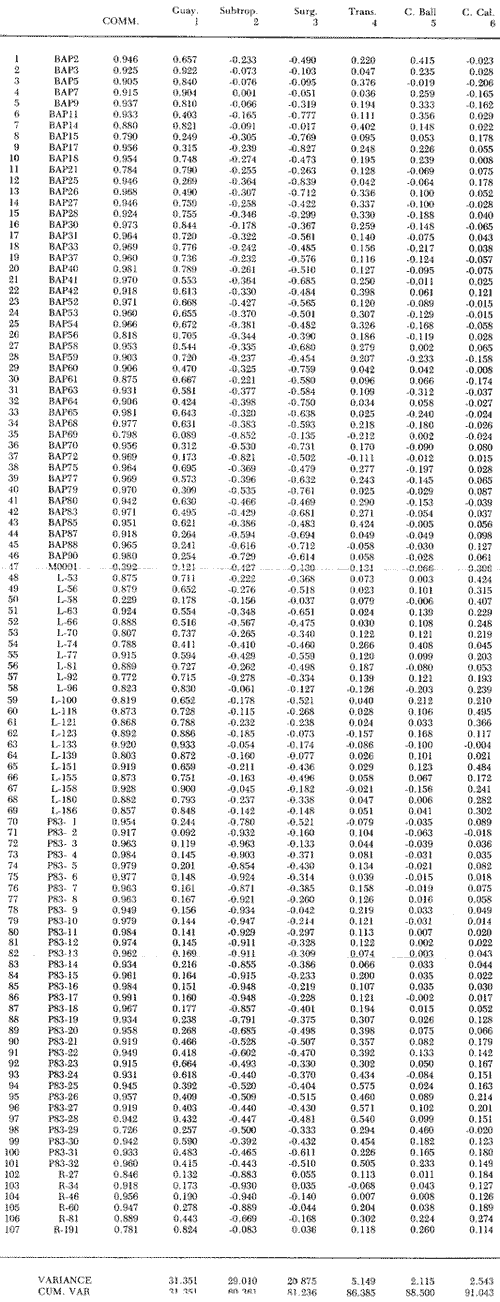 TABLA 4 MATRIZ DE FACTORES Contorneando los valores de la Tabla 4 en mapas del golfo (Fig. 6), se definen las dístribuciones de los conjuntos de radiolarios principales. Graficando estratigráficainente los valores de la Tabla 6 (Fig. 8), se muestra la variación en tiempo de dichos conjuntos. 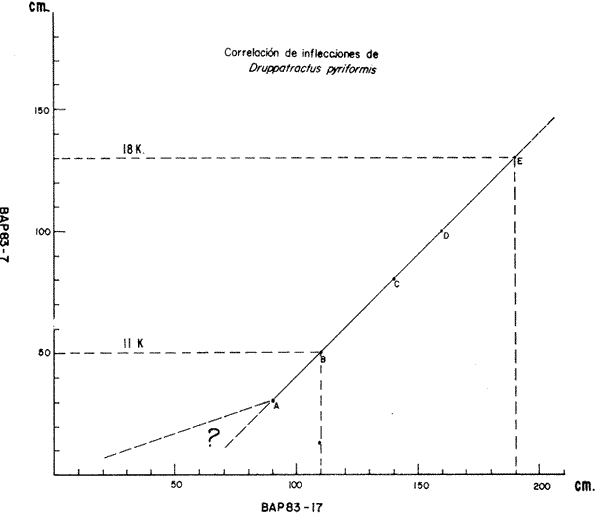 Fig. 4. Correlación estratigráfica entre los núcleos BAP83-7 y BAP83-17, mediante el método de Shaw (1964); utilizando máximos y mínimos de la abundancia relativa (porcentaje en relación a la población total de radiolarios) de D. pyriformis. Nótese la probabilidad de que los 30 cm más superficiales del núcleo BAP83-7, representen una tasa de sedimentación menor. Porque los conjuntos de radiolarios han sido definidos matemáticamente, también se les denomina factores. Las especies afines de cada uno de estos factores son indicados en la Tabla 5, usando la clave que es señalada en la Tabla 3. El comportamiento particular de algunas especies "importantes" se investigó estructurando mapas (Fig. 7) y secciones estratigráficas (Figs. 9 y 10) que muestran su abundancia relativa. RESULTADOS Y DISCUSIÓN
"DISTRIBUCIÓN SUPERFICIAL" DE CONJUNTOS DE RADIOLARIOS"Las "huellas" de las masas de agua en los sedimentos del Golfo de California fueron definidas, mediante el contenido mieropaleontológico, inicialmente, entre otros, por Bandy (1961). Los trabajos de Benson (1966), Round (1967), Murray y Schrader (1982; 1984), Schrader et al. (1984) y Álvares-Arellano y Molina-Cruz (1986)representan un magnífico avance en tal sentido; sin embargo, como se juzgará a continuación, la mayor densidad de muestreo, así como la técnica empleada en este estudio, han contribuido a una definición de las masas del golfo con más detalle. 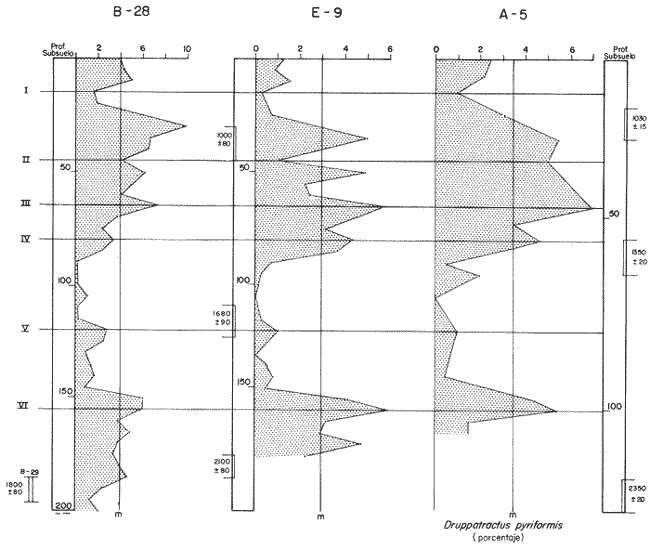 Fig. 5. Correlación estratigráfica de los núcleos B-28, E-9 y A-5, utilizando máximos y mínimos de la abundancia relativa de D. pyriformis. Inferencia paleoceanográfica referente a estos núcleos, localizados en la Cuenca de Guaymas, se ha discutido en el trabajo de Alvarez-Arellano y Molina-Cruz (1984). Sobre la columna: "Prof. subsuelo" se indican fechaciones determinadas mediante C^14. Nótese la coherencia entre dichas fechaciones y la correlación bioestratigráfica. El análisis de factores en su modo "Q" ha producido una matriz (Tabla 4) que explica el 91 % de la variancia total de los datos y define 6 conjuntos (factores) de radiolarios. Éstos de acuerdo a su distribución geográfica (Fig. 6) han sido denominados: 1) "Factor Guaymas”, 2) "Factor Subtropical", 3) "Factor Surgencias", 4) "factor Transicional”, 5) “Factor Canal de Ballenas", y 6) "Factor Corriente de California". El factor "Guaymas" está distribuido principalmente sobre la cuenca del mismo nombre (Fig. 6A), valora el 31.3 % de la variancia de los datos, y contiene cómo componentes principales a las especies Lithomelissa hystrix (N16) y Lithomealissa thoracites (N17; Tabla 5). La afinidad de L. hystríx es tal, que su distribución (Fig. 7A) semeja mucho a la del conjunto en si. Tanto la distribución del factor como la de L. hystrix muestran su mayor abundancia sobre el norte de la cuenca de Guaymas, que es un lugar de cambio batimétrico y morfológico importante en el golfo. Al norte de dicha cuenca, se localiza a las grandes islas Tiburón y Ángel de la Guarda, y las profundidades generalmente son menores a 200 m; al sur, en contraste, éstas alcanzan hasta 2,100 m (Bischoff y Niemitz, 1980). Este hecho batimétrico, así como la obstaculación de circulación, por medio de las islas, provoca un mayor rango en los niveles de las mareas y sus corrientes (Roden, 1964); consecuentemente, una intensa mezcla. En esta región; durante el verano se observó un gradiente térmico perpendicular al eje del Golfo que desciende hacia el sureste y que en invierno se invierte. No obstante tal inversión, el gradiente sugiere una continua mezcla entre las aguas al norte y al sur. Dicha masa de agua, aparentemente, es desplazada por la circulación oceánica invernal hacia el sur, sobre la costa oriental del golfo (Fig. 2B); luego entonces, es posible explicar porque la distribución del conjunto "Guaymas" (Fig. 6A) también se extiende hacia el sureste en dichas localidades. El segundo conjunto de radiolarios en importancia es el "Subtropical”. Éste explica el 29.0 % de la variancia de los datos. Su distribución, extendiéndose desde el Océano Pacífico, se interna notoriamente en el golfo hasta la región norte de la cuenca Farallón (Fig. 6B). Sus especies características: Tetrapyle octacantha (S25; Fig. 7B) y Pterocanium zancleus (N25) son afines a las aguas subtropicales del Pacífico oriental (Nigrini, 1967; Sachs, 1973; Molina-Cruz, 1977; Wenkam, 1977). Druppatractus pyriformis (S6; Fig. 7C) es también aparentemente una especie importante en el factor "Subtropical" (Tabla 5). No obstante, analizando el trabajo de Wenkam (1977), es observado que la distribución de esta especie no se extiende concordantemente con la distribución de las aguas subtropicales, sino más bien que se restringe a las de la región circundante a la boca del golfo. En esta región, la Corriente de California se mezcla de manera compleja con el agua subtropical del Pacífico y la propia del golfo (Álvarez-Sánchez, 1974). Esta condición oceanográfica es análoga a la observada en la región donde la Corriente de Chile (Oceánica del Perú; Wyrtki, 1977) se mezcla con el agua subtropical. Ahí, Cenosphera sp (Molina-Cruz, 1977), una especie cuya teca cortical es muy similar a la D. pyriformis, ocurre abundantemente. (Molina-Cruz, 1984). De acuerdo a lo anteriormente expuesto, D. pyriformis no es estrictamente una especie característica del ambiente subtropical; por consiguiente, su significativa importancia en la composición del Factor Subtropical estructurado en este trabajo (Tabla 5), debe tomarse como el resultado de: 1) que la especie ocurre abundantemente en la región de la boca del golfo (Fig. 7C) y 2) que la definición matemática del factor en cuestión esta restringida a muestras provenientes de dicha región (Fig. 6B). Las zonas del golfo donde ocurren frecuentemente surgencias (Byrne y Emery, 1960; Roden y Groves, 1959; Roden, 1972), son señaladas por la distribución del Factor con ese nombre (Fig. 6C). Este factor comprende el 20.9 % de la variancia de los datos y ocurre notoriamente en las costas. Aparentemente, ha sido fácil definir la distribución del "Factor Surgencias"; sin embargo, esta distribución produce 2 preguntas cuya contestación está limitada en el presente estudio- 1) ¿El florecimiento del conjunto "Surgencias" ocurre al mismo tiempo en todas las localidades?; 2) ¿Las aguas de la subsuperficie que alimentan a las surgencias de la costa oriental del golfo, tienen las mismas características que las que lo hacen en la costa occidental? Respecto a la primera pregunta, es posible argumentar que debido a que las surgencias de la costa occidental ocurren en verano y las de la costa oriental ocurren en invierno (véase la sección MARCO OCEANOGRAFICO), el florecimiento del "factor Surgencias" debe presentarse alternadamente entre el margen occidental y el margen oriental del golfo. Respecto a la segunda pregunta, se especula que no, basados en la dinámica circulatoria del golfo y en la distribución de especies afines a las aguas de la cuenca de Guaymas y de la Corriente de California: L. hystrix, Fig. 7A) y C. davísiana (Fig. 7D), respectivamente. Las surgencias de la costa oriental son invernales y coincidentes con un desplazamiento superficial hacia el exterior del golfo, consecuentemente, con mayores posibilidades de ser alimentadas, al menos en parte, por aguas originales de la cuenca de Guaymas, tal como es evidenciado por la distribución de L. hystrix. Las surgencias de la costa occidental ocurren en verano, cuando el desplazamiento superficial es hacia el interior del golfo; luego enconces, con mayores posibilidades de que estas surgencias sean alimentadas, al menos en parte, por aguas subsuperficiales de la Corriente de California, tal como es evidenciado por la ocurrencia de C. davisiana en esas zonas. 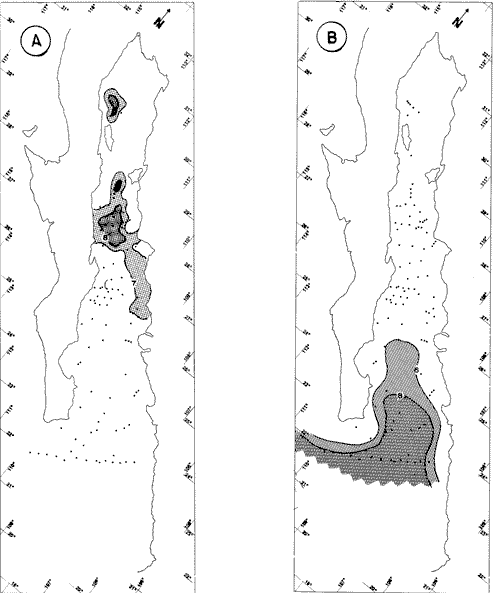 Fig. 6. Distribución de factores de radíolarios en el Golfo de California. A) Factor "Guaymas",B) Factor "Subtropical", C) Factor "Surgencias" y D) Factores: "Transicional", "Canal de Ballenas" y "Corriente de California". Los valores de los contornos indican "pesos" del factor por 10. 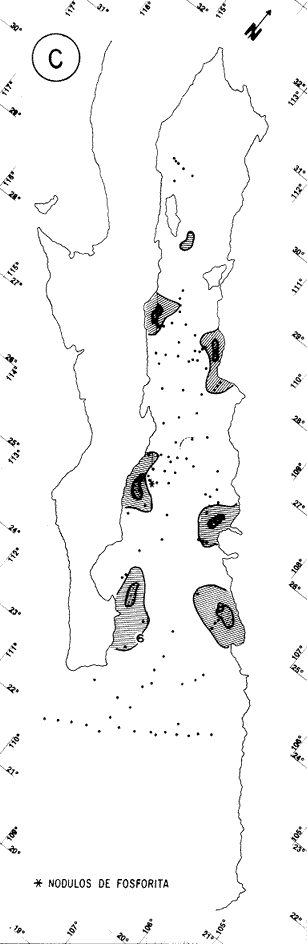 Fig. 6-C 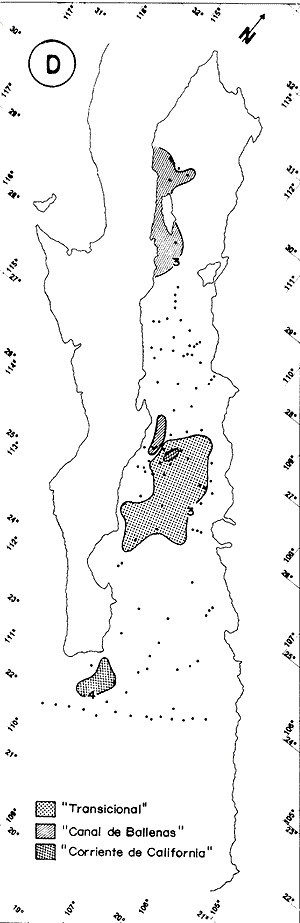 Fig. 6-D 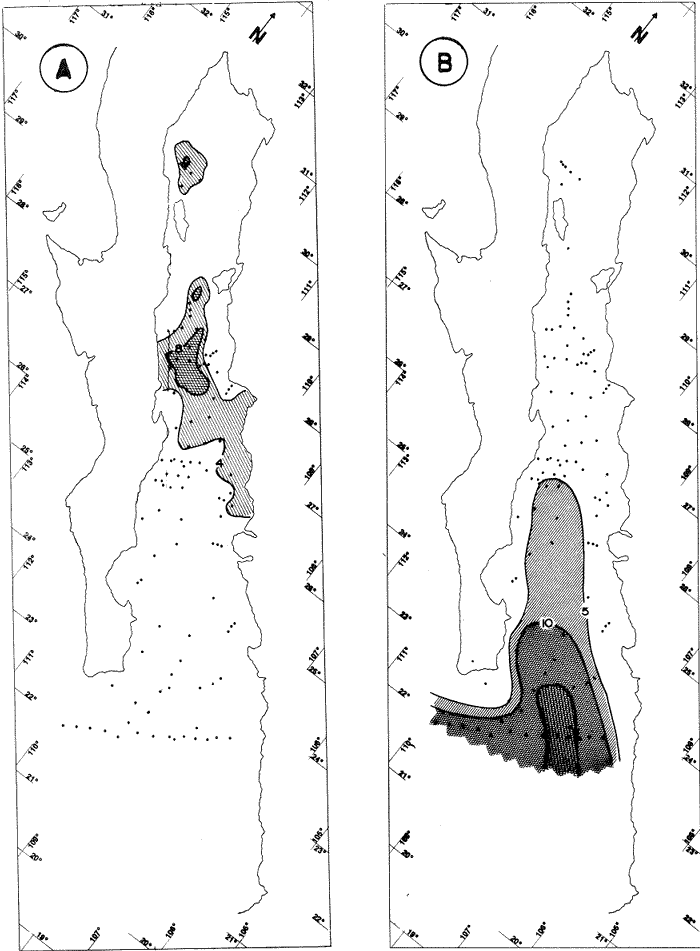 Fig. 7. Distribución de algunas especies de radiolarios en el Golfo de California. A) Lithomelissa hystrix, B) Tetrapyle octacantha, C. Druppatractus pyriformis, y D) Hexacontium entacanthum y CycIadophora (?) davisiana. Los valores de los contornos indican abundancia relativa (porcentaje en relación a la población total de radiolarios). 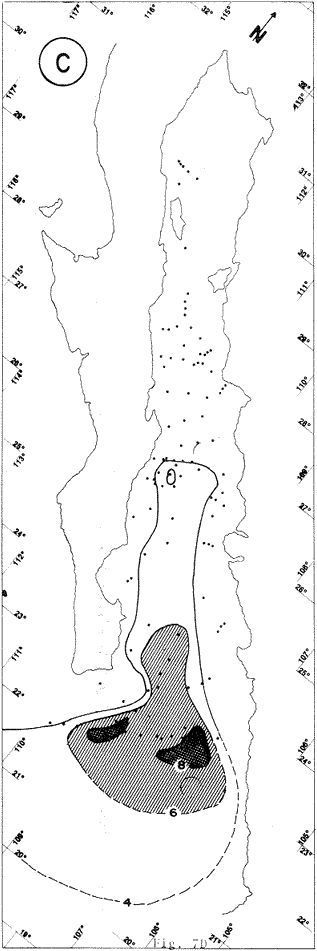 Fig. 7-C 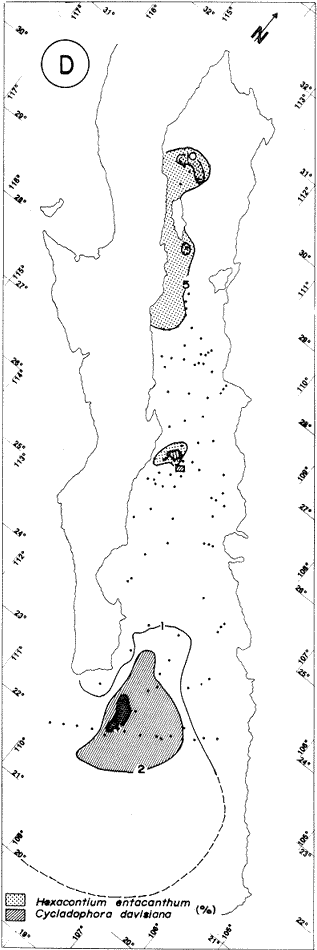 Fig. 7-D Es importante hacer notar que bajo una de las zonas de surgencias de la costa occidental (Fig. 6C), se han recuperado nódulos de fósforita. Aunque el origen de dichos nódulos a un es motivo de discusión, es generalmente aceptado que para su formación se requiere de la conjunción de varios factores ambientales, tales como alta productividad orgánica, baja tasa de sedimentación y condiciones anóxicas (Seibold y Berger, 1982; p. 73). No está al alcance de este trabajo evaluar detalladamente a dichos factores en la región; pero sí el de: 1) sugerir que la Corriente de California, al igual que en la Costa Pacífica de Baja California, contribuye al carácter altamente productivo de las surgencias en el golfo; 2) el mencionar que las márgenes continentales del Golfo de California, las condiciones anóxícas son comúnes (Calvert, 1964) y 3) que el flujo detrical terrígeno de la península de Baja California hacía el golfo es bajo (Van Andel, 1964). Aunque han sido sugeridas algunas explicaciones para las cuestiones anteriores, éstas no deben ser tomadas como concluyentes, hasta que no se efectúen hidrocalas y muestreos planetónicos periódicos en esas regiones. Phortícíum pylonium clevei (S16) es abrumadoramente la especie más importante dentro del "Factor Surgencias” (Tabla 5); sin embargo, se renonoce que su identificación fue en ocasiones dudosa. Los restantes 3 factores del modelo matemático (Tabla 4), los factores "Canal de Ballenas", ''Transicional'' y "Corriente de California" (Fig. 8D), valoran conjuntamente una mínima parte de la variancia de los datos (9.8 %). No obstante, aportan información que es coherente con la dinámica oceanográfica del golfo. Sobre la costa occidental del golfo, principal mente de su parte media al norte, hay una serie de canales formados entre islas y la península de Baja California. De éstos, el Canal de Ballenas, localizado al occidente de la isla Ángel de la Guarda, es el más conspicuo y el más estudiado (Badan et al., 1985). Ahí, las corrientes, principalmente las de marea, alcanzan velocidades hasta de 3 m/seg (Álvarez et al., 1984), produciendo intensas mezclas que son expresadas en la superficie por temperaturas relativamente más frías que las adyacentes (Fig. 2, Robinson, 1973), y en la subsuperficie, por una disminución de la estratificación. Por ejemplo, en el Canal de Ballenas se observan temperaturas de 11.3°C a 1,000 m de profundidad, mientras que en la cuenca de Guaymas, a esa profundidad, las temperaturas son de 5°C (Gaxiola, 1978). La extenxión geográfica de las particularidades oceanográficas del Canal de Ballenas son señaladas por la distribución del Factor del mismo nombre (Fig. 6D), a pesar de los limitado del muestreo en esa región. El "factor Canal de Ballenas” tiene como especie afín a Hexapyle enthacanthum (S9; Tabla 5, Fig. 7D), y presenta manifestaciones también sobre los canales formados entre las islas de la cuenca del Carmen y la península de Baja California. Entre el agua de Guaymas y la subtropical, sobre la cuenca del Carmen, se define moderadamente el "Factor Transicional" (var. 5.1 %, Fig. 6D). Este factor es llamado así, porque las especies que actúan como sus componentes principales, Eucyrtidium siphonostoma (N11), Lithomelissa hystríx (N16), Pseudocubus obeliscus (N22), y Druppatractus pyriformis (S6), también están significativamente presentes en el Factor Guaymas, al norte, o en los "Factores subtropícal" y "Corriente de California", al sur (Tabla 5). Este hecho sugiere que la cuenca del Carmen es una región en el golfo, donde las aguas incursionantes del Pacífico son diluidas por las formadas en la cuenca de Guaymas o viceversa. Esta zona de transición de masas de agua fue propuesta por Round (1967), y descrita por Roden (1964) como una región donde las aguas subtropicales del Pacífico son modificadas mediante evaporación, principalmente. De acuerdo a la condición impuesta en la Tabla 5, ninguna especie puede considerarse afín al "Factor Transicional", sin embargo, cabe mencionarse que Eucyrtidium siphonostoma (N11) ocurre abundantemente en este ambiente. 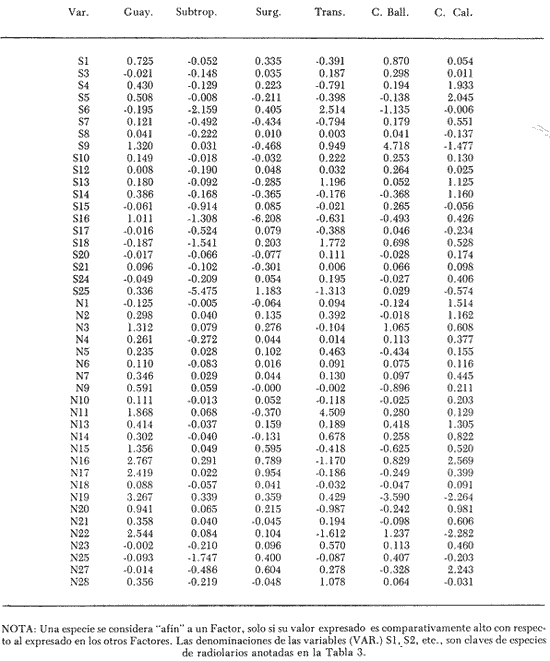 TABLA 5 SCORES DE LAS VARIABLES (ESPECIES DE RADIOLARIOS) EN LOS FACTORES La Corriente de California generalmente, presenta una clara expresión de flujo hasta la punta de la península de Baja California (Fig. 2). Por tal motivo, se asume que el factor que ahí se describe le corresponde y ha sido llamado "Factor Corriente de California" (Fig. 6D). Esta decisión es apoyada porque las especies principales que integran a este factor (Tabla 5) Cycladophora davisíana (N27), Druppatractus irregularis (S5), Botryostrobus aquilonaris (N1) y Botríostrobus aurítus/australís group (N2), han sido reportadas en varias ocasiones como afines a la Corriente de California (Kling, 1973; Wenkan, 1977; Moore, 1973a). Aunque el "Factor Corriente de California" ha sido definido precariamente (var. 2.5 %), su presencia en el margen occidental de la cuenca del Carmen, así como la de una de sus especies afines: C. davisíana, apoya las indicaciones de que la Corriente de California incursiona en el golfo, al menos subsuperficialmente en el lado occidente (Roden, 1972). Tal hecho proporciona posibilidades de que las aguas de dicha corríente participen en las surgencias de la costa occidental del golfo, que ocurren en verano. Cabe mencionar, que las aguas de la Corriente de California pueden ser inducidas hacia el interior del golfo en verano, porque en esta estación el patrón de circulación es hacia el noreste (Fig. 2, Robinson, 1973). Aun cuando todos los elementos manejados en este estudio indican que Cycladophora davisiana (N27) es una especie afin a la Corriente de California, es importante hacer notar que su abundancia se incrementa significativamente, en la región donde la mencionada corriente participa en la formación de "frentes oceánicos", principalmente en la boca del golfo (Fig. 7D). Esta observación, por consiguiente apoya la sugerencia de Molina-Cruz (1984) de que C. davisíana "florece" en regiones con estructuras oceanográficas donde participan al menos 2 masas de agua con características termales contrastantes. ESTUDIO ESTRATIGRÁFICOTomando en cuenta que la localización de los núcleos BAP83-7 y BAP83-17 es la boca del golfo (Fig. 2), el análisis de "Para factores" indica que el modelo definido en el "Estudio Superficial" no ha sido perturbado desmesuradamente en el tiempo considerado; ya que sólo los valores de los factores "Subtropical”, "Surgencias" y "Corriente de California" son significativos (Tabla 6). Las fluctuaciones de estos factores (Fig. 8), sin embargo, permiten observar cambios oceanográficos relevantes que han ocurrido en la región, desde el máximo del último estadio glacial, ocurrido hace 18,000 años, hasta el reciente. La contraposición de fluctuaciones entre los registros del factor “Subtropical"(Fig. 8A) y del factor "Corriente de California" (Fig. 8B), referidas a un marco estratigráfico establecido (ver la sección de métodos), permite a primera instancia hacer deducciones paleoceanográficas: Los valores altos del "Factor Subtropical" (Fig. 8A) indican que sus aguas correspondientes son las que principalmente han inundado la región de la boca del golfo, al menos durante los últimos 20,000 años. Este dominio, sin embargo, ha sido frecuentemente disminuido por incursiones de la Corriente de California, tal como es evidenciado por el factor del mismo nombre (Fig. 8B). Dichas incursiones aparecen más severas durante la parte tardía del último estadio glacial (desde hace aproximadamente 20,000 a 11,000 años) que durante el postglacial (iniciado hace 11,000 años); particularmente, alrededor del datum 18 k. En el registro del núcleo BAP83-17 se sugiere inclusive, una mayor incursión hacia el interior del golfo. El agua subtropical, por el contrario, ha observado una mayor estabilidad en el Holoceno (postglacial presente), que durante la parte tardía del último estadio glacial; mostrando en consecuencia que sus valores más altos han ocurrido muy recientemente (Fig. 8A). El registro del "Factor Surgencias" (Fig. 8C) no sólo sugiere eventos paleoceanográficos, sino que apoya la correlación estratigráfica establecida entre los dos núcleos considerados (ver métodos), a través de un mínimo notorio situado aproximadamente en el año 15 K de la columna cronoestratigráfica. Las columnas sedimentarias de los núcleos BAP837 y BAP83-17 no registran idealmente los cambios dinámicos de las surgencias, porque se localizan lejos de la costa (Fig. 2). No obstante, lo "marcado" de las fluctuaciones del "Factor '-Surgencias" en el intervalo comprendido entre los años 20,000 y 11,000 antes del presente (A.P.), sugieren un cuadro elimático oceanográfico más dinámico durante ese tiempo que recientemente; entonces, acorde con las deducciones hechas antes sobre la Corriente de California. 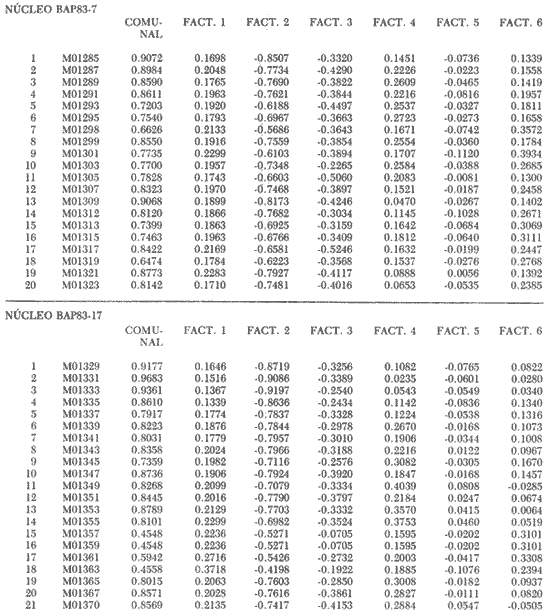 TABLA 6 MATRIZ DE "PARAFACTORES" PARA LOS NÚCLEOS: BAP83-7: Y BAP83-17 FACT. I-GUAY. FACT. 2-SUBTROP. FACT. 3-SURG. FACT. 4-TRANS, FACT. 5-C. BALL. FACT. 6-C. CAL. La paleoceanografía regional hasta ahora estructurada en este estudio, encaja en la paleoceanografía de carácter global, y no difiere de aquella propuesta para regiones cercanas al Golfo de California. El estudio de "CLIMAP” (1976), comprendiendo una extensión mundial, señala que durante el máximo del último estadio glacial, la circulación eólica y la oceánica fueron más rápidas. Por consiguiente, el haber definido incursiones "severas” de la Corriente de California en la boca del golfo, durante la parte tardía del último estadio glacial, es coherente con dicho cuadro paleoceanográfico. 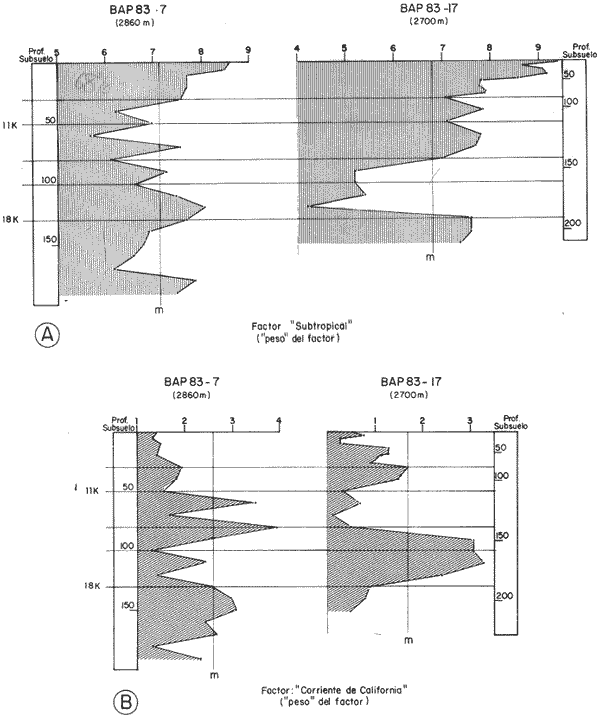 Fig. 8 Distribución estratigráfica, en los núcleos BAP83-7 y BAP83-17, de los factores: A) "Subtropical", B) "Corriente de California y C) "Surgencias". La correlación entre los núcleos, así como los datums 11K y 18K, fueron determinados en las figuras 3 y 4. 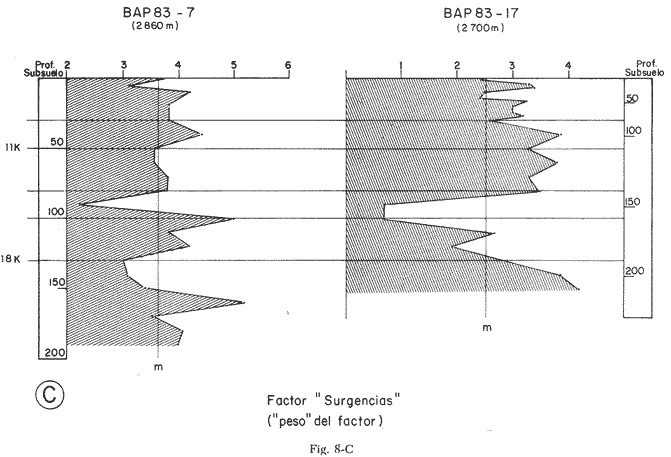 Fig. 8-C Los estudios regionales de Moore (1973a) y Wenkan (1977), señalando que la Corriente de California fue más rápida durante el último estadio glacial, apoyan las deducciones argumentadas arriba. La respectiva afinidad de las especies Tetrapyle octacantha y Cyladophora davisiana a los factores "Subtropical" y "Corriente de California”, discutida en el estudio superficial, es confirmada por el estudio estratigráfico, ya que las curvas de los registros de estas especies (Fig. 9) son paralelas a la de sus factores respectivos (Fig. 8). Dicha confirmación, por consiguiente, sugiere que es válido utilizar especies afines a un ambiente para formular deducciones paleoceanográficas. De acuerdo a las perforaciones realizadas por el DSDP, los sedimentos más antiguos que sobreyacen sobre el basamento del Golfo de California son los recobrados en el sitio 476 (23°02'N; 109°05'W). Cuando se eligió tal sitio para perforación, se consideró que siendo las márgenes del golfo del "tipo pasivo”, en ese lugar se encontraría a los sedimentos depositados durante la iniciación de la dispersión del piso oceánico, que está originando al golfo (Curray et al., 1982). Por consiguiente, graficando las fluctuaciones de las especies Druppatroctus pyriformis y CycIadophora davisiana (Fig. 10), con respecto a la biocronoestratigrafía del sitio 476 (Aubry et al., 1982) se han formulado inferencias paleoceanográficas que datan desde los comienzos de la formación del golfo. En el presente, Druppatractus pyriformis es común en los alrededores de la boca del golfo, como consecuencia de la mezcla que ocurre entre las aguas subtropicales y las de la Corriente de California (véase descripción del factor Subtropical"). Luego, la abundante presencia de esta especie en los inicios de la formación del golfo (Fig. 10A), sugiere que tal proceso ha venido ocurriendo en las latitudes correspondientes a la boca del golfo, al menos desde ese entonces. Esta sugerencia es viable, ya que la Corriente de California fluye desde aproximadamente el Cretásico (Sliter, 1972). No obstante, las incursiones de dicha corriente al interior del golfo, empiezan a ocurrir desde hace aproximadamente 1.4 millones de años, de acuerdo al comportamiento estratigráfico de Cycladophora davisiana (Fig. 10B). Este datum, sin embargo, es tentativo, porque una gran dilución de radiolarios por medio de terrígenos, en el contacto Pleistoceno/Plioceno, no permite deducir si dichas incursiones ocurren desde antes. (Molina-Cruz, 1982). 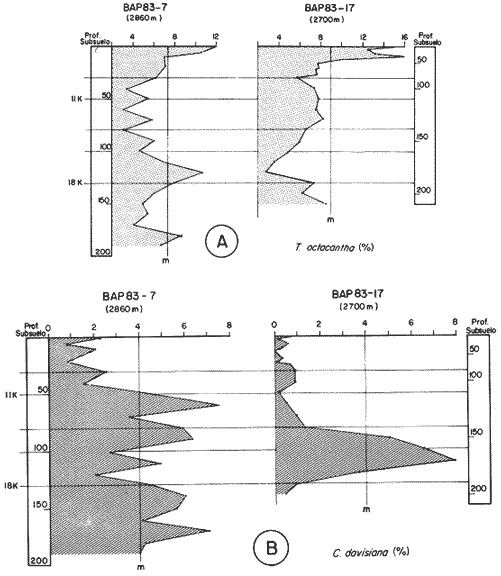 Fig. 9. Distribución estatigráfica, en los núcleos BAP83-7 y BAP83-17, de la abundancia relativa (porcentaje en relación a la población total de radiotarios) de las especies T. octacantha y C. (?) davisina. La correlación entre los núcleos, así como los datums 11K. y 18K, fueron determinados en las figuras 3 y 4. 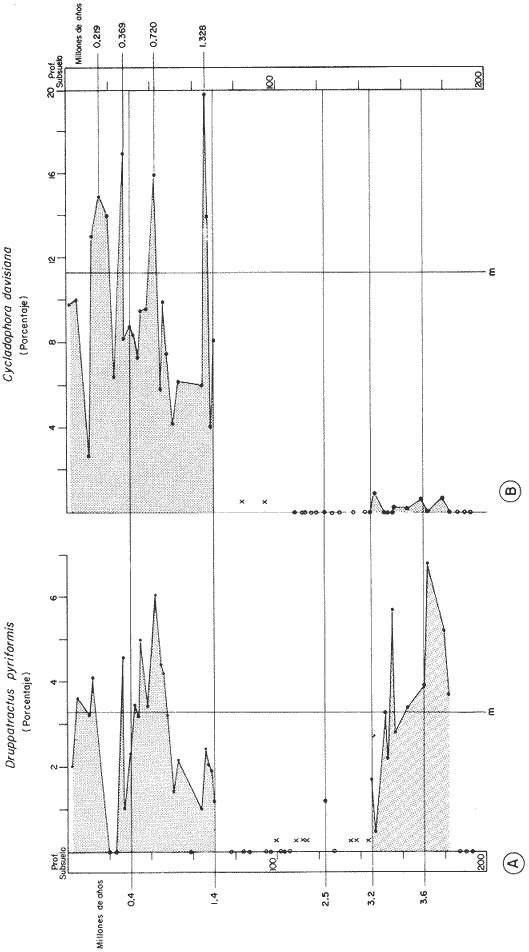 Fig. 10. Distribución estratigráfica, en el sitio 476 del "Deep Sea Drilling Project" (D.S.D.P.), de la abundancia relativa (porcentaje en relación a la población total de radiolarios) de las especies D. pyriformis y C. (?) davisina. Las edades del subsuelo han sido determinadas de acuerdo a los trabajos de Aubry et al. (1982) y Molina-Cruz (1982). Los puntos -vacíos- corresponden a muestras que no contiene radiolarios. Las muestras indicadas por una "x" contienen muy escasos radiolarios; por lo que no se estimó su porcentaje. Apoyados en el comportamiento de D. davisina en la Fig. 9B. antes se argumentó que una marcada incursión de la Corriente de California al golfo, ocurrido hace 18,000 años, durante el máximo del último estadio glacial. Por consiguiente, considerando dicha deducción, en el registro de C. davisiana descrito en la Fig. 10B, se sugiere que alrededor de los años 1.328,000, 720,000, 369,000 y 219,000 antes del presente, ocurrieron también incursiones notorias de la corriente en cuestión. Se hace notar que el datum 720,000 k corresponde aproximadamente a la edad del contacto paleomagnético Brunhes/Matuyama. Esta inversión magnética, aparentemente fue acompañada por condiciones glaciales (Molina-Cruz, 1979a). ConclusionesLa distribución geográfica de seis conjuntos (factores) de radiolarios, en los sedimentos del Golfo de California, "refleja" la ubicación y extensión de algunos fenómenos oceanográficos, tales como surgencias, frentes oceánicos y formación de masas de agua. Estos conjuntos, a su vez, tienen especies "representativas"; por ejemplo: L. hystríx es afin al "Factor Guaymas", T. octacantha al “Factor Subtropical”, P. pylonium clevei al "Factor Surgencias", H. Enthacantum” al "Factor Canal de Ballenas", y C. (?) davisiana al “Factor Corriente de California". Los factores "Guaymas", "canal de Ballenas" y "Surgencias" se desarrollan bajo influencias de fenómenos oceanográficos que ocurren en el interior del golfo; mientras que, los factores "Subtropical" y Corriente de California" provienen del Océano Pacífico. La Corriente de California fluye sobre las latitudes 20°-24' norte, desde antes de la formación del Golfo de California, inicia da aproximadamente en el contacto Plioceno/Mioceno. Sin embargo, dicha corriente no incursionó al golfo, sino hasta el Pleistoceno; aproximadamente hace 1.4 millones de años. Las aguas de la Corriente de California han participado en las surgencias que ocurren al sur de la costa occidental del golfo, particularmente durante los máximos de estadios glaciales, en los que las incursiones de dicha corriente fueron más evidentes. Esta circulación oceanográfica, más dinámica durante el último estadio glacial que recientemente, es coherente con la evidenciada en una extensión mundial. AgradecimientosA Alejandro Álvarez Arellano por permitir usar parte de los datos de su tesis de maestría y criticar este trabajo. A Ana María Pérez Guzmán por la revisión y sugerencias al manuscrito. Mis agradecimientos se extienden a Alejandro Pineda M., por su ayuda en laboratorio. A Alejandra Estrada, por mecanografiar el manuscrito. Este estudio contó con el apoyo económico del CONACyT, bajo el convenio "Evolución Oceanográfica del Golfo de California", PCECBNA021993. LITERATURAÁLVAREZ-SÁNCHEZ, L. G. M.S. Thesis Currents and Water Masses at the Entrance to the Gulf of California, spring 1970. Oregon State University, Corvallis 1974 75 p. ALVAREZ-ARELLANO A. y A. MOLINA-CRUZ An. Inst. Cienc. del Mar y Limnol. Aspectos poleoceanográficos cuaternarios del Golfo de California evidenciados por conjuntos de radiolaríos. Univ. Nal. Autón. México 1986 67-94 13 (2) ALVAREZ, L. G., A. BADAN-DARGON y J. M. ROBLES Lagrangian observations of near-surface currents in Canal de Ballenas. CALCOFI, Rep. 1984 35-42 25 AGUAYO-CAMARGO, E. Rev. del Inst. Mex. del Petróleo Origen y distribución de sedimentos en el Golfo de California. 1981 5-19 13 (3) AUBRY, M. P., MATOBA, A. MOLINA-CRUZ y H. SCHRADIGNI Inítíal Reports of the Deep Sea Drilling Project Synthesis of Leg 64 biostratígraphy. 1982 1057-1064 64 (2) BADAN-DANGON, A., C. J. KOBLINSKY y T. BAUMGARTNER Spring and summer in the Gulf of California: observations of surface thermal patterns. Oceanológica Acta 1985 13-22 8 (1) BANDY, O. L. Micropaleontology Distribution of foraminifera, radiolaría and díatoms in sediments of the Gulf of California 1961 1-26 7 BENSON, R. N. Recent radiolaria from the Gulf of California Ph. D. Thesis University of Minnesota 1966 5-7 p. BENSON, R. N. Initial Reports of the Deep Sea Drilling Project Quaternary radiolarian from the mounth of the Gulf of California, deep sea drilling project. Leg. 65. 1983 491-523 65 BISCHOFF, J. L. y J. W. NIEMITZ Bathymetric maps of the Gulf of California. Geol. Surv. Misc. Investing. Ser (U.S.) 1980 Map, 1-1244, 4 plates. BYRNE, J. V. y L. P. EMERY Geol. Soc. Am. Bull Sediments of the Gulf of California. 1960 983-1010 71 CALVERT, S. E. Marine Geol. of the Gulf of California: a symposium. Factors affecting distribution of laminated diatomaceous sediments in the Gulf of California. Van Andel, Tj. H. and G. G. Shor, Jr. Amer. Assoc. Petrol. Geologist. Memoir 1964 311-330 3 CALVERT, S. M. Geol. Soc. Am. Bull. Aceumulation of diatomaceus siliea in the sediments of the Gulf of California. 1966 569-596 77 CLIMAP PROYECT MEMBERS Science The Surface of the leeage Earth. 1976 1131-1137 191 CRONWELL, T. y J. REID, Jr. A study of oceanic fronts. Tellus 1956 1131-1137 8 CURRAY, J. EN COLABORACIÓN CON LOS PARTICIPANTES DE LA ETAPA 64 DEL D.S.P.P. Geotimes In the Gulf of California, Leg 64 seeks Evidence on Develompent of Basins. 1979 18-20 24 (17) CURRAY, J. R., D. G. MOORE, K. KELTS y G. EINSELE Initial reports of the Deep Sea Drilling Project Tectonics and geological history of the passive continental margin at the tip of Baja California. 1982 1089-1116 64 (2) DONEGAN, D. y H. SCHRADER Marine Geol. Biogenic and abiogenic components of laminated sediments in the central Gulf of California. 1982 214-237 48 GAXIOLA-CASTRO Tesis del Lic. Sistema de bióxido de carbono en el Golfo de California. Esc. Sup. de Ciencias Marinas. Univ. Aut. de Baja Cal. Ensenada, B.C. 1978 57p. GRIFFITHS, R. C. Studies of oceanic fronts in the mounth of the Gulf of California, an area of Tuna migrations. F.A.O. Fish Rep. 1963 1583-1609 6 (3) HASTING, J. R. y R. M. TURNER Geografiska Annaler Seasonal precipitation regimes in Baja California, México. 1965 204-223 47 HAYS, J. D. y N. J., SHACKLETON Geology Globally synchronous extinction of the radiolarian Stylatractus universus. 1976 649-652 4 IMBRIE, J. y Tj. H. VAN ANDEL Geol. Soc. Amer. Bull. Vector analysis of heavy mineral data. 1964 1131-1156 75 IMBRIE, J. y N. G. KIPP The Late Cenozoíc Glacial Ages. A new mieropaleonotological method for quantitative paleoclimatology: Application to a late pleistocene Caribbean core K. K. Turekian. Yale University Press New York 1971 p. 71-181 INGLE, J. C. Neotectonícs and Sea level Variations in the Gulf of California Area. A Symposium. Neogene stratigraphy and tectonic history of the Gulf of California Inst. Geol. Univ. Nal. Autón. México 1984 p. 111. KLING, S. A. Inítial of the Deep Sea Drílling Prolect Radiolaria from the eastern north Pacific, Leg 18, Deep Sea Drilling Project. 1973 617-673 18 KLOVAN, J. E. y J. IMBRIE Mathematical Geol. An algorithm and FortramIV program for large-scale Q-mode factor analysis and calculation of factor scores. 1971 61-77 3 (1) MOLINA-CRUZ, A. Marine Micropal. Radiolarian assemblages and their relationship to the oceanography of the subtropical S.E. Pacific. 1977 315-352 21 (4) MOLINA-CRUZ, A. The relation of the southern trade winds to upwelling proccesses during the last 75,000 years. Quat. Res. 1977b 324-338 8 (3) MOLINA-CRUZ, A. Memorias del Symposium, El Golfo de California: Origen, Evolución, Aguas, Vida Marina y Recursos. Mazatlán, Sinaloa, México, noviembre, 1979. Registro micropaleontológico de las masas de agua en la región central del Golfo de California. 1979 (en prensa) MOLINA-CRUZ, A. Ciencias Marinas El Pacífico suroriental durante la época paleomagnética Brunhes: breve consideración paleoceanográfica. 1979a 65-73 6 (1 y 2) MOLINA-CRUZ, A. Initíal Reports of the Deep Sea Drilling Project Radiolaria in the Gulf of Calífornia: Deep sea drilling project, Leg 64 1982 983-1002 64 (2) MOLINA-CRUZ, A. Marine Micropal The radiolarian remains as indicators of upweIling processes: The peruvian connection. 1984 53-75 9 MOORE, T. C., Jr. Method of random1y distributing grains for microscopic examination. Jour. Sed. Petrol. 1973 904-906 43 (3) MOORE, T. C., Jr. Late Pleistocene-Holocene oceanographic changes in the northeastern Pacific. Quat. Res. 1973a 99-109 3 (1) MORLEY, J. J. A transfer function for estimating paleoceanographic conditions, based on deep-sea surface distribution of radiolarian assemblages in the south Atlantic. Quat. Res. 1979 381-395 12 MURRAY, D. y H. SCHRADER Marine Micropal Distribution of silicoflagellates in plankton and core top samples from the Gulf of California. 1982 517-539 7 MURRAY, y H. SCHRADER Marine Micropal. (en prensa) Silicofiagellate assemblages in the Gulf of California during the last glacial maximum and the present: Oceanographic implications. 1984 NIGRINI, C. Mieropaleontology Radiolaria from the eastern tropical Pacific sediments. 1967 51-63 14 (1) NIGRINI, C. y T. C. MOORE, Jr. Spec. Publ. Cushman Found Foraminiferal Res. A Guide to Modern Radiolaria. 1978 16 PISIAS, N., P. DAUPHIN, T. C. MOORE y C. R. HEATH CLIMAP Data Reduction and Processing Programs. Oregon State University. Spec. Publ. 1973 153 p. ROBINSON, M. K. Atlas of Monthly Mean Sea Surface and Subsurface Temperatures in the Gulf of California, Mexico. Society of Natural History, San Diego, Cal., Memoir 5 1973 90 figs., 19 p. RODEN, G. I. Geological of the Gulf of California. Oceanographic aspects of the Gulf of California. Van Andel, Tj. y G. G. Shor Am. Assoc. Petrol. Geol. Mem. 1964 30-58 3 RODEN, G. I. Thermohaline and baroclinic flow across the Gulf of California entrance and in the Revillagigedo islans region. J. Phys. Oceanog. 1972 177-183 2 (2) RODEN, G. I. y G. W. GROVES J. Marine Res. Recent oceanograpihe investigations in the Gulf of California. 1959 10-35 18 (1) ROUND, R. E. J. Esp. Mar. Biol. Ecol. The phytoplankton of the Gulf of California. Part I: Its composition, distribution to the sediments. 1967 76-96 1 SACHS, H. M. Ph. D. Thesis Quantitative radiolarian-based paleoceanography in late Pleistocene subartic Pacific sediment. Brown University, Providence Rhode Island 1973 208 p. SACHS, H. M. North Pacific radiolarian assemblages and relationship to oceanographic parameters. Quat. Res. 1973b 73-88 3 SACHS, H. M. Late Pleistocene history of the north Pacific: Evidence from a quantitative study of radiolarian in core V21-173´ Quat. Res. 1973c 89-98 3 SHACKLETON, N. J. y N. D. OPDYKE Oxygen isotope and paleomagnetic stratigraphy of equatorial Pacific core V28-238 oxygen isotope temperatures and ice volumes of 105 year scale. Quat. Res. 1973 39-55 3 (1) SHAW, A. B. Time in Stratigraphy. McGraw-Hill Book Co. New York, N.Y. 1964 365 p. SCHRADER, H. y T. BAUMGARTNER Coastal upwelling: Its Sediment Record, Plenum Publishing Corporation. Decadal variation of upwelling in the central Gulf of California. Thiede J., y E. Suess 1983 SCHRADER, H. R. MAHOOD, y G. CHENG Marine Micropaleontology Gulf of California: Silicoflagellates in phytoplankton samples collected in june 1982, Part. 2. (en prensa). 1984 SHWARTZLOSE, R. A. y J. R. HENDRICKSON Inst. Cienc. del Mar y Limnol. Univ. Nal. Auton. Mexico. Publ. Espec. Bibliografía del Golfo de California: Ciencias Marinas. (Hasta el final de 1981). 1983 1-212 7 SEIBOLD, E. y W. H. BERGER Berlin, Heidelberg. The Sea Flor. An Introduction to Marine Geology. Springer Verlag New York 1982 288 p. SIMPSON, J. H., D. J. EDELSTEN, A. EDWARDS, N. C. G. MORRIS y P. B. TELT The islay front: Physical structure and phytoplankton distribution. Est. and Coast. Mar. Sci. 1979 713-726 9 SLITER, W. V. Palaeogeogr. Palaeoclimatol. Paleo. Upper Cretaceous planktonic foraminiferal zoogenography and ecology-eastern Pacific margin. 1972 12 (3) SOUTAR, A., S. R. JOHNSON y T. R. BAUNGARTNER The Monterey Formation and Related Siliceous Rocks of California. In search of modern depositional analogs to the Monterey formation. Garrison, G. E. y R. G. Douglas Society of Economic Paleontologists and Mineralogists. 1981 p. 123-147. SPIKER, E. C. y B. R. T. SIMONEIT Initial Reports of the Deep Sea Drilling Project Radiocarbon dating of recent sediments from Leg 64, Gulf of California 1982 757-758 64 (2) VAN ANDEL, Tj. H. Marine Geology of the Gulf of California a symposium. Recent marine sediments of Gulf of California Van Andel, Tj. H. y G. G. Shor The American Association of Petroleum Geologist. Memoir 1964 216-310 3 WARSH, C. E., K. L. WARSH y R. C. STANLEY Nutrients and water masses at the mounth of the Gulf of California. Deep-Sea Res. 1973 561-570 20 WENKAM, CH. M. S. Thesis Late quaternary changes in the oceanography of the eastern tropical Pacific. School of Oceanography, Oregon State University. Corvallis Or. 1977 143 p. WIRTKI, K. Inter-Amer. Trop. Tuna Comm. Bull. Surface currents of the eastern tropical Pacific. 1965 207-304 9 (5) WYIRTKI, K. Advection in the Peru current as observed by satellite. J. Geoph. Res. 1977 3939-3943 82 27
|